
ITS DNA Barcoding Reveals That Halophila stipulacea Still Remains the Only Non-Indigenous Seagrass of the Mediterranean Sea

 , , , and
, , , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Morphological Identification
2.2. Sample Collection
2.3. DNA Isolation, PCR Amplification, and Sequencing
2.4. Data Analysis
3. Results
3.1. Morphological Identification
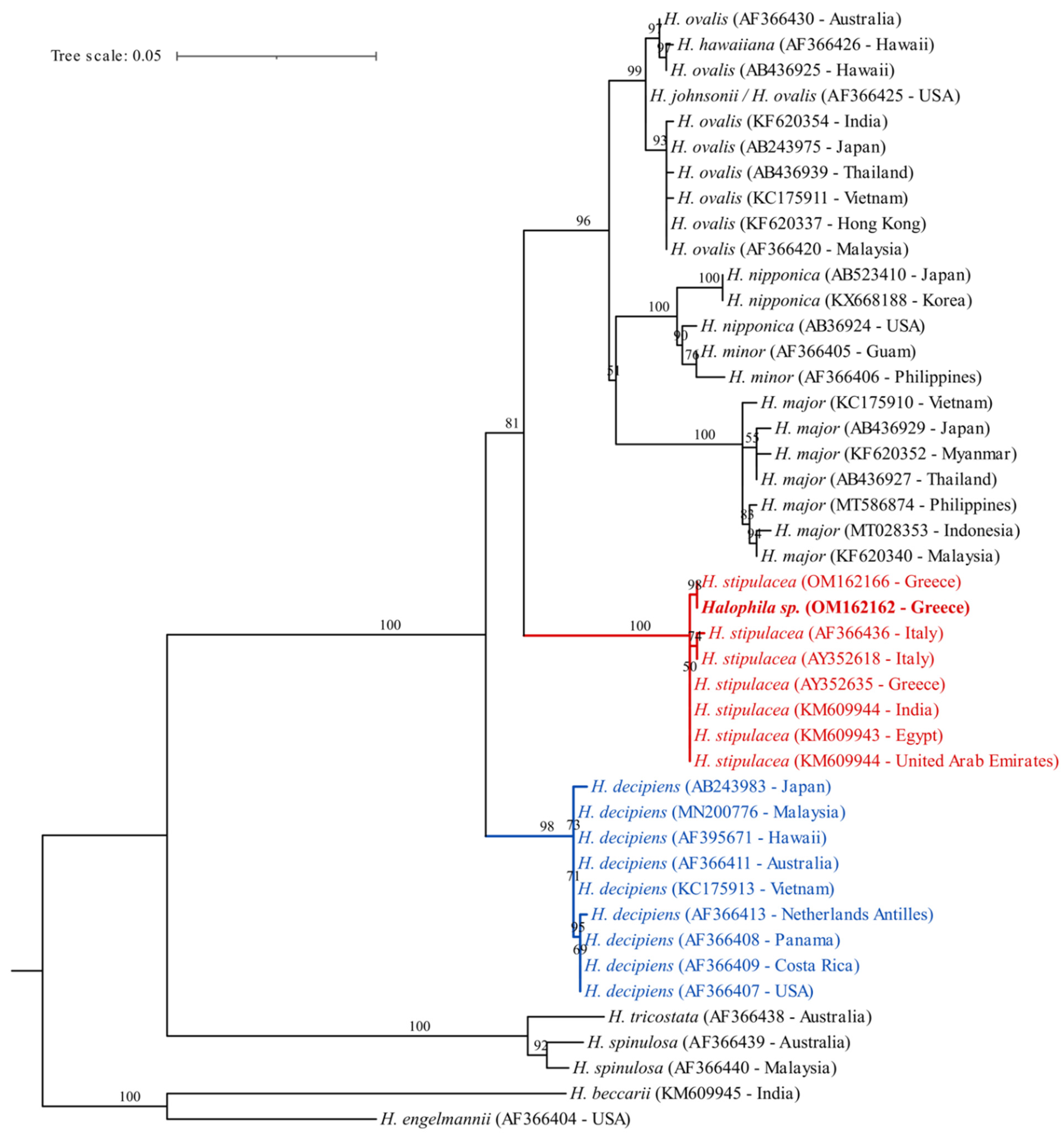
3.2. Genetic Identification
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hulme, P.E. Trade, Transport and Trouble: Managing Invasive Species Pathways in an Era of Globalization. J. Appl. Ecol. 2009, 46, 10–18. [Google Scholar] [CrossRef]
- Seebens, H.; Blackburn, T.M.; Dyer, E.E.; Genovesi, P.; Hulme, P.E.; Jeschke, J.M.; Pagad, S.; Pyšek, P.; Winter, M.; Arianoutsou, M.; et al. No Saturation in the Accumulation of Alien Species Worldwide. Nat. Commun. 2017, 8, 14435. [Google Scholar] [CrossRef]
- Pecl, G.; Araujo, M.B.; Bell, J.D.; Blanchard, J.; Bonebrake, T.C.; Pecl, G.T.; Araujo, M.B.; Bell, J. Biodiversity Redistribution under Climate Change: Impacts on Ecosystems and Human Well-Being Publication Details. Science 2017, 355, 1–9. [Google Scholar] [CrossRef]
- Gallardo, B.; Clavero, M.; Sánchez, M.I.; Vilà, M. Global Ecological Impacts of Invasive Species in Aquatic Ecosystems. Glob. Chang. Biol. 2016, 22, 151–163. [Google Scholar] [CrossRef]
- Worm, B.; Barbier, E.B.; Beaumont, N.; Duffy, E.; Folke, C.; Halpern, B.S.; Jackson, J.B.C.; Lotze, H.K.; Micheli, F.; Palumbi, S.R.; et al. Impacts of Biodiversity Loss on Ocean Ecosystem Services. Science 2006, 314, 787–790. [Google Scholar] [CrossRef] [Green Version]
- Costello, M.J.; Coll, M.; Danovaro, R.; Halpin, P.; Ojaveer, H.; Miloslavich, P. A Census of Marine Biodiversity Knowledge, Resources, and Future Challenges. PLoS ONE 2010, 5, e12110. [Google Scholar] [CrossRef] [Green Version]
- Edelist, D.; Rilov, G.; Golani, D.; Carlton, J.T.; Spanier, E. Restructuring the Sea: Profound Shifts in the World’s Most Invaded Marine Ecosystem. Divers. Distrib. 2013, 19, 69–77. [Google Scholar] [CrossRef]
- Zenetos, A.; Galanidi, M. Mediterranean Non Indigenous Species at the Start of the 2020s: Recent Changes. Mar. Biodivers. Rec. 2020, 13, 10. [Google Scholar] [CrossRef]
- Orfanidis, S.; Alvito, A.; Azzurro, E.; Badreddine, A.; Souissi, J.B.; Chamorro, M.; Crocetta, F.; Dalyan, C.; FortiČ, A.; Galanti, L.; et al. New Alien Mediterranean Biodiversity Records (March 2021). Mediterr. Mar. Sci. 2021, 22, 180–198. [Google Scholar] [CrossRef]
- Lodge, D.M. Biological Invasions: Lessons for Ecology. Trends Ecol. Evol. 1993, 8, 133–137. [Google Scholar] [CrossRef]
- Galil, B.S. A Sea under Siege-Alien Species in the Mediterranean. Biol. Invasions 2000, 2, 177–186. [Google Scholar] [CrossRef]
- Zenetos, A.; Gofas, S.; Morri, C.; Rosso, A.; Violanti, D.; García Raso, J.E.; Çinar, M.E.; Almogi-Labin, A.; Ates, A.S.; Azzurro, E.; et al. Alien Species in the Mediterranean Sea by 2012. A Contribution to the Application of European Union’s Marine Strategy Framework Directive (MSFD). Part 2. Introduction Trends and Pathways. Mediterr. Mar. Sci. 2012, 13, 328–352. [Google Scholar] [CrossRef] [Green Version]
- Molnar, J.L.; Gamboa, R.L.; Revenga, C.; Spalding, M.D. Assessing the Global Threat of Invasive Species to Marine Biodiversity. Front. Ecol. Environ. 2008, 6, 485–492. [Google Scholar] [CrossRef]
- Bianchi, C.N.; Morri, C. Global Sea Warming and “Tropicalization” of the Mediterranean Sea: Biogeographic and Ecological Aspects. Biogeogr.-J. Integr. Biogeogr. 2003, 24. [Google Scholar] [CrossRef] [Green Version]
- Coll, M.; Piroddi, C.; Steenbeek, J.; Kaschner, K.; Lasram, F.B.R.; Aguzzi, J.; Ballesteros, E.; Bianchi, C.N.; Corbera, J.; Dailianis, T.; et al. The Biodiversity of the Mediterranean Sea: Estimates, Patterns, and Threats. PLoS ONE 2010, 5, e11842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reverdin, G.; Bosse, A.; Juza, M.; Tintoré, J. Multivariate Sub-Regional Ocean Indicators in the Mediterranean Sea: From Event Detection to Climate Change Estimations. Front. Mar. Sci. 2021, 8, 233. [Google Scholar] [CrossRef]
- Raitsos, D.E.; Beaugrand, G.; Georgopoulos, D.; Zenetos, A.; Pancucci-Papadopoulou, A.M.; Theocharis, A.; Papathanassiou, E. Global Climate Change Amplifies the Entry of Tropical Species into the Eastern Mediterranean Sea. Limnol. Oceanogr. 2010, 55, 1478–1484. [Google Scholar] [CrossRef]
- Albano, P.G.; Steger, J.; Bošnjak, M.; Dunne, B.; Guifarro, Z.; Turapova, E.; Hua, Q.; Kaufman, D.S.; Rilov, G.; Zuschin, M. Native Biodiversity Collapse in the Eastern Mediterranean. Proc. R. Soc. B Biol. Sci. 2021, 288, 20202469. [Google Scholar] [CrossRef]
- Hoffman, R. Alien Benthic Algae and Seagrasses in the Mediterranean Sea and Their Connection to Global Warming. In The Mediterranean Sea; Springer: Dordrecht, The Netherlands, 2014; pp. 159–181. [Google Scholar] [CrossRef]
- Cullen-Unsworth, L.C.; Nordlund, L.M.; Paddock, J.; Baker, S.; McKenzie, L.J.; Unsworth, R.K.F. Seagrass Meadows Globally as a Coupled Social-Ecological System: Implications for Human Wellbeing. Mar. Pollut. Bull. 2014, 83, 387–397. [Google Scholar] [CrossRef] [PubMed]
- Nordlund, L.M.; Koch, E.W.; Barbier, E.B.; Creed, J.C. Seagrass Ecosystem Services and Their Variability across Genera and Geographical Regions. PLoS ONE 2016, 11, e0163091. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- den Hartog, C. Range Extension of Halophila stipulacea (Hydrocharitaceae) in the Mediterranean. Blumea 1972, 20, 154–156. [Google Scholar]
- Lipkin, Y. Halophila stipulacea, a Review of a Successful Immigration. Aquat. Bot. 1975, 1, 203–215. [Google Scholar] [CrossRef]
- Lipkin, Y. Halophila stipulacea in Cyprus and Rhodes, 1967–1970. Auqat. Bot. 1975, 1, 309–320. [Google Scholar] [CrossRef]
- den Hartog, C. The Sea-Grasses of the World; North-Holland Publishing Company: Amsterdam, The Netherlands, 1970; Volume 59. [Google Scholar]
- Fritsch, C. Ueber Die Auffindung Einer Marinen Hydrocharideeim Mittelmeer. Verh. Zool. Bot. Ges. 1895, 45, 104–106. [Google Scholar]
- Winters, G.; Beer, S.; Willette, D.A.; Viana, I.G.; Chiquillo, K.L.; Beca-Carretero, P.; Villamayor, B.; Azcárate-García, T.; Shem-Tov, R.; Mwabvu, B.; et al. The Tropical Seagrass Halophila stipulacea: Reviewing What We Know from Its Native and Invasive Habitats, alongside Identifying Knowledge Gaps. Front. Mar. Sci. 2020, 7, 1–28. [Google Scholar] [CrossRef]
- Gambi, M.C.; Barbieri, F.; Bianchi, C.N. New Record of the Alien Seagrass Halophila stipulacea (Hydrocharitaceae) in the Western Mediterranean: A Further Clue to Changing Mediterranean Sea Biogeography. Mar. Biodivers. Rec. 2009, 2, 1–7. [Google Scholar] [CrossRef]
- Biliotti, M.; Abdelahad, N. Halophila stipulacea (Forssk.) Aschers.(Hydrocharitaceae): Espèce Nouvelle Pour l’Italie. Posidonia Newsl. 1990, 3, 23–26. [Google Scholar]
- Missaoui, H.; Mahjoub, S.; Chalghaf, M. Sur La Présence de La Phanérogame Marine Halophila stipulacea (Forsskål) Dans Le Golfe de Gabes. Bull. Inst. Nat. Sci. Technol. Mer Salammbô 2003, 30, 111–114. [Google Scholar]
- Sghaier, Y.R.; Zakhama-Sraieb, R.; Benamer, I.; Charfi-Cheikhrouha, F. Occurrence of the Seagrass Halophila stipulacea (Hydrocharitaceae) in the Southern Mediterranean Sea. Bot. Mar. 2011, 54, 575–582. [Google Scholar] [CrossRef]
- Thibaut, T.; Blanfuné, A.; Boudouresque, C.F.; Holon, F.; Agel, N.; Descamps, P.; Deter, J.; Pavy, T.; Delaruelle, G.; Verlaque, M. Distribution of the Seagrass Halophila stipulacea: A Big Jump to the Northwestern Mediterranean Sea. Aquat. Bot. 2022, 176, 103465. [Google Scholar] [CrossRef]
- Willette, D.A.; Chalifour, J.; Debrot, A.O.D.; Engel, M.S.; Miller, J.; Oxenford, H.A.; Short, F.T.; Steiner, S.C.C.; Védie, F. Continued Expansion of the Trans-Atlantic Invasive Marine Angiosperm Halophila stipulacea in the Eastern Caribbean. Aquat. Bot. 2014, 112, 98–102. [Google Scholar] [CrossRef]
- Beca-Carretero, P.; Teichberg, M.; Winters, G.; Procaccini, G.; Reuter, H. Projected Rapid Habitat Expansion of Tropical Seagrass Species in the Mediterranean Sea as Climate Change Progresses. Front. Plant Sci. 2020, 11, 555376. [Google Scholar] [CrossRef]
- Telesca, L.; Belluscio, A.; Criscoli, A.; Ardizzone, G.; Apostolaki, E.T.; Fraschetti, S.; Gristina, M.; Knittweis, L.; Martin, C.S.; Pergent, G.; et al. Seagrass Meadows (Posidonia oceanica) Distribution and Trajectories of Change. Sci. Rep. 2015, 5, 12505. [Google Scholar] [CrossRef] [Green Version]
- Boudouresque, C.F.; Bernard, G.; Pergent, G.; Shili, A.; Verlaque, M. Regression of Mediterranean Seagrasses Caused by Natural Processes and Anthropogenic Disturbances and Stress: A Critical Review. Bot. Mar. 2009, 52, 395–418. [Google Scholar] [CrossRef]
- Montefalcone, M.; Morri, C.; Peirano, A.; Albertelli, G.; Bianchi, C.N. Substitution and Phase Shift within the Posidonia oceanica Seagrass Meadows of NW Mediterranean Sea. Estuar. Coast. Shelf Sci. 2007, 75, 63–71. [Google Scholar] [CrossRef]
- Montefalcone, M.; Albertelli, G.; Morri, C.; Bianchi, C.N. Patterns of Wide-Scale Substitution within Meadows of the Seagrass Posidonia oceanica in NW Mediterranean Sea: Invaders Are Stronger than Natives. Aquat. Conserv. Mar. Freshw. Ecosyst. 2010, 20, 507–515. [Google Scholar] [CrossRef]
- Gerakaris, V.; Lardi, P.L.; Issaris, Y. First Record of the Tropical Seagrass Species Halophila decipiens Ostenfeld in the Mediterranean Sea. Aquat. Bot. 2020, 160, 103151. [Google Scholar] [CrossRef]
- Short, F.; Carruthers, T.; Dennison, W.; Waycott, M. Global Seagrass Distribution and Diversity: A Bioregional Model. J. Exp. Mar. Biol. Ecol. 2007, 350, 3–20. [Google Scholar] [CrossRef]
- Green, E.P.; Short, F.T.; Frederick, T. World Atlas of Seagrasses; University of California Press: Berkeley, CA, USA, 2003. [Google Scholar]
- Gorman, D.; Turra, A.; Bergstrom, E.R.; Horta, P.A. Population Expansion of a Tropical Seagrass (Halophila decipiens) in the Southwest Atlantic (Brazil). Aquat. Bot. 2016, 132, 30–36. [Google Scholar] [CrossRef]
- Schubert, N.; Demes, K. Phenotypic Plasticity in the Marine Angiosperm Halophila decipiens (Hydrocharitaceae, Streptophyta). Mar. Ecol. Prog. Ser. 2017, 575, 81–93. [Google Scholar] [CrossRef]
- Rozaimi, M.; Arina, N.; Zainee, N.F.A.; Raynusha, C.; Hidayah, N. An Uncommon Intertidal Record of Halophila decipiens Ostenfeld in the Vicinity of Mangroves. Aquat. Bot. 2020, 160, 103168. [Google Scholar] [CrossRef]
- Thresher, R.E.; Kuris, A.M. Options for Managing Invasive Marine Species. Biol. Invasions 2004, 6, 295–300. [Google Scholar] [CrossRef]
- Mehta, S.V.; Haight, R.G.; Homans, F.R.; Polasky, S.; Venette, R.C. Optimal Detection and Control Strategies for Invasive Species Management. Ecol. Econ. 2007, 61, 237–245. [Google Scholar] [CrossRef]
- Westfall, K.M.; Therriault, T.W.; Abbott, C.L. A New Approach to Molecular Biosurveillance of Invasive Species Using DNA Metabarcoding. Glob. Chang. Biol. 2020, 26, 1012–1022. [Google Scholar] [CrossRef] [PubMed]
- Darling, J.A.; Blum, M.J. DNA-Based Methods for Monitoring Invasive Species: A Review and Prospectus. Biol. Invasions 2007, 9, 751–765. [Google Scholar] [CrossRef]
- Dayrat, B. Towards Integrative Taxonomy. Biol. J. Linn. Soc. 2005, 85, 407–417. [Google Scholar] [CrossRef]
- Hebert, P.D.N.; Cywinska, A.; Ball, S.L.; DeWaard, J.R. Biological Identifications through DNA Barcodes. Proc. R. Soc. B Biol. Sci. 2003, 270, 313–321. [Google Scholar] [CrossRef] [Green Version]
- Andújar, C.; Arribas, P.; Yu, D.W.; Vogler, A.P.; Emerson, B.C. Why the COI Barcode Should Be the Community DNA Metabarcode for the Metazoa. Mol. Ecol. 2018, 27, 3968–3975. [Google Scholar] [CrossRef]
- Kress, W.J.; Wurdack, K.J.; Zimmer, E.A.; Weigt, L.A.; Janzen, D.H. Use of DNA Barcodes to Identify Flowering Plants. Proc. Natl. Acad. Sci. USA 2005, 102, 8369–8374. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Yang, Y.; Henry, R.J.; Rossetto, M.; Wang, Y.; Chen, S. Plant DNA Barcoding: From Gene to Genome. Biol. Rev. Camb. Philos. Soc. 2015, 90, 157–166. [Google Scholar] [CrossRef]
- Chase, M.W.; Salamin, N.; Wilkinson, M.; Dunwell, J.M.; Kesanakurthi, R.P.; Haidar, N.; Savolainen, V. Land Plants and DNA Barcodes: Short-Term and Long-Term Goals. Philos. Trans. R. Soc. B Biol. Sci. 2005, 360, 1889. [Google Scholar] [CrossRef] [Green Version]
- Trivedi, S.; Aloufi, A.A.; Ansari, A.A.; Ghosh, S.K. Role of DNA Barcoding in Marine Biodiversity Assessment and Conservation: An Update. Saudi J. Biol. Sci. 2016, 23, 161–171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, X.-V.; Höfler, S.; Glasenapp, Y.; Thangaradjou, T.; Lucas, C.; Papenbrock, J. New insights into DNA barcoding of seagrasses. Syst. Biodivers. 2015, 13, 496–508. [Google Scholar] [CrossRef]
- Uchimura, M.; Jean Faye, E.; Shimada, S.; Inoue, T.; Nakamura, Y. A Reassessment of Halophila Species (Hydrocharitaceae) Diversity with Special Reference to Japanese Representatives. Bot. Mar. 2008, 51, 258–268. [Google Scholar] [CrossRef]
- Hollingsworth, P.M.; Forrest, L.L.; Spouge, J.L.; Hajibabaei, M.; Ratnasingham, S.; van der Bank, M.; Chase, M.W.; Cowan, R.S.; Erickson, D.L.; Fazekas, A.J.; et al. A DNA Barcode for Land Plants. Proc. Natl. Acad. Sci. USA 2009, 106, 12794–12797. [Google Scholar] [CrossRef] [Green Version]
- Li, D.Z.; Gao, L.M.; Li, H.T.; Wang, H.; Ge, X.J.; Liu, J.Q.; Chen, Z.D.; Zhou, S.L.; Chen, S.L.; Yang, J.B.; et al. Comparative Analysis of a Large Dataset Indicates That Internal Transcribed Spacer (ITS) Should Be Incorporated into the Core Barcode for Seed Plants. Proc. Natl. Acad. Sci. USA 2011, 108, 19641–19646. [Google Scholar] [CrossRef]
- Waycott, M.; Freshwater, W.D.; York, R.A.; Calladine, A.; Kenworthy, J.W. Evolutionary Trends in the Seagrass Genus Halophila (Thouars): Insights from Molecular Phylogeny. Bull. Mar. Sci. 2002, 71, 1299–1308. [Google Scholar]
- Lucas, C.; Thangaradjou, T.; Papenbrock, J. Development of a DNA Barcoding System for Seagrasses: Successful but Not Simple. PLoS ONE 2012, 7, e29987. [Google Scholar] [CrossRef] [Green Version]
- Kuo, J. Taxonomy of the Genus Halophila Thouars (Hydocharitaceae): A Review. Plants 2020, 9, 1732. [Google Scholar] [CrossRef] [PubMed]
- Kuo, J.; den Hartog, C. “Seagrass Taxonomy and Identification Key” Global Seagrass Research Methods. In Global Seagrass Research Methods; Short, F.T., Coler, R.G., Eds.; Elsevier Science B.V: Amsterdam, The Netherlands, 2001; pp. 31–58. [Google Scholar]
- Kim, Y.K.; Kim, S.H.; Yi, J.M.; Kang, C.K.; Short, F.; Lee, K.S. Genetic Identification and Evolutionary Trends of the Seagrass Halophila nipponica in Temperate Coastal Waters of Korea. PLoS ONE 2017, 12, e0177772. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, X.-V.; Holzmeyer, L.; Papenbrock, J. New Record of the Seagrass Species Halophila Major (Zoll.) Miquel in Vietnam: Evidence from Leaf Morphology and ITS Analysis. Bot. Mar. 2013, 56, 313–321. [Google Scholar] [CrossRef] [Green Version]
- Phillips, R.C.; Mehez, E.G. Seagrasses, 34th ed.; Smithsonian Institution Scholarly Press: Washington, DC, USA, 1988. [Google Scholar]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the Sensitivity of Progressive Multiple Sequence Alignment through Sequence Weighting, Position-Specific Gap Penalties and Weight Matrix Choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. JModelTest 2: More Models, New Heuristics and Parallel Computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [Green Version]
- Trifinopoulos, J.; Nguyen, L.T.; von Haeseler, A.; Minh, B.Q. W-IQ-TREE: A Fast Online Phylogenetic Tool for Maximum Likelihood Analysis. Nucleic Acids Res. 2016, 44, W232–W235. [Google Scholar] [CrossRef] [Green Version]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; von Haeseler, A.; Lanfear, R.; Teeling, E. IQ-TREE 2: New Models and Efficient Methods for Phylogenetic Inference in the Genomic Era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A Fast and Effective Stochastic Algorithm for Estimating Maximum-Likelihood Phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive Tree of Life (ITOL) v5: An Online Tool for Phylogenetic Tree Display and Annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef] [PubMed]
- Puillandre, N.; Lambert, A.; Brouillet, S.; Achaz, G. ABGD, Automatic Barcode Gap Discovery for Primary Species Delimitation. Mol. Ecol. 2012, 21, 1864–1877. [Google Scholar] [CrossRef] [PubMed]
- Ruggiero, M.V.; Zoologica, S.; Dohrn, A.; Procaccini, G. The rDNA ITS Region in the Lessepsian Marine Angiosperm Halophila stipulacea (Forssk.) Aschers. (Hydrocharitaceae): Intragenomic Variability and Putat Climate Change Effects on Seagrass Secondary Metabolism: Ecological Implications View Project RITMARE View Project. Artic. J. Mol. Evol. 2004, 58, 115–121. [Google Scholar] [CrossRef]
- Nguyen, V.X.; Detcharoen, M.; Tuntiprapas, P.; Soe-Htun, U.; Sidik, J.B.; Harah, M.Z.; Prathep, A.; Papenbrock, J. Genetic Species Identification and Population Structure of Halophila (Hydrocharitaceae) from the Western Pacific to the Eastern Indian Ocean. BMC Evol. Biol. 2014, 14, 1–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolátková, V.; Čepička, I.; Hoffman, R.; Vohník, M. Marinomyxa Gen. Nov. Accommodates Gall-Forming Parasites of the Tropical to Subtropical Seagrass Genus Halophila and Constitutes a Novel Deep-Branching Lineage Within Phytomyxea (Rhizaria: Endomyxa). Microb. Ecol. 2021, 81, 673–686. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.Y.V.; Kumara, T.P.; Hsu, C.H. Genetic Identification and Hybridization in the Seagrass Genus Halophila (Hydrocharitaceae) in Sri Lankan Waters. PeerJ 2020, 8, e10027. [Google Scholar] [CrossRef]
- Waycott, M.; van Dijk, K.; Calladine, A.; Bricker, E.; Biffin, E. Genomics-Based Phylogenetic and Population Genetic Analysis of Global Samples Confirms Halophila johnsonii Eiseman as Halophila ovalis (R.Br.) Hook.f. Front. Mar. Sci. 2021, 8. [Google Scholar] [CrossRef]
- Singh, S.; Southgate, P.C.; Lal, M.M. Morphological Plasticity in a Fijian Seagrass: Halophila ovalis Subsp. Bullosa. Reg. Stud. Mar. Sci. 2019, 32, 100809. [Google Scholar] [CrossRef]
- McMahon, K.M.; Waycott, M. New Record for Halophila decipiens Ostenfeld in Kenya Based on Morphological and Molecular Evidence. Aquat. Bot. 2009, 91, 318–320. [Google Scholar] [CrossRef]
- Hubert, N.; Hanner, R. DNA Barcoding, Species Delineation and Taxonomy: A Historical Perspective. DNA Barcodes 2016, 3, 44–58. [Google Scholar] [CrossRef] [Green Version]
- Hebert, P.D.N.; Stoeckle, M.Y.; Zemlak, T.S.; Francis, C.M. Identification of Birds through DNA Barcodes. PLoS Biol. 2004, 2, e312. [Google Scholar] [CrossRef] [Green Version]
- Ekrem, T.; Willassen, E.; Stur, E. A Comprehensive DNA Sequence Library Is Essential for Identification with DNA Barcodes. Mol. Phylogenet. Evol. 2007, 43, 530–542. [Google Scholar] [CrossRef]
- Demes, K.W.; Graham, M.H.; Suskiewicz, T.S. Phenotypic plasticity reconciles incongruous molecular and morphological taxonomies: The giant kelp, macrocystis (laminariales, phaeophyceae), is a monospecific genus 1. J. Phycol. 2009, 45, 1266–1269. [Google Scholar] [CrossRef]
- Debiasse, M.B.; Hellberg, M.E. Discordance between Morphological and Molecular Species Boundaries among Caribbean Species of the Reef Sponge Callyspongia. Ecol. Evol. 2015, 5, 663–675. [Google Scholar] [CrossRef]
- Goonesekera, K.; Lee, P.L.M.; van der Poorten, G.; Ranawaka, G. A Global Comparison of DNA Sequences of Pelopidas (Lepidoptera: Hesperiidae) Reveals Discordance between Morphological and Genetic Data, and an Insular ‘Ghost’ Population. Insect Conserv. Divers. 2021, 14, 81–94. [Google Scholar] [CrossRef]
- Yurchenko, V.Y.; Lukeš, J.; Tesařová, M.; Jirků, M.; Maslov, D.A. Morphological Discordance of the New Trypanosomatid Species Phylogenetically Associated with the Genus Crithidia. Protist 2008, 159, 99–114. [Google Scholar] [CrossRef]
- Trujillo-Arias, N.; Rodríguez-Cajarville, M.J.; Sari, E.; Miyaki, C.Y.; Santos, F.R.; Witt, C.C.; Barreira, A.S.; Gómez, I.; Naoki, K.; Tubaro, P.L.; et al. Evolution between Forest Macrorefugia Is Linked to Discordance between Genetic and Morphological Variation in Neotropical Passerines. Mol. Phylogenet. Evol. 2020, 149, 106849. [Google Scholar] [CrossRef] [PubMed]
- McManus, G.B.; Katz, L.A. Molecular and Morphological Methods for Identifying Plankton: What Makes a Successful Marriage? J. Plankton Res. 2009, 31, 1119–1129. [Google Scholar] [CrossRef] [Green Version]
- McDonald, A.M.; Prado, P.; Heck, K.L.; Fourqurean, J.W.; Frankovich, T.A.; Dunton, K.H.; Cebrian, J. Seagrass Growth, Reproductive, and Morphological Plasticity across Environmental Gradients over a Large Spatial Scale. Aquat. Bot. 2016, 134, 87–96. [Google Scholar] [CrossRef]
- Pazzaglia, J.; Reusch, T.B.H.; Terlizzi, A.; Marín-Guirao, L.; Procaccini, G. Phenotypic Plasticity under Rapid Global Changes: The Intrinsic Force for Future Seagrasses Survival. Evol. Appl. 2021, 14, 1181–1201. [Google Scholar] [CrossRef] [PubMed]
- Pyšek, P.; Richardson, D.M. 7 Traits Associated with Invasiveness in Alien Plants: Where Do We Stand? Biol. Invasions 2008, 193, 97–125. [Google Scholar]
- Daehler, C.C. Performance Comparisons of Co-Occurring Native and Alien Invasive Plants: Implications for Conservation and Restoration. Annu. Rev. Ecol. Evol. Syst. 2003, 34, 183–211. [Google Scholar] [CrossRef]
- Ortega, A.; Geraldi, N.R.; Díaz-Rúa, R.; Oberg, S.B.; Wesselmann, M.; Kreuse-Jensen, D.; Duarte, C.M. A DNA Mini-barcode for marine macrophytes. Mol. Ecol. Resour. 2020, 20, 920–935. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, H.M.; Yadav, N.S.; Barak, S.; Lima, F.P.; Sapir, Y.; Winters, G. Responses of Invasive and Native Populations of the Seagrass Halophila stipulacea to Simulated Climate Change. Front. Mar. Sci. 2020, 6, 812. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Nº | Species | GenBank Accession | Location | Source | Ref. |
|---|---|---|---|---|---|
| 1 | Halophila decipiens | AF395671 | Hawaii | Waycott et al., 2002 | [60] |
| 2 | Halophila decipiens | AF366411 | Australia | Waycott et al., 2002 | [60] |
| 3 | Halophila decipiens | AF366407 | USA | Waycott et al., 2002 | [60] |
| 4 | Halophila decipiens | AF366413 | Curaçao | Waycott et al., 2002 | [60] |
| 5 | Halophila decipiens | AF366409 | Costa Rica | Waycott et al., 2002 | [60] |
| 6 | Halophila decipiens | AF366408 | Panama | Waycott et al., 2002 | [60] |
| 7 | Halophila decipiens | AB243983 | Japan | Uchimura et al., 2008 | [57] |
| 8 | Halophila decipiens | KC175913 | Vietnam | Nguyen et al., 2013 | [65] |
| 9 | Halophila decipiens | MN200776 | Malaysia | Rozaimi et al., 2020 | [44] |
| 10 | Halophila sp. | OM162162 | Greece | This study | - |
| 11 | Halophila stipulacea | OM162166 | Greece | This study | - |
| 12 | Halophila stipulacea | AF366436 | Italy | Waycott et al., 2002 | [60] |
| 13 | Halophila stipulacea | AY352618 | Italy | Ruggiero et al., 2004 | [75] |
| 14 | Halophila stipulacea | AY352635 | Greece | Ruggiero et al., 2004 | [75] |
| 15 | Halophila stipulacea | KM609943 | Egypt | Nguyen et al., 2015 | [56] |
| 16 | Halophila stipulacea | KM609944 | United Arab Emirates | Nguyen et al., 2015 | [56] |
| 17 | Halophila stipulacea | KM609944 | India | Nguyen et al., 2015 | [56] |
| 18 | Halophila ovalis | KF620337 | Hong Kong | Nguyen et al., 2014 | [76] |
| 19 | Halophila ovalis | AF366430 | Australia | Waycott et al., 2002 | [60] |
| 20 | Halophila ovalis | AF366420 | Malaysia | Waycott et al., 2002 | [60] |
| 21 | Halophila ovalis | AB243975 | Japan | Uchimura et al., 2008 | [57] |
| 22 | Halophila ovalis | AB436939 | Thailand | Uchimura et al., 2008 | [57] |
| 23 | Halophila ovalis | AB436925 | Hawaii | Uchimura et al., 2008 | [57] |
| 24 | Halophila ovalis | KF620354 | India | Nguyen et al., 2014 | [76] |
| 25 | Halophila ovalis | KC175911 | Vietnam | Nguyen et al., 2013 | [65] |
| 26 | Halophila hawaiiana | AF366414 | Hawaii | Waycott et al., 2002 | [60] |
| 27 | Halophila johnsonii | AF366425 | USA | Waycott et al., 2002 | [60] |
| 28 | Halophila major | AB436929 | Japan | Uchimura et al., 2008 | [57] |
| 29 | Halophila major | AB436927 | Thailand | Uchimura et al., 2008 | [57] |
| 30 | Halophila major | KC175910 | Vietnam | Nguyen et al., 2013 | [65] |
| 31 | Halophila major | KF620340 | Malaysia | Nguyen et al., 2014 | [76] |
| 32 | Halophila major | KF620352 | Myanmar | Nguyen et al., 2014 | [76] |
| 33 | Halophila major | MT586874 | Philippines | Kolátková et al., 2021 | [77] |
| 34 | Halophila major | MT028353 | Indonesia | Kolátková et al., 2021 | [77] |
| 35 | Halophila minor | AF366406 | Philippines | Waycott et al., 2002 | [60] |
| 36 | Halophila minor | AF366405 | Guam | Waycott et al., 2002 | [60] |
| 37 | Halophila nipponica | AB36924 | USA | Uchimura et al., 2008 | [57] |
| 38 | Halophila nipponica | AB523410 | Japan | Uchimura et al., 2008 | [57] |
| 39 | Halophila nipponica | KX668188 | Korea | Kim et al., 2017 | [64] |
| 40 | Halophila spinulosa | AF366440 | Malaysia | Waycott et al., 2002 | [60] |
| 41 | Halophila spinulosa | AF366439 | Australia | Waycott et al., 2002 | [60] |
| 42 | Halophila tricostata | AF366438 | Australia | Waycott et al., 2002 | [60] |
| 43 | Halophila engelmannii | AF366404 | USA | Waycott et al., 2002 | [60] |
| 44 | Halophila becarii | KM609945 | India | Nguyen et al., 2015 | [56] |
| Phillips & Mehez (1988) [66]; Kuo & Den Hartog (2001 [63]); Kuo (2020) [62] | This Study | ||
|---|---|---|---|
| Character | Halophila decipiens | Halophila stipulacea | Halophila sp. |
| Rhizome | Thin, fragile, fleshy, elongated, 1 mm diameter | 0.5–2 mm wide | Thin, fleshy, smooth, elongated, <1 mm diameter |
| Leaf shape | Oblong to elliptic, apex obtuse or rounded, base cuneate | Linear to oblong, elliptic, cartilaginous to membranous, apex obtuse, base cuneate or gradually decurrent-petiolate | Oblong to elliptic, base cuneate, apex obtuse |
| Leaf dimensions | 10–25 mm long, 2.5–6.5 mm wide | Up to 60 mm long, 10 mm wide | 7–20 mm long, 2–4 mm wide |
| Cross-veins | 5–9 pairs ascending, unbranched | 10–40 pairs, branched, ascending at 45–60 degrees | 6–9 pairs ascending, unbranched |
| Leaf margin | finely serrulate | Finely serrulate | Finely serrulate |
| Leaf surfaces | Membranous, hairy on both sides or only on the ventral side, sometimes glabrous | Glabrous, or with minute hairs; not papillous; occasionally bullate | Both surfaces covered in minute unicellular hairs |
| Petioles | Not sheathing, shorter than the blades, 3–15 mm long | Sheathing lopsidedly at the base, shorter than the blades, 5–15 mm long | Shorter than the blades, 1–26 mm long |
| Scales | Transparent, usually hairy outside | Large, elliptic, or obovate transparent scales; 12–17 mm long; 6–10 mm wide; folded at the rhizome nodes covering (sheathing) petioles | Short, obovate, transparent, not sheathing the petioles lopsidedly |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
García-Escudero, C.A.; Tsigenopoulos, C.S.; Gerakaris, V.; Tsakogiannis, A.; Apostolaki, E.T. ITS DNA Barcoding Reveals That Halophila stipulacea Still Remains the Only Non-Indigenous Seagrass of the Mediterranean Sea. Diversity 2022, 14, 76. https://doi.org/10.3390/d14020076
García-Escudero CA, Tsigenopoulos CS, Gerakaris V, Tsakogiannis A, Apostolaki ET. ITS DNA Barcoding Reveals That Halophila stipulacea Still Remains the Only Non-Indigenous Seagrass of the Mediterranean Sea. Diversity. 2022; 14(2):76. https://doi.org/10.3390/d14020076
Chicago/Turabian StyleGarcía-Escudero, Catalina A., Costas S. Tsigenopoulos, Vasilis Gerakaris, Alexandros Tsakogiannis, and Eugenia T. Apostolaki. 2022. "ITS DNA Barcoding Reveals That Halophila stipulacea Still Remains the Only Non-Indigenous Seagrass of the Mediterranean Sea" Diversity 14, no. 2: 76. https://doi.org/10.3390/d14020076