
Characterization of Two New Apodemus Mitogenomes (Rodentia: Muridae) and Mitochondrial Phylogeny of Muridae


1
School of Resources and Environmental Engineering, Anhui University, Hefei 230601, China
2
Anhui Province Key Laboratory of Wetland Ecosystem Protection and Restoration, Anhui University, Hefei 230601, China
*
Author to whom correspondence should be addressed.
Diversity 2022, 14(12), 1089; https://doi.org/10.3390/d14121089
Submission received: 16 October 2022
/
Revised: 26 November 2022
/
Accepted: 6 December 2022
/
Published: 9 December 2022
(This article belongs to the Collection Feature Papers in Phylogeny and Evolution)
Abstract
:Apodemus is the most common small rodent species in the Palearctic realm and an ideal species for biogeographical research and understanding environmental changes. Elucidating phylogenetic relationships will help us better understand species adaptation and genetic evolution. Due to its stable structure, maternal inheritance, and rapid evolution, the mitogenome has become a hot spot for taxonomic and evolutionary studies. In this research, we determined the mitochondrial genome of Apodemus agrarius ningpoensis and Apodemus draco draco and studied the phylogeny of Muridae using ML and BI trees based on all known complete mitogenomes. The mitochondrial genome of Apodemus agrarius ningpoensis was 16,262 bp, whereas that of Apodemus draco draco was 16,222 bp, and both encoded 13 protein-coding genes, 2 ribosomal RNA genes, and 22 transfer RNA genes. Analysis of base composition showed a clear A-T preference. All tRNAs except tRNASer and tRNALys formed a typical trilobal structure. All protein-coding genes contained T- and TAA as stop codons. Phylogeny analysis revealed two main branches in the Muridae family. Apodemus agrarius ningpoensis formed sister species with Apodemus chevrieri, whereas Apodemus draco draco with Apodemus latronum. Our findings provide theoretical basis for future studies focusing on the mitogenome evolution of Apodemus.
1. Introduction
With the continuous improvement of molecular biology technology in recent years, the construction of molecular phylogenetic trees has become an indispensable component of systematics and taxonomy [1]. An increasing number of species differentiation analyses rely on molecular phylogenetic analyses. Therefore, establishing reliable phylogenetic relationships is a prerequisite for accurately distinguishing between species and revealing the mechanisms of species formation.
Many researchers have attempted to determine the evolutionary relationships of species using different methods. Early traditional taxonomies that relied on morphology severely underestimated species richness [2]. Later researchers began to use protein electrophoresis, DNA hybridization and other molecular biology tools to explore the relationship between biological systems [3]. In recent decades, mitochondrial gene sequences have become a hotspot of taxonomic and evolutionary research of different animal groups [4,5,6]. Mitochondria have their own unique genetic system and thus have become a widely used molecular marker in the study of phylogeny of species because of their matrilineal inheritance and high mutation rate [7,8,9,10,11,12,13]. Mitochondria provide energy for the daily cellular activities, and their genes change under the influence of the environment in which the species lives [14,15]. Physiological changes and evolutionary pressures on species are directly reflected in their mitochondrial genome sequence [14]. However, single molecular markers of mitochondrial gene sequences often have limited phylogenetic information [6,16,17,18]. Increasing use of two, three, or more mitochondrial markers has been found in phylogenetic studies [19,20]. Researchers can gain more information from the whole mitotic genome sequences owing to their multiple structural genomic characteristics in different groups [21], which are reflected in functionally dependent differences associated with structural differences or phylogenetically distant organisms [22]. The mitochondrial genome contains information on gene rearrangements, tRNAs structure, genetic system that are lacking in single gene fragments [16], hence improving the precision of the study of evolutionary relationships among related species [6,10,23,24]. The ongoing refinement of the composition and structural characteristics of the mitochondrial genome of species help us understand the evolutionary history of species. This will better promote the study of species taxonomy [16,25].
The genus Apodemus of the Muridae family species is abundant and widely distributed in the Palearctic [26]. Members of this genus are highly adaptable in almost all habitats in the mountains, and are the most common small rodent in the field [4,27,28]. However, these rodents are the most important hosts of zoonotic diseases and parasites [12,29] and have been associated with multiple hazards to humans, such as disease transmission and crop destruction [30]. Their wide distribution and unique adaptations make members of the genus Apodemus ideal candidates for understanding the relationship between environmental change, biogeography, and rodent species evolution [27]. Approximately 7.17–9.95 Mya ago, Apodemus underwent two stages of radiation evolution to produce the agrarius and draco subgroups [28]. Moreover, detailed descriptions of different genetic lineages are lacking, which is not conducive to identifying these species based on morphology alone [12].
Apodemus agrarius ningpoensis and Apodemus draco draco are the two common Apodemus subspecies in the northern margin of the Oriental and southern margins of the Palearctic in eastern China. However, their molecular characteristics are limited.
Phylogenetic relationships of Apodemus agrarius ningpoensis and Apodemus draco draco are difficult to determine owing to the paucity of mitochondrial genomic data. Typically, phylogenetic errors in the taxonomy of Apodemus have been attributed to recessive cytochrome b pseudogenes [31]. In this study, we sequenced the mitogenomes to explore their phylogenetic position and the composition and evolution of the mitochondrial genomes of Apodemus agrarius ningpoensis and Apodemus draco draco. Their genome sequences, nucleotide composition, genome size, codon usage, gene overlap, and tRNA secondary structure were analyzed. The mitochondrial genome data generated herein provide a reference for future research focusing on the evolution of Apodemus mitochondrial genome. In addition, our study will lay a foundation for the identification and classification of species, better promote the study of rodent taxonomy.
2. Materials and Methods
2.1. Experimental Samples
Specimens of Apodemus agrarius ningpoensis and Apodemus draco draco were collected from two counties in Dabie Mountains, Yue-xi and Huo-shan, in eastern China. Samples were collected using the clip-night method, with fresh peanut kernel as bait. Combining with habitat characteristics, 100 standard iron plate clips were placed in each sample plot. The clips were placed at dusk and collected next morning. The captured rodents were first weighed and numbered, and then the conventional data such as head–body length, tail length, ear length, and hind foot length were measured, and the sex was identified. After morphological identification, BLAST analysis in NCBI (http://ncbi.nlm.gov/nuccore?db=nucleotide, accessed on 3 February 2022) using DNA barcoding sequences verified their classification. Specimens were stored in 95% ethanol and placed in a −80 °C refrigerator.
2.2. DNA Extraction and Sequencing
The SPIN easy DNA Kit for Tissue and Bacteria (with lysing matrix) (MP Biomedicals, Hefei, China) was used to obtain the required DNA from muscle tissue. High-quality DNA samples were library constructed and sequenced by the Genepioneer Biotechnology (Nanjing, China) on the Illumina HiSeq2500 platform [32].
2.3. Assembly and Annotation
The raw sequences in FASTA format were extracted from the FASTQ format obtained by sequencing using Python. The mitochondrial genome was assembled using the SPAdes 3.15.3 software [33] by referring to the KY851939.1 sequence in NCBI. Using the MITOS web server (http://mitos.bioinf.unileipzig.de/index.py, accessed on 10 May 2022) [34] to annotate the assembled sequence. Annotation results were compared with those of related species, the final annotation results were obtained after manual correction [35]. tRNA structures were predicted using the Mitos web server and tRNAscan-SE 2.0 [36,37]. The mitochondrial genome of the two Apodemus obtained in this experiment was mapped using OGDRAW (https://chlorobox.mpimp-golm.mpg.de/OGDraw, accessed on 15 May 2022).
2.4. Sequence Analysis
The base composition of the mitogenome of the two Apodemus species was calculated by MEGA X [38]. AT-skew = (A − T)/(A + T) and GC skew = (G − C)/(G + C) [39] was used to calculate the relative numbers of AT skew and GC skew, respectively. The relative synonymous codon usage (RSCU) of the PCGs of the two Apodemus species was obtained using PhyloSuite v1.2.2 [40]. After using the PhyloSuite v1.2.2 to obtain the coding sequence (CDS) and remove the stop codon from the sequence based on the codon matching pattern of the MAFFT algorithm and G-INS-i (accurate) strategy, the specific value of nonsynonymous to synonymous substitutions Ka/Ks values of the two Apodemus subspecies was calculated using the DnaSP software [41].
2.5. Phylogenetic Analysis
There are 21 genera in the Muridae family that are distributed in China [42]. To determine the phylogenetic position of Apodemus agrarius ningpoensis and Apodemus draco draco, we constructed a phylogenetic tree using the whole mitogenome sequences from 63 species of 14 genera in the Muridae family provided by NCBI and this study. Ratufa bicolor and Pteromys volans of the Sciuridae family were selected for outgroups [43] (Table 1). Mitochondrial genome sequences of 67 species were aligned using the MAFFT plug-in in PhyloSuitev1.2.2 based on the codon pattern and G-INS-i strategy for comparison. The aligned data were trimmed using trimA [41]. Bayesian inference (BI) phylogenetic analysis was based on the GTR model using MrBayes v3.2.2 [44,45]. The Markov chain Monte Carlo (MCMC) for BI analysis was started with a random tree, run 4 Markov chains, 1,000,000 generations, taking a sample every 1000 generations with a burn-in of 25% trees. The RAxMLv.7.3.0 [46] was used to construct the maximum likelihood (ML) tree, with GTRGAMMA as the default model; 1000 bootstrap repeat tests were used to verify branch reliability. The ChiPlot (https://www.chiplot.online/, accessed on 12 June 2022) was used to visualize and beautify the phylogenetic trees.
3. Results
3.1. Features of Mitogenomes
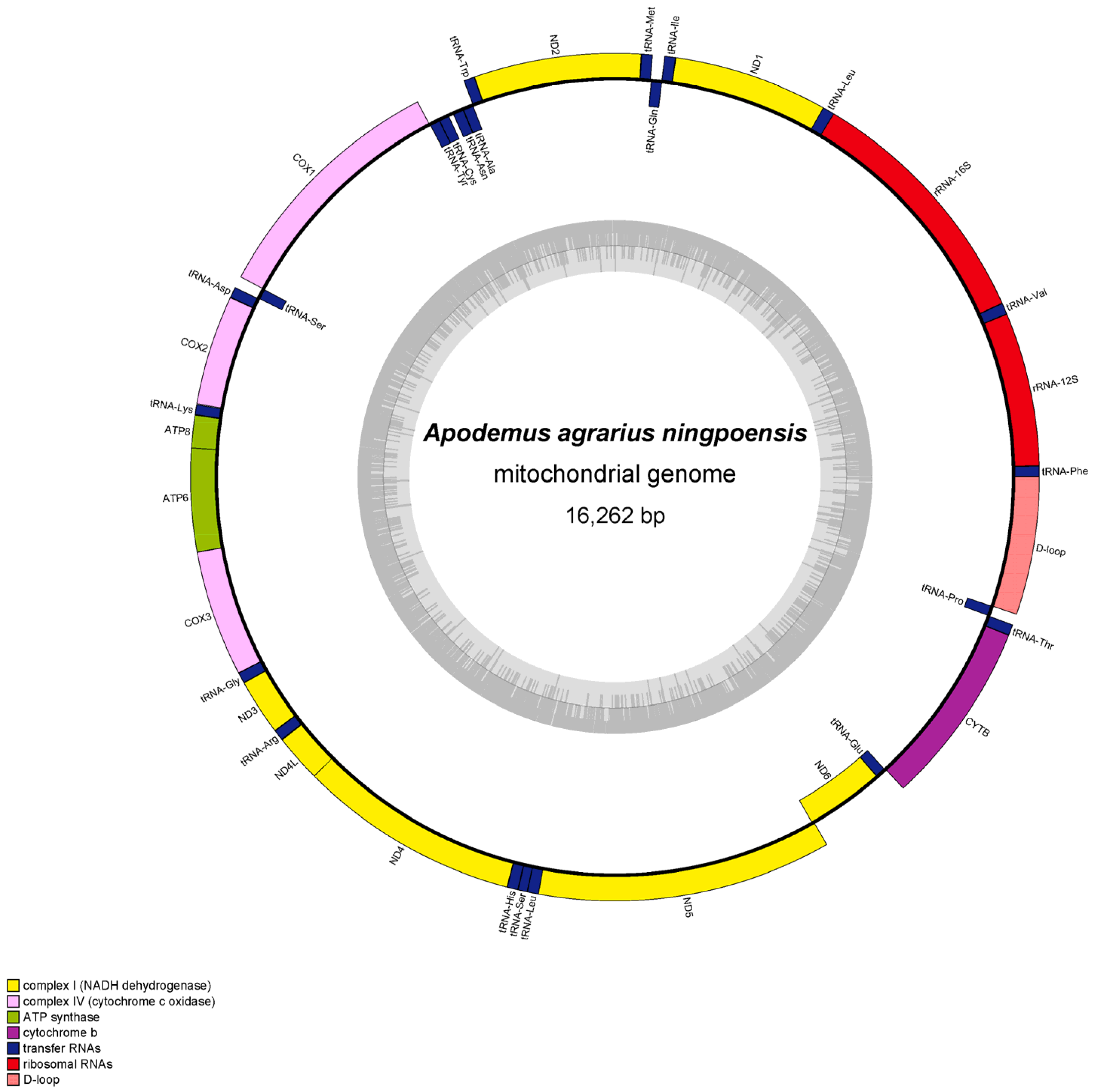
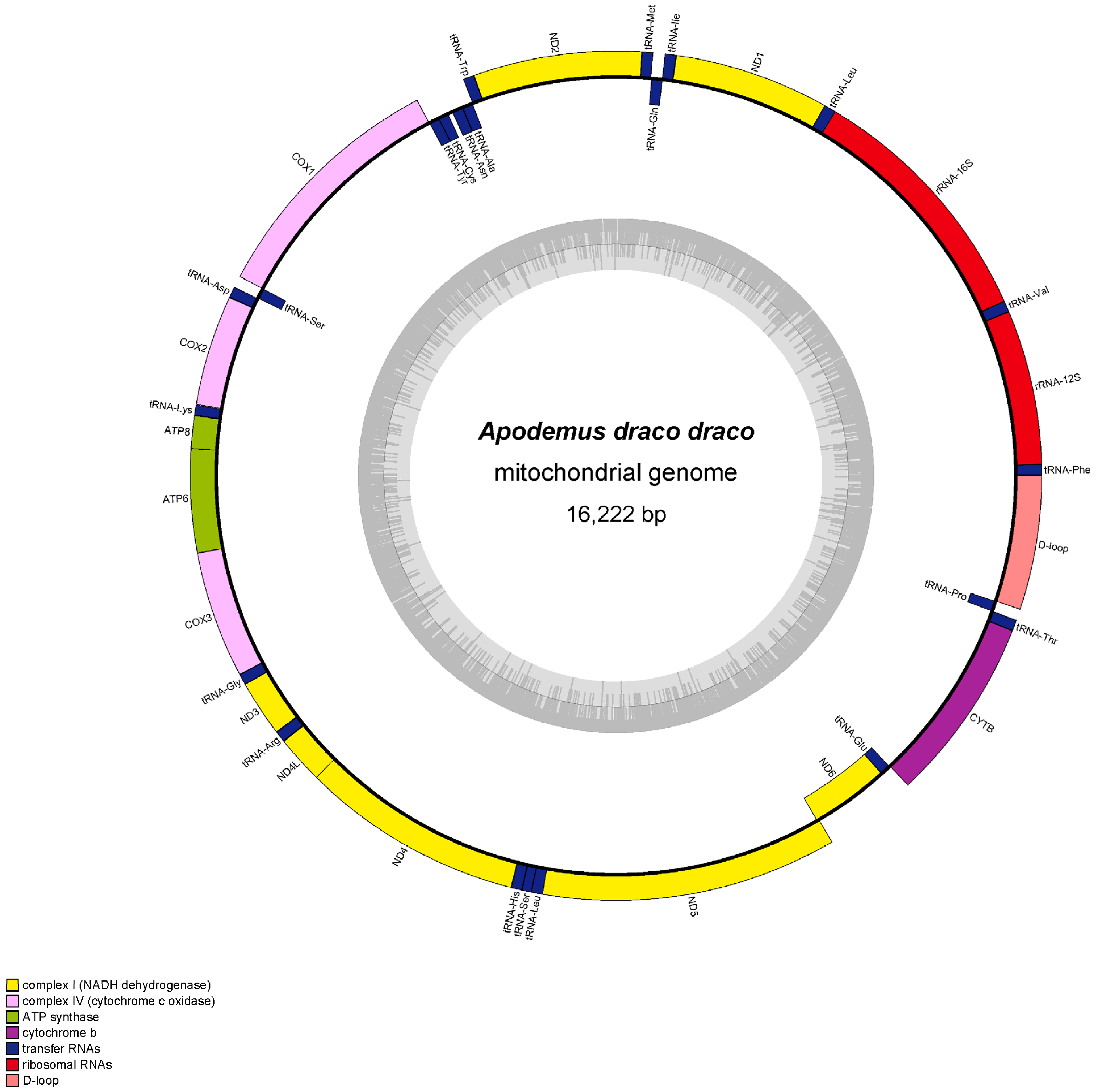
The mitochondrial genome length of Apodemus agrarius ningpoensis was 16,262 bp, and Apodemus draco draco was 16,222 bp. Both mitochondrial genomes were composed of 37 genes (13 protein-coding genes (PCGs), 22 tRNAs, and 2 rRNAs) (Figure 1 and Figure 2). Of these 37 genes, the light chain encoded 9 genes ND6, tRNAGlu, tRNAPro, tRNAGln, tRNAAla, tRNAAsn, tRNACys, tRNATyr, and tRNASer, whereas the rest were detected in the heavy chain.
The overlapping and gapping regions in the mitogenomes of Apodemus agrarius ningpoensis and Apodemus draco draco, ranged from 1 bp to 7 bp. ATP8 and ATP6 (Table 2) had the longest overlap area. The completed mitochondrial genomes had a similar composition, sequences, and localization to other vertebrates. Interestingly, both mitochondrial genome sequences were biased toward A and T. The AT content of Apodemus agrarius ningpoensis was 63.74%, and that of Apodemus draco draco was 64.12%. The AT-skew value was greater than 0, whereas the GC skew value was less than 0, indicating that the base composition of Apodemus agrarius ningpoensis and Apodemus draco draco showed a strong A-bias and C-bias (Table 3 and Table 4).
3.2. Protein-Coding Genes and Codon Usage
The full length of the protein-coding genes of Apodemus agrarius ningpoensis and Apodemus draco draco was 11,406 bp and 11,202 bp, respectively. In the Apodemus agrarius ningpoensis mitogenome, the start codon of all PCGs was ATN, ATG was the most common start codon, identified in 9 PCGs (COX1, COX2, ATP8, ATP6, COX3, ND4L, ND4, ND6, and CYTB), whereas ATA was the start codon in the remaining PCGs. We observed that the ND1, ND2, COX1, COX3, ND4, and CYTB genes had the stop codon T-, whereas in the remaining seven PCGs had TAA. Interestingly, we detected that the third codon of PCGs tended to select base A. In the case of Apodemus draco draco, ND1, ND2, ND3, and ND5 started with an ATT codon, unlike the rest PCGs that used ATG as the start codon. Similar to Apodemus agrarius ningpoensis, ND1, ND2, COX1, COX3, ND4, and CYTB had the stop codon T-, whereas the rest had TAA.
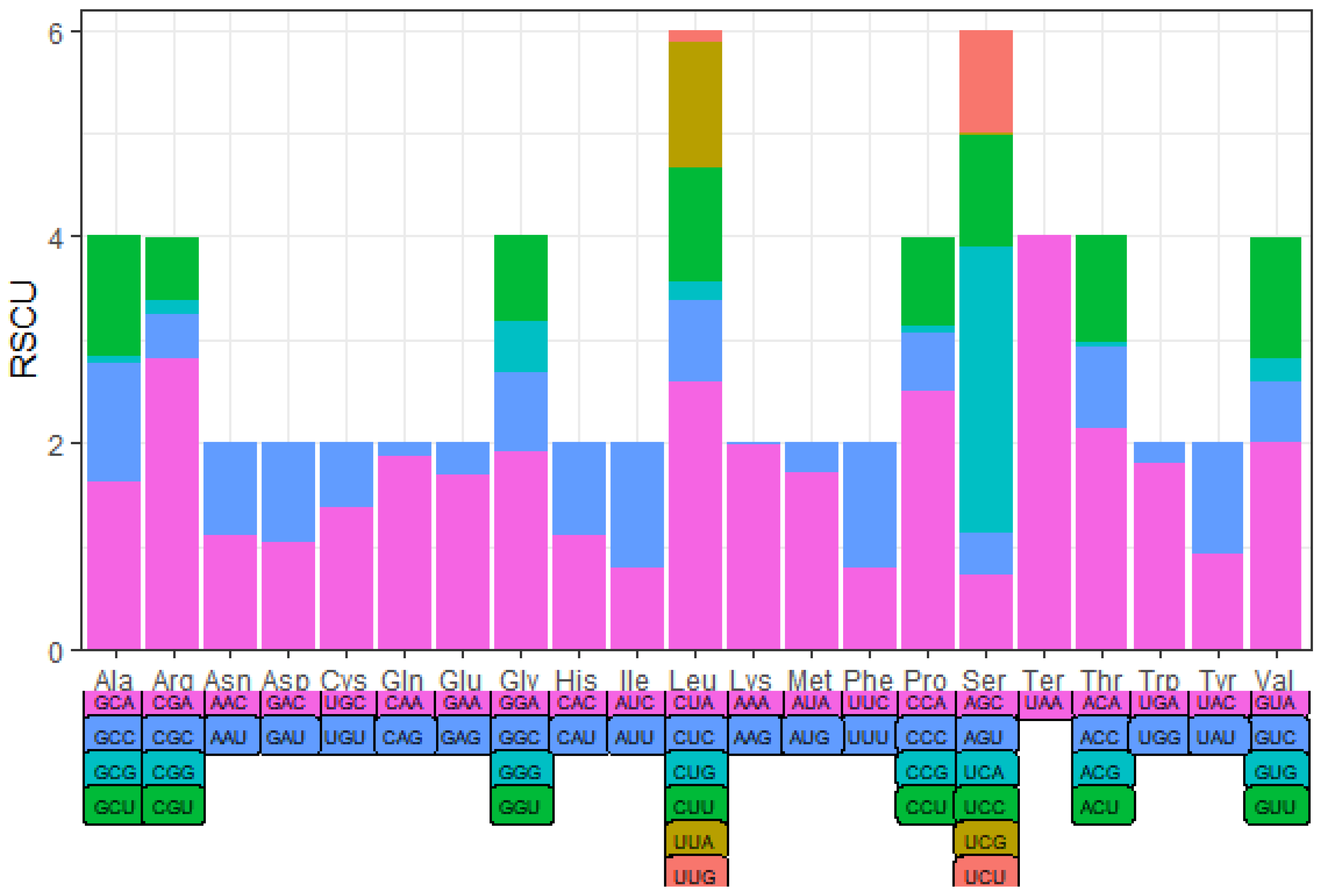
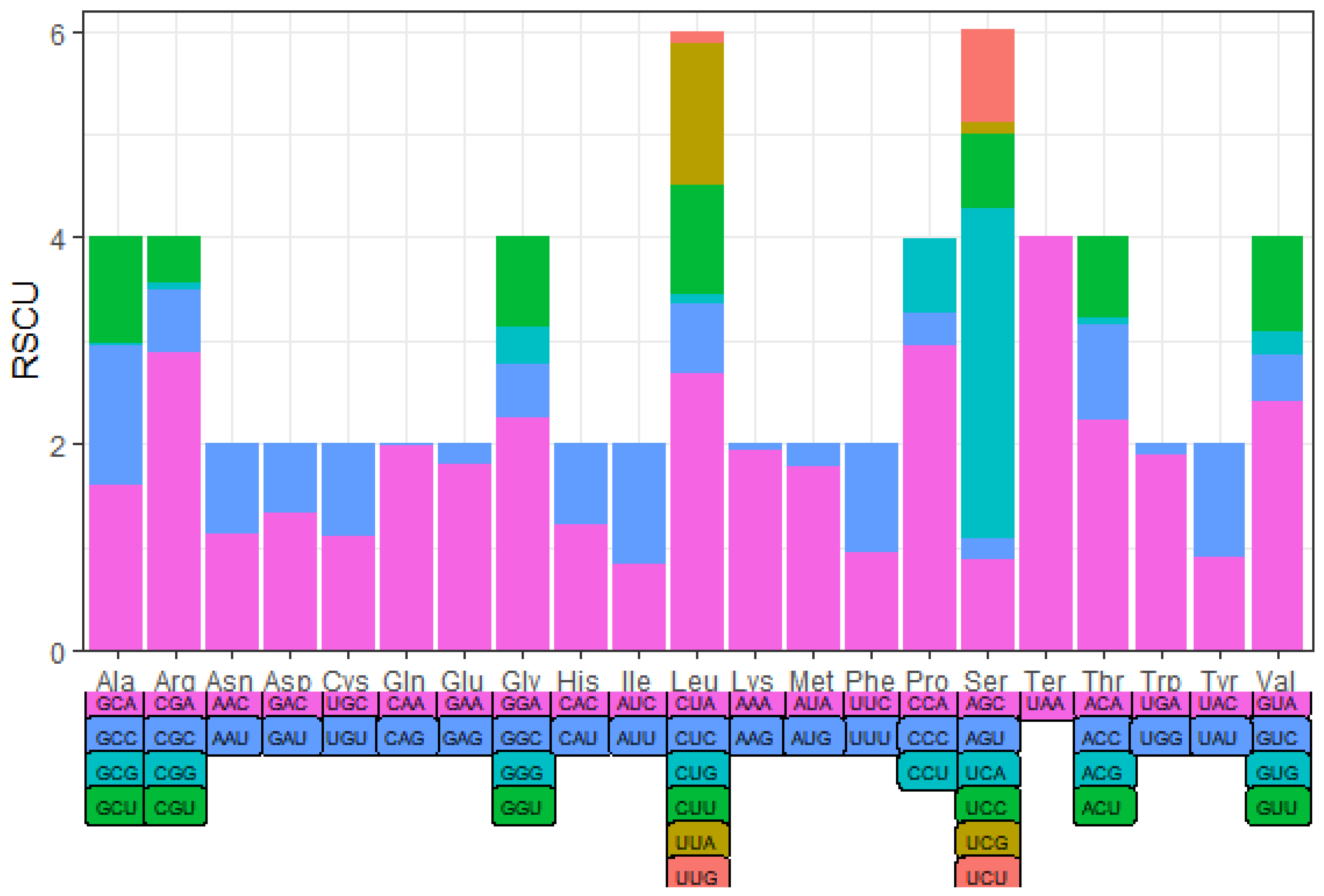
The codon usage of PCGs was evaluated based on RSCU values (Table 5 and Table 6, Figure 3 and Figure 4), CUA of Leu, CGA of Arg, and UCA of Ser were the most frequent codons among all 13 PCGs in Apodemus agrarius ningpoensis. In contrast, CGA of Arg, CCA of Pro, and UCA of Ser were the most commonly used codons in Apodemus draco draco PCGs.
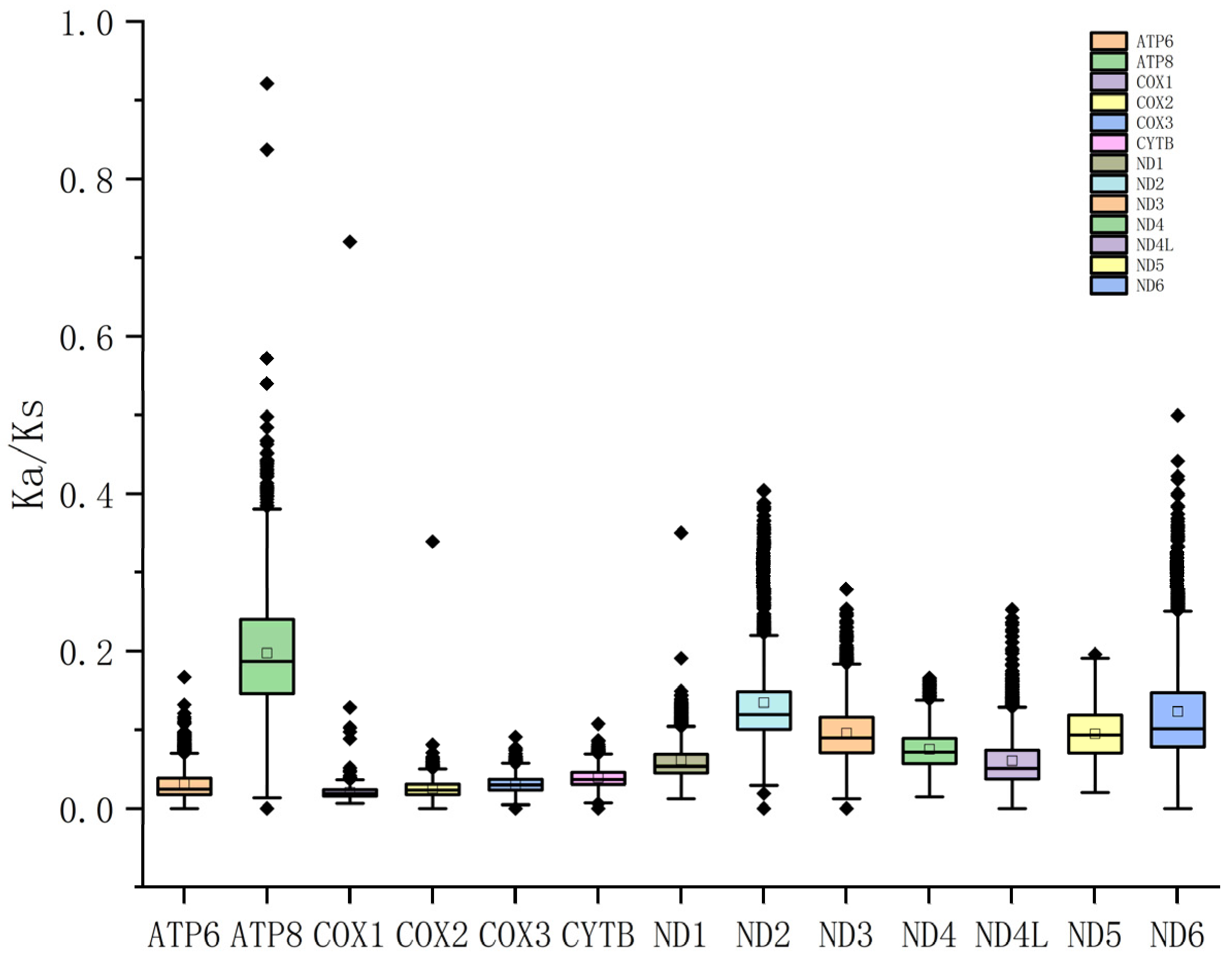
We calculated the Ka/Ks values for 13 PCGs from 65 Muridae species (Figure 5). All Ka/Ks values were smaller than 1, indicating genes subjected to purification selection. In particular, the mean and median Ka/Ks values were the highest for ATP8, suggesting that ATP8 had the highest evolution speed, the lowest for COX1, indicating the slowest evolution.
3.3. tRNAs and rRNAs
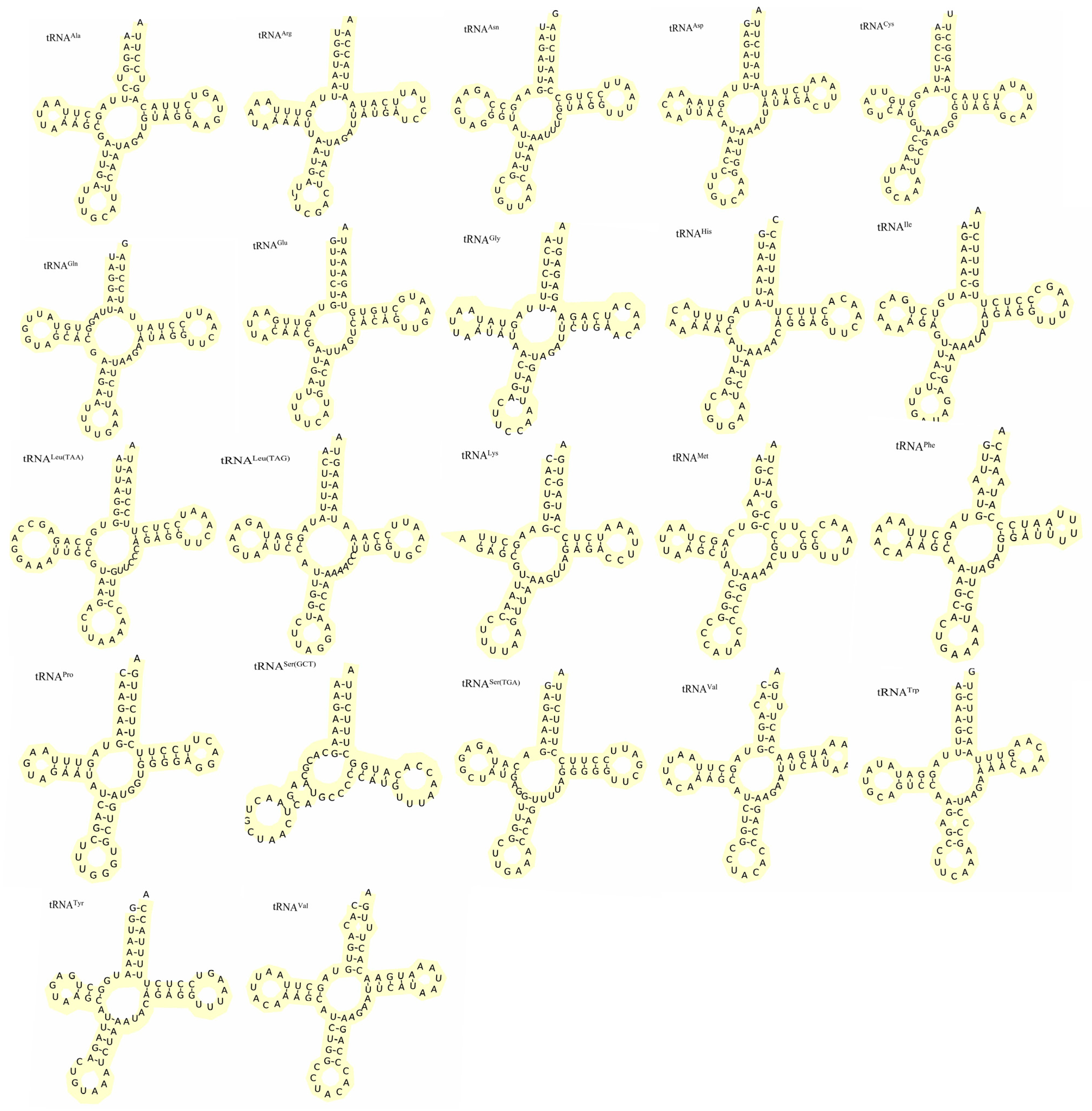
As expected, both Apodemus agrarius ningpoensis and Apodemus draco draco had 22 tRNA genes in their mitochondrial genomes, of which 9 were encoded on the negative chain, and the remaining 13 were encoded on the positive chain (Figure 6 and Figure 7). The 12S rRNA and 16S rRNA genes were located between the tRNAPhe and tRNALeu genes and segregated by the tRNAVal gene. In addition, tRNASer lacked the entire dihydrouracil (DHU) arm, tRNALys had a DHU arm lacking the D-loop to form a stable secondary structure, while the rest could form a cloverleaf structure. We further identified 36 mismatches in the Apodemus agrarius ningpoensis mitogenome, including 28 G-U, forming weak hydrogen bonds in tRNAs, as well as atypical A-A2, U-U1, A-G1, U-U2, C-U1, and A-C1. Similarly, there were 33 mismatches in the Apodemus draco draco mitogenome, including G-U26, U-U1, A-A2, A-G1, C-A2, and C-U1.
3.4. Phylogenetic Analysis
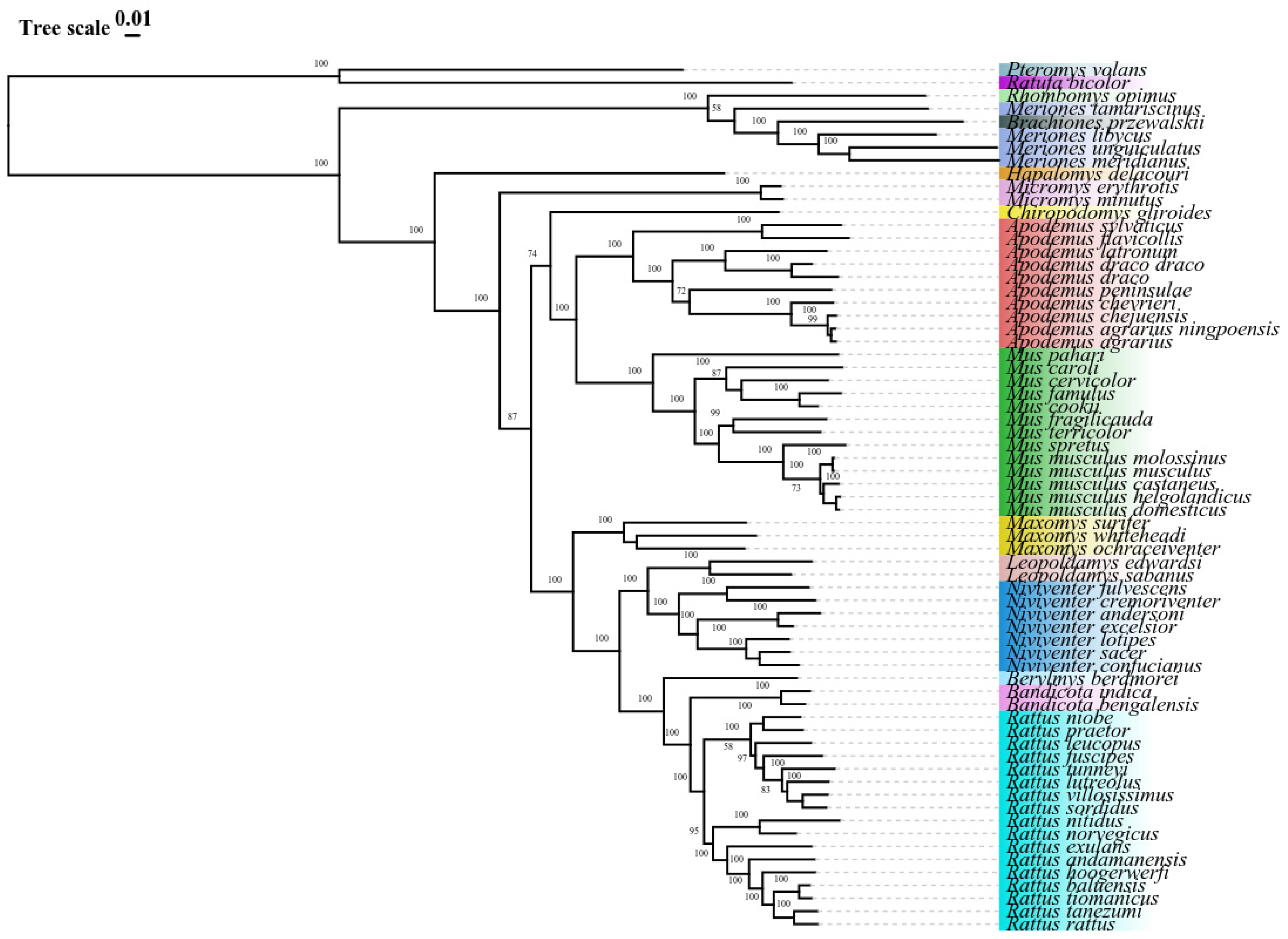
Phylogenetic trees were constructed based on ML and BI methods and yielded a consistent topology (Figure 8 and Figure 9). Most branches of the BI and ML trees received high node support. We identified two major clades in the Muridae family. One clade consisted of the genera Rhombomys, Meriones, Brachiones, and Hapalomys, whereas the others were composed of the genera Micromys, Chiropodomys, Apodemus, Mus, Maxomys, Leopoldamys, Niviventer, Berylmys, Bandicota, and Rattus. Interestingly, the genera Hapalomys, Micromys, Chiropodomys, Apodemus, Mus, Maxomys, Leopoldamys, Niviventer, Berylmys, Bandicota, and Rattus exhibited monophyly. However, the genus Meriones did not form a monophyletic group, and was first grouped with the genus Brachiones, followed by the genus Rhombomys g, and finally with the genus Meriones. The other branch of the relationship was (Apodemus + Mus) + (Maxomys + ((Leopoldamys + Niviventer) + (Berylmys + (Bandicota + Rattus))). Several members of the genus Apodemus formed an evident monophyletic group, which was divided into two small branches, among which Apodemus agrarius ningpoensis and Apodemus chevrieri were first clustered into a clade. Subsequently, it was grouped with Apodemus peninsulae, constituting a monophyletic group. The second group was composed of Apodemus flavicollis and Apodemus sylvaticus. The monophyletic lineage of the genus Apodemus was well supported by the sisters of the genus Mus; moreover, the first genus to be grouped with the genus Apodemus was the genus Mus.
4. Discussion
The length of the mitochondrial genome analyzed in this study was consistent with that of mammals, which is generally approximately 16,000 bp long [47]. The mitochondrial genome structure of Apodemus agrarius ningpoensis and Apodemus draco draco is a double-stranded closed loop, containing a noncoding control region sequence and encoding 37 genes. The distribution of mtDNA genes was not homogeneous. Most mitochondrial genes were encoded on the heavy chain, except for the ND6 gene and eight tRNA genes, which are encoded on the light chain. The apparent A-T preference in base composition was consistent with the basic characteristics of the Muridae mitochondrial genome, which is related to a base composition bias associated with the shedding of purine or pyrimidine bases from heavy chains during replication [48]. This high AT content and mutation rate of the mitochondrial genome have also been suggested to accelerate the evolutionary process in rodents [49,50]. Most animal mitochondrial genomes usually have obvious strand-specific biases, as expressed by AT and GC skews. The mitogenomes of the two species examined in this research showed positive AT, and negative GC skews, suggesting that the base composition of the whole mitogenomes of Apodemus agrarius ningpoensis and Apodemus draco draco is biased toward A and C. Synonymous codon frequency is generally biased owing to natural selection or mutational reasons, with different pressures on gene evolution. RSCU calculations showed that the protein-coding gene region of Apodemus agrarius ningpoensis and Apodemus draco draco had 3800 and 3799 codons, respectively. The codon CUA, CUU, UUA, and UCA determining Leu and Ser with RSCU values greater than 1, occupied a higher percentage in these mitogenomes, thus explaining the negative GC skew exhibited by the PCGs. The RSCU values indicate that there are significantly more bases in the third position of the codon for A and T than for G and C. This may be attributed to the AT preference for selective avoidance of termination codon formation and amino acid loss [51].
The calculation of Ka/Ks values revealed that ATP8 had the highest mean and median Ka/Ks values, implying that ATP8 had the fastest evolution among all PCGs, and is thus likely to be under relaxed selective pressure, accumulating more mutations [52]. In contrast, COX1 had the lowest mean and median Ka/Ks values, indicating it was under greater evolutionary pressure. All 13 PCGs had Ka/Ks values smaller than 1, indicating that sites underwent purification selection. It determines the development of the mitogenome [53].
Among the predicted 22 tRNA structures, we observed that tRNASer lacked the DHU arm and the DHU loop of tRNALys failed to form a typical cloverleaf structure. Most mammalian mitochondrial genomes lack DHU arms, which might be related to the compensatory mechanism between tRNA arms [22,54,55].
The presence of gene overlap and spacing in both species and gene overlap regions implied that Muridae species need to continuously improve DNA utilization from a supply-side perspective to meet evolutionary demands, which is extremely beneficial to the evolutionary process.
Both sequences had multiple G-U mismatches, which might be mainly attributed to the low free energy of the bond, which makes it stable and neutral [56]. This usually occurs in invertebrates and is rectified by post-transcriptional processing of RNA [57].
Currently, studies on the evolution and taxonomy of the Chinese Apodemus populations have concentrated on mitochondrial single gene sequences such as D-loop region and cytb. However, the mitochondrial genome can provide a better understanding of phylogenetic relationships [6,24]. The phylogenetic trees of the gene sequences of 63 rodent species in 14 genera of the Muridae family constructed using the BI and ML methods in this study were largely consistent, with bootstrap values of evolutionary branches >70% being considered indicative of strong relationships. This indicated a close phylogenetic relationship between species. The phylogenetic analysis of Muridae in this study was consistent with a system based on COB, IRBP, and GHR genes [43,58], albeit with the use of different datasets. We identified two major clades in the Muridae family. One clade consisted of the genera Rhombomys, Meriones, Brachiones, and Hapalomys, whereas the others were composed of the genera Micromys, Chiropodomys, Apodemus, Mus, Maxomys, Leopoldamys, Niviventer, Berylmys, Bandicota, and Rattus. Except for Rhombomys opimus, which originated from the same ancestor with the genera Brachiones and Meriones, the majority of Muridae species formed their own branches on a genus basis, which was consistent with previous studies [43]. Brachiones przewalskii has been reported to be nested in a diverse taxon corresponding to Rhombomys opimus and separated from Meriones [22]. In addition, Meriones tamariscinus formation predated the separation of Brachiones from Rhombomys [59], with Meriones tamariscinus being closely related to Rhombomys but more distantly related to any other Meriones species. The results of the study by Ito et al. supported our findings [60]. These morphological features might be associated with surrounding habitats. Ecological specialization allows for molecular and phenotypic evolution independent of each other [22,61]. Interestingly, the topology of the phylogenetic tree changed when other mitochondrial genomes were added to the study. The topology of the phylogenetic tree also changed when different species were selected for tree building.
Parallel analyses in the nuclear genome and mitogenome may probably influence phylogenetic studies in the future. The cytogenetic characteristics of the analyzed species are interesting [62]. Apodemus agrarius and Apodemus draco belong to the Apodemus group and Draco group, respectively, based on morphological characteristics. The karyotype of Apodemus agrarius had 3–5 biarmed metacentric autosome pairs (2n = 48, FNa = 54), whereas that of Apodemus draco had one small metacentric pair (2n = 48, FNa = 48). The karyotype of Apodemus chevrieri, a sister species of Apodemus agrarius, possesses four small metacentric pairs (2n = 48, FNa = 54), which was consistent with the standard karyotype of Apodemus agrarius. The sister species of Apodemus draco, Apodemus latronum, has a similar karyotype to Apodemus draco. However, they have different chromosome histotypes, and central heterochromatin is present in many distal center pairs. Two DNA phylogenetic species groups were distinct. Chromosomal rearrangements at the cytological level have contributed to the formation of post-mating reproductive isolation.
5. Conclusions
We reported and analyzed the mitochondrial genomes of Apodemus agrarius ningpoensis and Apodemus draco draco for the first time. The basic features of the mitogenomes of Apodemus agrarius ningpoensis and Apodemus draco draco, such as base content, gene arrangement, codon usage, and structures of tRNA, were highly conserved, also showing a relative abundance of AT content and a relative lack of GC content, which were consistent with the basic features of the Muridae mitochondrial genome. The new mitochondrial genome data contributed to shedding light on the phylogenetic relationship of the family Muridae and also revealed the phylogenetic position of Apodemus agrarius ningpoensis and Apodemus draco draco. The phylogenetic relationships among the various groups are consistent with related studies. It will provide genetic data for further evolutionary study of Apodemus.
In the future, developing primers using the mitochondrial genome sequence can help identify the most abundant loci in different rodent polymorphic types and combine the data with the construction of DNA barcode candidate markers to distinguish rodent populations. The data can be used for comparative analysis of mitochondrial genomes of different species, to understand disease information related to the source of the samples, and to provide a reference basis for the conservation and utilization of germplasm resources of the genus Apodemus. However, because of their wide distribution, representative Apodemus species exhibit particularly high levels of morphological and genetic variations. Expanding the sample size of the geographical population of Apodemus will help to better understand the genetic differentiation, dispersal, and phylogenetic relationships of Apodemus, as well as the influence of environmental mechanisms on its evolution.
Author Contributions
Conceptualization, L.Z.; data curation, D.W.; methodology, D.W.; software, D.W.; formal analysis, D.W.; visualization, D.W.; validation, D.W.; investigation, D.W., J.X., Q.X. and L.M.; funding acquisition, L.Z.; project administration, L.Z.; resource, L.Z.; supervision, L.Z.; writing—original draft preparation, D.W.; writing—review and editing, D.W. and L.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by grants for the National Science and Technology Fundamental Resources Investigation Program of China (2019FY101804).
Institutional Review Board Statement
The animal study protocol was approved by the Institutional Animal Care and Use Committee of Anhui Zoological Society (2022102001) on 20 October 2022.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Tokumasa, H. An introduction to molecular phylogenetic analysis. Rev. Agric. Sci. 2016, 4, 36–45. [Google Scholar] [CrossRef]
- Kryštufek, B.; Lužnik, M.; Buzan, E.V. Mitochondrial cytochrome b sequences resolve the taxonomy of field mice (Apodemus) in the western Balkan refugium. Acta Theriol. 2012, 57, 1–7. [Google Scholar] [CrossRef]
- Atopkin, D.M.; Bogdanov, A.S.; Chelomina, G.N. Genetic variation and differentiation in striped field mouse Apodemus agrarius inferred from RAPD-PCR analysis. Genetika 2007, 43, 804–817. [Google Scholar] [CrossRef]
- Yue, H.; Fan, Z.; Liu, S.; Liu, Y.; Song, Z.; Zhang, X. A mitogenome of the chevrier’s field mouse (Apodemus Chevrieri) and genetic variations inferred from the cytochrome b gene. DNA Cell Biol. 2012, 31, 460–469. [Google Scholar] [CrossRef] [PubMed]
- Jing, M.; Yu, H.; Wu, S.; Wang, W.; Zheng, X. Phylogenetic relationships in genus Niviventer (Rodentia: Muridae) in China inferred from complete mitochondrial cytochrome b gene. Mol. Phylogenet. Evol. 2007, 44, 521–529. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Li, F.; Zhou, J.; Lv, J.; Tan, Z.; Zhang, Y.; Ge, X. The phylogeny of the Anderson’s white-bellied rat (Niviventer andersoni) based on complete mitochondrial genomes. Ecol. Evol. 2022, 12, e8663. [Google Scholar] [CrossRef]
- Galtier, N.; Nabholz, B.; Glemin, S.; Hurst, G.D.D. Mitochondrial DNA as a marker of molecular diversity: A reappraisal. Mol. Ecol. 2009, 18, 4541–4550. [Google Scholar] [CrossRef]
- An, J.; Fan, C.; Fu, Z.; Zhang, H.; Yang, P. Analyses of the complete mitochondrial genome of Paraconiothyrium sp. and gene rearrangement diversity in the Pleosporales. Diversity 2022, 14, 601. [Google Scholar] [CrossRef]
- Wang, X.; Pei, J.; Bao, P.; Cao, M.; Guo, S.; Song, R.; Song, W.; Liang, C.; Yan, P.; Guo, X. Mitogenomic diversity and phylogeny analysis of yak (Bos grunniens). BMC Genom. 2021, 22, 325. [Google Scholar] [CrossRef]
- Sullivan, K.A.; Platt, R.N.; Bradley, R.D.; Ray, D.A. Whole mitochondrial genomes provide increased resolution and indicate paraphyly in deer mice. BMC Zool. 2017, 2, 11. [Google Scholar] [CrossRef]
- Kim, H.R.; Park, Y.C. The complete mitochondrial genome of the striped field mouse, Apodemus agrarius (Rodentia, Murinae) from Korea. Mitochondrial DNA 2012, 23, 145–147. [Google Scholar] [CrossRef] [PubMed]
- Ge, D.; Feijó, A.; Cheng, J.; Lu, L.; Liu, R.; Abramov, A.V.; Xia, L.; Wen, Z.; Zhang, W.; Shi, L.; et al. Evolutionary history of field mice (Murinae: Apodemus), with emphasis on morphological variation among species in China and description of a new species. Zool. J. Linn. Soc. Lond. 2019, 187, 518–534. [Google Scholar] [CrossRef]
- Moshtaghi, S.; Darvish, J.; Mirshamsi, O.; Mahmoudi, A. Cryptic species diversity in the genus Allactaga (Rodentia: Dipodidae) at the edge of its distribution range. Folia Zool. 2016, 65, 142–147. [Google Scholar] [CrossRef]
- Plese, B.; Kenny, N.J.; Rossi, M.E.; Cárdenas, P.; Schuster, A.; Taboada, S.; Koutsouveli, V.; Koutsouveli, V. Mitochondrial evolution in the Demospongiae (Porifera): Phylogeny, divergence time, and genome biology. Mol. Phylogenet. Evol. 2020, 155, 107011. [Google Scholar] [CrossRef] [PubMed]
- Dowling, D.; Friberg, U.; Lindell, J. Evolutionary implications of non-neutral mitochondrial genetic variation. Trends Ecol. Evol. 2008, 23, 546–554. [Google Scholar] [CrossRef]
- Ye, W.; Wang, J.; Zhao, X.; Liu, H.; Zhu, S. Mitochondrial genomes of two Lycosa spiders (Araneae, Lycosidae): Genome description and phylogenetic implications. Diversity 2022, 14, 538. [Google Scholar] [CrossRef]
- Wu, B.; Zhou, P.; He, J.; Cui, B.; Chen, F.; Sun, L. The taxonomic research of rodents with related target genes. Chin. J. Vector Biol. Control 2016, 27, 520–524. [Google Scholar] [CrossRef]
- Steppan, S.J.; Schenk, J.J. Muroid rodent phylogenetics: 900-species tree reveals increasing diversification rates. PLoS ONE 2017, 12, e183070. [Google Scholar] [CrossRef] [Green Version]
- Bellinvia, E. A phylogenetic study of the genus Apodemus by sequencing the mitochondrial DNA control region. J. Zool. Syst. Evol. Res. 2004, 42, 289–297. [Google Scholar] [CrossRef]
- Yalkovskaya, L.; Sibiryakov, P.; Borodin, A. Phylogeography of the striped field mouse (Apodemus agrarius Pallas, 1771) in light of new data from central part of Northern Eurasia. PLoS ONE 2022, 17, e276466. [Google Scholar] [CrossRef]
- Moritz, C.; Dowling, T.E.; Brown, W.M. Evolution of animal mitochondrial DNA: Relevance for population biology and systematics. Annu. Rev. Ecol. Syst. 1987, 18, 269–292. [Google Scholar] [CrossRef]
- Ding, L.; Luo, G.; Zhou, Q.; Sun, Y.; Liao, J. Comparative mitogenome analysis of gerbils and the mitogenome phylogeny of Gerbillinae (Rodentia: Muridae). Biochem. Genet. 2022, 60, 2226–2249. [Google Scholar] [CrossRef] [PubMed]
- Yue, H.; Yan, C.; Tu, F.; Yang, C.; Ma, W.; Fan, Z.; Song, Z.; Owens, J.; Liu, S.; Zhang, X. Two novel mitogenomes of Dipodidae species and phylogeny of Rodentia inferred from the complete mitogenomes. Biochem. Syst. Ecol. 2015, 60, 123–130. [Google Scholar] [CrossRef]
- İbiş, O. Whole mitochondrial genome sequence and phylogenetic relationships of Williams’s jerboa (Scarturus williamsi) from Turk. PeerJ 2020, 8, e9569. [Google Scholar] [CrossRef]
- Chen, L.; Lin, Y.; Xiao, Q.; Lin, Y.; Du, Y.; Lin, C.; Ward-Fear, G.; Hu, C.; Qu, Y.; Li, H. Characterization of the complete mitochondrial genome of the many-lined sun skink (Eutropis multifasciata) and comparison with other Scincomorpha species. Genomics 2021, 113, 2526–2536. [Google Scholar] [CrossRef]
- Oh, D.; Kim, T.; Chang, M.; Han, S.; Oh, H.; Kim, S. Mitochondrial genome sequences of the striped field mice Apodemus agrarius coreae and Apodemus agrarius chejuensis. Mitochondrial DNA 2011, 22, 165–167. [Google Scholar] [CrossRef]
- Wei, H.; Jia, Q.; Li, F.; Liu, Y.; Chen, S.; Yong, B. Sequencing and analysis of complete mitochondrial genome of Apodemus draco (Rodentia: Arvicolinae). Mitochondrial DNA Part A 2016, 27, 2740–2741. [Google Scholar] [CrossRef]
- Liu, X.; Wei, F.; Li, M.; Jiang, X.; Feng, Z.; Hu, J. Molecular phylogeny and taxonomy of wood mice (genus Apodemus Kaup, 1829) based on complete mtDNA cytochrome b sequences, with emphasis on Chinese species. Mol. Phylogenet. Evol. 2004, 33, 1–15. [Google Scholar] [CrossRef]
- Liu, S.; He, K.; Chen, S.; Jin, W.; Murphy, R.W.; Tang, M.; Liao, R.; Li, F. How many species of Apodemus and Rattus occur in China? A survey based on mitochondrial cyt b and morphological analyses. Zool. Res. 2018, 39, 309–320. [Google Scholar] [CrossRef]
- Xiong, Y.; You, F.; Chen, X.; Chen, Y.; Wen, Y.; Chen, Q. Detection and phylogenetic analysis of porcine bocaviruses carried by murine rodents and house shrews in China. Transbound. Emerg. Dis. 2018, 66, 259–267. [Google Scholar] [CrossRef]
- Dubey, S.; Michaux, J.; Brünner, H.; Hutterer, R.; Vogel, P. False phylogenies on wood mice due to cryptic cytochrome-b pseudogene. Mol. Phylogenet. Evol. 2009, 50, 633–641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [Green Version]
- Bernt, M.; Donath, A.; Jühling, F.; Externbrink, F.; Florentz, C.; Fritzsch, G.; Pütz, J.; Middendorf, M.; Stadler, P.F. MITOS: Improved de novo metazoan mitochondrial genome annotation. Mol. Phylogenet. Evol. 2013, 69, 313–319. [Google Scholar] [CrossRef] [PubMed]
- Bernt, M.; Braband, A.; Schierwater, B.; Stadler, P.F. Genetic aspects of mitochondrial genome evolution. Mol. Phylogenet. Evol. 2013, 69, 328–338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lowe, T.M.; Chan, P.P. tRNAscan-SE On-line: Integrating search and context for analysis of transfer RNA genes. Nucleic Acids Res. 2016, 44, W54–W57. [Google Scholar] [CrossRef]
- Chan, P.P.; Lowe, T.M. tRNAscan-SE: Searching for tRNA genes in genomic sequences. Methods Mol. Biol. 2019, 1962, 1–14. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Perna, N.T.; Kocher, T.D. Patterns of nucleotide composition at fourfold degenerate sites of animal mitochondrial genomes. J. Mol. Evol. 1995, 3, 353–358. [Google Scholar] [CrossRef]
- Zhang, D.; Gao, F.; Jakovlić, I.; Zou, H.; Zhang, J.; Li, W.X.; Wang, G.T. PhyloSuite: An integrated and scalable desktop platform for streamlined molecular sequence data management and evolutionary phylogenetics studies. Mol. Ecol. Resour. 2019, 20, 348–355. [Google Scholar] [CrossRef]
- Librado, P.; Rozas, J. Dnasp v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar] [CrossRef] [Green Version]
- Wei, F.; Yang, Q.; Wu, Y.; Jiang, X.; Liu, S.; Li, B.; Yang, G.; Li, M.; Zhou, J.; Li, S.; et al. Catalogue of mammals in China (2021). Acta Theriol. Sin. 2021, 41, 487–501. [Google Scholar] [CrossRef]
- Song, S.; Yong, H.; Lim, P.; Tan, J. Mitochondrial genome of Rattus tiomanicus (Rodentia: Muridae) and molecular phylogeny of Murinae. Sains Malays. 2021, 50, 953–965. [Google Scholar] [CrossRef]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [Green Version]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Stamatakis, A. Raxml version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [Green Version]
- Balakirev, A.E.; Abramov, A.V.; Rozhnov, V.V. Revision of the genus Leopoldamys (Rodentia, Muridae) as inferred from morphological and molecular data, with a special emphasis on the species composition in continental Indochina. Zootaxa 2013, 3640, 521–549. [Google Scholar] [CrossRef] [Green Version]
- Reyes, A.; Gissi, C.; Pesole, G.; Saccone, C. Asymmetrical directional mutation pressure in the mitochondrial genome of mammals. Mol. Biol. Evol. 1998, 15, 957–966. [Google Scholar] [CrossRef] [Green Version]
- Triant, D.A.; DeWoody, J.A. Accelerated molecular evolution in Microtus (Rodentia) as assessed via complete mitochondrial genome sequences. Genetica 2006, 128, 95–108. [Google Scholar] [CrossRef]
- Yakovchuk, P.; Protozanova, E.; Frank-Kamenetski, M.D. Base-stacking and base-pairing contributions into thermal stability of the DNA double helix. Nucleic Acids Res. 2006, 34, 564–574. [Google Scholar] [CrossRef]
- Charneski, C.A.; Honti, F.; Bryant, J.M.; Hurst, L.D.; Feil, E.J. Atypical AT skew in firmicute genomes results from selection and not from mutation. PLoS Genet. 2011, 7, e1002283. [Google Scholar] [CrossRef] [Green Version]
- Hurst, L.D. The Ka/Ks ratio: Diagnosing the form of sequence evolution. Trends Genet. 2002, 18, 486. [Google Scholar] [CrossRef]
- Wei, H.; Li, F.; Wang, X.; Wang, Q.; Chen, G.; Zong, H.; Chen, S. The characterization of complete mitochondrial genome and phylogenetic relationship within Rattus genus (Rodentia: Muridae). Biochem. Syst. Ecol. 2017, 71, 179–186. [Google Scholar] [CrossRef]
- Jiang, X.; Gao, J.; Ni, L.; Hu, J.; Li, K.; Sun, F.; Xie, J.; Bo, X.; Gao, C.; Xiao, J.; et al. The complete mitochondrial genome of Microtus fortis calamorum (Arvicolinae, Rodentia) and its phylogenetic analysis. Gene 2012, 498, 288–295. [Google Scholar] [CrossRef]
- Wolstenholme, D.R. Animal mitochondria DNA: Structure and evolution. Int. Rev. Cytol. 1992, 141, 173–216. [Google Scholar] [CrossRef]
- Liu, H.; Chen, S.; Chen, Q.; Pu, D.; Chen, Z.; Liu, Y.; Liu, X. The first mitochondrial genomes of the family Haplodiplatyidae (Insecta: Dermaptera) reveal intraspecific variation and extensive gene rearrangement. Biology 2022, 11, 807. [Google Scholar] [CrossRef]
- Lavrov, D.V.; Brown, W.M.; Boore, J.L. A novel type of RNA editing occurs in the mitochondrial tRNAs of the centipede Lithobius forficatus. Proc. Natl. Acad. Sci. USA 2000, 97, 13738–13742. [Google Scholar] [CrossRef] [Green Version]
- Lecompte, E.; Aplin, K.; Denys, C.; Catzeflis, F.; Chades, M.; Chevret, P. Phylogeny and biogeography of African Murinae based on mitochondrial and nuclear gene sequences, with a new tribal classification of the subfamily. BMC Evol. Biol. 2008, 1, 199. [Google Scholar] [CrossRef] [Green Version]
- Alhajeri, B.H.; Hunt, O.J.; Steppan, S.J. Molecular systematics of gerbils and deomyines (Rodentia: Gerbillinae, Deomyinae) and a test of desert adaptation in the tympanic bulla. J. Zool. Syst. Evol. Res. 2015, 53, 312–330. [Google Scholar] [CrossRef]
- Ito, M.; Jiang, W.; Sato, J.J.; Zhen, Q.; Jiao, W.; Goto, K.; Sato, H.; Ishiwata, K.; Oku, Y.; Chai, J.; et al. Molecular phylogeny of the subfamily Gerbillinae (Muridae, Rodentia) with emphasis on species living in the Xinjiang-Uygur Autonomous Region of China and based on the mitochondrial cytochrome b and cytochrome c oxidase subunit II genes. Zool. Sci. 2010, 27, 269–278. [Google Scholar] [CrossRef]
- Renaud, S.; Chevret, P.; Michaux, J. Morphological vs. molecular evolution: Ecology and phylogeny both shape the mandible of rodents. Zool. Scr. 2007, 36, 525–535. [Google Scholar] [CrossRef]
- Motokawa, M.; Wu, Y.; Harada, M.; Shintaku, Y.; Jiang, X.L.; Li, Y.C. Karyotypes of field mice of the genus Apodemus (Mammalia: Rodentia) from China. Zool. Res. 2018, 39, 348–355. [Google Scholar] [CrossRef]
Figure 1.
Annotated mitogenome of Apodemus agrarius ningpoensis.

Figure 2.
Annotated mitogenome of Apodemus draco draco.

Figure 3.
RSCU values of Apodemus agrarius ningpoensis.

Figure 4.
The RSCU values of Apodemus draco draco.

Figure 5.
The Ka/Ks values for the 13 PCGs of 65 Muridae mitogenomes.

Figure 6.
Structures of tRNA of Apodemus agrarius ningpoensis.

Figure 7.
Structures of tRNA of Apodemus draco draco.

Figure 8.
Phylogenetic tree inferred from mitochondrial genome sequences using ML. Bootstrap support values are indicated on branches. Different genera are distinguished by different colors.
Figure 8.
Phylogenetic tree inferred from mitochondrial genome sequences using ML. Bootstrap support values are indicated on branches. Different genera are distinguished by different colors.

Figure 9.
Phylogenetic tree inferred from mitochondrial genome sequences using BI. Bootstrap support values are indicated on branches. Different genera are distinguished by different colors.
Figure 9.
Phylogenetic tree inferred from mitochondrial genome sequences using BI. Bootstrap support values are indicated on branches. Different genera are distinguished by different colors.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Mitogenomes used in the analyses.
| Genus | Species | Accession No. |
|---|---|---|
| Apodemus | Apodemus chevrieri | NC_017599.1 |
| Apodemus | Apodemus latronum | NC_019585.1 |
| Apodemus | Apodemus flavicollis | MN122902.1 |
| Apodemus | Apodemus peninsulae | NC_016060.1 |
| Apodemus | Apodemus draco | NC_019584.1 |
| Apodemus | Apodemus chejuensis | NC_016662.1 |
| Apodemus | Apodemus agrarius | NC_016428.1 |
| Apodemus | Apodemus sylvaticus | NC_049122.1 |
| Rattus | Rattus andamanensis | NC_046686.1 |
| Rattus | Rattus tiomanicus | NC_029888.1 |
| Rattus | Rattus hoogerwerfi | NC_049040.1 |
| Rattus | Rattus baluensis | NC_035621.1 |
| Rattus | Rattus sordidus | NC_014871.1 |
| Rattus | Rattus lutreolus | NC_014858.1 |
| Rattus | Rattus villosissimus | NC_014864.1 |
| Rattus | Rattus tunneyi | NC_014861.1 |
| Rattus | Rattus fuscipes | NC_014867.1 |
| Rattus | Rattus leucopus | NC_014855.1 |
| Rattus | Rattus niobe | NC_023347.1 |
| Rattus | Rattus rattus | NC_012374.1 |
| Rattus | Rattus praetor | NC_012461.1 |
| Rattus | Rattus norvegicus | NC_001665.2 |
| Rattus | Rattus tanezumi | NC_011638.1 |
| Rattus | Rattus exulans | NC_012389.1 |
| Rattus | Rattus nitidus | NC_040919.1 |
| Hapalomys | Hapalomys delacouri | MZ159976.1 |
| Niviventer | Niviventer cremoriventer | NC_035822.1 |
| Niviventer | Niviventer lotipes | NC_065402.1 |
| Niviventer | Niviventer sacer | MZ935252.1 |
| Niviventer | Niviventer excelsior | NC_019617.1 |
| Niviventer | Niviventer confucianus | NC_023960.1 |
| Niviventer | Niviventer fulvescens | NC_028715.1 |
| Niviventer | Niviventer andersoni | NC_060500.1 |
| Leopoldamys | Leopoldamys sabanus | NC_035819.1 |
| Leopoldamys | Leopoldamys edwardsi | NC_025670.1 |
| Mus | Mus musculus musculus | NC_010339.1 |
| Mus | Mus cervicolor | NC_025269.1 |
| Mus | Mus cookii | NC_025270.1 |
| Mus | Mus fragilicauda | NC_025287.1 |
| Mus | Mus terricolor | NC_010650.1 |
| Mus | Mus musculus helgolandicus | KP877610.1 |
| Mus | Mus musculus castaneus | NC_012387.1 |
| Mus | Mus musculus domesticus | NC_006914.1 |
| Mus | Mus musculus molossinus | NC_006915.1 |
| Mus | Mus spretus | NC_025952.1 |
| Mus | Mus famulus | NC_030342.1 |
| Mus | Mus pahari | NC_036680.1 |
| Mus | Mus caroli | NC_025268.1 |
| Berylmys | Berylmys berdmorei | NC_036730.1 |
| Brachiones | Brachiones przewalskii | KT834972.1 |
| Maxomys | Maxomys ochraceiventer | NC_056988.1 |
| Maxomys | Maxomys surifer | NC_036732.1 |
| Maxomys | Maxomys whiteheadi | NC_049119.1 |
| Bandicota | Bandicota bengalensis | NC_057104.1 |
| Bandicota | Bandicota indica | KT029807.1 |
| Micromys | Micromys erythrotis | NC_060316.1 |
| Micromys | Micromys minutus | NC_027932.1 |
| Chiropodomys | Chiropodomys gliroides | NC_049121.1 |
| Meriones | Meriones unguiculatus | KF425526.1 |
| Meriones | Meriones tamariscinus | NC_034314.1 |
| Meriones | Meriones meridianus | NC_027684.1 |
| Meriones | Meriones libycus | NC_027683.1 |
| Rhombomys | Rhombomys opimus | MK359635.1 |
| Pteromys | Pteromys volans | NC_019612.1 |
| Ratufa | Ratufa bicolor | NC_023780.1 |
Table 2.
Mitochondrial composition of Apodemus agrarius ningpoensis (left) and Apodemus draco draco (right).
Table 2.
Mitochondrial composition of Apodemus agrarius ningpoensis (left) and Apodemus draco draco (right).
| Gene | Direction | Position (bp) | Size (bp) | Start Codon | Stop Codon | Intergenic Nucleotides |
|---|---|---|---|---|---|---|
| tRNAPhe | + | 1–67\1–68 | 67\68 | 0\0 | ||
| 12S rRNA | + | 68–1022\69–1023 | 955\955 | 0\0 | ||
| tRNAVal | + | 1023–1091\1024–1091 | 69\68 | 0\0 | ||
| 16S rRNA | + | 1092–2665\1092–2660 | 1547\1569 | 0\0 | ||
| tRNALeu | + | 2664–2738\2659–2733 | 75\75 | −2\−2 | ||
| ND1 | + | 2736–3693\2731–3688 | 958\958 | ATA\ATA | T-\T- | −3\−3 |
| tRNAIle | + | 3694–3762\3689–3757 | 69\69 | 0\0 | ||
| tRNAGln | − | 3760–3830\3755–3825 | 71\71 | −3\−3 | ||
| tRNAMet | + | 3835–3903\3830–3898 | 69\69 | 4\4 | ||
| ND2 | + | 3094–4939\3899–4934 | 1036\1036 | ATA\ATA | T-\T- | 0\0 |
| tRNATrp | + | 4940–5007\4935–5002 | 68\68 | 0\0 | ||
| tRNAAla | − | 5009–5077\5004–5072 | 69\69 | 1\1 | ||
| tRNAAsn | − | 5079–5149\5074–5144 | 71\71 | 1\1 | ||
| tRNACys | − | 5181–5248\5176–5241 | 68\66 | 31\31 | ||
| tRNATyr | − | 5249–5315\5242–5308 | 67\67 | 0\0 | ||
| COX1 | + | 5317–6859\5310–6852 | 1543\1543 | ATG\ATG | T-\T- | 1\1 |
| tRNASer | − | 6859–6927\6852–6920 | 69\69 | −1\−1 | ||
| tRNAAsp | + | 6931–6998\6924–6991 | 68\68 | 3\3 | ||
| COX2 | + | 7000–7683\6993–7676 | 684\684 | ATG\ATG | TAA\TAA | 1\1 |
| tRNALys | + | 7687–7750\7680–7743 | 64\64 | 3\3 | ||
| ATP8 | + | 7752–7955\7745–7948 | 204\204 | ATG\ATG | TAA\TAA | 1\1 |
| ATP6 | + | 7913–8593\7906–8586 | 681\681 | ATG\ATG | TAA\TAA | −43\−43 |
| COX3 | + | 8593–9376\8586–9369 | 784\784 | ATG\ATG | T-\T- | −1\−1 |
| tRNAGly | + | 9377–9444\9370–9436 | 68\67 | 0\0 | ||
| ND3 | + | 9445–9792\9437–9784 | 348\348 | ATA\ATT | TAA\TAA | 0\0 |
| tRNAArg | + | 9794–9861\9786–9853 | 68\68 | 1\1 | ||
| ND4L | + | 9864–10160\9856–10152 | 297\297 | ATG\ATG | TAA\TAA | 2\2 |
| ND4 | + | 10154–11531\10146–11523 | 1378\1378 | ATG\ATG | T-\T- | −7\−7 |
| tRNAHis | + | 11532–11599\11524–11590 | 68\67 | 0\0 | ||
| tRNASer | + | 11600–11658\11591–11649 | 59\59 | 0\0 | ||
| tRNALeu | + | 11658–11728\11649–11719 | 71\71 | −1\−1 | ||
| ND5 | + | 11720–13549\11711–13540 | 1830\1830 | ATA\ATA | TAA\TAA | −9\−9 |
| ND6 | − | 13536–14054\13527–14042 | 519\516 | ATG\ATG | TAA\TAA | −14\−14 |
| tRNAGlu | − | 14055–14123\14043–14111 | 69\69 | 0\0 | ||
| CYTB | + | 14127–15270\14115–15258 | 1144\1144 | ATG\ATG | T-\T- | 3\3 |
| tRNAThr | + | 15271–15339\15259–15326 | 69\68 | 0\0 | ||
| tRNAPro | − | 15340–15406\15327–15393 | 67\67 | 0\0 | ||
| D-loop | + | 15407–16262\15394–16222 | 856\829 | 0\0 |
Table 3.
Base content of the mitochondrial genome of Apodemus agrarius ningpoensis.
| Apodemus agrarius ningpoensis | Size (bp) | A% | T% | G% | C% | A + T% | G + C% | AT Skew | GC Skew |
|---|---|---|---|---|---|---|---|---|---|
| Mitogenome | 16,262 | 34.01 | 29.74 | 12.47 | 23.79 | 63.74 | 36.26 | 0.067 | −0.312 |
| PCGs | 11,406 | 32.05 | 31.45 | 12.1 | 24.4 | 63.5 | 36.5 | 0.01 | −0.337 |
| tRNAs | 1503 | 34.13 | 30.81 | 18.43 | 16.63 | 64.94 | 35.06 | 0.051 | 0.051 |
| rRNAs | 2529 | 37.92 | 26.49 | 16.77 | 18.82 | 64.41 | 35.59 | 0.177 | −0.058 |
| D-loop | 856 | 32.71 | 31.66 | 11.92 | 23.71 | 64.37 | 35.63 | 0.016 | −0.331 |
Table 4.
Base content of the mitochondrial genome of Apodemus draco draco.
| Apodemus draco draco | Size (bp) | A% | T% | G% | C% | A + T% | G + C% | AT Skew | GC Skew |
|---|---|---|---|---|---|---|---|---|---|
| Mitogenome | 16,222 | 35.15 | 28.97 | 12.08 | 23.8 | 64.12 | 35.88 | 0.096 | −0.327 |
| PCGs | 11,403 | 33.33 | 30.64 | 11.68 | 24.34 | 63.97 | 36.03 | 0.042 | −0.352 |
| tRNAs | 1498 | 34.31 | 31.51 | 18.29 | 15.89 | 65.82 | 34.18 | 0.043 | 0.07 |
| rRNAs | 2524 | 39.26 | 25.79 | 16.32 | 18.62 | 65.06 | 34.94 | 0.207 | −0.066 |
| D-loop | 829 | 31.97 | 29.67 | 13.15 | 25.21 | 61.64 | 38.36 | 0.037 | −0.314 |
Table 5.
The codon usages of Apodemus agrarius ningpoensis.
| Codon | No. | RSCU | Codon | No. | RSCU | Codon | No. | RSCU |
|---|---|---|---|---|---|---|---|---|
| UAA(*) | 7 | 4 | AAA(K) | 98 | 1.98 | CGU(R) | 10 | 0.62 |
| GCA(A) | 93 | 1.62 | AAG(K) | 1 | 0.02 | AGC(S) | 36 | 0.72 |
| GCC(A) | 66 | 1.15 | CUA(L) | 258 | 2.58 | AGU(S) | 20 | 0.4 |
| GCG(A) | 4 | 0.07 | CUC(L) | 80 | 0.8 | UCA(S) | 138 | 2.77 |
| GCU(A) | 67 | 1.17 | CUG(L) | 19 | 0.19 | UCC(S) | 54 | 1.08 |
| UGC(C) | 20 | 1.38 | CUU(L) | 109 | 1.09 | UCG(S) | 2 | 0.04 |
| UGU(C) | 9 | 0.62 | UUA(L) | 123 | 1.23 | UCU(S) | 49 | 0.98 |
| GAC(D) | 37 | 1.04 | UUG(L) | 10 | 0.1 | ACA(T) | 161 | 2.13 |
| GAU(D) | 34 | 0.96 | AUA(M) | 198 | 1.7 | ACC(T) | 60 | 0.79 |
| GAA(E) | 78 | 1.68 | AUG(M) | 35 | 0.3 | ACG(T) | 4 | 0.05 |
| GAG(E) | 15 | 0.32 | AAC(N) | 92 | 1.1 | ACU(T) | 78 | 1.03 |
| UUC(F) | 97 | 0.79 | AAU(N) | 76 | 0.9 | GUA(V) | 85 | 2.01 |
| UUU(F) | 148 | 1.21 | CCA(P) | 125 | 2.51 | GUC(V) | 25 | 0.59 |
| GGA(G) | 101 | 1.92 | CCC(P) | 28 | 0.56 | GUG(V) | 9 | 0.21 |
| GGC(G) | 40 | 0.76 | CCG(P) | 3 | 0.06 | GUU(V) | 50 | 1.18 |
| GGG(G) | 26 | 0.5 | CCU(P) | 43 | 0.86 | UGA(W) | 93 | 1.81 |
| GGU(G) | 43 | 0.82 | CAA(Q) | 77 | 1.88 | UGG(W) | 10 | 0.19 |
| CAC(H) | 53 | 1.1 | CAG(Q) | 5 | 0.12 | UAC(Y) | 60 | 0.92 |
| CAU(H) | 43 | 0.9 | CGA(R) | 45 | 2.81 | UAU(Y) | 70 | 1.08 |
| AUC(I) | 146 | 0.79 | CGC(R) | 7 | 0.44 | |||
| AUU(I) | 225 | 1.21 | CGG(R) | 2 | 0.12 |
Asterisk “*” indicates that it is a stop codon and has no corresponding amino acid.
Table 6.
The codon usages of Apodemus draco draco.
| Codon | No. | RSCU | Codon | No. | RSCU | Codon | No. | RSCU |
|---|---|---|---|---|---|---|---|---|
| UAA(*) | 7 | 4 | AUU(I) | 216 | 1.16 | CGU(R) | 7 | 0.44 |
| GCA(A) | 89 | 1.6 | AAG(K) | 3 | 0.06 | CGA(R) | 46 | 2.88 |
| GCC(A) | 76 | 1.36 | AAA(K) | 99 | 1.94 | AGU(S) | 10 | 0.2 |
| GCG(A) | 1 | 0.02 | CUC(L) | 66 | 0.67 | UCA(S) | 163 | 3.21 |
| GCU(A) | 57 | 1.02 | CUG(L) | 8 | 0.08 | UCC(S) | 37 | 0.73 |
| UGC(C) | 16 | 1.1 | CUU(L) | 104 | 1.06 | UCG(S) | 5 | 0.1 |
| UGU(C) | 13 | 0.9 | UUA(L) | 134 | 1.37 | UCU(S) | 46 | 0.9 |
| GAC(D) | 48 | 1.33 | UUG(L) | 13 | 0.13 | AGC(S) | 44 | 0.87 |
| GAU(D) | 24 | 0.67 | CUA(L) | 264 | 2.69 | ACC(T) | 71 | 0.92 |
| GAA(E) | 84 | 1.79 | AUG(M) | 28 | 0.23 | ACG(T) | 4 | 0.25 |
| GAG(E) | 10 | 0.21 | AUA(M) | 211 | 1.77 | ACU(T) | 61 | 0.79 |
| UUC(F) | 116 | 0.95 | AAU(N) | 72 | 0.88 | ACA(T) | 173 | 2.24 |
| GGA(G) | 118 | 2.26 | CCC(P) | 16 | 0.32 | GUG(V) | 10 | 0.24 |
| GGC(G) | 27 | 0.52 | CCU(P) | 36 | 0.73 | GUU(V) | 38 | 0.91 |
| GGG(G) | 19 | 0.36 | CCA(P) | 145 | 2.94 | GUA(V) | 101 | 2.42 |
| GGU(G) | 45 | 0.86 | CAG(Q) | 1 | 0.02 | UGA(W) | 98 | 1.9 |
| CAC(H) | 60 | 1.21 | CAA(Q) | 81 | 1.98 | UGG(W) | 5 | 0.1 |
| CAU(H) | 39 | 0.79 | CGC(R) | 10 | 0.62 | UAC(Y) | 59 | 0.91 |
| AUC(I) | 157 | 0.84 | CGG(R) | 1 | 0.06 | UAU(Y) | 70 | 1.09 |
Asterisk “*” indicates that it is a stop codon and has no corresponding amino acid.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Wu, D.; Zhou, L.; Xue, J.; Xia, Q.; Meng, L. Characterization of Two New Apodemus Mitogenomes (Rodentia: Muridae) and Mitochondrial Phylogeny of Muridae. Diversity 2022, 14, 1089. https://doi.org/10.3390/d14121089
AMA Style
Wu D, Zhou L, Xue J, Xia Q, Meng L. Characterization of Two New Apodemus Mitogenomes (Rodentia: Muridae) and Mitochondrial Phylogeny of Muridae. Diversity. 2022; 14(12):1089. https://doi.org/10.3390/d14121089
Chicago/Turabian StyleWu, Di, Lizhi Zhou, Jiezhong Xue, Qiliang Xia, and Lei Meng. 2022. "Characterization of Two New Apodemus Mitogenomes (Rodentia: Muridae) and Mitochondrial Phylogeny of Muridae" Diversity 14, no. 12: 1089. https://doi.org/10.3390/d14121089
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.