
Mechanisms of Seed-To-Seed Interactions between Dominant Species in the Yangtze River Estuary under Saline Condition

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Experimental Design
2.3. Data Analysis
3. Results
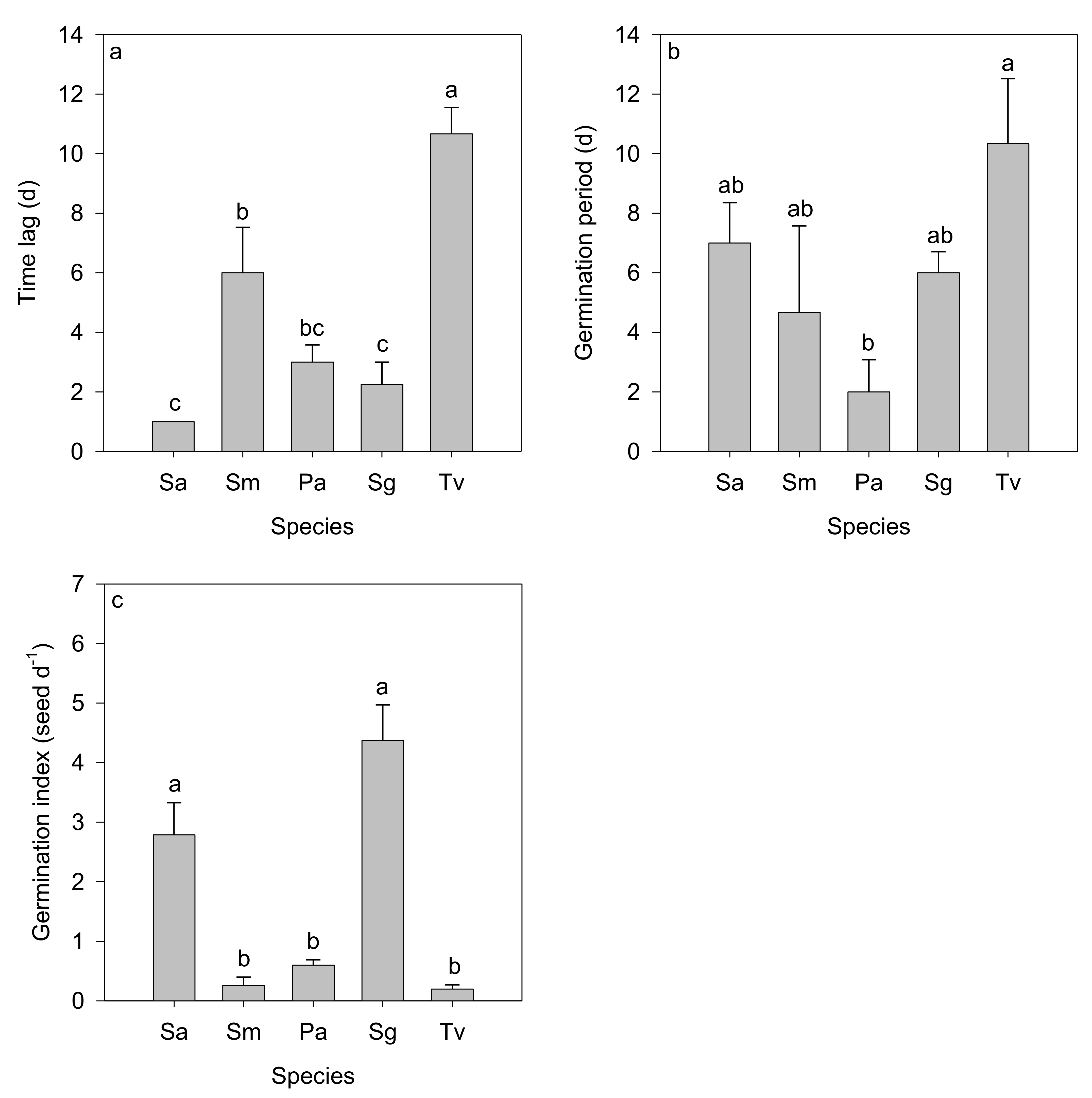
3.1. Germination Traits of Dominant Species
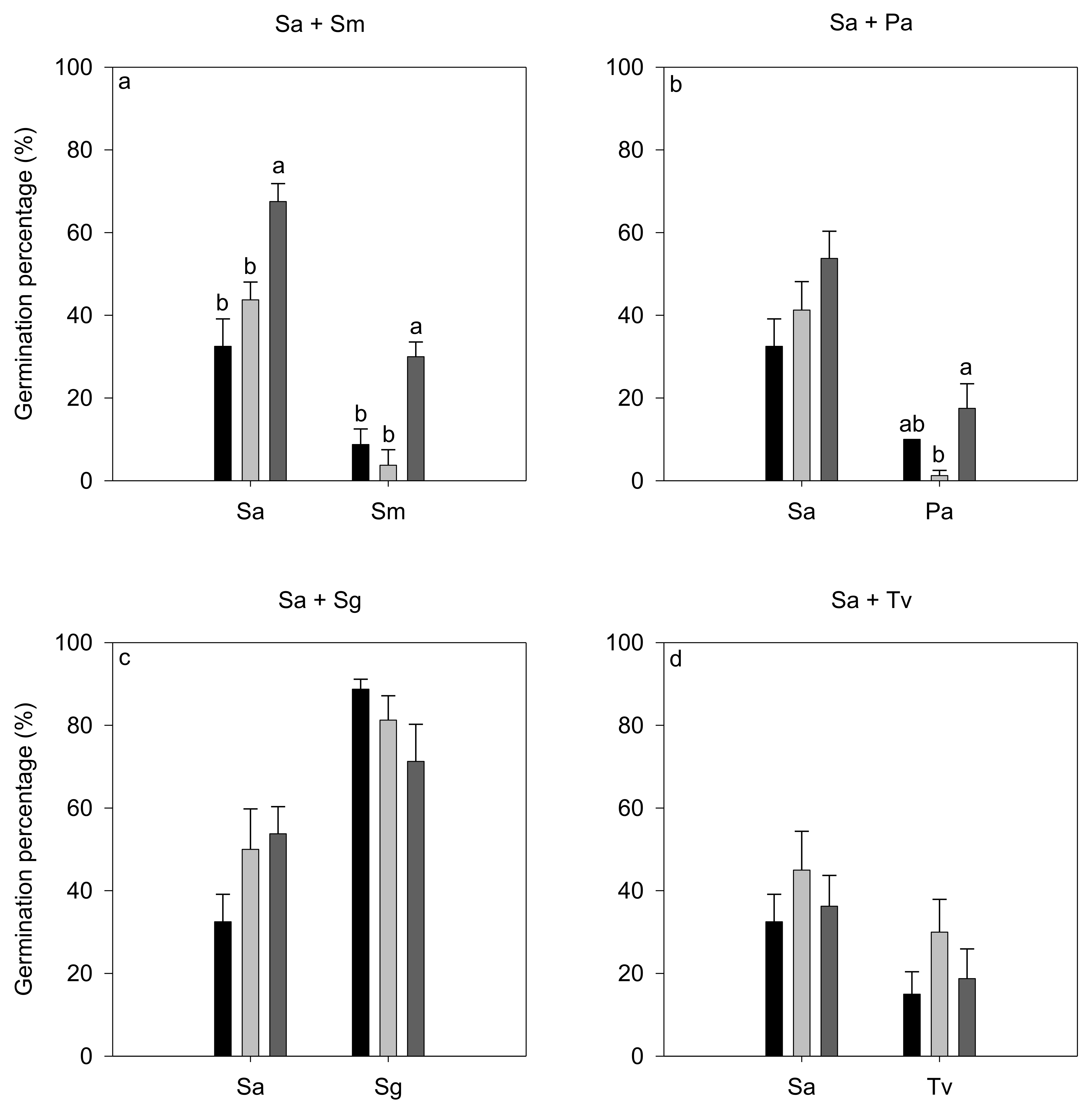
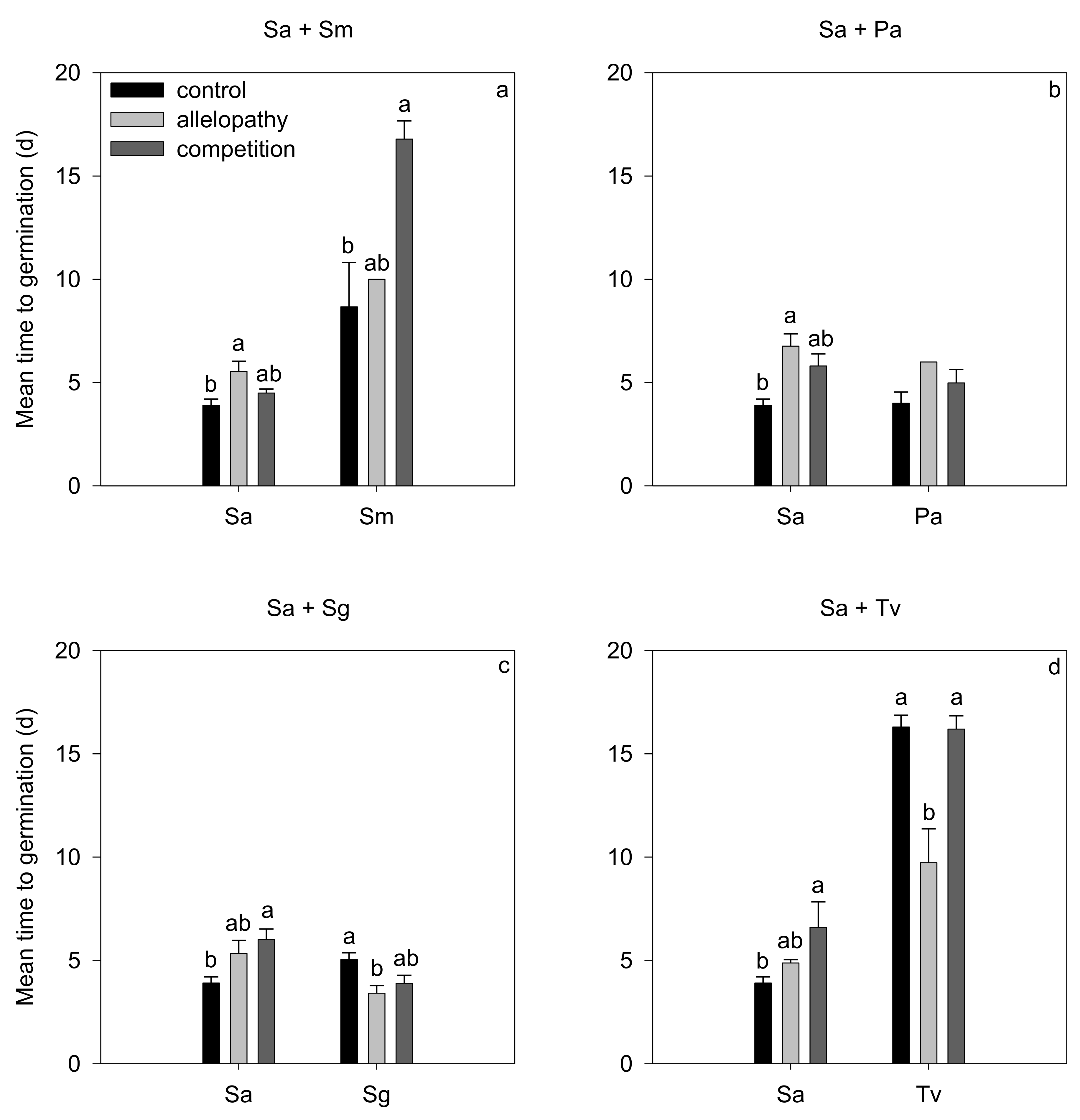
3.2. Effects of Interspecific Interaction on Germination
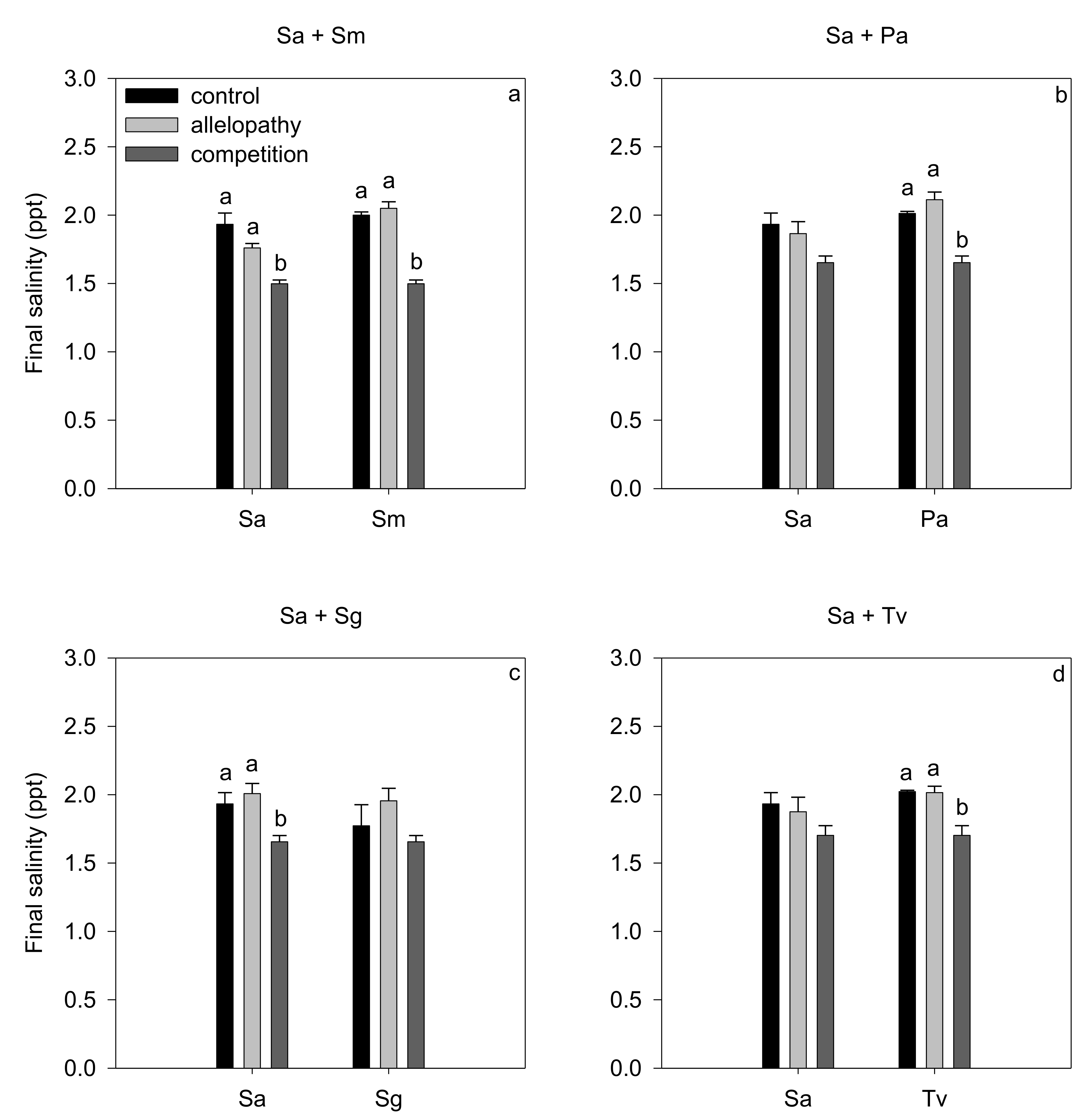
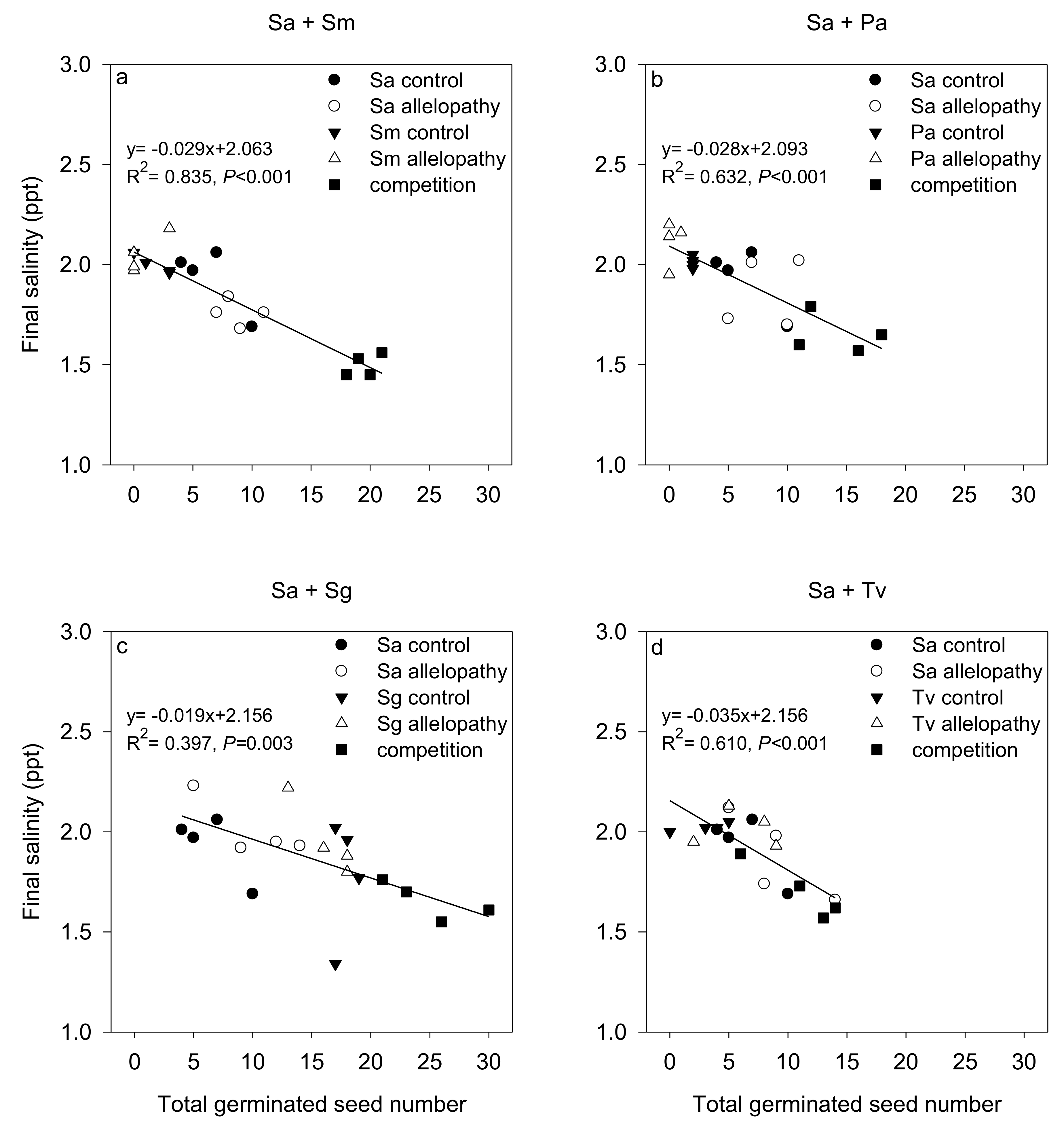
3.3. Salinity Modification during Germination
4. Discussion
4.1. Effects of Allelopathy on Seed Germination
4.2. Effects of Salinity Modification on Seed Germination
4.3. Implications for Community Assembly
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rosindell, J.; Hubbell, S.P.; Etienne, R.S. The Unified Neutral Theory of Biodiversity and Biogeography at Age Ten. Trends Ecol. Evol. 2011, 26, 340–348. [Google Scholar] [CrossRef] [PubMed]
- Rosenzweig, M.L. Species Diversity in Space and Time; Cambridge University Press: Cambridge, UK, 1995. [Google Scholar]
- Purschke, O.; Schmid, B.C.; Sykes, M.T.; Poschlod, P.; Michalski, S.G.; Durka, W.; Kuehn, I.; Winter, M.; Prentice, H.C. Contrasting changes in taxonomic, phylogenetic and functional diversity during a long-term succession: Insights into assembly processes. J. Ecol. 2013, 101, 857–866. [Google Scholar] [CrossRef]
- Xu, Y.; Dong, S.; Li, S.; Shen, H. Research progress on ecological filtering mechanisms for plant community assembly. Acta Ecol. Sin. 2019, 39, 2267–2281. [Google Scholar]
- Wang, X.; Liang, C.; Wang, W. Balance between facilitation and competition determines spatial patterns in a plant population. Chin. Sci. Bull. 2014, 59, 1405–1415. [Google Scholar] [CrossRef]
- Smith, A.L.; Atwater, D.Z.; Kim, W.; Haak, D.C.; Barney, J.N. Invasive plant rhizome production and competitiveness vary based on neighbor identity. J. Plant Ecol. 2021, 14, 638–647. [Google Scholar] [CrossRef]
- Wang, C.-h.; Li, B. Salinity and disturbance mediate direct and indirect plant-plant interactions in an assembled marsh community. Oecologia 2016, 182, 139–152. [Google Scholar] [CrossRef]
- Wehnert, A.; Wagner, S.; Huth, F. Spatio-Temporal Distribution of Carabids Influenced by Small-Scale Admixture of Oak Trees in Pine Stands. Diversity 2020, 12, 398. [Google Scholar] [CrossRef]
- Mikami, O.K.; Kawata, M. Does interspecific territoriality reflect the intensity of ecological interactions—A theoretical model for interspecific territoriality. Evol. Ecol. Res. 2004, 6, 765–775. [Google Scholar]
- Le Roux, P.C.; Shaw, J.D.; Chown, S.L. Ontogenetic shifts in plant interactions vary with environmental severity and affect population structure. New Phytol. 2013, 200, 241–250. [Google Scholar] [CrossRef] [Green Version]
- Schiffers, K.; Tielborger, K. Ontogenetic shifts in interactions among annual plants. J. Ecol. 2006, 94, 336–341. [Google Scholar] [CrossRef]
- Brooker, R.W.; Callaghan, T.V. The balance between positive and negative plant interactions and its relationship to environmental gradients: A model. Oikos 1998, 81, 196–207. [Google Scholar] [CrossRef]
- Dai, W.-l.; Wang, T.-h.; Wang, C.-h. Effects of interspecific interactions on seed germination between dominant species in the Yangtze River Estuary. Estuar. Coast. Shelf Sci. 2020, 232, 106483. [Google Scholar] [CrossRef]
- Margreiter, V.; Walde, J.; Erschbamer, B. Competition-free gaps are essential for the germination and recruitment of alpine species along an elevation gradient in the European Alps. Alp. Bot. 2021, 131, 135–150. [Google Scholar] [CrossRef]
- Elsalahy, H.; Bellingrath-Kimura, S.; Kautz, T.; Doering, T. Effects of mixing two legume species at seedling stage under different environmental conditions. Peerj 2021, 9, e10615. [Google Scholar] [CrossRef]
- Afonso, A.; Castro, S.; Loureiro, J.; Mota, L.; de Oliveira, J.C.; Torices, R. The Effects of Achene Type and Germination Time on Plant Performance in the Heterocarpic Anacyclus clavatus (Asteraceae). Am. J. Bot. 2014, 101, 892–898. [Google Scholar] [CrossRef]
- Zacchello, G.; Vinyeta, M.; Agren, J. Strong stabilizing selection on timing of germination in a Mediterranean population of Arabidopsis thaliana. Am. J. Bot. 2020, 107, 1518–1526. [Google Scholar] [CrossRef]
- Cheng, H.; Wu, B.; Yu, Y.; Wang, S.; Wei, M.; Wang, C.; Du, D. The allelopathy of horseweed with different invasion degrees in three provinces along the Yangtze River in China. Physiol. Mol. Biol. Plants 2021, 27, 483–495. [Google Scholar] [CrossRef]
- Roman, A.M.; Truta, A.M.; Viman, O.; Morar, I.M.; Spalevic, V.; Dan, C.; Sestras, R.E.; Holonec, L.; Sestras, A.F. Seed Germination and Seedling Growth of Robinia pseudoacacia Depending on the Origin of Different Geographic Provenances. Diversity 2022, 14, 34. [Google Scholar] [CrossRef]
- Fernandez-Torquemada, Y.; Luis Sanchez-Lizaso, J. Effects of salinity on seed germination and early seedling growth of the Mediterranean seagrass Posidonia oceanica (L.) Delile. Estuar. Coast. Shelf Sci. 2013, 119, 64–70. [Google Scholar] [CrossRef] [Green Version]
- Hayasaka, D.; Nakagawa, M.; Maebara, Y.; Kurazono, T.; Hashimoto, K. Seed germination characteristics of invasive Spartina alterniflora Loisel in Japan: Implications for its effective management. Sci. Rep. 2020, 10, 2116. [Google Scholar] [CrossRef] [Green Version]
- Xiaojun, Y.; Shangli, S.; Ruijun, L.; Fang, W.; Benjian, C. Research progress on effects of ecological factors on seed germination. Pratacultural Sci. 2006, 23, 44–49. [Google Scholar]
- Reynolds, C.; Cumming, G.S. Seed traits and bird species influence the dispersal parameters of wetland plants. Freshw. Biol. 2016, 61, 1157–1170. [Google Scholar] [CrossRef]
- Soons, M.B. Wind dispersal in freshwater wetlands: Knowledge for conservation and restoration. Appl. Veg. Sci. 2006, 9, 271–278. [Google Scholar] [CrossRef]
- Partzsch, M. Warming differently affects the inter- and intraspecific interactions among semi-dry grassland species. Perspect. Plant Ecol. Evol. Syst. 2019, 40, 125481. [Google Scholar] [CrossRef]
- Thorpe, A.S.; Aschehoug, E.T.; Atwater, D.Z.; Callaway, R.M. Interactions among plants and evolution. J. Ecol. 2011, 99, 729–740. [Google Scholar] [CrossRef]
- Rashidi, S.; Reza Yousefi, A.; Goicoechea, N.; Pouryousef, M.; Moradi, P.; Vitalini, S.; Iriti, M. Allelopathic Interactions between Seeds of Portulaca oleracea L. and Crop Species. Appl. Sci. 2021, 11, 3539. [Google Scholar] [CrossRef]
- Laterra, P.; Bazzalo, M.E. Seed-to-seed allelopathic effects between two invaders of burned Pampa grasslands. Weed Res. 1999, 39, 297–308. [Google Scholar] [CrossRef]
- Suman, A.; Shahi, H.N.; Singh, P.; Gaur, A. Allelopathic influence of Vigna mungo (black gram) seeds on germination and radical growth of some crop plants. Plant Growth Regul. 2002, 38, 69–74. [Google Scholar] [CrossRef]
- Zhang, S.; Liu, J.; Bao, X.; Niu, K. Seed-to-seed potential allelopathic effects between Ligularia virgaurea and native grass species of Tibetan alpine grasslands. Ecol. Res. 2011, 26, 47–52. [Google Scholar] [CrossRef]
- Mercedes Aguilar-Franco, Z.; Flores-Palacios, A.; Flores-Morales, A.; Perea-Arango, I.; de Jesus Arellano-Garcia, J.; Valencia-Diaz, S. Density-dependent effect of allelopathy on germination and seedling emergence in two Ipomoea species. Rev. Chil. De Hist. Nat. 2019, 92, 7. [Google Scholar] [CrossRef] [Green Version]
- Badano, E.I.; Cavieres, L.A. Ecosystem engineering across ecosystems: Do engineer species sharing common features have generalized or idiosyncratic effects on species diversity? J. Biogeogr. 2006, 33, 304–313. [Google Scholar] [CrossRef]
- Ma, Y.A.; Ma, Z.J. Chongming Dongtan Wetland of International Importance; China Forestry Publishing House: Beijing, China, 2006. [Google Scholar]
- Xie, B.; Han, G.; Qiao, P.; Mei, B.; Wang, Q.; Zhou, Y.; Zhang, A.; Song, W.; Guan, B. Effects of mechanical and chemical control on invasive Spartina alterniflora in the Yellow River Delta, China. Peerj 2019, 7, e7655. [Google Scholar] [CrossRef] [Green Version]
- Xiao, D.; Zhang, L.; Zhu, Z. A study on seed characteristics and seed bank of Spartina alterniflora at saltmarshes in the Yangtze Estuary, China. Estuar. Coast. Shelf Sci. 2009, 83, 105–110. [Google Scholar] [CrossRef]
- Wang, C.-H.; Tang, L.; Fei, S.-F.; Wang, J.-Q.; Gao, Y.; Wang, Q.; Chen, J.-K.; Li, B. Determinants of seed bank dynamics of two dominant helophytes in a tidal salt marsh. Ecol. Eng. 2009, 35, 800–809. [Google Scholar] [CrossRef]
- Chen, Z.Y.; Li, B.; Zhong, Y.; Chen, J.K. Local competitive effects of introduced Spartina alterniflora on Scirpus mariqueter at Dongtan of Chongming Island, the Yangtze River estuary and their potential ecological consequences. Hydrobiologia 2004, 528, 99–106. [Google Scholar] [CrossRef]
- Wang, Q.; Wang, C.H.; Zhao, B.; Ma, Z.J.; Luo, Y.Q.; Chen, J.K.; Li, B. Effects of growing conditions on the growth of and interactions between salt marsh plants: Implications for invasibility of habitats. Biol. Invasions 2006, 8, 1547–1560. [Google Scholar] [CrossRef]
- Yuan, Y.; Wang, K.; Li, D.; Pan, Y.; Lv, Y.; Zhao, M.; Gao, J. Interspecific Interactions between Phragmites australis and Spartina alterniflora along a Tidal Gradient in the Dongtan Wetland, Eastern China. PLoS ONE 2013, 8, e53843. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Z.; Zhang, L.; Xiao, D. Seed production of Spartina alterniflora and its response of germination to temperature at Chongming Dongtan, Shanghai. Acta Ecol. Sin. 2011, 31, 1574–1581. [Google Scholar]
- Wang, Q.; Shi, H.H.; Yu, Z.L.; Wang, T.H.; Wang, C.H. Effects of salinity and interspecific interaction on germination and growth of Scirpus mariqueter and Spartina alterniflora. Acta Ecol. Sin. 2022, 42, 8300–8310. [Google Scholar] [CrossRef]
- Lissner, J.; Schierup, H.H. Effects of salinity on the growth of Phragmites australis. Aquat. Bot. 1997, 55, 247–260. [Google Scholar] [CrossRef]
- Peng, Y.; Xie, T.; Zhou, F.; Wan, H.; Zhang, C.; Zhai, R.; Zheng, Q.; Zheng, C.; Liu, Z. Response of plant growth and photosynthetic characteristics in Suaeda glauca and Atriplex triangularis seedlings to different concentrations of salt treatments. Acta Prataculturae Sin. 2012, 21, 64–74. [Google Scholar]
- Gong, J.-n.; Wang, K.-y.; Zhang, C.; Ma, Y.-l. Invasion and its effects of xerarch halophytes in reclaimed tidal wetlands. J. Appl. Ecol. 2009, 20, 33–39. [Google Scholar]
- De Souza Coelho, L.C.; Broch Mignoni, D.S.; Barbedo, C.J.; Braga, M.R. Seed leachates of the tropical legume Sesbania virgata: Their effects on germination and seedling growth of tomato and rice. Acta Physiol. Plant. 2022, 44, 96. [Google Scholar] [CrossRef]
- Schmer, M.R.; Xue, Q.; Hendrickson, J.R. Salinity effects on perennial, warm-season (C-4) grass germination adapted to the northern Great Plains. Can. J. Plant Sci. 2012, 92, 873–881. [Google Scholar] [CrossRef] [Green Version]
- Brenchley, J.L.; Probert, R.J. Seed germination responses to some environmental factors in the seagrass Zostera capricorni from eastern Australia. Aquat. Bot. 1998, 62, 177–188. [Google Scholar] [CrossRef]
- Pellissier, F. Improved germination bioassays for allelopathy research. Acta Physiol. Plant. 2013, 35, 23–30. [Google Scholar] [CrossRef]
- Guido, A.; Hoss, D.; Pillar, V.D. Exploring seed to seed effects for understanding invasive species success. Perspect. Ecol. Conserv. 2017, 15, 234–238. [Google Scholar] [CrossRef]
- Baskin, C.C.; Baskin, J.M.; Chester, D.W.; Smith, M. Ethylene as a possible cue for seed germination of Schoenoplectus hallii (Cyperaceae), a rare summer annual of occasionally flooded sites. Am. J. Bot. 2003, 90, 620–627. [Google Scholar] [CrossRef]
- Song, J.; Shi, W.; Liu, R.; Xu, Y.; Sui, N.; Zhou, J.; Feng, G. The role of the seed coat in adaptation of dimorphic seeds of the euhalophyte Suaeda salsa to salinity. Plant Species Biol. 2017, 32, 107–114. [Google Scholar] [CrossRef]
- Sun, S.C.; Gao, X.M.; Cai, Y.L. Variations in sexual and asexual reproduction of Scirpus mariqueter along an elevational gradient. Ecol. Res. 2001, 16, 263–274. [Google Scholar] [CrossRef]
- Wu, G.-L.; Du, G.-Z.; Shi, Z.-H. Germination strategies of 20 alpine species with varying seed mass and light availability. Aust. J. Bot. 2013, 61, 404–411. [Google Scholar] [CrossRef]
- McKee, J.; Richards, A.J. Variation in seed production and germinability in common reed (Phragmites australis) in Britain and France with respect to climate. New Phytol. 1996, 133, 233–243. [Google Scholar] [CrossRef]
- Zhang, H.; Irving, L.J.; McGill, C.; Matthew, C.; Zhou, D.; Kemp, P. The effects of salinity and osmotic stress on barley germination rate: Sodium as an osmotic regulator. Ann. Bot. 2010, 106, 1027–1035. [Google Scholar] [CrossRef] [Green Version]
- Rehman, S.; Harris, P.J.C.; Bourne, W.F.; Wilkin, J. The effect of sodium chloride on germination and the potassium and calcium contents of Acacia seeds. Seed Sci. Technol. 1997, 25, 45–57. [Google Scholar]
- Dyer, A.R.; Fenech, A.; Rice, K.J. Accelerated seedling emergence in interspecific competitive neighbourhoods. Ecol. Lett. 2000, 3, 523–529. [Google Scholar] [CrossRef]
- Williams, L.K.; Kristiansen, P.; Sindel, B.M.; Wilson, S.C.; Shaw, J.D. Quantifying the seed bank of an invasive grass in the sub-Antarctic: Seed density, depth, persistence and viability. Biol. Invasions 2016, 18, 2093–2106. [Google Scholar] [CrossRef]
- Orrock, J.L.; Christopher, C.C. Density of Intraspecific Competitors Determines the Occurrence and Benefits of Accelerated Germination. Am. J. Bot. 2010, 97, 694–699. [Google Scholar] [CrossRef]
- Fenesi, A.; Kelemen, K.; Sandor, D.; Ruprecht, E. Influential neighbours: Seeds of dominant species affect the germination of common grassland species. J. Veg. Sci. 2020, 31, 1028–1038. [Google Scholar] [CrossRef]
- Houseman, G.R.; Mahoney, A.K. Intraspecific seed interactions alter seedling emergence of Lespedeza cuneata under field conditions. Popul. Ecol. 2015, 57, 539–544. [Google Scholar] [CrossRef]
- Tielboerger, K.; Prasse, R. Do seeds sense each other? Testing for density-dependent germination in desert perennial plants. Oikos 2009, 118, 792–800. [Google Scholar] [CrossRef]
- Tang, L.; Wolf, A.A.; Gao, Y.; Wang, C.H. Photosynthetic tolerance to non-resource stress influences competition importance and intensity in an invaded estuary. Ecology 2018, 99, 1327–1337. [Google Scholar] [CrossRef]
- Li, B.; Liao, C.-H.; Zhang, X.-D.; Chen, H.-L.; Wang, Q.; Chen, Z.-Y.; Gan, X.-J.; Wu, J.-H.; Zhao, B.; Ma, Z.-J.; et al. Spartina alterniflora invasions in the Yangtze River estuary, China: An overview of current status and ecosystem effects. Ecol. Eng. 2009, 35, 511–520. [Google Scholar] [CrossRef]
- Hu, Z.; Ma, Q.; Cao, H.; Zhang, Z.; Tang, C.; Zhang, L.; Ge, Z. A trial study on revegetation of the native Scirpus mariqueter population in the coastal wetland of the Yangtze Estuary. Ecol. Sci. 2016, 35, 1–7. [Google Scholar]
- Shi, H.H.; Xue, Q.; Yu, Z.L.; Wang, C.H. Effects of density and species proportion on intraspecific and interspecific interactions between salt marsh plants during germination. Chin. J. Plant Ecol. 2022, in press. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source of Variation | Sa + Sm (Sa) | Sa + Pa (Sa) | Sa + Sg (Sa) | Sa + Tv (Sa) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Df | F | p | Df | F | p | Df | F | p | Df | F | p | |
| treatment | 2 | 11.86 | 0.003 | 2 | 2.55 | 0.133 | 2 | 2.11 | 0.177 | 2 | 0.30 | 0.746 |
| Source of variation | Sa + Sm (Sm) | Sa + Pa (Pa) | Sa + Sg (Sg) | Sa + Tv (Tv) | ||||||||
| Df | F | p | Df | F | p | Df | F | p | Df | F | p | |
| treatment | 2 | 14.35 | 0.002 | 2 | 4.52 | 0.044 | 2 | 1.91 | 0.204 | 2 | 1.28 | 0.325 |
| Source of Variation | Sa + Sm (Sa) | Sa + Pa (Sa) | Sa + Sg (Sa) | Sa + Tv (Sa) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Df | F | p | Df | F | p | Df | F | p | Df | F | p | |
| treatment | 2 | 5.40 | 0.029 | 2 | 8.03 | 0.010 | 2 | 4.51 | 0.044 | 2 | 5.27 | 0.030 |
| Source of variation | Sa + Sm (Sm) | Sa + Pa (Pa) | Sa + Sg (Sg) | Sa + Tv (Tv) | ||||||||
| Df | F | p | Df | F | p | Df | F | p | Df | F | p | |
| treatment | 2 | 8.31 | 0.026 | 2 | 1.60 | 0.291 | 2 | 5.32 | 0.030 | 2 | 11.02 | 0.005 |
| Source of Variation | Sa + Sm (Sa) | Sa + Pa (Sa) | Sa + Sg (Sa) | Sa + Tv (Sa) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Df | F | p | Df | F | p | Df | F | p | Df | F | p | |
| treatment | 2 | 16.49 | 0.001 | 2 | 3.82 | 0.063 | 2 | 7.09 | 0.014 | 2 | 1.85 | 0.212 |
| Source of variation | Sa + Sm (Sm) | Sa + Pa (Pa) | Sa + Sg (Sg) | Sa + Tv (Tv) | ||||||||
| Df | F | p | Df | F | p | Df | F | p | Df | F | p | |
| treatment | 2 | 78.77 | <0.001 | 2 | 30.89 | <0.001 | 2 | 2.00 | 0.191 | 2 | 13.33 | 0.002 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, C.-H.; Yu, Z.-L.; Yasenjiang, Y.; Tang, L.; Gao, Y.; Zou, C.-J. Mechanisms of Seed-To-Seed Interactions between Dominant Species in the Yangtze River Estuary under Saline Condition. Diversity 2022, 14, 1017. https://doi.org/10.3390/d14121017
Wang C-H, Yu Z-L, Yasenjiang Y, Tang L, Gao Y, Zou C-J. Mechanisms of Seed-To-Seed Interactions between Dominant Species in the Yangtze River Estuary under Saline Condition. Diversity. 2022; 14(12):1017. https://doi.org/10.3390/d14121017
Chicago/Turabian StyleWang, Cheng-Huan, Zhen-Lin Yu, Yuerenisha Yasenjiang, Long Tang, Yang Gao, and Chun-Jing Zou. 2022. "Mechanisms of Seed-To-Seed Interactions between Dominant Species in the Yangtze River Estuary under Saline Condition" Diversity 14, no. 12: 1017. https://doi.org/10.3390/d14121017