
Effects of Landscape and Local Factors on the Diversity of Flower-Visitor Groups under an Urbanization Gradient, a Case Study in Wuhan, China

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
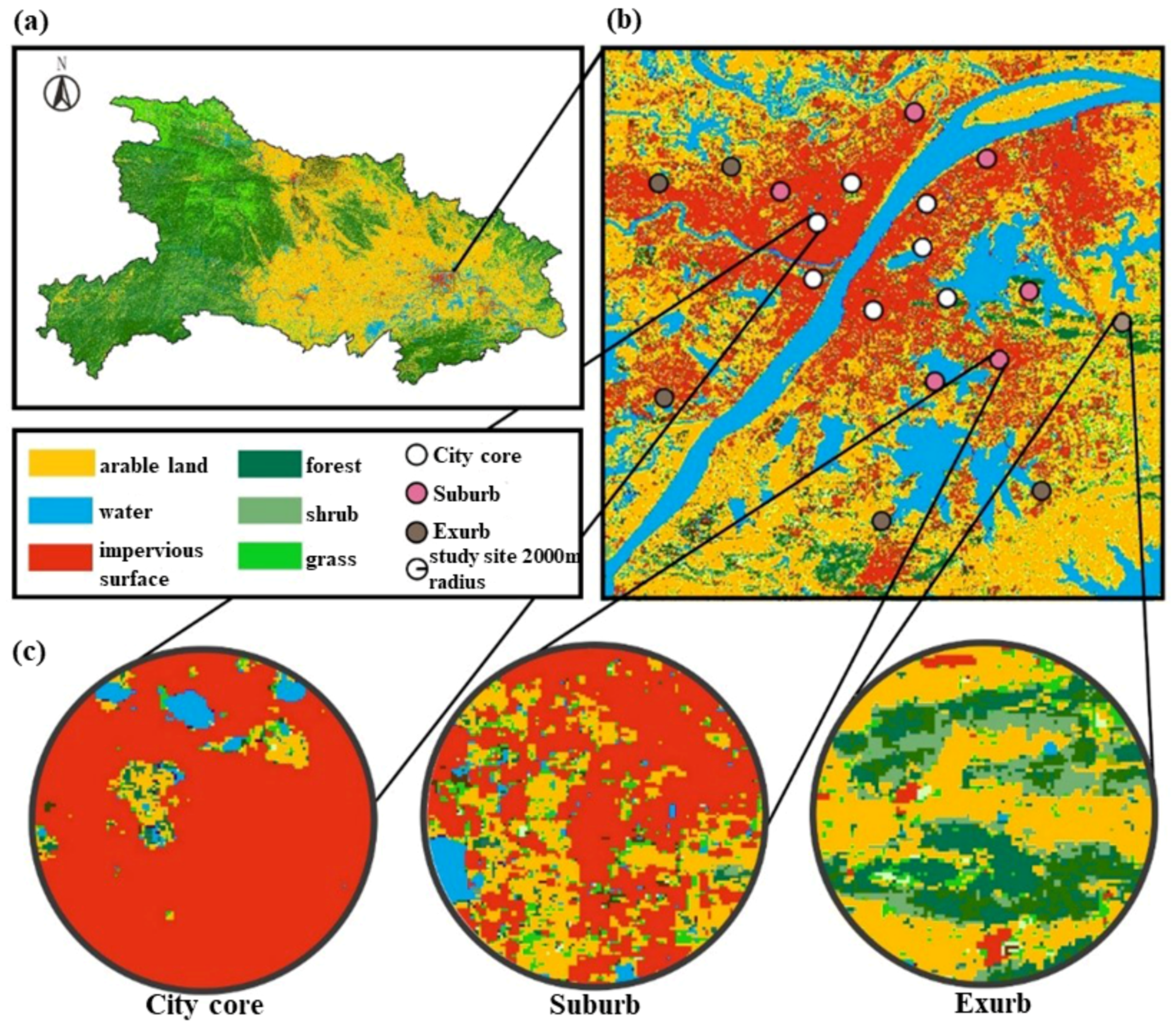
2.1. Study Area
2.2. Landscape Variables
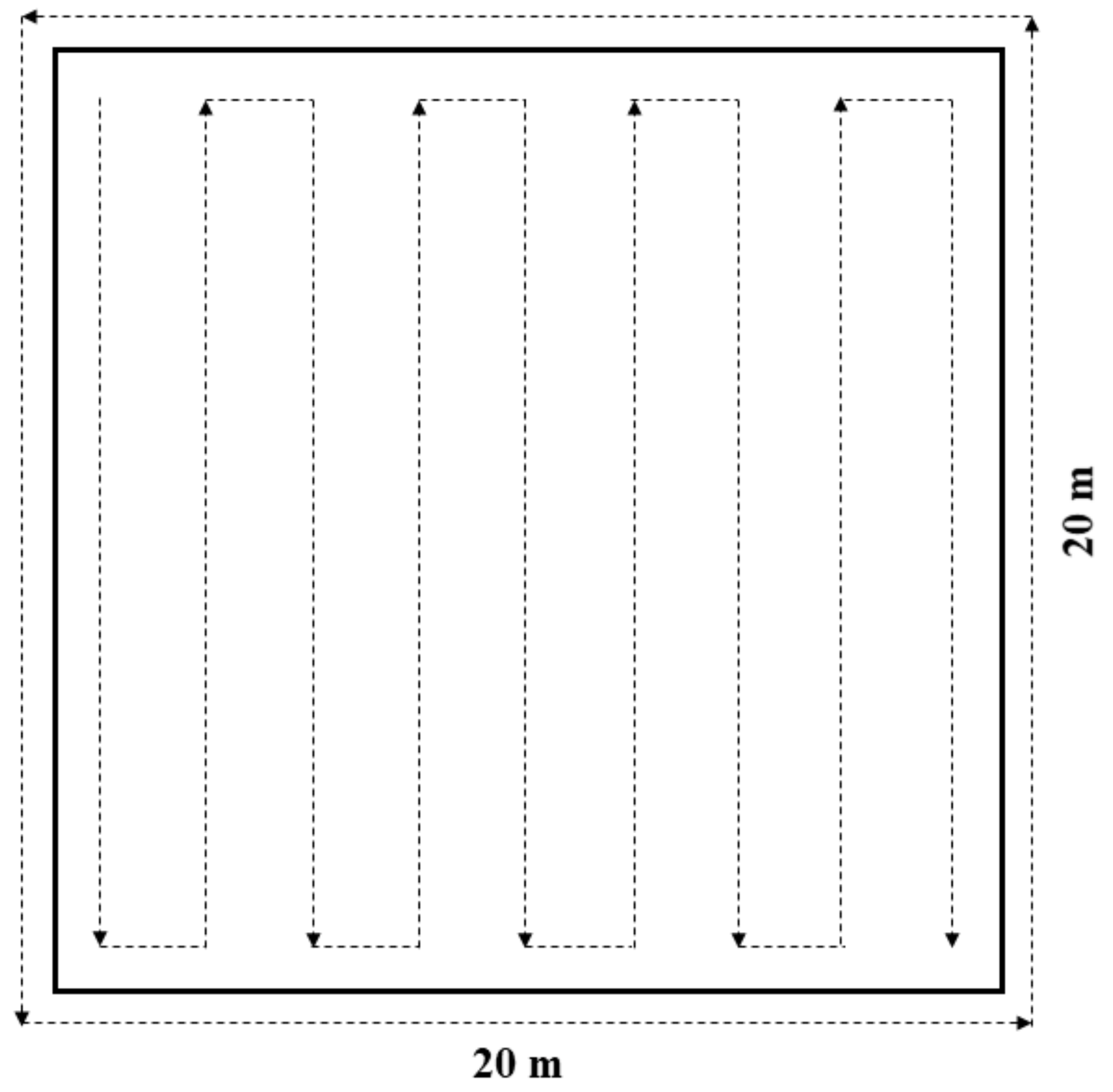
2.3. Local Floral Resources and Flower-Visitors
2.4. Data Analysis
3. Results
3.1. Survey Results
3.2. Factors Influencing the Number of Visits by Flower-Visitor Groups
3.3. Factors Influencing the Composition and Diversity of Flower-Visitor Groups
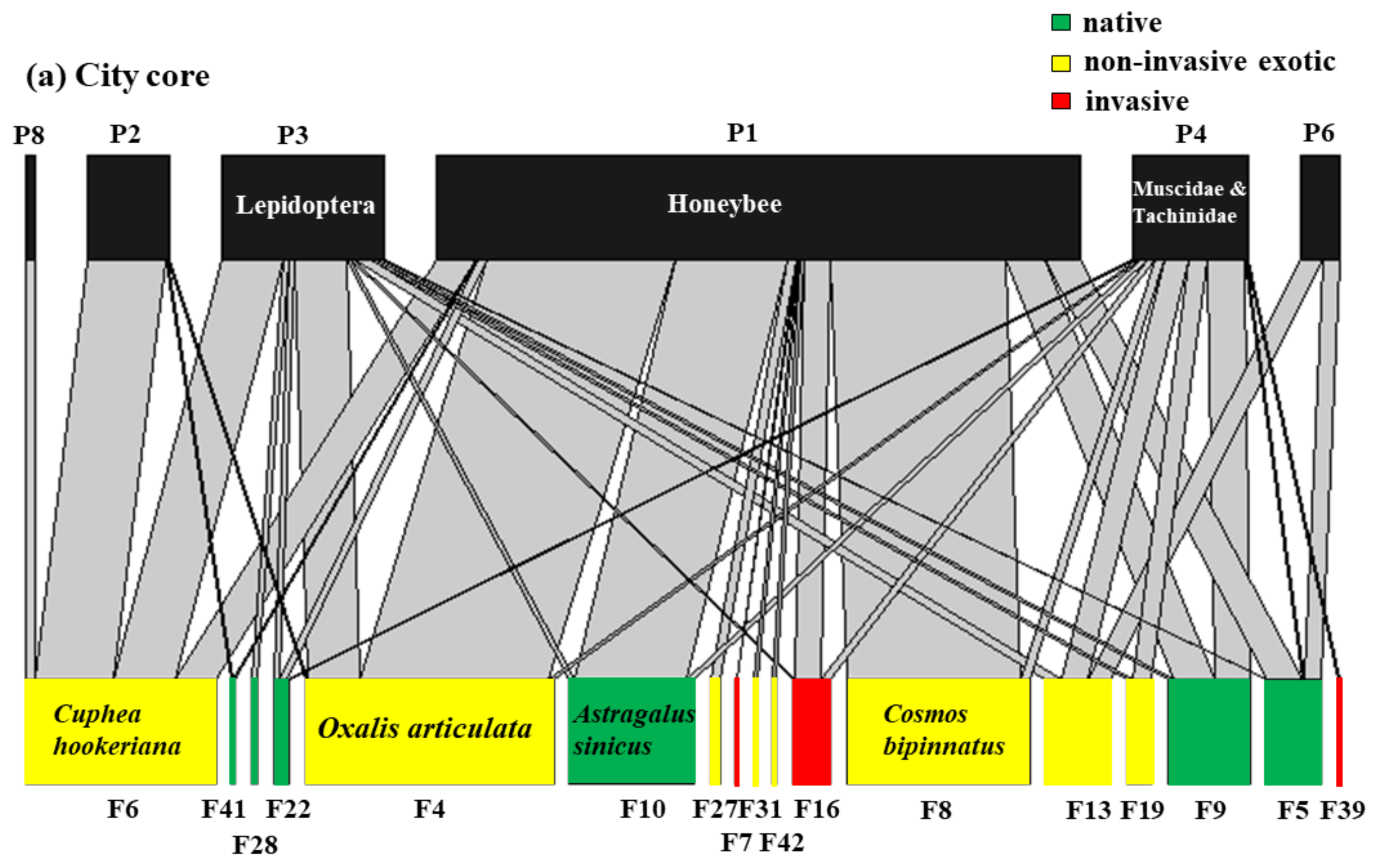
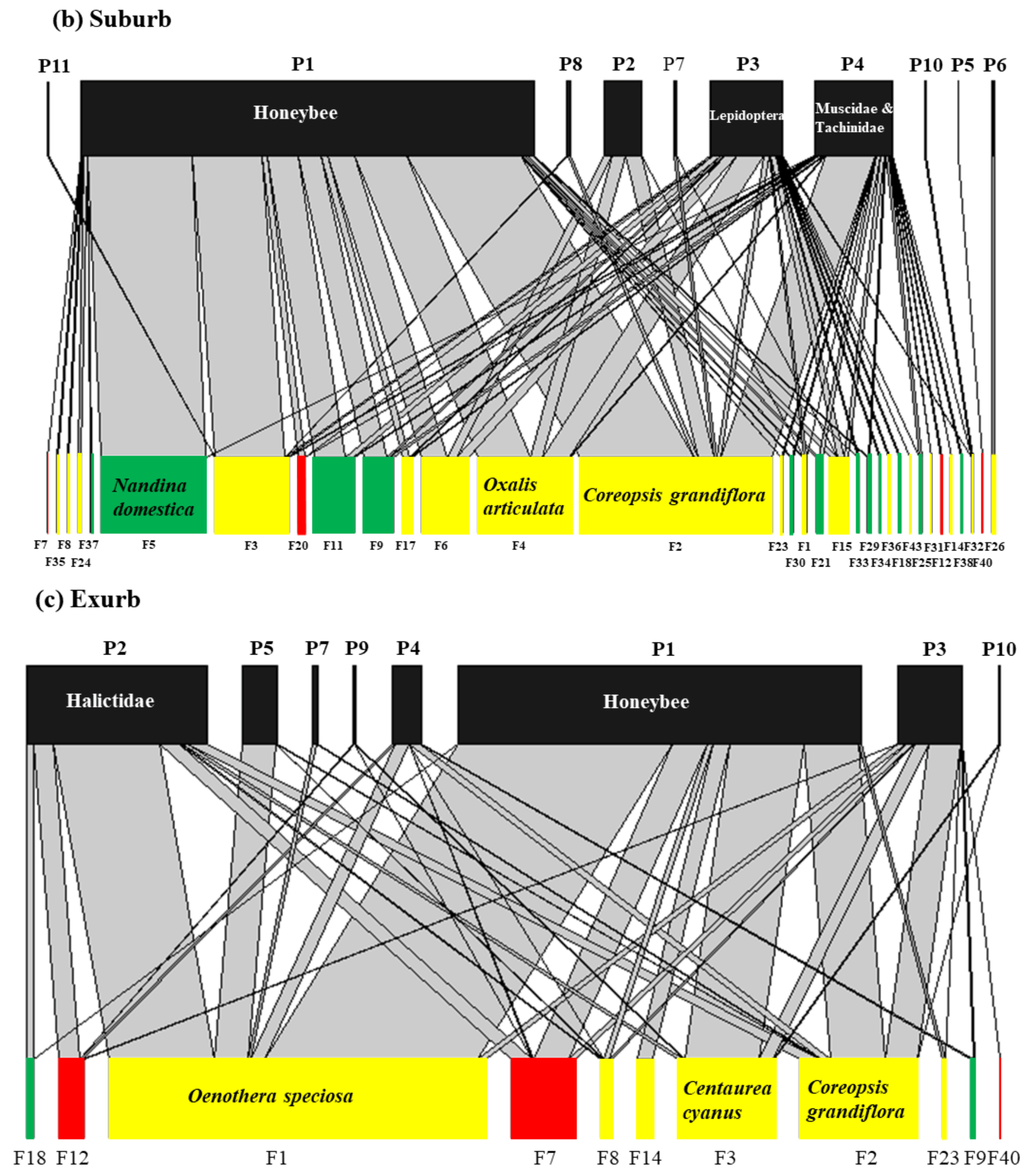
3.4. The Interaction between Flowering Plants and Flower-Visitor Groups under Three Urbanization Levels
4. Discussion
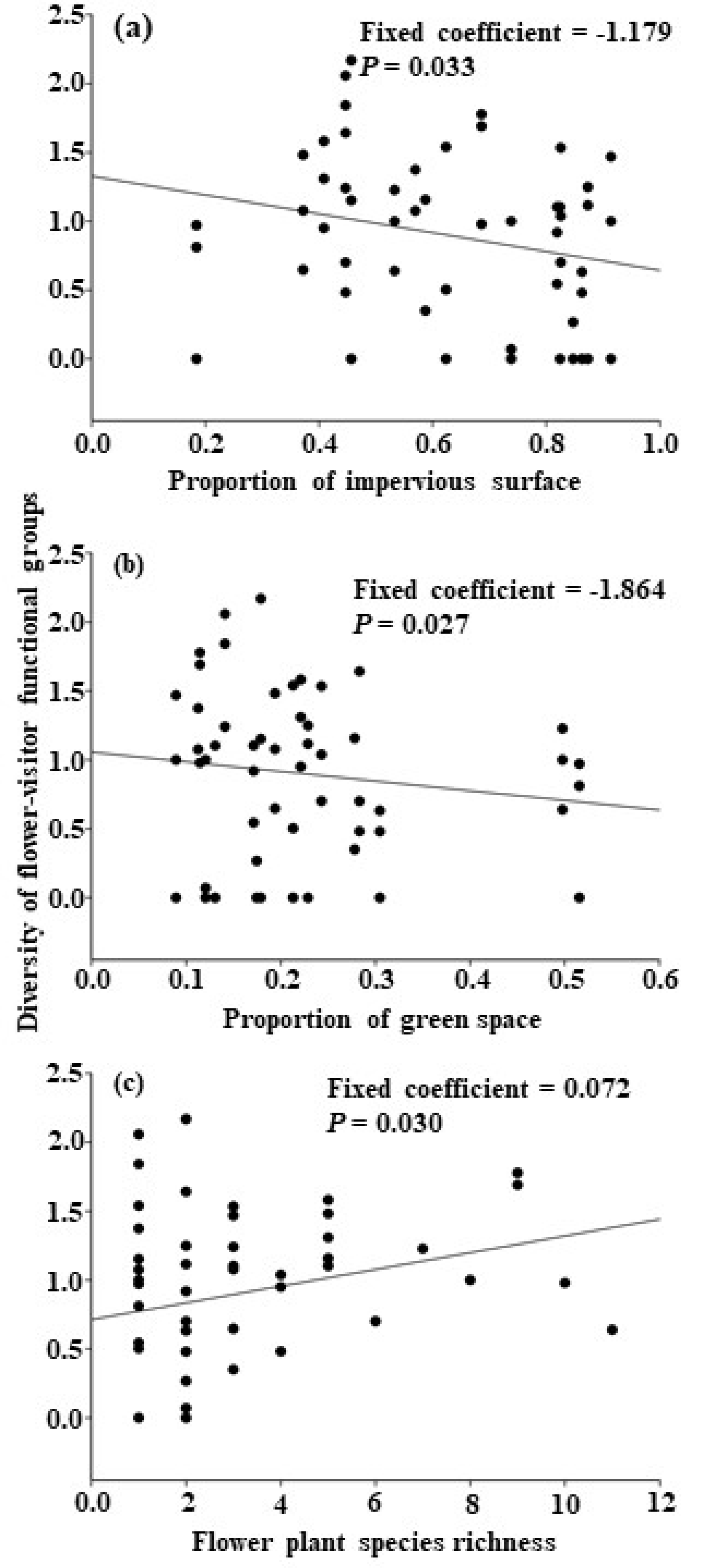
4.1. Effects of the Proportion of Impervious Surface to Flower-Visitor Groups
4.2. Effects of Proportion of Green Space on Flower-Visitor Groups
4.3. Effects of Night Light Intensity on Flower-Visitors
4.4. Effects of Local Flower Density and Flowering Plant Richness to Flower-Visitor Groups
4.5. Effects of Urbanization Level on Flowering Plant–Visitor Group Interactions and Core Species
4.6. Effects of Sampling Month to Flower-Visitor Groups
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study Site | Latitude | Longitude | Proportion of Impervious Surface | Proportion of Greenspace | Average Night Light Intensity (DN) |
|---|---|---|---|---|---|
| City core | |||||
| Jiefang Park | 30.6091 | 114.2911 | 87.35% | 22.87% | 45141 |
| Shahu Park | 30.5758 | 114.3416 | 81.95% | 17.13% | 50630 |
| Luojiashan Parkland | 30.5379 | 114.3561 | 86.32% | 30.47% | 30709 |
| Lianhuahu Park | 30.5517 | 114.2741 | 82.58% | 24.29% | 38430 |
| Shouyi Cultural Park | 30.5433 | 114.3006 | 84.75% | 17.45% | 45896 |
| Zhongshan Park | 30.5858 | 114.2674 | 91.42% | 8.93% | 45881 |
| Simeitang Park | 30.5965 | 114.3313 | 82.46% | 13.07% | 47913 |
| Suburb | |||||
| Shizishan Parkland | 30.4750 | 114.3380 | 58.76% | 27.81% | 25860 |
| Houxianghe Park | 30.6115 | 114.2482 | 68.63% | 11.45% | 47191 |
| Heping Park | 30.6369 | 114.3814 | 62.38% | 21.31% | 28522 |
| Wuhan Botanical Garden | 30.5483 | 114.4160 | 53.32% | 49.76% | 10606 |
| Dijiao Park | 30.6655 | 114.3331 | 56.96% | 11.27% | 21320 |
| Guanshan Holland Park | 30.4964 | 114.3927 | 73.86% | 12.05% | 27054 |
| Exurb | |||||
| Wuhan Garden Expo Park | 30.6209 | 114.2156 | 44.67% | 28.30% | 38582 |
| Shimenfeng Park | 30.5166 | 114.4735 | 18.43% | 51.55% | 16231 |
| Canglongdao Wetland Park | 30.4069 | 114.4188 | 37.20% | 19.38% | 21667 |
| Jiangxia Central Park | 30.3796 | 114.3159 | 44.67% | 14.10% | 24925 |
| Zhuyehai Park | 30.6237 | 114.1607 | 45.68% | 17.89% | 29033 |
| Tanghu Park | 30.4727 | 114.1556 | 40.86% | 22.10% | 22490 |
| Family | Species | Region of Origin | Status | Distribution Sites | Code in Pollination Network |
|---|---|---|---|---|---|
| Amaryllidaceae | Tulbaghia violacea Harv. | South Africa | non-invasive exotic | Suburb (Houxianghe Park, Wuhan Botanical Garden) | F36 |
| Amaryllidaceae | Zephyranthes carinata Herbert | South America | invasive | Suburb (Wuhan Botanical Garden) | F53 |
| Araceae | Acorus calamus L. | China | native | Suburb (Wuhan Botanical Garden) | F34 |
| Asteraceae | Carthamus tinctorius L. | Central Asia | non-invasive exotic | Suburb (Wuhan Botanical Garden) | F35 |
| Asteraceae | Centaurea cyanus L. | Europe, Russia, north America | non-invasive exotic | Suburb, Exurb (Shizishan Parkland, Wuhan Botanical Garden, Canglongdao Wetland Park) | F3 |
| Asteraceae | Cirsium arvense var. integrifolium Wimm. & Grab. | East and north Asia | native | Suburb (Wuhan Botanical Garden) | F11 |
| Asteraceae | Coreopsis grandiflora Hogg ex Sw. | America | non-invasive exotic | Suburb, Exurb (Houxianghe Park, Dijiao Park, Jiangxia Central Park, Tanghu Park) | F2 |
| Asteraceae | Cosmos bipinnatus Cav. | Mexico | non-invasive exotic | City, Suburb, Exurb (Shouyi Cultural Park, Shizishan Parkland, Houxianghe Park, Wuhan Garden Expo Park) | F8 |
| Asteraceae | Erigeron annuus (L.) Pers. | North America | invasive | City, Suburb, Exurb (Jiefang Park, Houxianghe Park, Canglongdao Wetland Park, Jiangxia Central Park, Tanghu Park) | F12 |
| Asteraceae | Euryops pectinatus (L.) Cass. | South Africa | non-invasive exotic | Suburb (Houxianghe Park) | F17 |
| Asteraceae | Helianthus annuus L. | North America | non-invasive exotic | Suburb (Houxianghe Park) | F26 |
| Asteraceae | Leucanthemum vulgare Lam. | West Europe | non-invasive exotic | Suburb, Exurb (Houxianghe Park, Jiangxia Central Park) | F15 |
| Asteraceae | Sanvitalia procumbens Lam. | Mexico | non-invasive exotic | Suburb, Exurb (Shizishan Parkland, Canglongdao Wetland Park) | F23 |
| Asteraceae | Silybum marianum (L.) Gaertn. | Europe, Mediterranean, north Africa, central Asia | invasive | Suburb (Wuhan Botanical Garden) | F48 |
| Asteraceae | Tagetes erecta L. | Mexico | non-invasive exotic | City, Suburb (Shouyi Cultural Park, Houxianghe Park) | F31 |
| Asteraceae | Taraxacum mongolicum Hand.-Mazz. | China, north Korea, Mongolia, Russia | native | City (Shahu Park, Lianhuahu Park) | F28 |
| Berberidaceae | Nandina domestica Thunb. | China | native | City, Suburb (Jiefang Park, Lianhuahu Park, Guanshan Holland Park) | F5 |
| Campanulaceae | Platycodon grandiflorus (Jacq.) A. DC. | East and north Asia | native | Suburb (Wuhan Botanical Garden) | F51 |
| Cannaceae | Canna indica L. | Japan | non-invasive exotic | Suburb (Wuhan Botanical Garden) | F52 |
| Caprifoliaceae | Weigela florida (Bunge) A. DC. | China, japan, india, america | native | City (Shahu Park) | F44 |
| Caprifoliaceae | Zabelia biflora (Turcz.) Makino | China, korea | native | Suburb, Exurb (Wuhan Botanical Garden, Tanghu Park) | F18 |
| Caryophyllaceae | Dianthus chinensis L. | China | native | Suburb (Houxianghe Park) | F29 |
| Caryophyllaceae | Vaccaria hispanica (Miller) Rauschert | Europe, asia | native | Suburb (Shizishan Parkland) | F38 |
| Crassulaceae | Sedum lineare Thunb. | China, japan, vietnam | native | Suburb (Houxianghe Park) | F25 |
| Cruciferae | Orychophragmus violaceus (Linnaeus) O. E. Schulz | China, north Korea | native | Suburb (Shizishan Parkland) | F37 |
| Geraniaceae | Geranium carolinianum L. | America | invasive | City (Lianhuahu Park) | F46 |
| Lamiaceae | Salvia miltiorrhiza Bunge | China, japan | native | Suburb (Wuhan Botanical Garden) | F47 |
| Lamiaceae | Scutellaria barbata D. Don | India, nepal, myanmar, laos, thailand | non-invasive exotic | City (Shouyi Cultural Park) | F42 |
| Leguminosae | Astragalus sinicus L. | China | native | City (Luojiashan Parkland) | F10 |
| Leguminosae | Medicago falcata L. | China | native | City (Shahu Park, Luojiashan Parkland, Lianhuahu Park) | F22 |
| Leguminosae | Trifolium pratense L. | Central Europe | invasive | City (Shahu Park) | F16 |
| Leguminosae | Trifolium repens L. | Europe, north Africa | invasive | City, Suburb, Exurb (Simeitang Park, Houxianghe Park, Zhuyehai Park, Tanghu Park) | F7 |
| Liliaceae | Hemerocallis fulva (L.) L. | China, south Europe | native | Suburb (Wuhan Botanical Garden) | F33 |
| Liliaceae | Ophiopogon japonicus (L. f.) Ker-Gawl. | China, japan, vietnam, india | native | City (Shahu Park) | F50 |
| Lythraceae | Cuphea hookeriana Walp. | Mexico | non-invasive exotic | City, Suburb (Lianhuahu Park, Houxianghe Park) | F6 |
| Malvaceae | Alcea rosea Linnaeus | China | native | Suburb (Wuhan Botanical Garden) | F30 |
| Oleaceae | Ligustrum sinense Lour. | China | native | Suburb (Houxianghe Park) | F21 |
| Onagraceae | Oenothera biennis L. | North America | invasive | Suburb (Wuhan Botanical Garden) | F20 |
| Onagraceae | Oenothera speciosa Nutt. | America | non-invasive exotic | Suburb, Exurb (Shizishan Parkland, Wuhan Botanical Garden, Wuhan Garden Expo Park, Zhuyehai Park, Tanghu Park) | F1 |
| Oxalidaceae | Oxalis articulata Savigny | South America | non-invasive exotic | City, Suburb (Shahu Park, Lianhuahu Park, Simeitang Park, Wuhan Botanical Garden | F4 |
| Oxalidaceae | Oxalis corniculata L. | Temperate and subtropical Asia, Europe, Mediterranean, north America | native | City (Lianhuahu Park) | F49 |
| Papaveraceae | Papaver rhoeas L. | Europe | non-invasive exotic | Suburb, Exurb (Shizishan Parkland, Wuhan Garden Expo Park, Canglongdao Wetland Park) | F14 |
| Plantaginaceae | Plantago depressa Willd. | China, north Korea, Russia, Kazakhstan | native | Exurb (Canglongdao Wetland Park) | F45 |
| Plantaginaceae | Veronica persica Poir. | Western Asia, Europe | invasive | City, Suburb, Exurb (Lianhuahu Park, Simeitang Park, Wuhan Botanical Garden, Wuhan Garden Expo Park) | F39 |
| Portulacaceae | Portulaca grandiflora Hook. | Brazil | non-invasive exotic | City (Shouyi Cultural Park) | F27 |
| Rosaceae | Kerria japonica (L.) DC. | China, japan | native | City (Lianhuahu Park) | F41 |
| Rosaceae | Rosa chinensis Jacq. | China | native | City, Suburb, Exurb (Jiefang Park, Zhongshan Park, Houxianghe Park, Heping Park, Guanshan Holland Park, Shimenfeng Park) | F9 |
| Saxifragaceae | Heuchera micrantha Douglas | Cenral america | non-invasive exotic | Suburb (Houxianghe Park) | F43 |
| Solanaceae | Petunia × atkinsiana D. Don ex Loudon | South America | non-invasive exotic | City (Zhongshan Park | F19 |
| Umbelliferae | Coriandrum sativum L. | Mediterranean | non-invasive exotic | Suburb (Wuhan Botanical Garden) | F24 |
| Umbelliferae | Daucus carota L. | Europe | invasive | Suburb, Exurb (Wuhan Botanical Garden, Canglongdao Wetland Park) | F40 |
| Verbenaceae | Glandularia × hybrida (Groenland & Rümpler) G. L.Nesom & Pruski | Panama, honduras, venezuela | non-invasive exotic | Suburb (Houxianghe Park) | F32 |
| Violaceae | Viola cornuta Desf. | Europe | non-invasive exotic | City (Zhongshan Park) | F13 |
| Code in Network | Functional Groups | Number of Links | Number of Linked Plant Species | Normalised Degree |
|---|---|---|---|---|
| P1 | Honeybee | 2534 | 30 | 0.6977 |
| P2 | Halictidae | 625 | 10 | 0.2326 |
| P3 | Lepidoptera | 449 | 29 | 0.6744 |
| P4 | Muscidae and Tachinidae | 335 | 28 | 0.6512 |
| P5 | Formicidae | 92 | 4 | 0.0930 |
| P6 | Anthophora spp. | 35 | 3 | 0.0698 |
| P7 | Coleoptera | 17 | 3 | 0.0698 |
| P8 | Syrphidae | 15 | 3 | 0.0698 |
| P9 | Vespidae and Ichneumonidae | 6 | 3 | 0.0698 |
| P10 | Odonata | 5 | 3 | 0.0698 |
| P11 | Orthoptera | 2 | 1 | 0.0233 |
References
- Garibaldi, L.A.; Steffan-Dewenter, I.; Winfree, R.; Aizen, M.A.; Bommarco, R.; Cunningham, S.A.; Kremen, C.; Carvalheiro, L.G.; Harder, L.D.; Afik, O.; et al. Wild Pollinators Enhance Fruit Set of Crops Regardless of Honey Bee Abundance. Science 2013, 339, 1608–1611. [Google Scholar] [CrossRef] [PubMed]
- Goulson, D.; Lye, G.C.; Darvill, B. Decline and Conservation of Bumble Bees. Annu. Rev. Ѐntomol. 2008, 53, 191–208. [Google Scholar] [CrossRef] [PubMed]
- Bartomeus, I.; Ascher, J.S.; Gibbs, J.; Danforth, B.N.; Wagner, D.L.; Hedtke, S.M.; Winfree, R. Historical changes in northeastern US bee pollinators related to shared ecological traits. Proc. Natl. Acad. Sci. USA 2013, 110, 4656–4660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carvalheiro, L.G.; Kunin, W.E.; Keil, P.; Aguirre-Gutiérrez, J.; Ellis, W.N.; Fox, R.; Groom, Q.; Hennekens, S.; Van Landuyt, W.; Maes, D.; et al. Species richness declines and biotic homogenisation have slowed down for NW-European pollinators and plants. Ecol. Lett. 2013, 16, 870–878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goulson, D.; Nicholls, E.; Botías, C.; Rotheray, E.L. Bee declines driven by combined stress from parasites, pesticides, and lack of flowers. Science 2015, 347, 1255957. [Google Scholar] [CrossRef] [PubMed]
- Johansson, V.; Gustafsson, L.; Andersson, P.; Hylander, K. Fewer butterflies and a different composition of bees, wasps and hoverflies on recently burned compared to unburned clear-cuts, regardless of burn severity. For. Ecol. Manag. 2020, 463, 118033. [Google Scholar] [CrossRef]
- Hallmann, C.A.; Sorg, M.; Jongejans, E.; Siepel, H.; Hofland, N.; Schwan, H.; Stenmans, W.; Müller, A.; Sumser, H.; Hörren, T.; et al. More than 75 percent decline over 27 years in total flying insect biomass in protected areas. PLoS ONE 2017, 12, e0185809. [Google Scholar] [CrossRef] [Green Version]
- Cameron, S.A.; Lozier, J.D.; Strange, J.P.; Koch, J.B.; Cordes, N.; Solter, L.F.; Griswold, T.L. Patterns of widespread decline in North American bumble bees. Proc. Natl. Acad. Sci. USA 2011, 108, 662–667. [Google Scholar] [CrossRef] [Green Version]
- Potts, S.G.; Biesmeijer, J.C.; Kremen, C.; Neumann, P.; Schweiger, O.; Kunin, W.E. Global pollinator declines: Trends, impacts and drivers. Trends Ecol. Evol. 2010, 25, 345–353. [Google Scholar] [CrossRef]
- González-Varo, J.P.; Biesmeijer, J.C.; Bommarco, R.; Potts, S.G.; Schweiger, O.; Smith, H.G.; Steffan-Dewenter, I.; Szentgyörgyi, H.; Woyciechowski, M.; Vilà, M. Combined effects of global change pressures on animal-mediated pollination. Trends Ecol. Evol. 2013, 28, 524–530. [Google Scholar] [CrossRef] [Green Version]
- Wagner, D.L.; Van Driesche, R.G. Threats Posed to Rare or Endangered Insects by Invasions of Nonnative Species. Annu. Rev. Ѐntomol. 2010, 55, 547–568. [Google Scholar] [CrossRef] [PubMed]
- Ahrné, K.; Bengtsson, J.; Elmqvist, T. Bumble Bees (Bombus spp.) along a gradient of increasing urbanization. PLoS ONE 2009, 4, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.; Li, J.; Kuang, S.; He, Y.; Chen, G.; Huang, Y.; Song, C.; Anderson, P.; Łowicki, D. Plant Diversity along the Urban-Rural Gradient and Its Relationship with Urbanization Degree in Shanghai, China. Forests 2020, 11, 171. [Google Scholar] [CrossRef] [Green Version]
- McKinney, M.L. Urbanization, Biodiversity, and Conservation. Bioscience 2002, 52, 883–888. [Google Scholar] [CrossRef]
- Tscharntke, T.; Klein, A.M.; Kruess, A.; Steffan-Dewenter, I.; Thies, C. Landscape perspectives on agricultural intensification and biodiversity—Ecosystem service management. Ecol. Lett. 2005, 8, 857–874. [Google Scholar] [CrossRef]
- Geslin, B.; Gauzens, B.; Thébault, E.; Dajoz, I. Plant Pollinator Networks along a Gradient of Urbanisation. PLoS ONE 2013, 8, e63421. [Google Scholar] [CrossRef]
- Fortel, L.; Henry, M.; Guilbaud, L.; Guirao, A.L.; Kuhlmann, M.; Mouret, H.; Rollin, O.; Vaissière, B.E. Decreasing Abundance, Increasing Diversity and Changing Structure of the Wild Bee Community (Hymenoptera: Anthophila) along an Urbanization Gradient. PLoS ONE 2014, 9, e104679. [Google Scholar] [CrossRef]
- Winfree, R.; Aguilar, R.; Vázquez, D.P.; Lebuhn, G.; Aizen, M.A. A meta-analysis of bees’ responses to anthropogenic disturbance. Ecology 2009, 90, 2068–2076. [Google Scholar] [CrossRef] [PubMed]
- Hausmann, S.L.; Petermann, J.; Rolff, J. Wild bees as pollinators of city trees. Insect Conserv. Divers. 2016, 9, 97–107. [Google Scholar] [CrossRef]
- Theodorou, P.; Radzeviciute, R.; Settele, J.; Schweiger, O.; Murray, T.E.; Paxton, R.J. Pollination services enhanced with urbanization despite increasing pollinator parasitism. Proc. R. Soc. B Boil. Sci. 2016, 283, 20160561. [Google Scholar] [CrossRef] [Green Version]
- Goulson, D.; Hughes, W.; Derwent, L.C.; Stout, J. Colony growth of the bumblebee, Bombus terrestris, in improved and conventional agricultural and suburban habitats. Oecologia 2002, 130, 267–273. [Google Scholar] [CrossRef] [PubMed]
- Osborne, J.L.; Martin, A.P.; Shortall, C.R.; Todd, A.D.; Goulson, D.; Knight, M.E.; Hale, R.J.; Sanderson, R.A. Quantifying and comparing bumblebee nest densities in gardens and countryside habitats. J. Appl. Ecol. 2008, 45, 784–792. [Google Scholar] [CrossRef]
- Goulson, D.; Lepais, O.; O’Connor, S.; Osborne, J.L.; Sanderson, R.A.; Cussans, J.; Goffe, L.; Darvill, B. Effects of land use at a landscape scale on bumblebee nest density and survival. J. Appl. Ecol. 2010, 47, 1207–1215. [Google Scholar] [CrossRef] [Green Version]
- Majewska, A.A.; Altizer, S. Planting gardens to support insect pollinators. Conserv. Biol. 2018, 34, 15–25. [Google Scholar] [CrossRef]
- Luppi, M.; Dondina, O.; Orioli, V.; Bani, L. Local and landscape drivers of butterfly richness and abundance in a human-dominated area. Agric. Ecosyst. Environ. 2018, 254, 138–148. [Google Scholar] [CrossRef]
- Bubová, T.; Vrabec, V.; Kulma, M.; Nowicki, P. Land management impacts on European butterflies of conservation concern: A review. J. Insect Conserv. 2015, 19, 805–821. [Google Scholar] [CrossRef] [Green Version]
- Knop, E.; Zoller, L.; Ryser, R.; Gerpe, C.; Hörler, M.; Fontaine, C. Artificial light at night as a new threat to pollination. Nat. 2017, 548, 206–209. [Google Scholar] [CrossRef]
- Giavi, S.; Fontaine, C.; Knop, E. Impact of artificial light at night on diurnal plant-pollinator interactions. Nat. Commun. 2021, 12, 1690. [Google Scholar] [CrossRef]
- Venjakob, C.; Klein, A.-M.; Ebeling, A.; Tscharntke, T.; Scherber, C. Plant diversity increases spatio-temporal niche complementarity in plant-pollinator interactions. Ecol. Evol. 2016, 6, 2249–2261. [Google Scholar] [CrossRef] [Green Version]
- Garbuzov, M.; Ratnieks, F.L.W. Quantifying variation among garden plants in attractiveness to bees and other flower-visiting insects. Funct. Ecol. 2014, 28, 364–374. [Google Scholar] [CrossRef]
- Tasker, P.; Reid, C.; Young, A.D.; Threlfall, C.G.; Latty, T. If you plant it, they will come: Quantifying attractiveness of exotic plants for winter-active flower visitors in community gardens. Urban Ecosyst. 2020, 23, 345–354. [Google Scholar] [CrossRef]
- Gong, Y.-B.; Huang, S.-Q. Temporal stability of pollinator preference in an alpine plant community and its implications for the evolution of floral traits. Oecologia 2011, 166, 671–680. [Google Scholar] [CrossRef] [PubMed]
- Wuhan Natural Resources and Planning Bureau. Wuhan’s General Plan for Land Use (2006–2020). 2011. Available online: http://zrzyhgh.wuhan.gov.cn/ (accessed on 20 January 2022).
- Gong, P.; Liu, H.; Zhang, M.; Li, C.; Wang, J.; Huang, H.; Clinton, N.; Ji, L.; Li, W.; Bai, Y.; et al. Stable classification with limited sample: Transferring a 30-m resolution sample set collected in 2015 to mapping 10-m resolution global land cover in 2017. Sci. Bull. 2019, 64, 370–373. [Google Scholar] [CrossRef] [Green Version]
- Hagen, M.; Wikelski, M.; Kissling, W.D. Space Use of Bumblebees (Bombus spp.) Revealed by Radio-Tracking. PLoS ONE 2011, 6, e19997. [Google Scholar] [CrossRef] [PubMed]
- Glaum, P.; Simao, M.-C.; Vaidya, C.; Fitch, G.; Iuliano, B. Big city Bombus: Using natural history and land-use history to find significant environmental drivers in bumble-bee declines in urban development. R. Soc. Open Sci. 2017, 4, 170156. [Google Scholar] [CrossRef] [Green Version]
- Xie, Q.J.; Chen, K.L.; Jin, G. Quantifying urbanization and its impact on urban heat island effect in Wuhan. Sci. Surv. Mapp. 2017, 42, 71–76+87. (In Chinese) [Google Scholar]
- McKinney, M.L. Effects of urbanization on species richness: A review of plants and animals. Urban Ecosyst. 2008, 11, 161–176. [Google Scholar] [CrossRef]
- Baldock, K.C.R.; Goddard, M.; Hicks, D.M.; Kunin, W.E.; Mitschunas, N.; Osgathorpe, L.M.; Potts, S.G.; Robertson, K.M.; Scott, A.V.; Stone, G.; et al. Where is the UK’s pollinator biodiversity? The importance of urban areas for flower-visiting insects. Proc. R. Soc. B Boil. Sci. 2015, 282, 20142849. [Google Scholar] [CrossRef] [Green Version]
- Catalogue of Life. Available online: https://www.catalogueoflife.org/ (accessed on 20 January 2022).
- Invasive Alien Species of China (ISAC). Available online: https://www.plantplus.cn/ias/protlist (accessed on 20 January 2022).
- Collado, M.Á.; Sol, D.; Bartomeus, I. Bees use anthropogenic habitats despite strong natural habitat preferences. Divers. Distrib. 2019, 25, 924–935. [Google Scholar] [CrossRef] [Green Version]
- Ollerton, J.; Waser, N.M. Plant-Pollinator Interactions: From Specialization to Generalization; University of Chicago Press: Chicago, IL, USA, 2006. [Google Scholar]
- Klein, S.; Pasquaretta, C.; He, X.J.; Perry, C.; Søvik, E.; Devaud, J.-M.; Barron, A.B.; Lihoreau, M. Honey bees increase their foraging performance and frequency of pollen trips through experience. Sci. Rep. 2019, 9, 6778. [Google Scholar] [CrossRef]
- Gathmann, A.; Tscharntke, T. Foraging ranges of solitary bees. J. Anim. Ecol. 2002, 71, 757–764. [Google Scholar] [CrossRef]
- Zurbuchen, A.; Cheesman, S.; Klaiber, J.; Müller, A.; Hein, S.; Dorn, S. Long foraging distances impose high costs on offspring production in solitary bees. J. Anim. Ecol. 2010, 79, 674–681. [Google Scholar] [CrossRef]
- Le Féon, V.; Schermann-Legionnet, A.; Delettre, Y.; Aviron, S.; Billeter, R.; Bugter, R.; Hendrickx, F.; Burel, F. Intensification of agriculture, landscape composition and wild bee communities: A large scale study in four European countries. Agric. Ecosyst. Environ. 2010, 137, 143–150. [Google Scholar] [CrossRef]
- Abbate, A.; Campbell, J.W.; Kimmel, C.B.; Kern, W.H. Urban development decreases bee abundance and diversity within coastal dune systems. Glob. Ecol. Conserv. 2019, 20, e00711. [Google Scholar] [CrossRef]
- Burdine, J.D.; McCluney, K.E. Interactive effects of urbanization and local habitat characteristics influence bee communities and flower visitation rates. Oecologia 2019, 190, 715–723. [Google Scholar] [CrossRef]
- Persson, A.S.; Ekroos, J.; Olsson, P.; Smith, H.G. Wild bees and hoverflies respond differently to urbanisation, human population density and urban form. Landsc. Urban Plan. 2020, 204, 103901. [Google Scholar] [CrossRef]
- Kratschmer, S.; Pachinger, B.; Schwantzer, M.; Paredes, D.; Guernion, M.; Burel, F.; Nicolai, A.; Strauss, P.; Bauer, T.; Kriechbaum, M.; et al. Tillage intensity or landscape features: What matters most for wild bee diversity in vineyards? Agric. Ecosyst. Environ. 2018, 266, 142–152. [Google Scholar] [CrossRef]
- Beyer, N.; Gabriel, D.; Kirsch, F.; Schulz-Kesting, K.; Dauber, J.; Westphal, C. Functional groups of wild bees respond differently to faba bean Vicia faba L. cultivation at landscape scale. J. Appl. Ecol. 2020, 57, 2499–2508. [Google Scholar] [CrossRef]
- Garibaldi, L.A.; Steffan-Dewenter, I.; Kremen, C.; Morales, J.M.; Bommarco, R.; Cunningham, S.; Carvalheiro, L.G.; Chacoff, N.P.; Dudenhöffer, J.-H.; Greenleaf, S.S.; et al. Stability of pollination services decreases with isolation from natural areas despite honey bee visits. Ecol. Lett. 2011, 14, 1062–1072. [Google Scholar] [CrossRef]
- Rader, R.; Bartomeus, I.; Garibaldi, L.A.; Garratt, M.P.D.; Howlett, B.G.; Winfree, R.; Cunningham, S.A.; Mayfield, M.M.; Arthur, A.D.; Andersson, G.K.S.; et al. Non-bee insects are important contributors to global crop pollination. Proc. Natl. Acad. Sci. USA 2016, 113, 146–151. [Google Scholar] [CrossRef] [Green Version]
- Threlfall, C.G.; Walker, K.; Williams, N.S.; Hahs, A.K.; Mata, L.; Stork, N.; Livesley, S.J. The conservation value of urban green space habitats for Australian native bee communities. Biol. Conserv. 2015, 187, 240–248. [Google Scholar] [CrossRef]
- MacGregor, C.J.; Pocock, M.J.O.; Fox, R.; Evans, D.M. Pollination by nocturnal L epidoptera, and the effects of light pollution: A review. Ecol. Ѐntomol. 2015, 40, 187–198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghazoul, J. Floral diversity and the facilitation of pollination. J. Ecol. 2006, 94, 295–304. [Google Scholar] [CrossRef]
- Blaauw, B.R.; Isaacs, R. Larger patches of diverse floral resources increase insect pollinator density, diversity, and their pollination of native wildflowers. Basic Appl. Ecol. 2014, 15, 701–711. [Google Scholar] [CrossRef]
- Vrdoljak, S.M.; Samways, M.J.; Simaika, J.P. Pollinator conservation at the local scale: Flower density, diversity and community structure increase flower visiting insect activity to mixed floral stands. J. Insect Conserv. 2016, 20, 711–721. [Google Scholar] [CrossRef]
- Myers, M.C.; Hoksch, B.J.; Mason, J.T. Butterfly response to floral resources during early establishment at a heterogeneous prairie biomass production site in Iowa, USA. J. Insect Conserv. 2012, 16, 457–472. [Google Scholar] [CrossRef]
- Theodorou, P.; Albig, K.; Radzeviciute, R.; Settele, J.; Schweiger, O.; Murray, T.E.; Paxton, R. The structure of flower visitor networks in relation to pollination across an agricultural to urban gradient. Funct. Ecol. 2017, 31, 838–847. [Google Scholar] [CrossRef]
- Senapathi, D.; Goddard, M.; Kunin, W.E.; Baldock, K.C.R. Landscape impacts on pollinator communities in temperate systems: Evidence and knowledge gaps. Funct. Ecol. 2017, 31, 26–37. [Google Scholar] [CrossRef] [Green Version]
- Stelzer, R.J.; Chittka, L.; Carlton, M.; Ings, T.C. Winter Active Bumblebees (Bombus terrestris) Achieve High Foraging Rates in Urban Britain. PLoS ONE 2010, 5, e9559. [Google Scholar] [CrossRef] [Green Version]
- Grass, I.; Berens, D.G.; Peter, F.; Farwig, N. Additive effects of exotic plant abundance and land-use intensity on plant–pollinator interactions. Oecologia 2013, 173, 913–923. [Google Scholar] [CrossRef]
- Stout, J.C.; Tiedeken, E.J. Direct interactions between invasive plants and native pollinators: Evidence, impacts and approaches. Funct. Ecol. 2017, 31, 38–46. [Google Scholar] [CrossRef] [Green Version]
- Hansen, S.; Roets, F.; Seymour, C.L.; Thébault, E.; Van Veen, F.F.; Pryke, J.S. Alien plants have greater impact than habitat fragmentation on native insect flower visitation networks. Divers. Distrib. 2018, 24, 58–68. [Google Scholar] [CrossRef] [Green Version]






| Factor | Scale | Description | Level |
|---|---|---|---|
| Proportion of impervious surface | r = 2000 m | Proportion of impervious surface around the park, as an indicator of urbanization level | Continuous variable, 0–100% |
| Proportion of green space | r = 2000 m | Proportion of green space around the study site, including parkland, protective green space, affiliated green space, regional green space, and other types | Continuous variable, 0–100% |
| Night light intensity | r = 2000 m | Average brightness of night light around the sites | Continuous variable |
| Density of flowers | 20 m × 20 m plot | Number of flowers per m2 | Continuous variable |
| Species richness of flowering plant | 20 m × 20 m plot | Number of flowering plant species | Discrete variable |
| Sampling time | The month of field survey | Nominal variable, May, June, July |
| Term | Factor | Fixed Coefficient | F | p |
|---|---|---|---|---|
| All flower-visitors | Proportion of impervious surface | −1.579 | 0.666 | 0.419 |
| Sampling time | — | 3.365 | 0.043 * | |
| Proportion of green space | −2.556 | 0.772 | 0.384 | |
| Night light intensity | <−0.001 | 0.176 | 0.677 | |
| Local flower density | 0.001 | 0.155 | 0.696 | |
| Flowering plant species richness | 0.154 | 2.001 | 0.164 | |
| Non-honeybee visitors | Proportion of impervious surface | −2.309 | 1.410 | 0.241 |
| Sampling time | — | 0.338 | 0.715 | |
| Proportion of green space | −4.499 | 2.372 | 0.131 | |
| Night light intensity | <−0.001 | 0.530 | 0.471 | |
| Local flower density | 0.003 | 1.021 | 0.318 | |
| Flowering plant species richness | 0.186 | 2.913 | 0.095 | |
| Honeybee | Proportion of impervious surface | −2.067 | 1.722 | 0.196 |
| Sampling time | — | 7.554 | 0.001 *** | |
| Proportion of green space | 0.250 | 0.011 | 0.917 | |
| Night light intensity | <0.001 | 0.121 | 0.730 | |
| Local flower density | −0.001 | 0.037 | 0.848 | |
| Flowering plant species richness | 0.119 | 1.601 | 0.212 | |
| Halictidae | Proportion of impervious surface | 1.121 | 0.152 | 0.699 |
| Sampling time | — | 1.288 | 0.286 | |
| Proportion of green space | −1.607 | 0.134 | 0.716 | |
| Night light intensity | <−0.001 | 0.043 | 0.837 | |
| Local flower density | 0.004 | 47.175 | <0.001 *** | |
| Flowering plant species richness | −0.116 | 0.448 | 0.507 | |
| Lepidoptera | Proportion of impervious surface | −1.108 | 0.265 | 0.609 |
| Sampling time | — | 0.525 | 0.595 | |
| Proportion of green space | −3.400 | 1.101 | 0.300 | |
| Night light intensity | <−0.001 | 0.641 | 0.428 | |
| Local flower density | 0.004 | 1.673 | 0.202 | |
| Flowering plant species richness | 0.298 | 6.317 | 0.016 * | |
| Muscidae | Proportion of impervious surface | −1.023 | 0.348 | 0.558 |
| Sampling time | — | 10.366 | <0.001 *** | |
| Proportion of green space | −5.384 | 4.393 | 0.042 * | |
| Night light intensity | <−0.001 | 2.087 | 0.155 | |
| Local flower density | 0.002 | 0.364 | 0.549 | |
| Flowering plant species richness | −0.005 | 0.003 | 0.956 |
| Factor | Fixed Coefficient | F | p |
|---|---|---|---|
| Proportion of impervious surface | −1.179 | 4.847 | 0.033 * |
| Sampling time | — | 0.172 | 0.842 |
| Proportion of green space | −1.864 | 5.223 | 0.027 * |
| Night light intensity | <0.001 | 0.008 | 0.929 |
| Local flower density | <0.001 | 0.199 | 0.658 |
| Flowering plant species richness | 0.072 | 5.005 | 0.030 * |
| Metrics | City | Suburb | Exurb | Total |
|---|---|---|---|---|
| Functional groups of flower-visitors | 6 | 10 | 8 | 11 |
| Plant Species | 18 | 34 | 13 | 43 |
| Links | 754 | 1433 | 1928 | 4115 |
| Max links of flower-visitor groups | 462 | 994 | 1078 | 2534 |
| Max links of plants | 177 | 420 | 1009 | 1018 |
| Abundance of links | 39 | 70 | 40 | 117 |
| Abundance of potential links | 126 | 340 | 104 | 473 |
| Connectance | 0.31 | 0.21 | 0.38 | 0.24 |
| Matrix temperature | 10.80 | 5.16 | 12.12 | 6.25 |
| Nestedness | 0.89 | 0.95 | 0.88 | 0.94 |
| Code in Network | Plant Species | Number of Links | Number of Linked Pollinator Groups | Normalized Degree |
|---|---|---|---|---|
| F5 | Nandina domestica | 269 | 4 | 0.3636 |
| F9 | Rosa chinensis | 135 | 3 | 0.2727 |
| F10 | Astragalus sinicus | 89 | 3 | 0.2727 |
| F11 | Cirsium arvense var. integrifolium | 87 | 3 | 0.2727 |
| F18 | Zabelia biflora | 19 | 3 | 0.2727 |
| F21 | Ligustrum sinense | 14 | 2 | 0.1818 |
| F22 | Medicago falcata | 12 | 3 | 0.2727 |
| F25 | Sedum lineare | 7 | 2 | 0.1818 |
| F28 | Taraxacum mongolicum | 4 | 1 | 0.0909 |
| F29 | Dianthus chinensis | 4 | 2 | 0.1818 |
| F30 | Alcea rosea Linnaeus | 4 | 2 | 0.1818 |
| F33 | Hemerocallis fulva | 3 | 2 | 0.1818 |
| F34 | Acorus calamus | 3 | 1 | 0.0909 |
| F37 | Orychophragmus violaceus | 2 | 1 | 0.0909 |
| F38 | Vaccaria hispanica | 2 | 1 | 0.0909 |
| F41 | Kerria japonica | 2 | 2 | 0.1818 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, M.; Ran, N.; Jiang, H.; Han, Z.; Dian, Y.; Li, X.; Xie, D.; Bowler, P.A.; Wang, H. Effects of Landscape and Local Factors on the Diversity of Flower-Visitor Groups under an Urbanization Gradient, a Case Study in Wuhan, China. Diversity 2022, 14, 208. https://doi.org/10.3390/d14030208
He M, Ran N, Jiang H, Han Z, Dian Y, Li X, Xie D, Bowler PA, Wang H. Effects of Landscape and Local Factors on the Diversity of Flower-Visitor Groups under an Urbanization Gradient, a Case Study in Wuhan, China. Diversity. 2022; 14(3):208. https://doi.org/10.3390/d14030208
Chicago/Turabian StyleHe, Mengyu, Nan Ran, Huiqian Jiang, Zemin Han, Yuanyong Dian, Xiaoxia Li, Dong Xie, Peter A. Bowler, and Hui Wang. 2022. "Effects of Landscape and Local Factors on the Diversity of Flower-Visitor Groups under an Urbanization Gradient, a Case Study in Wuhan, China" Diversity 14, no. 3: 208. https://doi.org/10.3390/d14030208