
Two New Species of the Family Canuellidae Lang, 1944 (Copepoda: Polyarthra), from Korea, with a Key to Species of the Genus Scottolana Huys, 2009 †


1
Division of Biological Sciences, Mokwon University, Daejeon 35349, Korea
2
Marine Art Platform, Daejeon 35349, Korea
3
Department of Taxonomy and Systematics, National Marine Biodiversity Institute of Korea, Seocheon 33662, Korea
*
Author to whom correspondence should be addressed.
†
urn:lsid:zoobank.org:pub:AFA1EE39-A46E-4745-8690-FE82C8D44E28.
Diversity 2022, 14(11), 967; https://doi.org/10.3390/d14110967
Submission received: 10 October 2022
/
Revised: 7 November 2022
/
Accepted: 8 November 2022
/
Published: 10 November 2022
(This article belongs to the Collection Collection of Experts’ Researches on Aquatic Life (CEREAL))
Abstract
:Two new species of the genus Scottolana Huys, 2009, were collected from Korean waters. S. daecheonensis sp. nov. is morphologically similar to S. jasani Song, Kangtia, Khim, and Chullasorn, 2018, from the Yellow Sea, Korea, and S. gomezi Nazari, Mirshamsi, Sari, Aliabadian, and Martinez Arbizu, 2018, from Bandar Abbas, Iran. However, it can clearly be distinguished from its congeners by rostrum with two tiny lateral sensilla, 4-segemented antennule in female, and 5-segmented antennule in male, 3-segmented mandibular exopod, P3 basis with an inner spinous process on the posterior surface, P3 endopod-3 without a modified tube-pore along the inner margin in male, P4 endopod-2 crumpled inner margin with strong spinules in male, P5 second outermost seta naked, caudal rami with an unguiform extension on the inner margins, and caudal seta V with a normal base in female. S. wonchoeli sp. nov. is similar to S. geei Mu and Huys, 2004 from the Bohai Sea, and S. longipes (Thompson I.C. and Scott A., 1903) sensu Wells and Rao (1987) from the Andaman Islands, but it can clearly be distinguished from its congeners by a combination of the following features: rostrum with a broadly rounded apex, 3-segemented antennule in female and 5-segmented in male, antennary exopod 8-segmented, female genital field relatively small, extending about one-third of genital double-somite, genital somite with paired lateral spinous processes in male, P6 with a long proximal element in male, and maxillular endopod with 11 setae in total. In a molecular analysis using the mitochondrial cytochrome c oxidase subunit I (COI) and 18S ribosomal RNA (18S rRNA) genes, the interspecific distance was 20.031% for COI and 3.214% for 18S rRNA between the two new species. A key for the identification of the species of Scottolana is included.
1. Introduction
Lang [1] proposed Polyarthra Lang, 1944 as a suborder of the order Harpacticoida Sars G.O., 1903, consisting of only two families, Canuellidae Lang, 1944, and Longipediidae Boeck, 1865. Dahms [2,3] provided valid reasons why Polyarthra should be excluded from the order Harpacticoida, and suggested raising it to full order rank. Khodami et al. [4] proposed the new order Canuelloida Khodami, MacArthur, Blanco-Bercial and Martinez Arbizu, 2017 comprising the family Canuellidae and the family Longipediidae, but data inaccuracies and data processing issues were discovered [5], prompting the authors to withdraw the paper. Therefore, the Polyarthra is the oldest and only available name for this taxon at present.
The family Canuellidae comprises about 62 valid species in 18 genera: Sunaristes Hesse, 1867 (4 species), Canuella Scott T. and Scott A., 1893 (5 species), Brianola Monard, 1926 (8 species), Canuellina Gurney, 1927 (6 species), Canuellopsis Lang, 1936 (four species), Ifanella Vervoort, 1964 (monospecific), Ellucana Coull, 1971 (2 species), Galapacanuella Mielke, 1979 (monospecific), Intercanuella Becker and Schriever, 1979 (monospecific), Parasunaristes Fiers, 1982 (2 species), Coullana Por, 1984 (monospecific), Elanella Por, 1984 (3 species), Nathaniella Por, 1984 (monospecific), Microcanuella Mielke, 1994 (2 species), Echinosunaristes Huys, 1995 (monospecific), Intersunaristes Huys, 1995 (2 species), Scottolana Huys, 2009 (17 species), and Indicanuella Huys, 2016 (monospecific) [6].
The most speciose genus Scottolana Huys, 2009 is currently composed of 17 valid species, showing a broad global distribution, and it has been found in the Pacific Ocean— (S. bulbifera (Chislenko, 1971) [7]; S. dissimilis Fiers, 1982 [8]; S. glabra Fiers, 1982 [8]; S. geei Mu and Huys, 2004 [9]; S. huysi Song, Kangtia, Khim, and Chullasorn, 2018 [10]; and S. jasani Song, Kangtia, Khim, and Chullasorn, 2018 [10]); the Indian Ocean—(S. inopinata (Thompson I.C. and Scott A., 1903) [11]; S. longipes (Thompson I.C. and Scott A., 1903) [11]; S. scotti (Sewell, 1940) [12]; S. brevifurca (Wells, 1967) [13]; S. longipes sensu Wells, 1967 [13]; S. longipes sensu Wells and Rao, 1987 [14]; S. oleosa, Wells and Rao (1987) [14]; S. rostrata Wells and Rao, 1987 [14]; and S. tumidiseta Wells and Rao, 1987 [14]); the Atlantic Ocean—(S. antillensis Fiers, 1984 [15]), and the Mediterranean Sea—(S. bulbosa (Por, 1964) [16]; S. longipes sensu Por (1964) [16]; S. uxoris Por, 1983 [17]; and S. gomezi Nazari, Mirshamsi, Sari, Aliabadian, and Martinez Arbizu, 2018 [18]).
Until now, 3 species of the genus Scottolana have been reported around the Korean peninsula—S. bulbifera from the Posyet Bay, Russia [7], the Bohai Sea, China [9], and the South Sea of Korea [19]; S. geei from the Bohai Sea, China [9]; and S. jasani from the Yellow Sea, Korea [10]. Two new species of Scottolana were collected from Korean waters. The present study provides the detailed morphological descriptions of two new species, the sequences of mitochondrial cytochrome c oxidase subunit I and the 18S ribosomal RNA gene as a DNA barcode, and the updated identification key to species of the genus.
2. Material and Methods
2.1. Sampling Locations and Methods
Specimens were collected using a light trap, hand net, or by SCUBA diving, and then fixed with 99% ethanol (Table 1). The Ludox centrifugation method was used to extract benthic copepods from the sediments [20]. Using a Leica M80 stereomicroscope, specimens were separated from copepod samples. The sorted specimens were stored at −20 °C until DNA extraction was performed. After DNA extraction, all specimens were morphologically described.
2.2. DNA Extraction and Sequencing
For non-destructive DNA extraction, each specimen was washed in distilled water before being placed in a 1.5 mL tube with 20 mL Proteinase K and 180 mL ATL buffer [21]. The procedure for extracting DNA was then carried out in accordance with the instructions given by Qiagen (DNeasy Blood and Tissue Kit, Qiagen, Hilden, Germany). Using an AccuPower HotStart PCR PreMix (Bioneer, Daejeon, Korea), COI and 18S rRNA sequences were amplified from the sample DNAs. Table 2 showed the conditions used for the PCR and the primers. The PCR products were sequenced using an ABI PRISM 3730XL Analyzer (Macrogen Inc., Seoul, Korea) in both directions. Geneious 10.1.3 (Biomatters, Auckland, New Zealand) was used to assemble the sequences [22]. In Geneious 10.1.3, pairwise distances were calculated using the Tamura–Nei distance model [23]. The sequences from GenBank were aligned using the MUSCLE algorithm, which is built into Geneious 10.1.3 [24]. For the maximum likelihood analysis, six additional sequences were downloaded from GenBank. The analysis was performed using the Hasegawa–Kishino–Yano (mitochondrial COI) model [25], based on the model test result from MEGA X (MEGA, University of Pennsylvania, Philadelphia, PA, USA) [26]. The bootstrap was performed with 1000 replications.
2.3. Morphological Characterization
Specimens were dissected in lactic acid and mounted on slides using lactophenol as a mounting medium. Transparent nail varnish was used to seal the preparations. All drawings were created on a Leica DM 2500 differential interference contrast microscope with a camera lucida. The descriptive terminology regarding the body and appendage morphology is adopted from Huys and Boxshall [27]. Abbreviations used in the text are: A1 for antennule; A2 for antenna; ae for aesthetasc; exp for exopod; enp for endopod; P1–P6 for the first to the sixth thoracopod; exp (enp)-1 (-2, -3) to denote the proximal (middle, distal) segment of a ramus. All specimens were deposited in the National Marine Biodiversity Institute of Korea (MABIK).
3. Results
- Order Polyarthra Lang, 1944
- Family Canuellidae Lang, 1944
- Genus Scottolana Huys, 2009
3.1. Scottolana Daecheonensis sp. nov.
Figure 1, Figure 2, Figure 3, Figure 4, Figure 5, Figure 6, Figure 7, Figure 8, Figure 9, Figure 10 and Figure 11 are listed bellow.
Type locality—small rock pool in intertidal area, Daecheon Beach, Boryeong, Korea (36°18′00″ N 126°31′02″ E).
Material examined—Holotype 1♀ (MABIK CR00252814) dissected on 8 slides. Paratype 2♂♂ (MABIK CR00252815, and CR00252816) dissected on 10 slides and 9 slides each, and 3♂♂ (MABIK CR00252811–CR00252813) in 99% alcohol.
Etymology—The specific name refers to the type locality, which is the west coast of Korea.
Genbank accession number—mitochondrial cytochrome c oxidase subunit I gene (OP452900–OP452905); 18S ribonucleic acid gene (OP454041, OP454042).
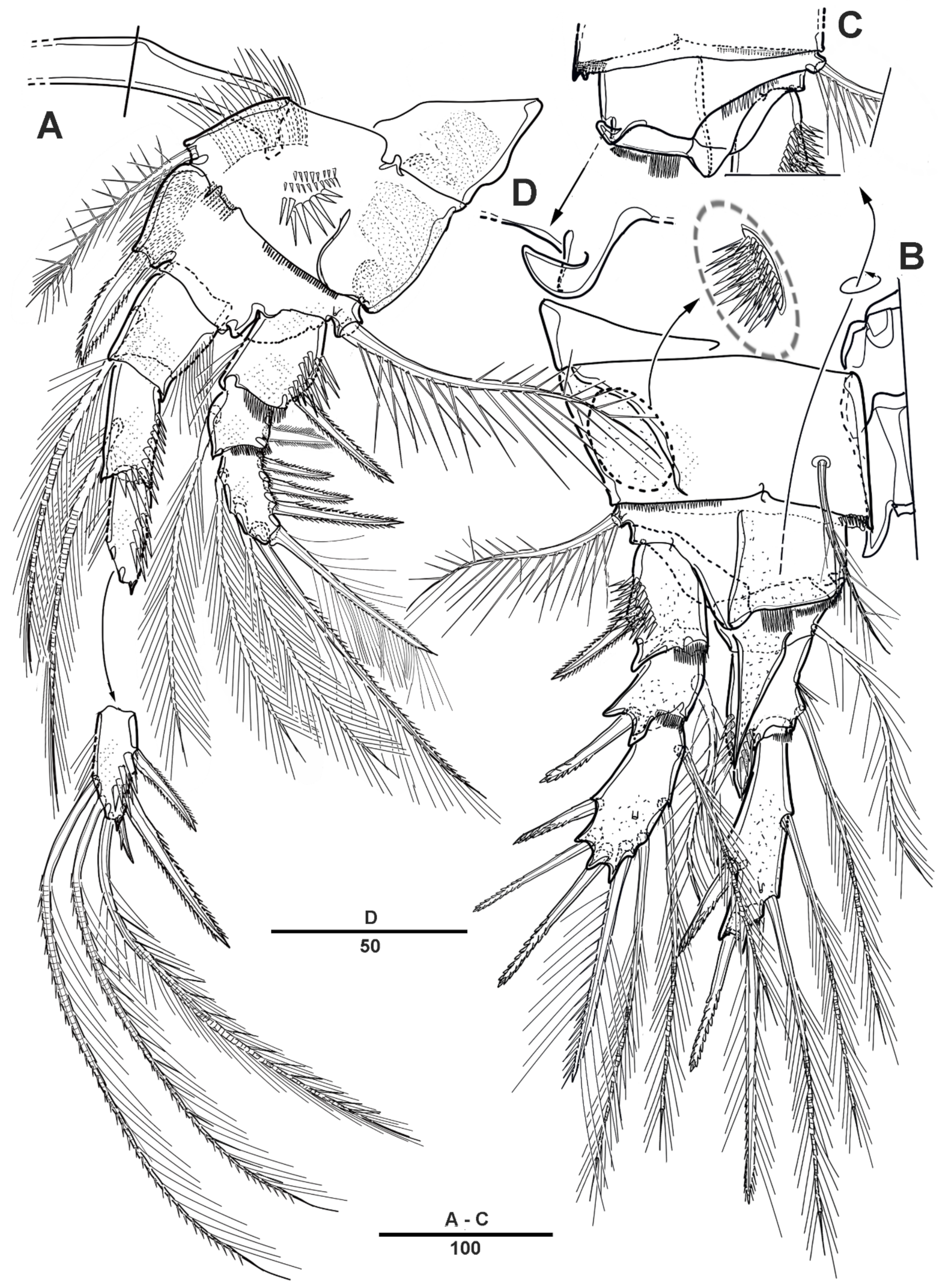
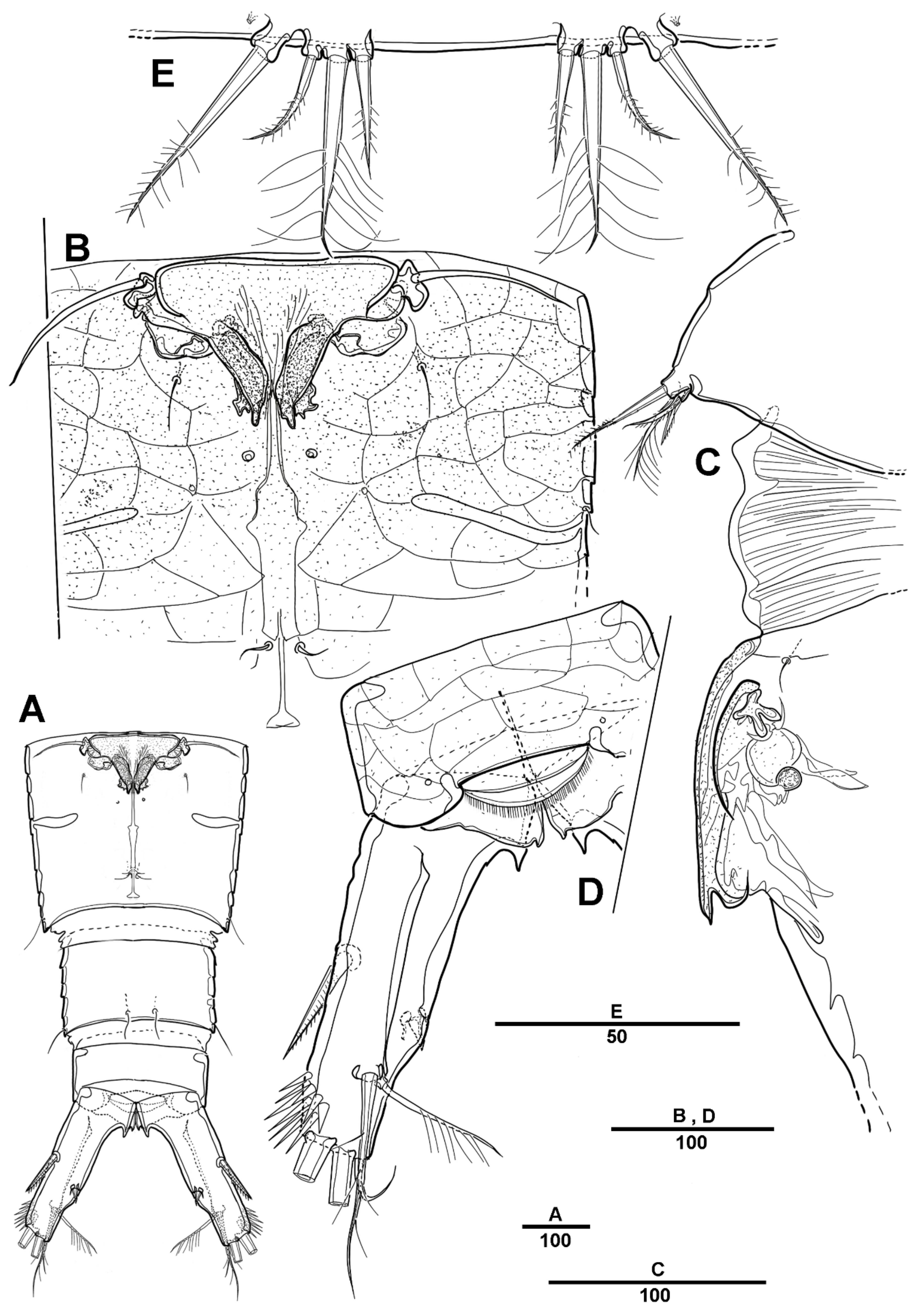
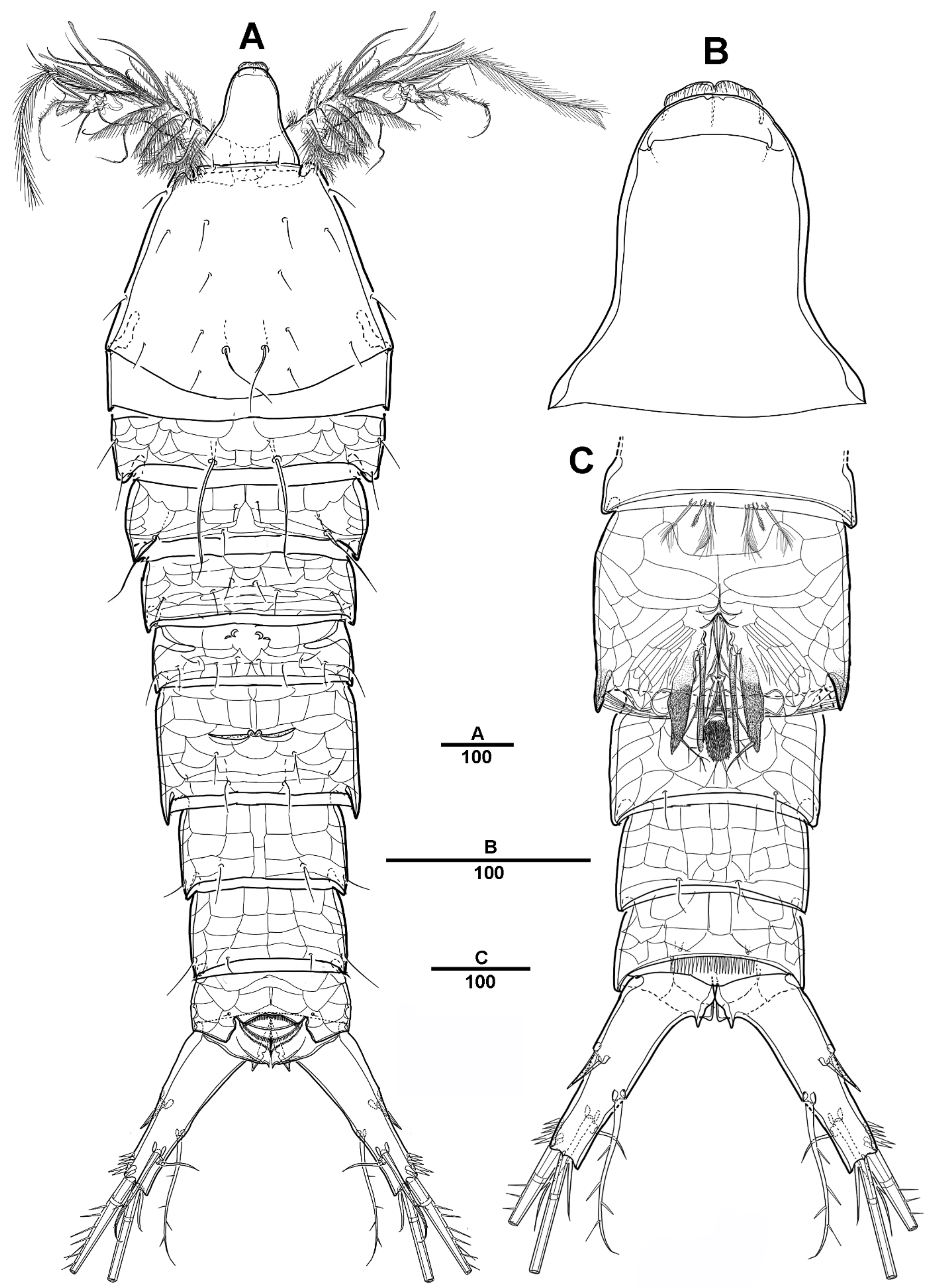
Female—habitus (Figure 1A,B). Total body length 1940 µm, measured from the anterior margin of rostrum to the posterior margin of caudal rami. Largest width measured at the posterior margin of cephalic shield: 490 µm. Body large, robust, semi-cylindrical, and the posterior margin with smooth hyaline frill. Prosome laterally compressed.
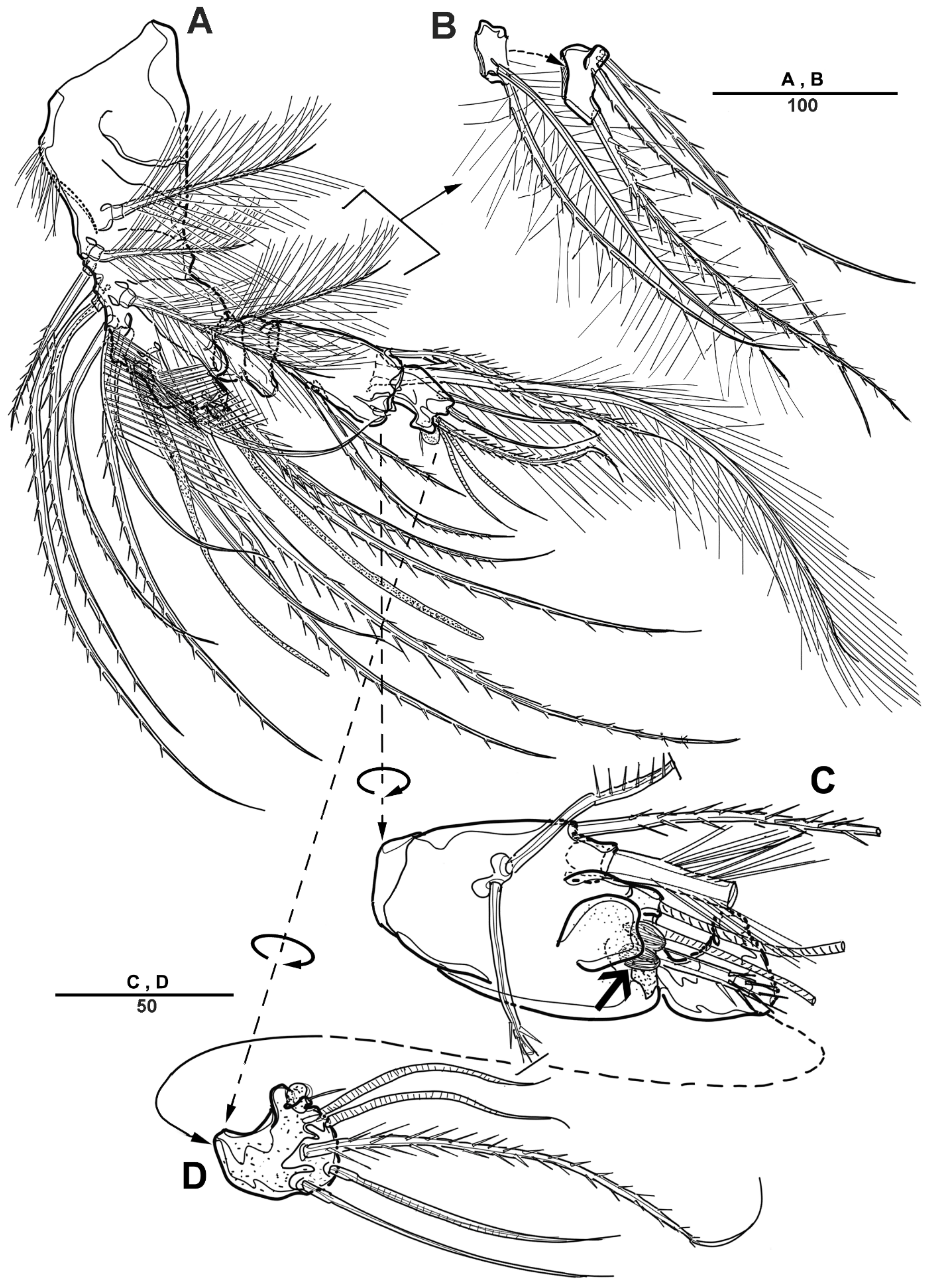
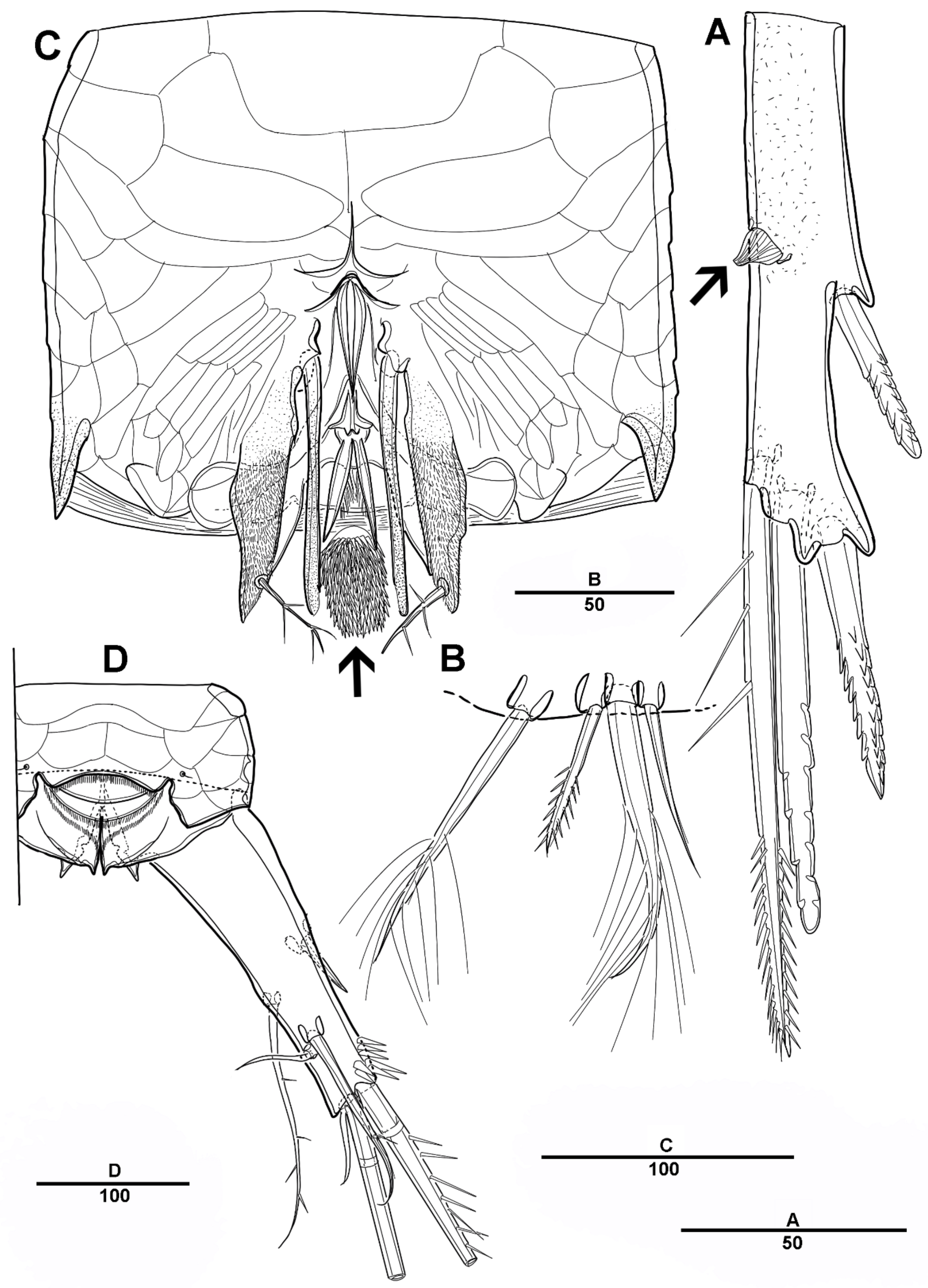
Cephalic shield gradually tapering anteriorly, and the dorsal surface with long sensilla and a huge pore, as shown in Figure 1A. Rostrum (Figure 1A) prominent, steeple-shaped, defined at base, with distal third tapering sharply, and with two tiny lateral sensilla. The first pedigerous somite not fused to cephalosome. Pedigerous somites with pores and sensillae as figured. Urosome slightly narrower than prosome. Urosome (Figure 1A,B and Figure 2A) consists of P5-bearing somite, genital double-somite and two free urosomites. Genital double-somite (Figure 2A), original segmentation marked by paired dorsolateral and ventrolateral internal chitinous ribs. Genital field (Figure 1C and Figure 2B) large, copulatory pores paired, and covered by flaps. Gonopores closed off by vestigial P6 bearing one long smooth seta and an internal projection. Anal operculum (Figure 1D) weakly developed, with a transverse row of setules. Caudal rami (Figure 1D and Figure 2C) about 2.3 times longer than the width at base, with an unguiform projection on the inner proximal margin; the outer margin and the posteroventral surface with strong spinules; Each ramus armed with seven setae: seta I short, spiniform, pinnate; seta II long, pinnate, located halfway along the inner margin; seta III pinnate; seta IV pinnate and long; seta V longest; seta VI bare and slender; seta VII short and smooth.
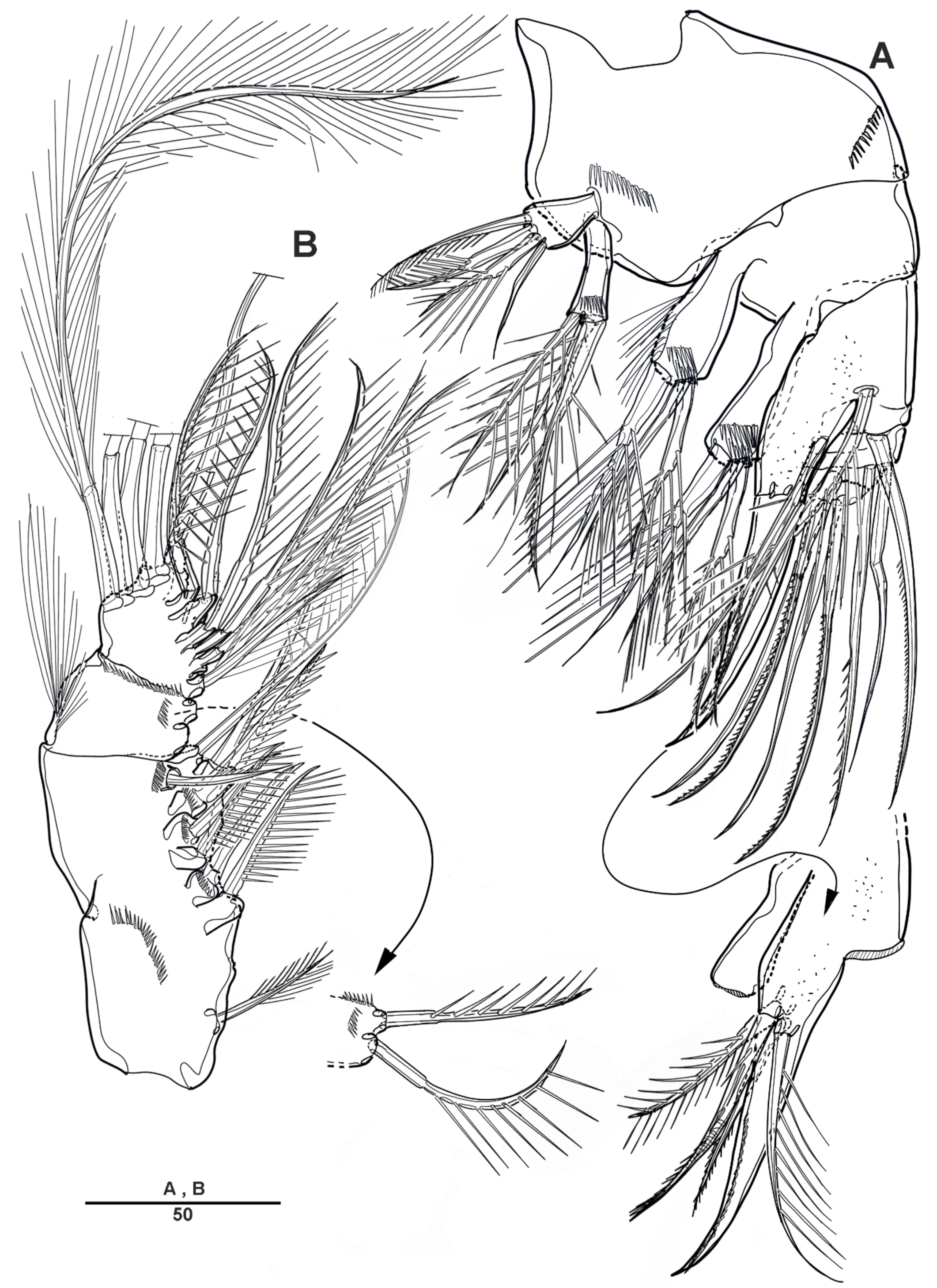
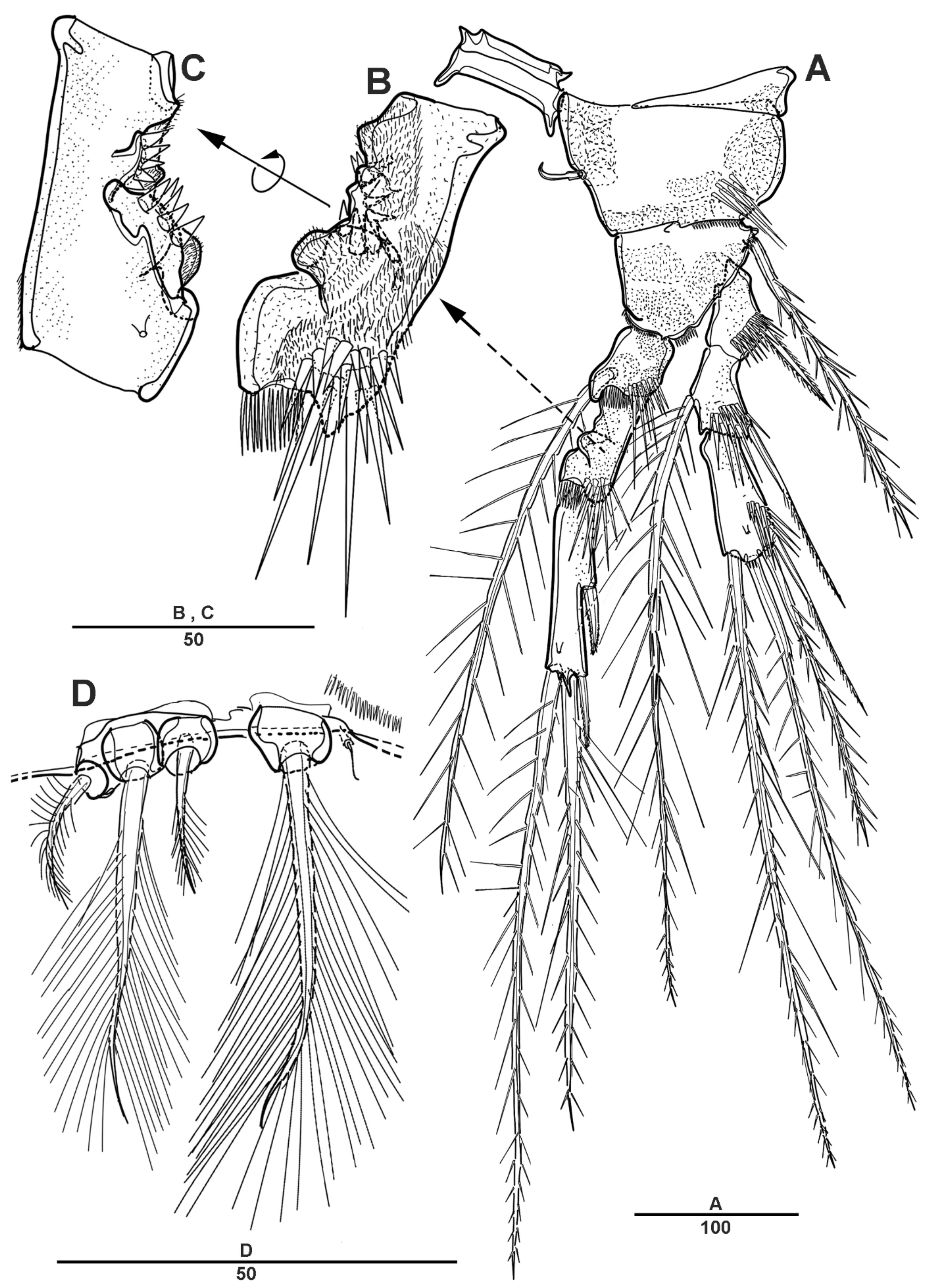
Antennule (Figure 3A) 4-segmented. Segment 1 with long setules around the anterior margin, the longest and the largest, with four incomplete sutures around the posterior margin, with eleven pinnate or plumose setae, four of them bi-articulate at base; segment 2, with four naked and eight spinulose setae, and two aesthetascs; segment 3 short, with two bare and two pinnate setae; and last segment with two long naked setae apically, one plumose seta posteriorly, one short and bare seta, and seven pinnate setae. Armature as follows: 1-[11], 2-[12 + 2ae], 3-[4], 4-[12].
Antenna (Figure 3B) biramous, coxa and basis fused to form short coxo-basis, with long setules around the distal margin. Exopod 8-segmented; first to seventh segments each with one seta, last segment with three pinnate and one plumose setae. Endopod 2-segmented; enp-1 with one pinnate and one small bare setae; enp-2 with incomplete suture indicating original segmentation, with one small bare, two long pinnate, and one long smooth setae laterally, and six pinnate, and one bare setae distally.
Mandible (Figure 4A) with well-developed gnathobase bearing six strong multicuspidate teeth as figured, dorsal corner with one pinnate spine, with small spinules around the dorsal margin. Basis with two pinnate inner setae. Palp well-developed, biramous. Exopod 3-segmented, all setae plumose; exp-1 with two long setae; exp-2 with row of spinules at the outer margin, with one long seta; exp-3 with one seta proximally, and two long setae along the distal margin. Endopod 2-segmented; enp-1 with three setae; enp-2 with eight setae.
Maxillule (Figure 4B). Praecoxa and coxa partially fused. Praecoxal arthrite with eight spines and two setae around the distal margin, and two setae on the anterior surface. Coxo-endite with five pinnate or plumose setae, and epipodite represented by two setae. Basis with spinular rows on the posterior surface, basal proximal endite with three setae, and distal endite with four setae. Exopod l-segmented, all setae plumose, with six inner setae, five distal setae, and one small outer seta. Endopod 2-segmented; enp-1 with three pinnate and two plumose setae; enp-2 with six plumose setae.
Maxilla (Figure 5A) comprising praecoxa, coxa, allobasis and 3-segmented endopod. Praecoxa with two endites; proximal endite with three pinnate and one bare setae; distal endite with three pinnate elements. Coxa with two endites, each with three spinulose elements. Allobasis forming strong unipinnate claw with four pinnate setae, and with two pinnate setae on the anterior surface. Endopod 3-segmented; enp-1 with one smooth and two spiniform and elements; enp-2 with one spinulose and two naked elements; enp-3 small, with two setae.
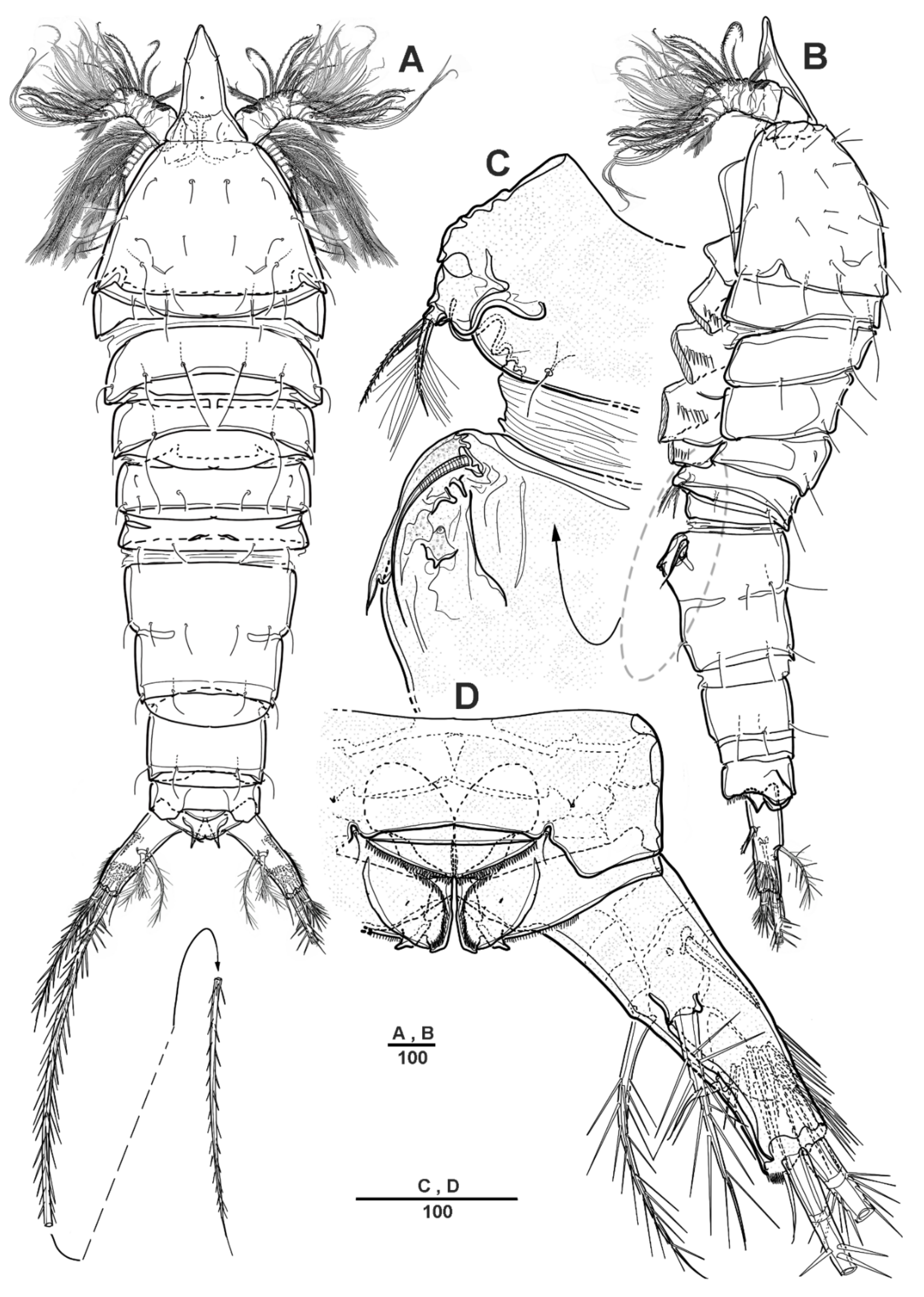
Figure 1.
Scottolana daecheonensis sp. nov. holotype female: (A) habitus, dorsal; (B) habitus, lateral; (C) P5 and genital field, lateral; (D) anal somite and right caudal ramus, dorsal. Scale bars in μm.
Figure 1.
Scottolana daecheonensis sp. nov. holotype female: (A) habitus, dorsal; (B) habitus, lateral; (C) P5 and genital field, lateral; (D) anal somite and right caudal ramus, dorsal. Scale bars in μm.

Figure 2.
Scottolana daecheonensis sp. nov. holotype female: (A) urosome, ventral; (B) genital field, ventral; (C) anal somite and left caudal ramus, ventral; (D) P5. Scale bars in μm.
Figure 2.
Scottolana daecheonensis sp. nov. holotype female: (A) urosome, ventral; (B) genital field, ventral; (C) anal somite and left caudal ramus, ventral; (D) P5. Scale bars in μm.

Figure 3.
Scottolana daecheonensis sp. nov. holotype female: (A) antennule (inset showing armature on segment 3); (B) antenna. Scale bars in μm.
Figure 3.
Scottolana daecheonensis sp. nov. holotype female: (A) antennule (inset showing armature on segment 3); (B) antenna. Scale bars in μm.

Figure 4.
Scottolana daecheonensis sp. nov. holotype female: (A) mandible; (B) maxillule. Scale bars in μm.
Figure 4.
Scottolana daecheonensis sp. nov. holotype female: (A) mandible; (B) maxillule. Scale bars in μm.

Figure 5.
Scottolana daecheonensis sp. nov. holotype female: (A) maxilla (inset showing armature on allobasis); (B) maxilliped. Scale bars in μm.
Figure 5.
Scottolana daecheonensis sp. nov. holotype female: (A) maxilla (inset showing armature on allobasis); (B) maxilliped. Scale bars in μm.

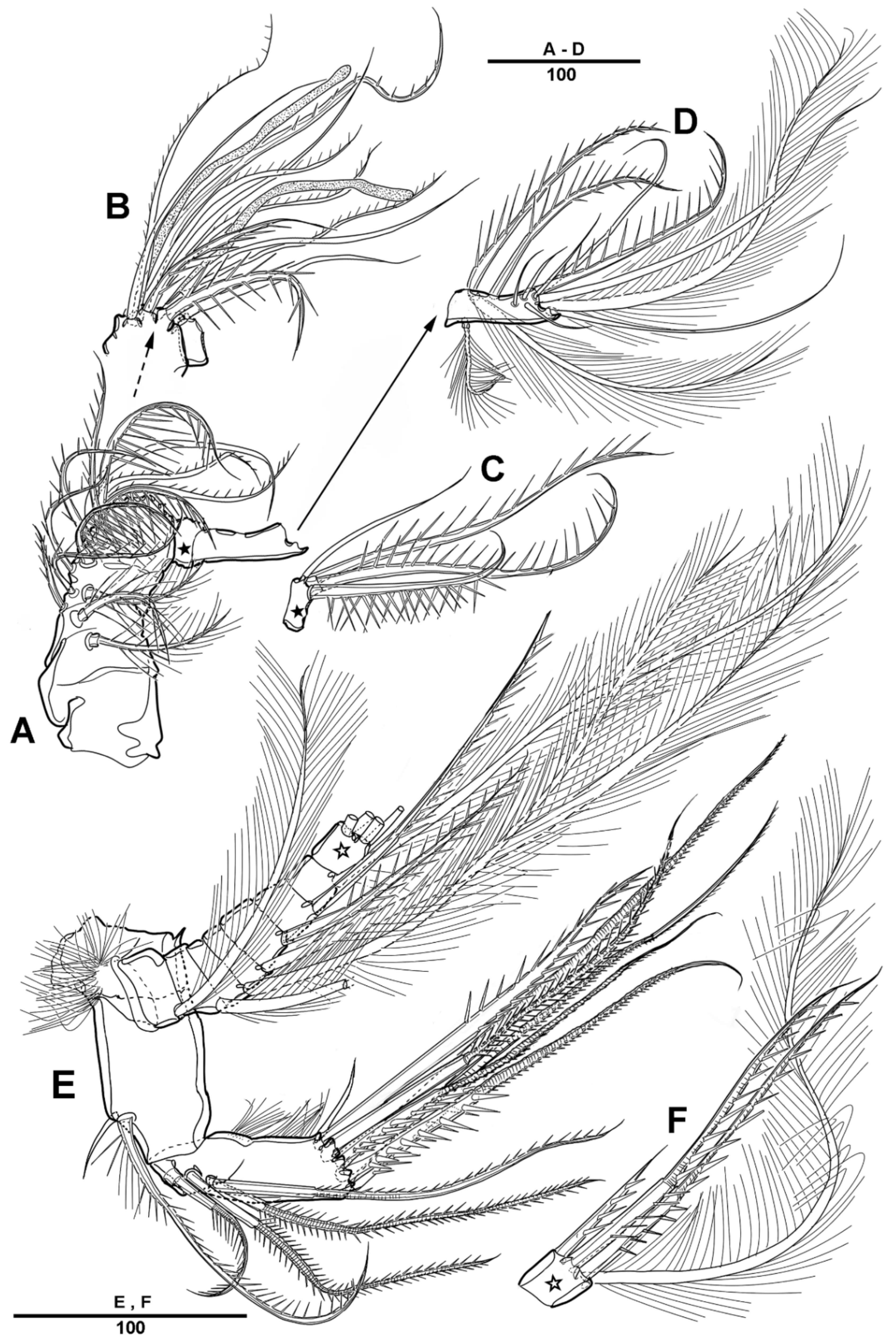
Maxilliped (Figure 5B) phyllopodial, comprising syncoxa, basis and 1-segmented endopod. Syncoxa with nine elements on the inner margin. Endopod with eleven elements.
Swimming legs 1–4 (Figure 6 and Figure 7) with intercoxal sclerites and praecoxae, both without ornamentation. Coxae and bases with a surface spinular pattern as figured. All swimming legs with 3-segmented exopods and endopods.
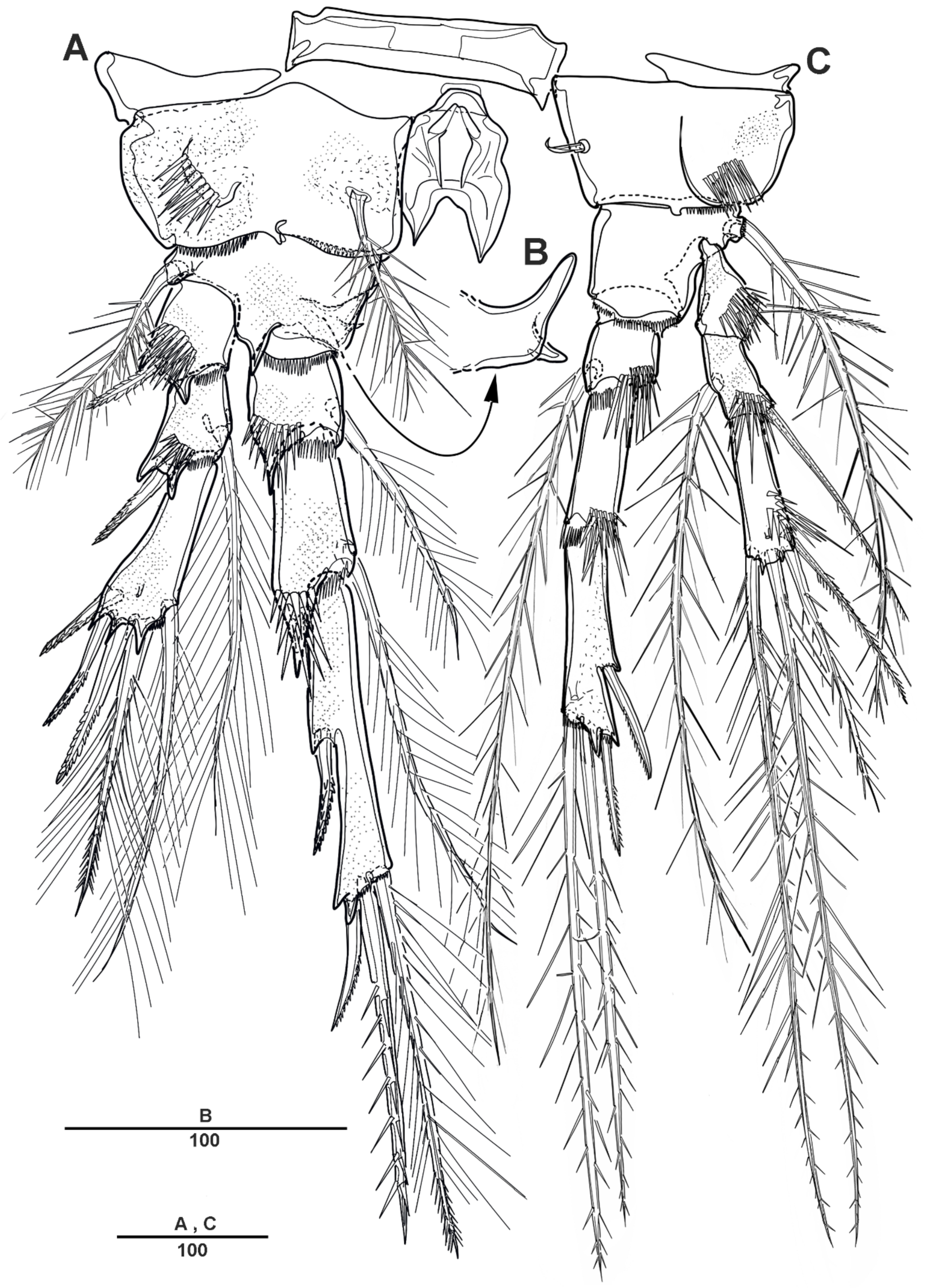
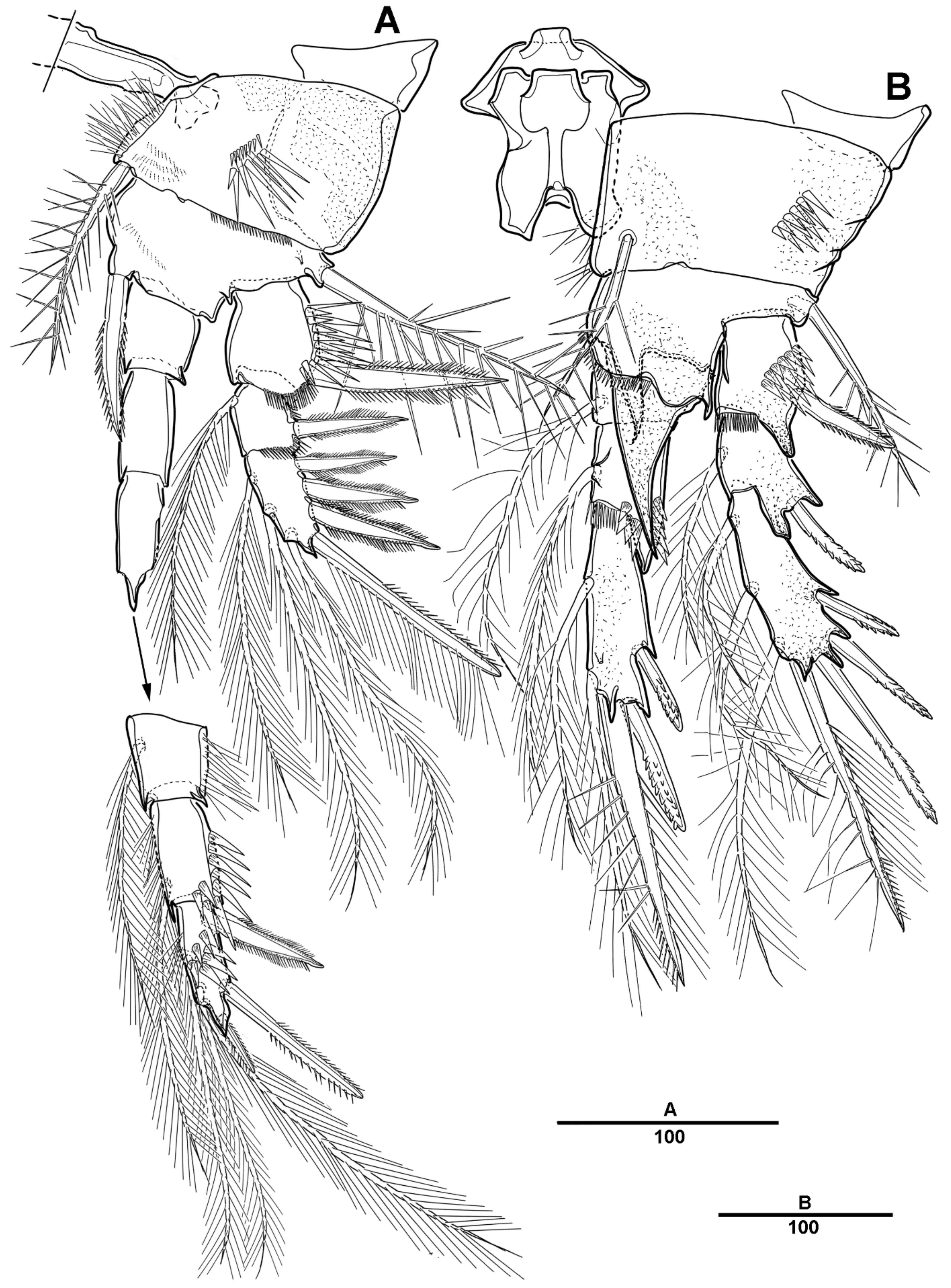
P1 (Figure 6A). Coxa large, with spinules and row of long setules on the anterior surface, and with pinnate inner seta. Basis with one pinnate long outer seta and one strong pinnate inner spine. First segment of exopod with one outer spine, and with spinules and setules as figured; exp-2 with one outer strong pinnate spine and one plumose long inner seta, and with tiny spinules at the distal margins; exp-3 slightly longer than exp-2, with three outer pinnate spines, two distal setae, and two plumose inner setae. Endopod longer than exopod; enp-1 with inner plumose seta, and long setules along the outer margin; enp-2 with one long inner plumose seta, and with strong spinules on distal and the outer margins; enp-3 with three long plumose inner setae, one small apical spine, and two pinnate outer spines.
P2 (Figure 6B–D). Coxa large, with dense patch of strong spinules near the outer margin, and with inner pinnate spine. Basis with one pinnate outer seta, with small spinules at base of endopod, and with small bent process at inner distal corner (Figure 6D). Exp-1 with one bipinnate outer spine, with strong spinules on the outer margin; exp-2 with one serrate outer spine and one plumose inner seta; exp-3 with four plumose setae and three serrate spines. Endopod slightly longer than exopod; enp-1 small, with plumose inner seta, and the anterior surface formed a large apophysis to reach the distal edge of enp-2; enp-2 with one plumose inner seta, and strong spinules along the outer distal margin; enp-3 with three plumose setae and two serrate spines.
P3 (Figure 7A,B). Coxa large, with one pinnate inner seta, with several strong spinules on the anterior surface. Basis with one pinnate outer seta, with minute spinules close to joint with endopod; the posterior surface with spinous processes (Figure 7B). Exp-1 with one serrate outer spine and strong spinules on the distal and outer margins; exp-2 with one serrate outer spine and one long plumose inner seta; exp-3 with two plumose setae and three serrate spines. Endopod much longer than exopod; enp-1 and enp-2 each with one inner plumose seta and strong spinules as figured; enp-3 with two long pinnate spines terminally, and two outer spines.
P4 (Figure 7C). Coxa with row of spinules, and with one small inner seta. Basis with long pinnate outer seta. Exp-1 with one pinnate outer seta, and with strong spinules; exp-2 with one outer spine and one long pinnate inner seta, setules on the distal margin; exp-3 with one inner pinnate spine, small pinnate seta inner side and long seta outer side at terminal, and very long pinnate outer seta. Endopod much longer than exopod; enp-1 shortest, with one very long inner spinulose seta; enp-2 unarmed; enp-3 with two long pinnate distal setae, and two outer spines.
Armature formula for swimming legs:
Exopod Endopod
P1 0.1.223 1.1.312
P2 0.1.322 1.1.221
P3 0.1.122 1.1.022
P4 0.1.112 1.0.022
P5 (Figure 2D) incorporated into somite, and the setae reduced to four; the outermost pinnate, and separated from others.
Figure 6.
Scottolana daecheonensis sp. nov. holotype female: (A) P1, anterior; (B) P2, anterior (inset showing dense patch of strong spinules); (C) P2 basis, posterior; (D) P2, basal inner process. Scale bars in μm.
Figure 6.
Scottolana daecheonensis sp. nov. holotype female: (A) P1, anterior; (B) P2, anterior (inset showing dense patch of strong spinules); (C) P2 basis, posterior; (D) P2, basal inner process. Scale bars in μm.

Figure 7.
Scottolana daecheonensis sp. nov. holotype female: (A) P3, anterior; (B) P3, basal inner process; (C) P4, anterior. Scale bars in μm.
Figure 7.
Scottolana daecheonensis sp. nov. holotype female: (A) P3, anterior; (B) P3, basal inner process; (C) P4, anterior. Scale bars in μm.

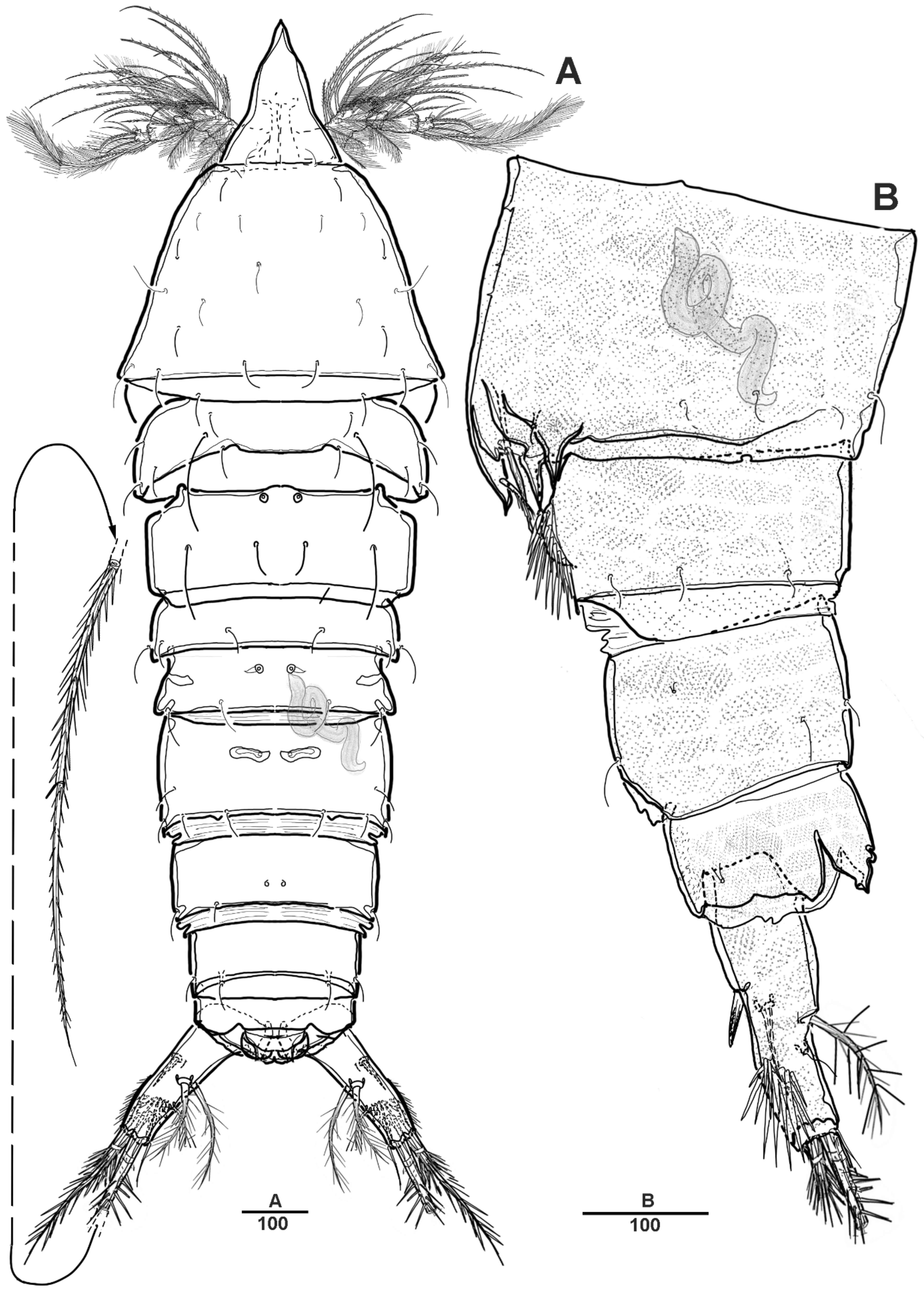
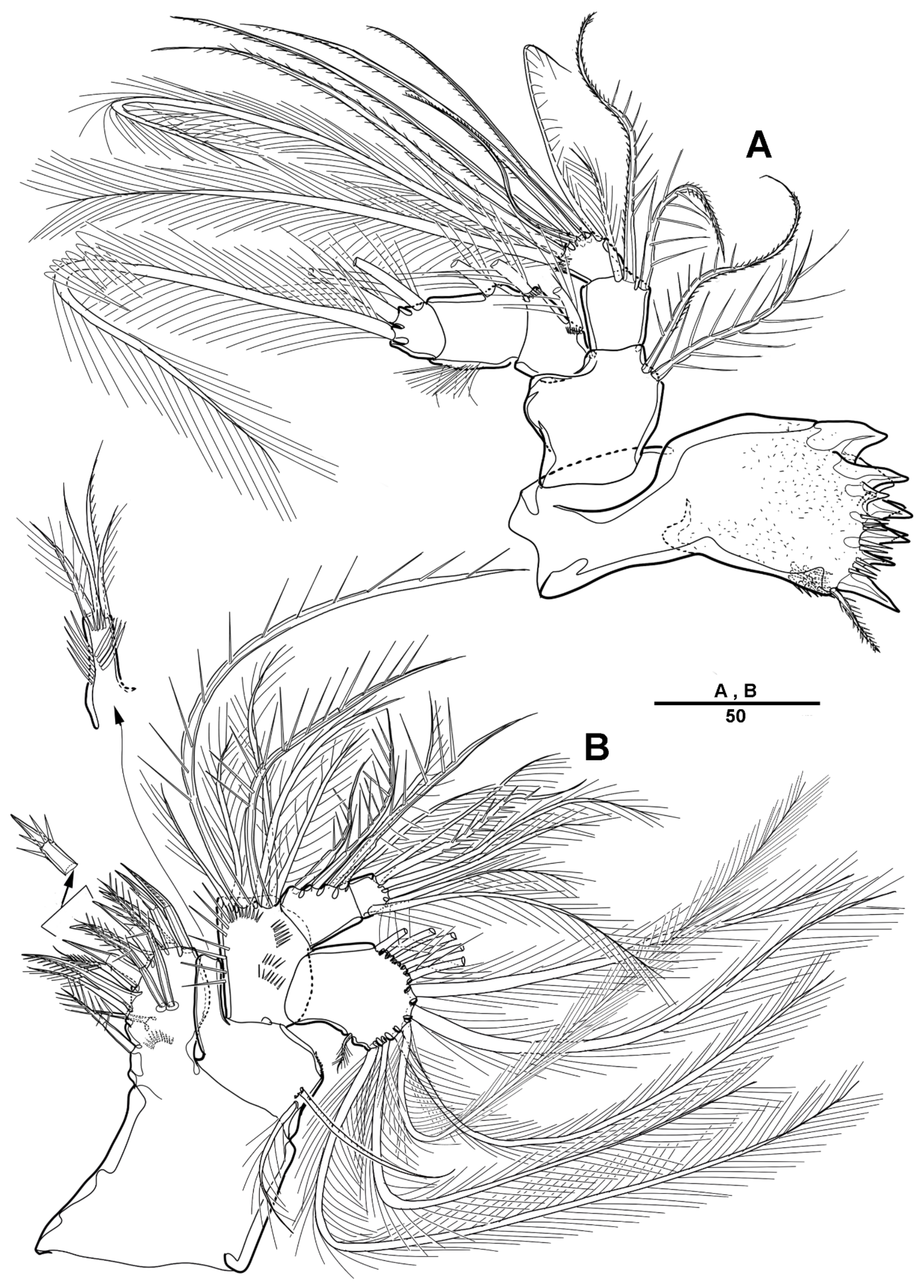
Male—habitus (Figure 8A). Total body length 1510 mm (N = 3; range: 1245–1593 mm). Sexual dimorphism in antennule, P4, P5, urosome and caudal ramus.
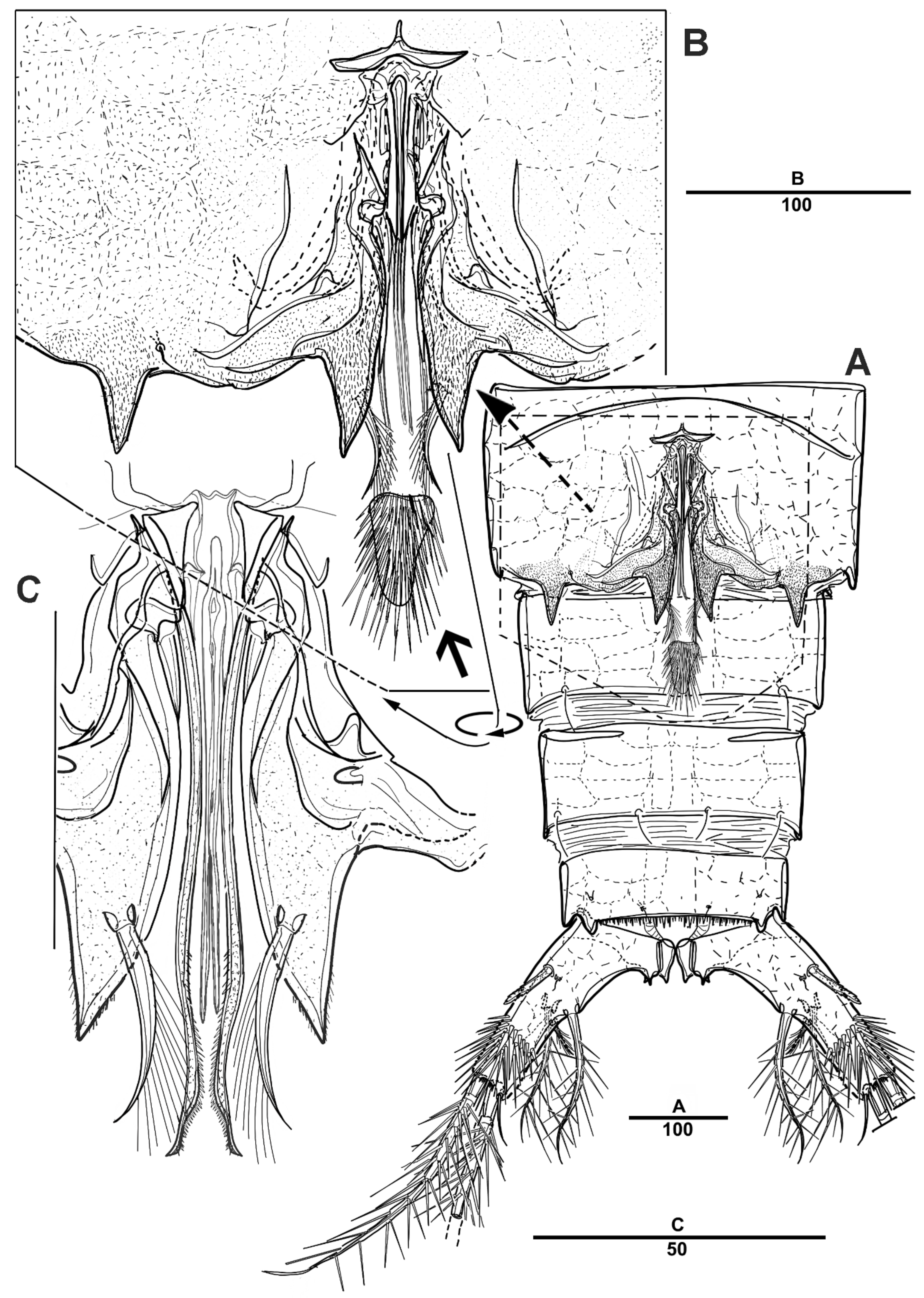
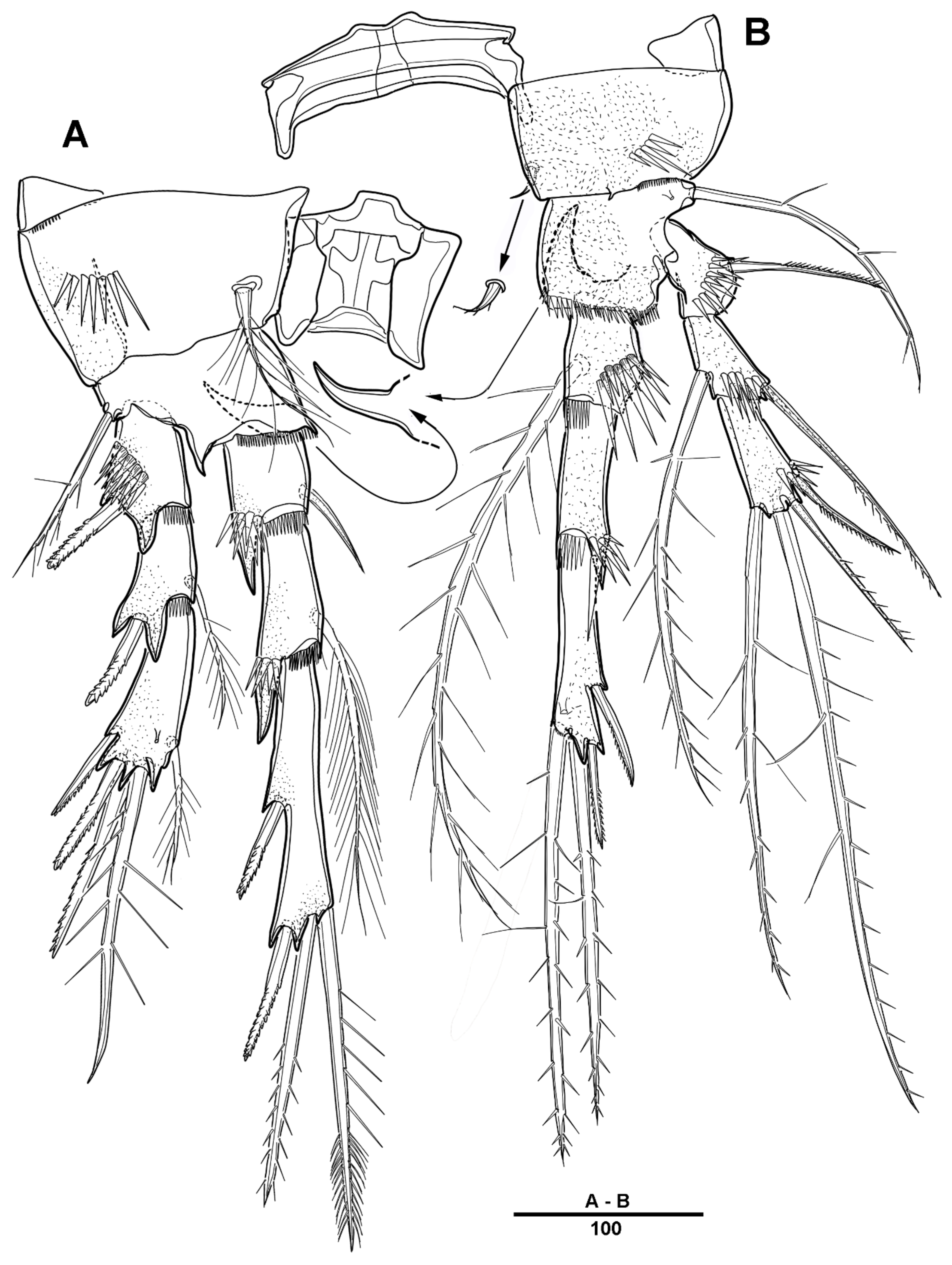
Antennule (Figure 9) 5-segmented. Segment 1 very large, with transverse surface sutures, with four bi-articulate plumose setae, twelve pinnate setae, and two aesthetascs (Figure 9A). Segment 2 short, with two pinnate setae (Figure 9B). Segment 3 small with three pinnate setae. Segment 4 with a blunt molar-like process on the distal three-quarters, with long distal plumose seta, six additional setae, and two modified elements near the outer margin (shown by an arrow in Figure 9C). Segment 5 small, with one pebble-like element and tiny seta anterior at the distal margin, and with one pinnate and four smooth setae (Figure 9D).
P4 (Figure 10A–C) as in female, except for a crumpled inner margin of enp-2, with strong spinules.
P5 (Figure 10D) with armature as in female, but outer basal seta longer and plumose, and a sensilla located near basal seta.
Urosome (Figure 8B and Figure 11A) 5-segmented, consisting of P5-bearing somite (omitted in Figure 11A), genital somite, and three free urosomites. Genital somite (Figure 11) with four spinous extensions covered by tiny spinules, with a long element near the proximal margin that reaches the edge of the first free urosomite, and with small distal setules. First free urosomite with patch of dense spinules on the ventral surface (shown by an arrow in Figure 11B). Genital pores closed off by the modified sixth legs (Figure 11B,C); P6 fused to genital somite, and forming a large triangular process with one pinnate seta; additional very long element arising from near the inner proximal margin of P6, articulated at base, and distally one-third denticulated.
Caudal rami (Figure 11A) as in female except that seta VII small and pinnate.
3.2. Scottolana Wonchoeli sp. nov.
Figure 12, Figure 13, Figure 14, Figure 15, Figure 16, Figure 17, Figure 18, Figure 19, Figure 20 and Figure 21 are listed bellow.
Type locality—subtidal zone near Ulleung-do Island, Korea (37°29′13″ N 130°55′20″ E).
Material examined—Holotype 1♀ (MABIK CR00252817) dissected on eleven slides; Paratype 1♂ (MABIK CR00252818) dissected on seven slides.
Etymology—the species is named in honor of Professor Wonchoel Lee, (Hanyang University, Korea), in recognition of his excellent contributions to the systematics of copepods.
Genbank accession number—mitochondrial cytochrome c oxidase subunit I gene (OP452899); 18S ribonucleic acid gene (OP454039, OP454040).
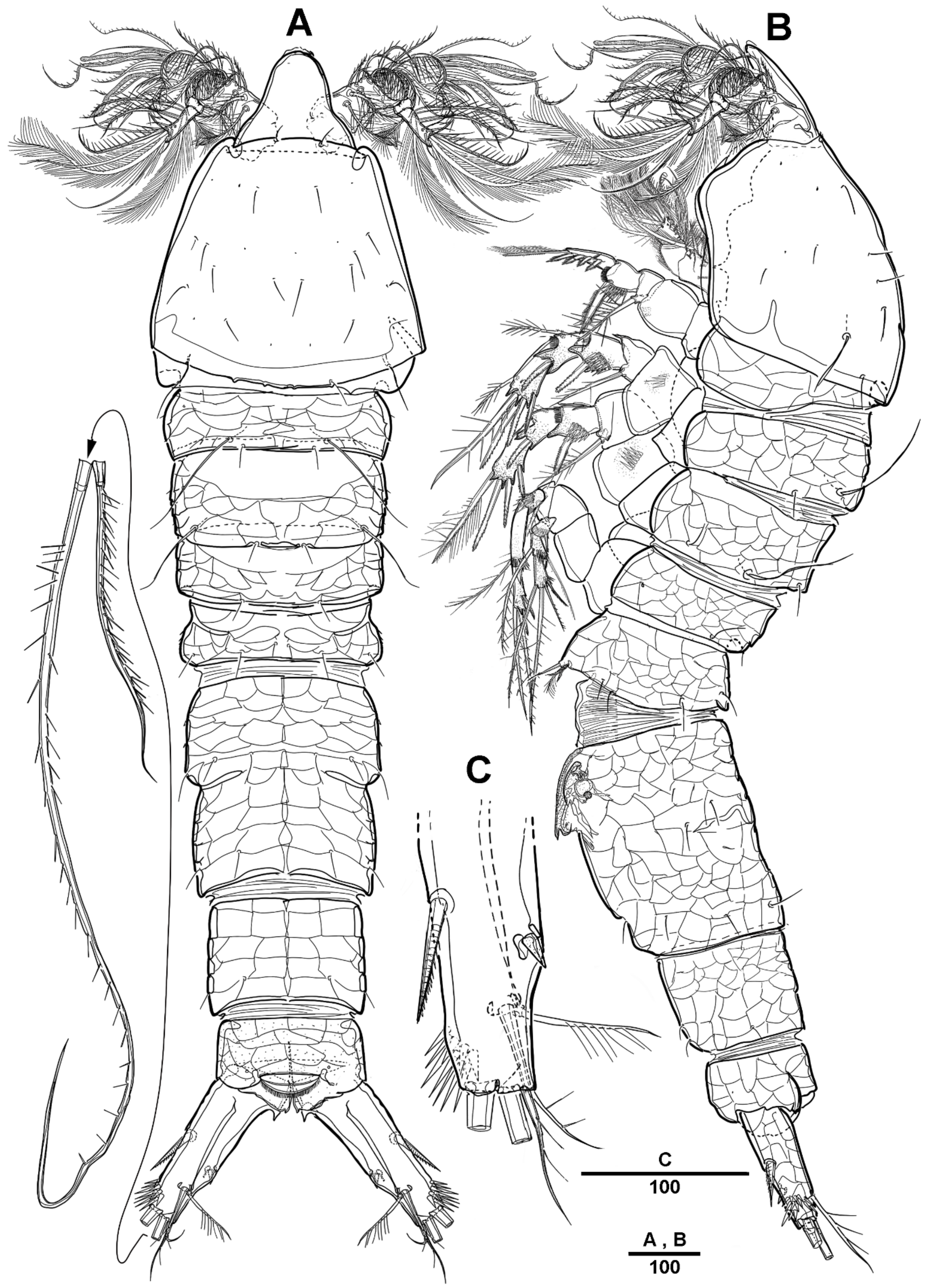
Female—habitus (Figure 12A,B). Total body length 1820 µm, measured from anterior margin of rostrum to the posterior margin of caudal rami. Largest width measured at the posterior margin of cephalic shield: 410 µm. Body large, robust, semi-cylindrical, and the posterior margin with smooth hyaline frill. Prosome slightly compressed laterally.
Cephalic shield gradually widens posteriorly, with long surface sensilla. Rostrum (Figure 12A and Figure 19B) large, defined at base, with two lateral sensilla, anterior margin with slightly pronounced ornamentation. Cephalosome separated from the first pedigerous somite. All prosomites and urosomites surface with distinct reticulation, and pedigerous somites with pores and sensilla as illustrated. Urosome (Figure 13A) 4-segmented, consisting of P5 bearing somite, genital double-somite, and two free urosomites. Genital double-somite completely fused; original segmentation is marked by internal chitinized ridges.
Genital field (Figure 13B,C), copulatory pores paired, and covered by epicopulatory flaps. Gonopores paired, anteriorly displaced, closed off by genital operculum derived from vestigial sixth legs, P6 with one long smooth seta and internal projections. Anal operculum (Figure 13D) weakly developed and with a spinular row along the distal margin.
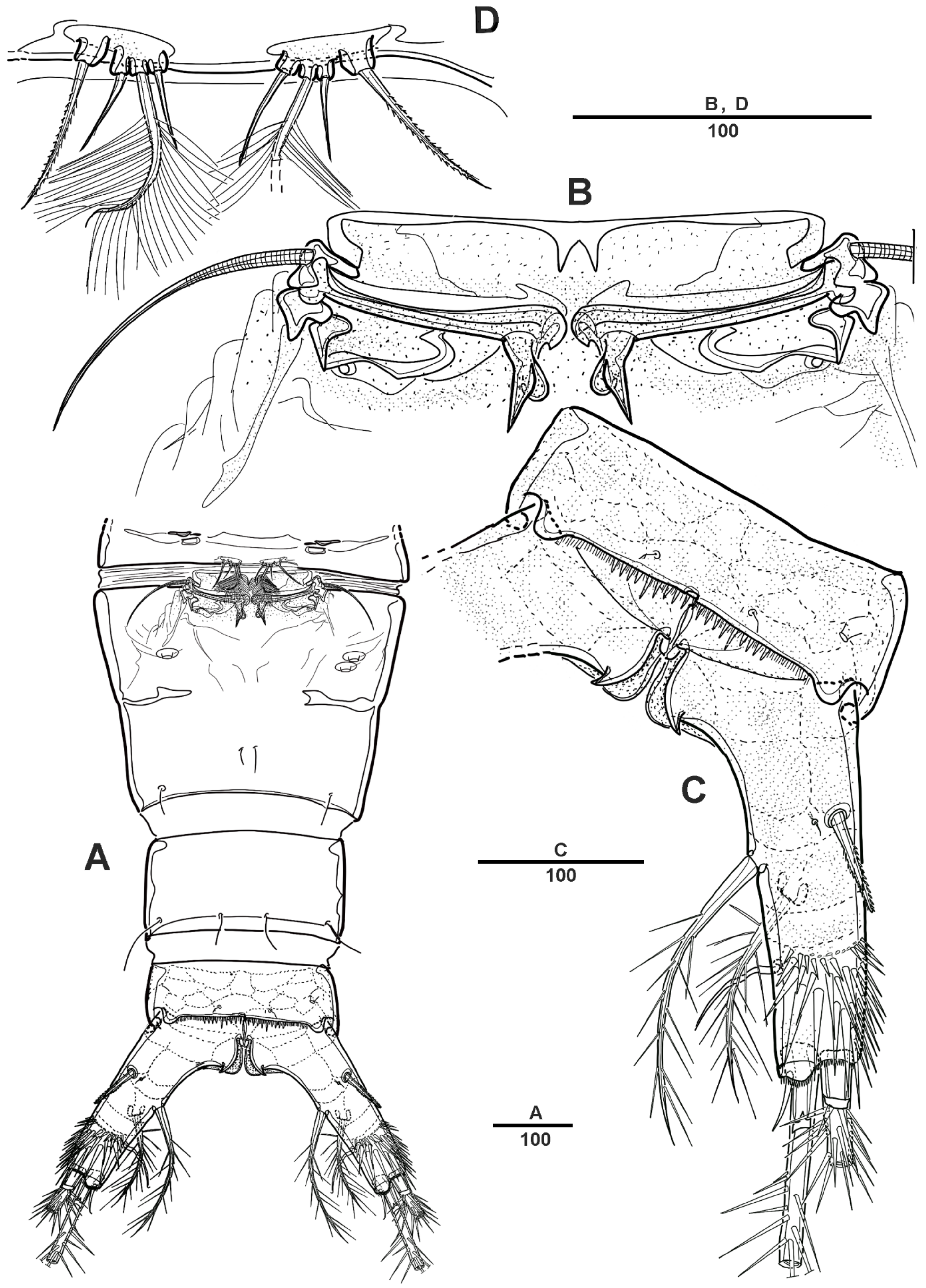
Figure 8.
Scottolana daecheonensis sp. nov. paratype male: (A) habitus, dorsal; (B) urosome, lateral. Scale bars in μm.
Figure 8.
Scottolana daecheonensis sp. nov. paratype male: (A) habitus, dorsal; (B) urosome, lateral. Scale bars in μm.

Figure 9.
Scottolana daecheonensis sp. nov. paratype male: (A) antennule, dorsal; (B) antennulary segment 2 and 3, dorsal; (C) antennulary segment 4 and 5, anterior (modified elements arrowed; armature of segment 5 omitted); (D) antennulary segment 5, anterior. Scale bars in μm.
Figure 9.
Scottolana daecheonensis sp. nov. paratype male: (A) antennule, dorsal; (B) antennulary segment 2 and 3, dorsal; (C) antennulary segment 4 and 5, anterior (modified elements arrowed; armature of segment 5 omitted); (D) antennulary segment 5, anterior. Scale bars in μm.

Figure 10.
Scottolana daecheonensis sp. nov. paratype male: (A) P4, anterior; (B) P4 enp-2, anterior; (C) P4 enp-2, posterior; (D) P5. Scale bars in μm.
Figure 10.
Scottolana daecheonensis sp. nov. paratype male: (A) P4, anterior; (B) P4 enp-2, anterior; (C) P4 enp-2, posterior; (D) P5. Scale bars in μm.

Figure 11.
Scottolana daecheonensis sp. nov. paratype male: (A) urosome, ventral; (B) genital field, ventral (patch of dense spinules arrowed); (C) detail of P6, dorsal. Scale bars in μm.
Figure 11.
Scottolana daecheonensis sp. nov. paratype male: (A) urosome, ventral; (B) genital field, ventral (patch of dense spinules arrowed); (C) detail of P6, dorsal. Scale bars in μm.

Figure 12.
Scottolana wonchoeli sp. nov. holotype female: (A) habitus, dorsal; (B) habitus, lateral; (C) right caudal ramus, ventral. Scale bars in μm.
Figure 12.
Scottolana wonchoeli sp. nov. holotype female: (A) habitus, dorsal; (B) habitus, lateral; (C) right caudal ramus, ventral. Scale bars in μm.

Figure 13.
Scottolana wonchoeli sp. nov. holotype female: (A) urosome, ventral; (B) genital field, ventral; (C) P5 and genital field, lateral; (D) anal somite and left caudal ramus, dorsal; (E) P5. Scale bars in μm.
Figure 13.
Scottolana wonchoeli sp. nov. holotype female: (A) urosome, ventral; (B) genital field, ventral; (C) P5 and genital field, lateral; (D) anal somite and left caudal ramus, dorsal; (E) P5. Scale bars in μm.

Caudal rami (Figure 12C and Figure 13D) about 2.6 times longer than the width at base, with a hook-like projection on the inner margins, and with spinular patch around the outer distal corner. Each ramus armed with 7 setae: seta I short and strong, pinnate spiniform; seta II small conical; seta III pinnate; seta IV pinnate and long; seta V longest, about three times longer than seta IV; seta VI slender and pinnate; seta VII plumose, and articulate at base.
Antennule (Figure 14A–D) 3-segmented. Segment 1 largest, with three incomplete sutures around the posterior margin, with five naked, seventeen pinnate, three bi-articulated plumose setae, and two aesthetascs; segment 2 short, with one bare and three pinnate setae; and last segment with six naked, three pinnate, three long plumose, and one short bi-articulated plumose seta posteriorly. Armature as follows: 1-[25 + 2ae], 2-[4], 3-[13].
Antenna (Figure 14E,F) biramous; coxo-basis short with long setules on the anterior surface. Exopod 8-segmented; first to fourth and seventh segments each with one plumose seta each; fifth segment with pinnate seta; last segment with one plumose and three unipinnate setae. Endopod 2-segmented; enp-1 with one pinnate and one small smooth setae; enp-2 with incomplete suture, with four pinnate setae laterally, and six pinnate and one bare setae apically.
Mandible (Figure 15A) with well-developed gnathobase bearing six strong teeth, with an additional row of spinules at the dorsal margin, and with the dorsal corner with one pinnate spine. Basis with two pinnate inner setae. Palp well-developed, biramous. Exopod 3-segmented, all setae plumose; exp-1 with two long setae; exp-2 with row of spinules at the outer margin, with one long seta; exp-3 with one proximal, and two apical setae. Endopod 2-segmented; enp-1 with two pinnate and one plumose setae; enp-2 with five pinnate and three plumose setae.
Maxillule (Figure 15B). Praecoxal arthrite with eight spines and two setae around the distal margin, and two setae on the anterior surface, with one row of spinules on the posterior surface. Coxal epipodite represented by one plumose and one bare setae; endite with four setae, and the anterior surface with two rows of spinules. Basis with closely set endites, with row of spinules; proximal endite with three setae, and distal endite with four setae. Exopod l-segmented, all setae plumose, with six inner setae, five distal setae, and one tiny outer seta. Endopod 2-segmented; enp-1 with five setae; enp-2 with six plumose setae.
Maxilla (Figure 16A) comprising praecoxa, coxa, allobasis and 3-segmented endopod. Praecoxa with row of spinules near base of endites and distal outer corner, with two endites; proximal endite with three pinnate and one bare setae; distal endite with two pinnate setae. Coxa with two endites, with three pinnate elements each. Allobasis endite transformed into a strong curved claw; four pinnate setae, and with two pinnate surface setae. Endopod 3-segmented; enp-1 with three pinnate setae; enp-2 with one spinulose and two naked elements; enp-3 with three naked setae.
Maxilliped (Figure 16B,D) phyllopodial. Syncoxa with rows of small spinules on the surface, with nine elements on the inner margin. Basis with two pinnate and one plumose setae on the inner margin. Endopod with seven pinnate and four plumose setae.
Swimming legs 1–4 (Figure 17 and Figure 18) with well-developed intercoxal sclerites and praecoxae, both without ornamentation. Coxae and bases with surface spinular pattern as illustrated. All swimming legs with 3-segmented exopods and endopods.
P1 (Figure 17A). Coxa large, with spinules and rows of setules on the anterior surface, and with tri-pinnate inner seta. Basis with strong pinnate outer seta and one bipinnate inner spine. Exp-1 with strong pinnate outer spine, and with spinules and setules on outer and the distal margins; exp-2 with one outer pinnate spine and one long plumose inner seta, and with tiny spinules at the outer and terminal margins; exp-3 with three outer pinnate spines, one distal spine, and three plumose inner setae. Endopod longer than exopod; enp-1 with plumose inner seta, and long setules along the outer margin; enp-2 with one inner plumose seta, and with strong spinules at distal and the outer margins; enp-3 with three long plumose inner setae, one small apical spine, and two pinnate outer spines, and with strong spinules on the anterior surface.
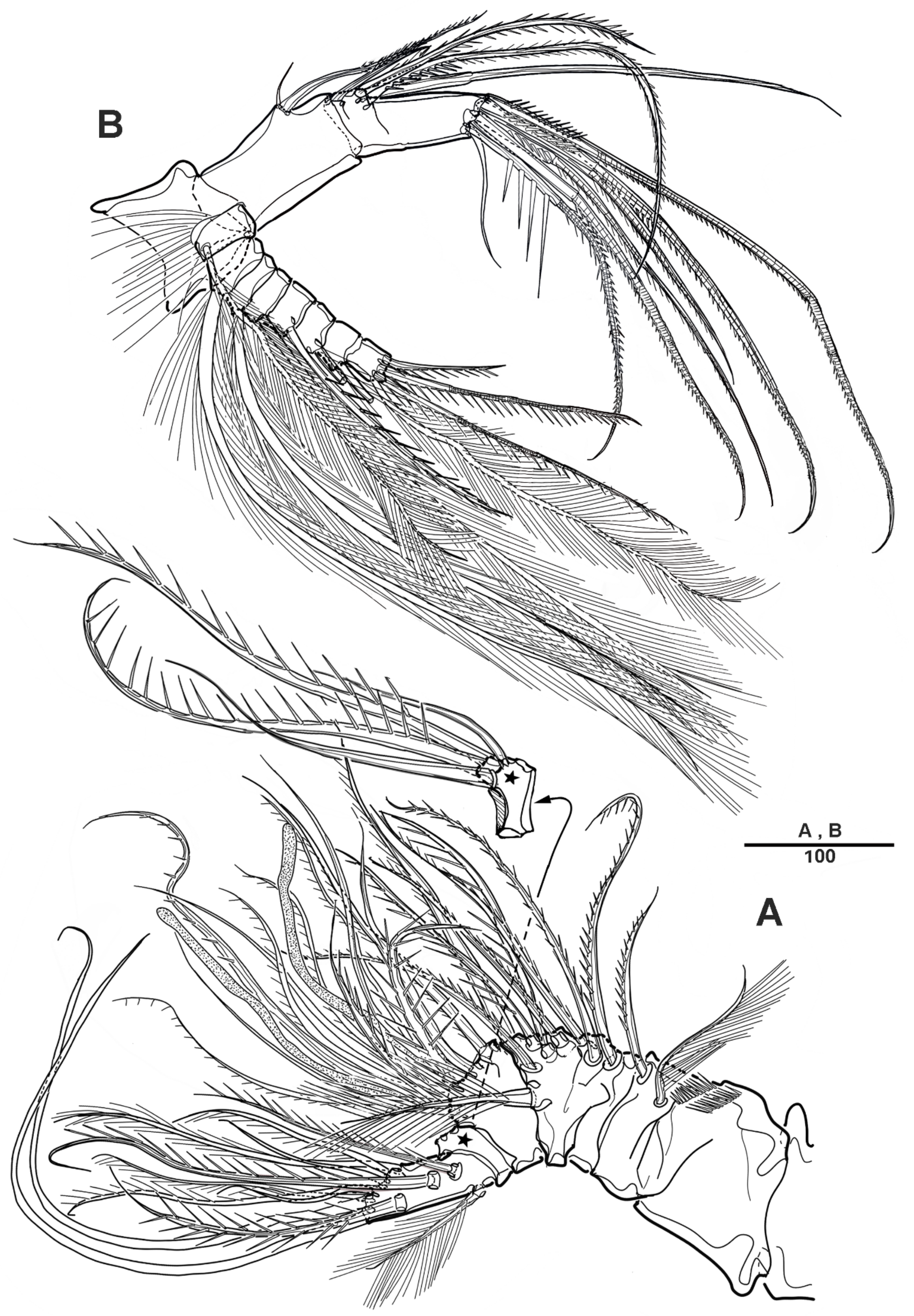
Figure 14.
Scottolana wonchoeli sp. nov. holotype female: (A) antennule; (B) distal part of first antennulary segment; (C) antennulary segment 2; (D) antennulary segment 3; (E) antenna; (F) distal antennary exopod. Scale bars in μm.
Figure 14.
Scottolana wonchoeli sp. nov. holotype female: (A) antennule; (B) distal part of first antennulary segment; (C) antennulary segment 2; (D) antennulary segment 3; (E) antenna; (F) distal antennary exopod. Scale bars in μm.

Figure 15.
Scottolana wonchoeli sp. nov. holotype female: (A) mandible; (B) maxillule. Scale bars in μm.
Figure 15.
Scottolana wonchoeli sp. nov. holotype female: (A) mandible; (B) maxillule. Scale bars in μm.

Figure 16.
Scottolana wonchoeli sp. nov. holotype female: (A) maxilla (asterisk indicated distal coxal endite); (B) maxilliped; (C) maxillipedal basis; (D) maxillipedal endopod. Scale bars in μm.
Figure 16.
Scottolana wonchoeli sp. nov. holotype female: (A) maxilla (asterisk indicated distal coxal endite); (B) maxilliped; (C) maxillipedal basis; (D) maxillipedal endopod. Scale bars in μm.

Figure 17.
Scottolana wonchoeli sp. nov. holotype female: (A) P1, anterior; (B) P2, anterior. Scale bars in μm.
Figure 17.
Scottolana wonchoeli sp. nov. holotype female: (A) P1, anterior; (B) P2, anterior. Scale bars in μm.

P2 (Figure 17B). Coxa large, rectangular, with inner pinnate seta, and with spinules near the outer margin. Basis with one pinnate outer seta, with small spinules along the inner distal margin, and a posterior surface formed large spinous process, extending beyond end of enp-1. Exp-1 with one bipinnate outer spine, with strong spinules at base of outer spine; exp-2 with one plumose inner seta and one pectinate outer spine; exp-3 with three plumose setae, one pinnate spine, and three serrate spines. Endopod not significantly longer than exopod; enp-1 small, with plumose inner seta, and an anterior surface produced into a large apophysis; enp-2 with one plumose inner seta, and strong spinules along the outer distal margin; enp-3 with two plumose setae, one strong pinnate spine, and two serrate spines.
P3 (Figure 18A). Coxa large, with one plumose inner seta, with row of spinules on the anterior surface. Basis with one pinnate outer seta, with tiny spinules near base of endopod; posterior surface with inner spinous process. Exp-1 with one outer spine and strong spinules on the outer margin; exp-2 with one small plumose inner seta, and one outer spine; exp-3 with one small plumose inner seta, one pinnate distal seta, and three serrate spines. Endopod longer than exopod; enp-1 with small naked seta, and with strong spinules; enp-2 with one inner plumose seta and strong spinules as figured; enp-3 with two pinnate distal spines, and with two outer spines.
P4 (Figure 18B). Coxa with one small inner seta, and with row of spinules. Basis with long pinnate outer seta, posterior surface with curved upward inner spinous process. Exp-1 with one pinnate outer seta, and with strong spinules; exp-2 with long pinnate inner seta, and with an outer pinnate spine; exp-3 with one pinnate inner seta, one long bipinnate and one short unipinnate setae distally, and one small pinnate outer seta. Endopod much longer than entire exopod; enp-1 short, with one long inner pinnate seta, and the outer margin with strong spinules; enp-2 unarmed; enp-3 with two long pinnate inner setae, and two outer spines.
Armature formula for swimming legs:
Exopod Endopod
P1 0.1.313 1.1.312
P2 0.1.322 1.1.221
P3 0.1.122 1.1.022
P4 0.1.121 1.0.022
P5 (Figure 13E) incorporated into fifth pedigerous somite, represented by four setae; the outermost seta (basal seta) is separated from others.
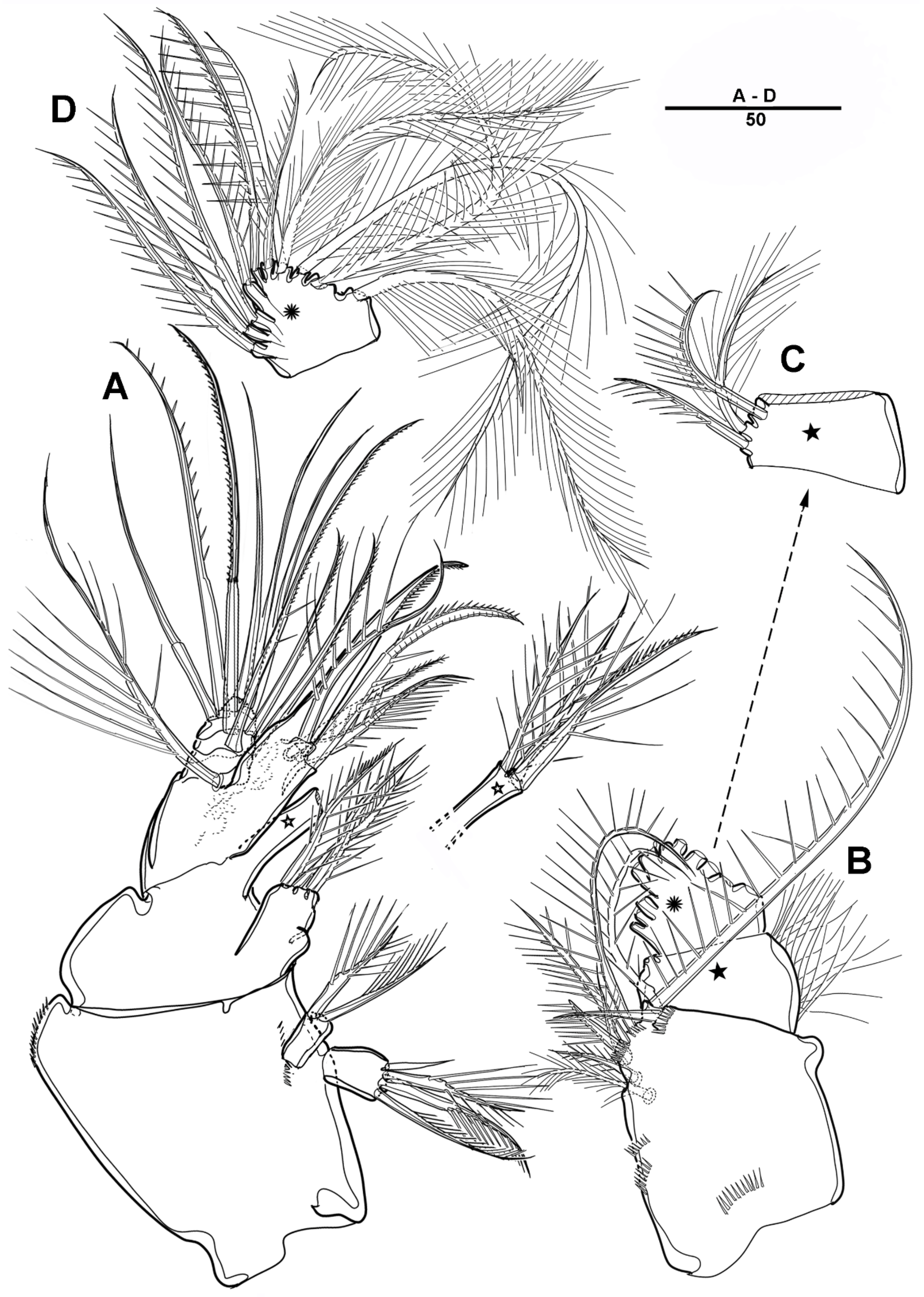
Male—habitus (Figure 19A). Total body length 1540 mm. Sexual dimorphism in antennule, P3, P5, urosome and caudal ramus.
Antennule (Figure 20) 5-segmented. Segment 1 large, with transverse surface sutures; with two aesthetascs, each fused basally to long smooth seta; with three bi-articulate pinnate, nine pinnate, one plumose, and five geniculated setae. Segment 2 short, with one pinnate and one plumose setae. Segment 3 small, with three pinnate setae and one bi-articulated plumose seta. Segment 4 large; with very long plumose and three long pinnate setae in proximal half; with modified element on the anterior surface; and with one bipinnate and two smooth setae near the distal margin. Segment 5 small, with two pinnate and five naked setae (two bi-articulate at base).
P3 enp-3 (Figure 21A) with large modified pore along the inner margin, and with pectinate outer distal spine.
P5 (Figure 21B) with armature as in female, but innermost seta smooth.
Urosome (Figure 19C) comprising P5-bearing somite, genital somite and three abdominal somites. Genital somite (Figure 21C) with paired lateral spinous processes, and with mid-ventral sabre-like process bearing fine setules. Genital pores closed off by the modified sixth legs. First free urosomite with patch of dense spinules on ventral surface (arrowed in Figure 21C). P6 fused to genital somite and formed into a large process with pinnate seta; additional long element arising from near the inner proximal margin of P6, defined at base, covered by tiny spinules.
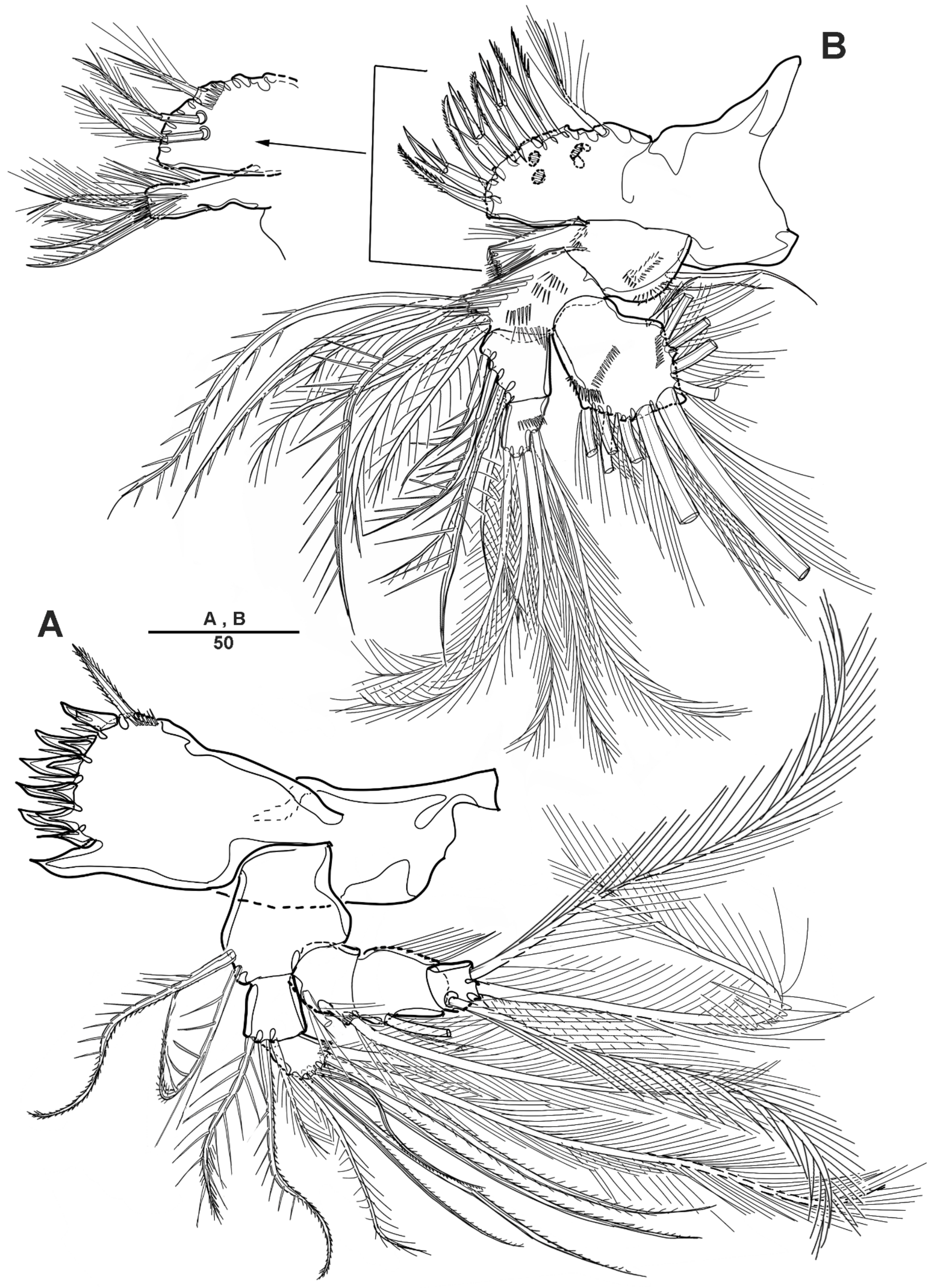
Figure 18.
Scottolana wonchoeli sp. nov. holotype female: (A) P3, anterior; (B) P4, anterior. Scale bars in μm.
Figure 18.
Scottolana wonchoeli sp. nov. holotype female: (A) P3, anterior; (B) P4, anterior. Scale bars in μm.

Figure 19.
Scottolana wonchoeli sp. nov. paratype male: (A) habitus, dorsal; (B) rostrum, dorsal; (C) urosome, ventral. Scale bars in μm.
Figure 19.
Scottolana wonchoeli sp. nov. paratype male: (A) habitus, dorsal; (B) rostrum, dorsal; (C) urosome, ventral. Scale bars in μm.

Figure 20.
Scottolana wonchoeli sp. nov. paratype male: (A) antennule, dorsal (asterisk showing armature on proximal half of segment 4); (B) antennulary segment 2–4, anterior; (C) antennulary segment 4, anterior (armature of segment 5 omitted); (D) antennulary segment 5, anterior. Scale bars in μm.
Figure 20.
Scottolana wonchoeli sp. nov. paratype male: (A) antennule, dorsal (asterisk showing armature on proximal half of segment 4); (B) antennulary segment 2–4, anterior; (C) antennulary segment 4, anterior (armature of segment 5 omitted); (D) antennulary segment 5, anterior. Scale bars in μm.

Figure 21.
Scottolana wonchoeli sp. nov. paratype male: (A) P3 enp-3, anterior (modified pore arrowed); (B) P5; (C) genital somite, ventral (patch of dense spinules arrowed); (D) anal somite and right caudal ramus, dorsal. Scale bars in μm.
Figure 21.
Scottolana wonchoeli sp. nov. paratype male: (A) P3 enp-3, anterior (modified pore arrowed); (B) P5; (C) genital somite, ventral (patch of dense spinules arrowed); (D) anal somite and right caudal ramus, dorsal. Scale bars in μm.

Caudal rami (Figure 21D) as in female except seta II not modified, being long and pinnate, and seta VII short and smooth.
3.3. DNA Sequences in the Family Canuellidae
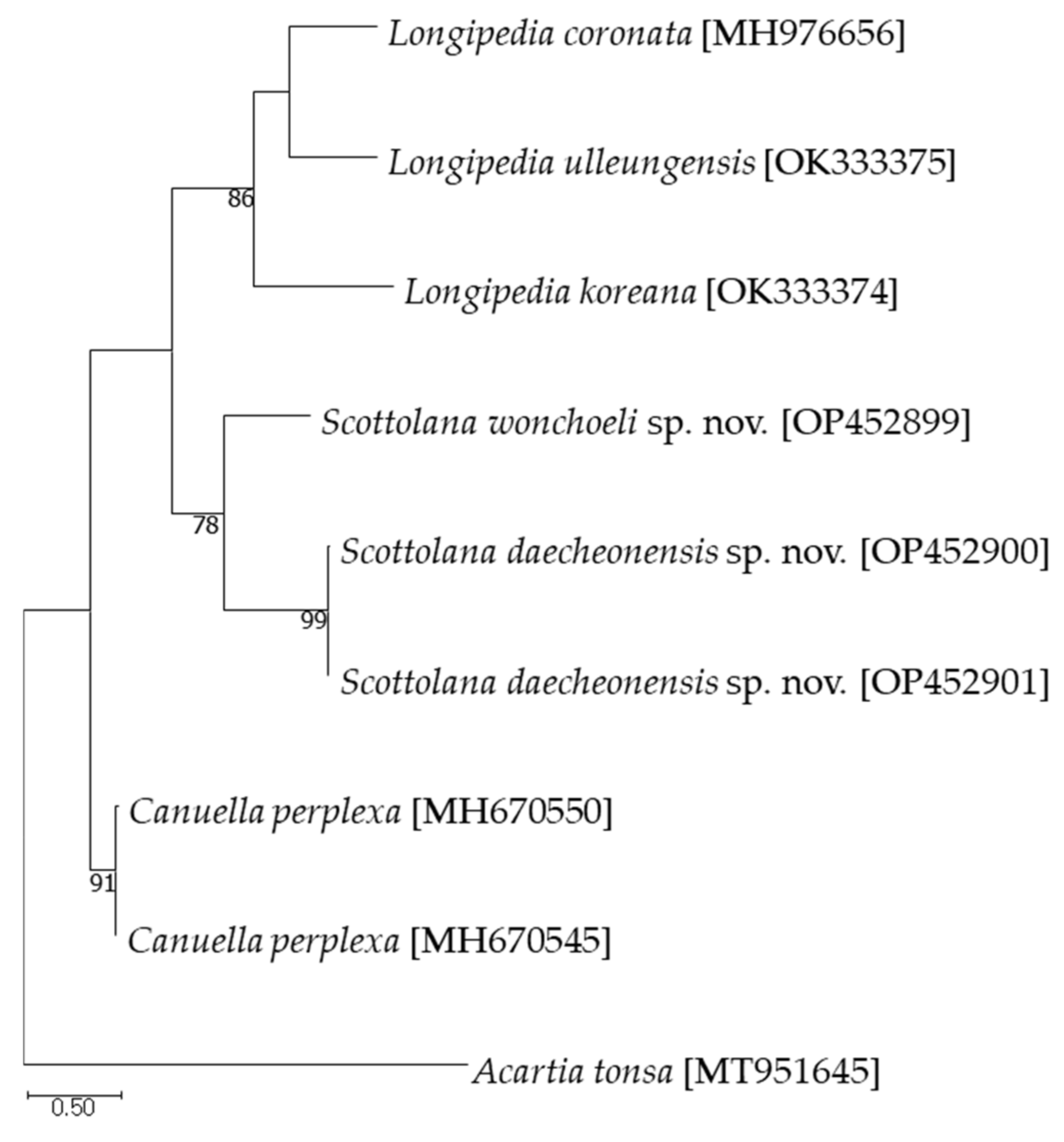
The partial mitochondrial cytochrome c oxidase subunit I (COI) and the 18S ribosomal RNA (18S rRNA) sequences were successfully obtained from two new species. In addition, the average pairwise distance was calculated by adding two sequence of the Canuella perplexa Scott T. and Scott A., 1893 found in NCBI. As a result of calculating the average pairwise distance, with nine COI sequences and six 18S rRNA sequences (Tables S1 and S2), the maximum intraspecific distance for COI was 0.917% in S. daecheonensis sp. nov. and the pairwise distance between the two new species was 20.031%. In the case of the 18S rRNA sequences, the intra-species distances were close to 0%, the maximum interspecific distance was 3.214% in Scottolana, and the maximum distances was 8.565% between Scottolana and Canuella. The maximum-likelihood phylogeny is shown in Figure 22.
4. Discussion
Por [28] established the genus Scottolana Por, 1967, in the family Canuellidae for 3 species, which were originally placed in Sunaristes Hesse, 1867: Sunaristes inopinata Thompson and Scott, 1903, Sunaristes longipes Thompson and Scott, 1903, and Sunaristes curticaudata Thompson and Scott, 1903, but unfortunately, he neglected to fix a type species, consequently rendering the generic name unavailable. Huys [29] re-established the generic name Scottolana Huys, 2009, and formally designated Scottolana geei Mu and Huys, 2004 as the type species of the genus. He also recognized 14 valid species within the genus Scottolana; S. antillensis, S. brevifurca, S. bulbifera, S. bulbosa, S. dissimilis, S. glabra, S. geei, S. inopinata, S. longipes, S. oleosa, S. rostrata, S. scotti, S. tumidiseta, and S. uxoris. After that, 3 species, S. gomezi [18], S. huysi, and S. jasani [10], were added to this genus. Currently, the genus contains 19 valid species, including two new species described in this study, S. daecheonensis sp. nov. and S. wonchoeli sp. nov.
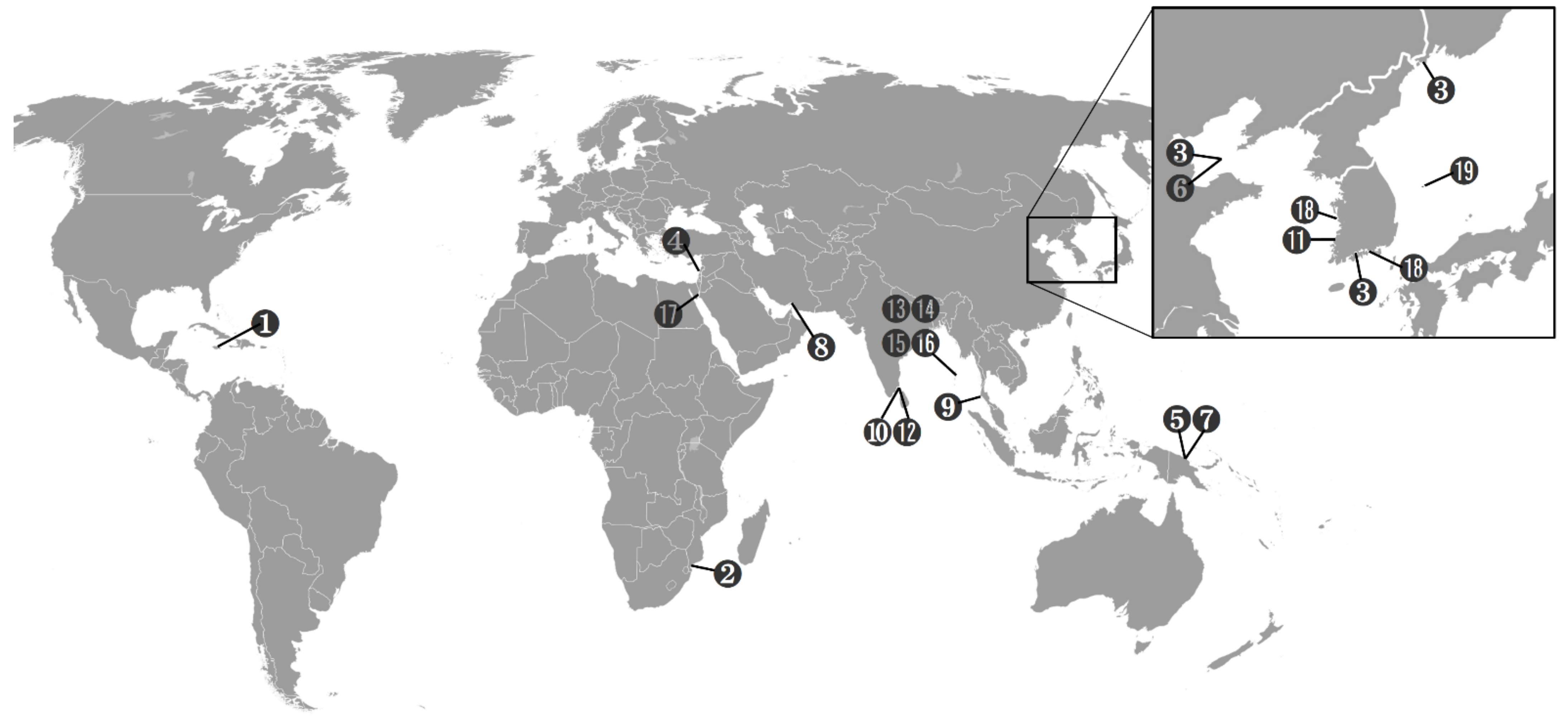
The genus Scottolana is widespread in tropical and subtropical waters (Figure 23), and is mainly found in soft sediments, but also in rockpools, macroalgae, and invertebrates. Scottolana is distributed worldwide, but has never been reported in the central and eastern Pacific Ocean and the Atlantic Ocean (except for S. antillensis, from the coast of Jamaica). In this genus, all species except two have been reported only in type locality. Scottolana bulbifera has been found around the Korean Peninsula; the Posyet Bay, Russia (type locality) [7]; the Bohai Sea, China [9]; and the South Sea of Korea [19]. S. longipes was first reported in the Gulf of Mannar in Sri Lanka (type locality) [11], and was found on the Mediterranean coast of Israel [16], Inhaca Island in Mozambique [13], and the Andaman Islands [14].
Scottolana daecheonensis sp. nov. and S. wonchoeli sp. nov. are assigned to the genus Scottolana Huys, 2009, in the family Canuellidae Lang, 1944, according to the following combination of morphological characters: P1-bearing somite not fused to cephalosome, antennary exopod 8-segmented, anal operculum weakly developed, P1–P4 with 3-segmented rami, P1–P4 exp-3 with 7, 7, 5, 4, and enp-3 with 6, 5, 4, 4 elements each [29,30].
Figure 23.
Distribution of Scottolana species. (1) S. antillensis; (2) S. brevifurca; (3) S. bulbifera; (4) S. bulbosa; (5) S. dissimilis; (6) S. geei; (7) S. glabra; (8) S. gomezi; (9) S. huysi; (10) S. inopinata; (11) S. jasani; (12) S. longipes; (13) S. oleosa; (14) S. rostrata; (15) S. scotti; (16) S. tumidiseta; (17) S. uxoris; (18) S. daecheonensis sp. nov.; (19) S. wonchoeli sp. nov.
Figure 23.
Distribution of Scottolana species. (1) S. antillensis; (2) S. brevifurca; (3) S. bulbifera; (4) S. bulbosa; (5) S. dissimilis; (6) S. geei; (7) S. glabra; (8) S. gomezi; (9) S. huysi; (10) S. inopinata; (11) S. jasani; (12) S. longipes; (13) S. oleosa; (14) S. rostrata; (15) S. scotti; (16) S. tumidiseta; (17) S. uxoris; (18) S. daecheonensis sp. nov.; (19) S. wonchoeli sp. nov.

According to Mu and Huys [9], two new species are placed in the longipes-group on the basis of the following apomorphic characteristics: (1) urosome with only two free abdominal somites in female and three in male; (2) P4 modified in both sexes, including the outward relocation of exopod and the elongation of several armature elements (e.g., P4 enp-1 inner seta longer than the entire endopod); (3) P2–P3 basis with curved spinous process; (4) caudal ramus showing a hook-like projection on the inner margin and a spinular patch around the outer distal corner. As stated in the World of Copepods database [6], 17 species of Scottolana have been reported to date, 7 of which belong to the longipes-group. Including these two new species, the longipes-group contains the following 9 species: S. longipes, S. longipes sensu Por (1964), S. longipes sensu Wells (1967), S. longipes sensu Wells and Rao (1987), S. dissimilis, S. uxoris, S. geei, S. jasani, S. huysi, S. gomezi, S. daecheonensis sp. nov. and S. wonchoeli sp. nov.
However, S. daecheonensis sp. nov. does not satisfy all of the apomorphies for the longipes-group proposed by Mu and Huys [9], due to: (1) a lack of modified tube-pore on male P3, and (2) no sexual dimorphism in caudal seta II. Despite these differences, S. daecheonensis sp. nov. clearly belongs to the longipes-group, and we agree with Nazari et al. [18], who argued that for the same reasons as ours the above two characters should be excluded from the apomorphies of the longipes-group.
Scottolana daecheonensis sp. nov. is the largest species of the genus (1940 µm) and is most closely related to S. jasani from the Yellow Sea, Korea, and S. gomezi from Bandar Abbas, Iran. It differs from the congeners in the following ways: (1) rostrum has two tiny lateral sensilla (the absence of sensilla in S. gomezi); (2) 4-segemented antennule in female and the 5-segmented in male; (3) 3-segmented mandibular exopod (4-segmented in S. jasani and indistinctly 4-segmented in S. gomezi); (4) P3 basis with an inner spinous process on the posterior surface; (5) P3 enp-3 without a modified tube-pore along the inner margin in male (with a modified tube-pore in S. jasani); (6) P4 enp-2 crumpled inner margin with strong spinules in male; (7) P5 second-outermost seta naked (plumose in S. jasani and S. gomezi); (8) caudal rami with an unguiform extension on the inner margins (without extension along the anterior inner margin in male S. gomezi); and (9) caudal seta V with a normal base in female (modified to a bulbous process in S. gomezi).
Based on previous descriptions [13,31], Scottolana wonchoeli sp. nov. is similar to S. geei from the Bohai Sea and S. longipes sensu Wells and Rao (1987) from the Andaman Islands. However, S. wonchoeli sp. nov. can be differentiated from the congeners by (1) rostrum with a broadly rounded apex (with a narrowly rounded apex in S. geei); (2) 3-segemented antennule in female (7-segmented in S. longipes sensu Wells and Rao (1987)), and 5-segmented in male (7-segmented in S. longipes sensu Wells and Rao (1987) and 4-segmented in S. geei); (3) 8-segmented antennary exopod (9-segmented in S. geei); (4) a relatively small female genital field, extending about one-third of genital double-somite (extending over three-quarters of genital double-somite in S. geei); (5) male genital somite with paired lateral spinous processes (strongly developed in S. geei and minute in S. longipes); (6) male P6 with a long proximal element (much longer and twisted in S. geei); and (7) maxillular endopod with eleven setae in total.
It is known that the intraspecific mitochondrial COI sequence distance of copepods varies from about 0.26 to 1.6% [32,33]. The intraspecific COI sequence distance of S. daecheonensis sp. nov. is 0.609–0.917%, which supports the results of previous studies. In the genus Scottolana, the interspecific maximum distance of the COI sequence (20.031%) is similar [21,34] or greater [32] than that in other genera. The 18S rRNA genes are highly conserved intra-species [35]. Moreover, in this study, the intraspecific distances of the 18S rRNA sequences are close to 0%, similar to the results in previous studies [21,34]. Since the distances of the sequences between species or within species are different for each taxon [33], it is necessary to secure more information about the average distance for each taxon in order to identify copepods using the distance of the DNA sequence.
A key for the identification of the species of the genus Scottolana is provided. It is amended from Wells [31].
| 1. | Urosome with two free abdominal somites in female and three in male; P4 modified in both sexes, including an outward relocation of exopod, and the elongation of several armature elements; P2–P3 basis with a curved spinous process; caudal ramus with a hook-like projection on the inner margin and a spinular patch around the outer distal corner ........................................................................................................ longipes-group ... 2 |
| — | These characters not combined ............................................................................... 9 |
| 2. | Male P3 endopod-3 with modified tube-pore along inner margin ……........... 3 |
| — | Male P3 endopod-3 without modified tube-pore …............................................ 6 |
| 3. | Antennary exopod 9-segmented; female caudal ramus seta II long and plumose with a bulbiform base ...................................................................... S. geei |
| — | Antennary exopod 8-segmented; female caudal ramus seta II with normal base or very short ........................................................................................ 4 |
| 4. | Female antennule 6- or 7-segmented; maxillular endopod with ten setae in total ................................................................................................ S. longipes |
| — | Female antennule 3-segmented; maxillular endopod with eleven setae in total ............................................................................................................... 5 |
| 5. | Rostrum with acutely pointed apex; caudal ramus approximately twice as long as wide .................................................................................... S. jasani |
| — | Rostrum with rounded apex; caudal ramus at least 2.5 times as long as wide ....................................................................................... S. wonchoeli sp. nov. |
| 6. | Female caudal seta II very short ............................................................ S. dissimilis |
| — | Female caudal seta II with bulbiform base ................................................ S. huysi |
| — | Female caudal seta II with normal base ................................................................. 7 |
| 7. | Caudal ramus less than 1.3 times as long as wide; maxillulary endopod with ten setae in total; P4 exopod extends to about the end of endopod-3 .......................................................................................... S. uxoris |
| — | Caudal ramus about twice as long as wide; maxillulary endopod with eleven setae in total; P4 exopod extends to about halfway along endopod-3 .................................................................................................................. 8 |
| 8. | Rostrum without lateral sensilla; female antennule 3-segmented; mandibular exopod 4-segmented; caudal seta V modified to a bulbous process in female .......................................................................... S. gomezi |
| — | Rostrum with two lateral sensilla; female antennule 4-segmented; mandibular exopod 4-segmented; caudal seta V with normal base in female ............................................................................. S. daecheonensis sp. nov. |
| 9. | P2 endopod-3 with six setae; P4 exopod-3 with five setae ................... S. bulbosa |
| — | P2 endopod-3 with five setae; P4 exopod-3 with four setae ............................. 10 |
| 10. | P1 endopod-1 without inner seta ................................................................. S. scotti |
| — | P1 endopod-1 with an inner seta .......................................................................... 11 |
| 11. | P4 exopod extends only to about the end of endopod-2 .................. S. brevifurca |
| — | P4 exopod extends to about halfway along endopod-3 .................................... 12 |
| — | P4 exopod extends to about the end of endopod-3 ............................................ 13 |
| 12. | Caudal seta II long and smooth ............................................................. S. inopinata |
| — | Caudal seta II very thick ....................................................................... S. antillensis |
| 13. | Caudal ramus about as long as wide ......................................................... S. glabra |
| — | Caudal ramus about 1.5 times as long as wide ...................................... S. rostrata |
| — | Caudal ramus about twice as long as wide ......................................................... 14 |
| 14. | Antennary exopod 8-segmented ............................................................ S. bulbifera |
| — | Antennary exopod 7-segmented .......................................................................... 15 |
| 15. | Caudal setae all slender and filiform ......................................................... S. oleosa |
| — | Caudal ramus with two bulbiform setae ........................................... S. tumidiseta |
Supplementary Materials
The following are available online at https://www.mdpi.com/article/10.3390/d14110967/s1, Table S1: Pairwise distances (Tamura–Nei distance) based on 658 bp between COI sequences among species in Canuellidae; Table S2: Pairwise distances (Tamura–Nei distance) based on 1728 bp between 18S rRNA sequences among species in Canuellidae.
Author Contributions
Conceptualization, H.W.B. and J.B.; data curation, H.M.; funding acquisition, H.W.B. and J.B.; investigation, H.M.; methodology, H.W.B. and J.B.; project administration, J.B.; visualization, H.M.; writing—original draft, H.W.B.; writing—review and editing, H.W.B. and J.B. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the Operation of Depository, Registry, and Reserve (MABIK 2022M01100) and the Marine Biological Resources Management and Systematic Research (MABIK 2022M00200), sponsored by the National Marine Biodiversity Institute of Korea.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Lang, K. Monographie der Harpacticiden (Vorläufige Mitteilung); Almqvist & Wiksells Boktryckeri Ab: Uppsala, Sweden, 1944; pp. 1–39. [Google Scholar]
- Dahms, H.U. Usefulness of postembryonic characters for phylogenetic reconstruction in Harpacticoida (Crustacea, Copepoda). Bull. Plankt. Soc. Jpn. Spec. 1991, 1991, 87–104. [Google Scholar]
- Dahms, H.U. Exclusion of the Polyarthra from Harpacticoida and its reallocation as an underived branch of the Copepoda (Arthropoda, Crustacea). Invertebr. Zool. 2004, 1, 29–51. [Google Scholar] [CrossRef]
- Khodami, S.; McArthur, J.V.; Blanco-Bercial, L.; Arbizu, P.M. RETRACTED ARTICLE: Molecular Phylogeny and Revision of Copepod Orders (Crustacea: Copepoda). Sci. Rep. 2017, 7, 9164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mikhailov, K.V.; Ivanenko, V.N. Low support values and lack of reproducibility of molecular phylogenetic analysis of Copepoda orders. bioRxiv 2019. [Google Scholar] [CrossRef]
- Walter, T.C.; Boxshall, G. World of Copepods database. Canuellidae Lang. 1944. Available online: https://www.marinespecies.org/aphia.php?p=taxdetails&id=115141 (accessed on 23 September 2022).
- Chislenko, L.L. Novye massovye formy garpaktitsid (Copepoda, Harpacticoida) iz zaliva Pos’eta Yaponskogo morya. [New common forms of harpacticids (Copepoda, Harpacticoida) from Possjet Bay of the Sea of Japan.]. Issled. Fauny Morei 1971, 8, 151–181. (In Russian) [Google Scholar]
- Fiers, F. New Canuellidae from the northern coast of Papua New Guinea (Copepoda: Harpacticoida). Bull. Inst. Royal Sci. Nat. Belg. Biologie. 1982, 54, 1–32. [Google Scholar]
- Mu, F.H.; Huys, R. Canuellidae (Copepoda, Harpacticoida) from the Bohai Sea, China. J. Nat. Hist. 2004, 38, 1–36. [Google Scholar] [CrossRef]
- Song, S.J.; Kangtia, P.; Khim, J.S.; Chullasorn, S. Two new Asian species of the genus Scottolana Huys, 2009 (Copepoda: Canuelloida: Canuellidae). J. Nat. Hist. 2018, 52, 377–403. [Google Scholar] [CrossRef]
- Thompson, I.C.; Scott, A. Report on the Copepoda collected by Professor Herdman, at Ceylon, in 1902. Rep. Govt. Ceylon Pearl Oyster Fish. Gulf Manaar 1903, 1 (Suppl. S7), 227–307. [Google Scholar]
- Sewell, R.B.S. Copepoda Harpacticoida. Sci. Rep. John Murray Exped. 1940, 7, 117–382. [Google Scholar]
- Wells, J.B. VII. The Littoral Copepoda (Crustacea) of Inhaca Island, Mozambique. Earth Environ. Sci. Trans. R. Soc. Edinb. 1967, 67, 189–358. [Google Scholar] [CrossRef]
- Wells, J.B.J.; Rao, C.G. Littoral Harpacticoida (Crustacea: Copepoda) from Andaman and Nicobar Islands. Mem. Zool. Surv. India 1987, 16, 1–385. [Google Scholar]
- Fiers, F. Harpacticoid copepods from the West Indian Islands: Canuellidae and Longipediidae (Copepoda, Harpacticoida). Bijdr. Tot Dierkd. 1984, 54, 197–210. [Google Scholar] [CrossRef] [Green Version]
- Por, F.D. A study of the levantine and pontic harpacticoida (Crustacea, Copepoda). Zool. Verh. 1964, 64, 1–128. [Google Scholar]
- Por, F.D. A note on two new species of Canuellidae (Copepoda, Harpacticoida) from the Red Sea. Crustaceana 1983, 44, 187–197. [Google Scholar] [CrossRef]
- Nazari, F.; Mirshamsi, O.; Sari, A.; Aliabadian, M.; Martinez Arbizu, P. Three new Canuellidae (Copepoda: Canuelloida) from Iran. Zootaxa 2018, 4446, 401–441. [Google Scholar] [CrossRef]
- Park, E.O.; Lee, W. New record of Scottolana bulbifera (Copepoda: Harpacticoida: Canuellidae) from Korea. Bull. NIBR 2011, 2, 66–75. [Google Scholar]
- Burgess, R. An improved protocol for separating meiofauna from sediments using colloidal silica sols. Mar. Ecol. Prog. Ser. 2001, 214, 161–165. [Google Scholar] [CrossRef]
- Lim, B.J.; Bang, H.W.; Moon, H.; Back, J. Integrative description of Diosaccus koreanus sp. nov. (Hexanauplia, Harpacticoida, Miraciidae) and integrative information on further Korean species. ZooKeys 2020, 927, 1–35. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Nei, M. Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Mol. Biol. Evol. 1993, 10, 512–526. [Google Scholar] [PubMed] [Green Version]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasegawa, M.; Kishino, H.; Yano, T. Dating the human-ape split by a molecular clock of mitochondrial DNA. J. Mol. Evol. 1985, 22, 160–174. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Huys, R.; Boxshall, G.A. Copepod Evolution; The Ray Society: London, UK, 1991; pp. 1–468. [Google Scholar]
- Por, F.D. Level bottom Harpacticoida (Crustacea, Copepoda) from Elat (Red Sea), part I. Isr. J. Zool. 1967, 16, 101–165. [Google Scholar]
- Huys, R. Unresolved cases of type fixation, synonymy and homonymy in harpacticoid copepod nomenclature (Crustacea: Copepoda). Zootaxa 2009, 2183, 1–99. [Google Scholar] [CrossRef]
- Huys, R. Harpacticoid copepods—Their symbiotic associations and biogenic substrata: A review. Zootaxa 2016, 4174, 448–729. [Google Scholar] [CrossRef]
- Wells, J.B. An annotated checklist and keys to the species of Copepoda Harpacticoida (Crustacea). Zootaxa 2007, 1568, 1–872. [Google Scholar] [CrossRef] [Green Version]
- Baek, S.Y.; Jang, K.H.; Choi, E.H.; Ryu, S.H.; Kim, S.K.; Lee, J.H.; Lim, Y.J.; Lee, J.; Jun, J.; Kwak, M.; et al. DNA Barcoding of Metazoan Zooplankton Copepods from South Korea. PLoS ONE 2016, 11, e0157307. [Google Scholar] [CrossRef]
- Yeom, J.; Park, N.; Jeong, R.; Lee, W. Integrative description of cryptic Tigriopus species from Korea using MALDI-TOF MS and DNA barcoding. Front. Mar. Sci. 2021, 8, 648197. [Google Scholar] [CrossRef]
- Bang, H.W.; Moon, H.; Back, J. Two New Species of the Genus Longipedia Claus, 1863 (Copepoda: Harpacticoida: Longipediidae) from Korea, with an Update and a Key to Species. Diversity 2021, 13, 590. [Google Scholar] [CrossRef]
- Wu, S.; Xiong, J.; Yu, Y. Taxonomic resolutions based on 18S rRNA genes: A case study of subclass Copepoda. PLoS ONE 2015, 10, e0131498. [Google Scholar]
Figure 22.
Maximum likelihood tree based on the mitochondrial COI sequences. Numbers above the branches indicate bootstrap values from 1000 replications.
Figure 22.
Maximum likelihood tree based on the mitochondrial COI sequences. Numbers above the branches indicate bootstrap values from 1000 replications.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Information regarding collection and specimens in this study.
| Species Name | Date | Locality | Methods/ Sediment/Depth | Specimen Nos. |
|---|---|---|---|---|
| Genus Scottolana Huys, 2009 | ||||
| S. daecheonensis sp. nov. | 13 May 2021 | 36°13′18″ N 126° 31′32″ E | Light trap/ Muddy/2 m | MABIK CR00252814, MABIK CR00252811 |
| 05 June 2019 | 36°18′00″ N 126°31′02″ E | Hand net/ Rock pool/0 m | MABIK CR00252815, MABIK CR00252812 | |
| 27 March 2018 | 34°54′28″ N 128°9′ 31″ E | Hand net/ Sand beach/0 m | MABIK CR00252816, MABIK CR00252813 | |
| S. wonchoeli sp. nov. | 26 July 2017 | 37°29′13″ N 130°55′20″ E | SCUBA/ Sandy/5 m | MABIK CR00252817, MABIK CR00252818 |
Table 2.
Primer sequences and PCR conditions used in the present study.
| Gene | Primer | Primer Sequence | PCR Condition |
|---|---|---|---|
| COI | MaxilloF MaxilloR | CWAAYCATAAAGAYATTG ACTTCAGGRTGNCCAAAR | 94 °C, 300 s; 40 cycles × (94 °C, 60 s; 46 °C, 120 s; 72 °C, 180 s; 72 °C, 600 s) |
| 18S rRNA | 18S F1 18S R9 18S F2 18S F3 18S F4 18S R6 18S R7 18S R8 | TACCTGGTTGATCCTGCCAG GATCCTTCCGCAGGTTCACCTAC CCTGAGAAACGGCTRCCACAT GYGRTCAGATACCRCCSTAGTT GGTCTGTGATGCCCTYAGATGT TYTCTCRKGCTBCCTCTCC GYYARAACTAGGGCGGTATCTG ACATCTRAGGGCATCACAGACC | 94 °C, 300 s; 40 cycle × (94 °C, 30 s; 50 °C, 30 s; 72 °C, 60 s); 72 °C, 420 s Internal primers, 18S F2–F4 and 18S R6–8, were used for primer walking to sequence over 1700 bps. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Bang, H.W.; Moon, H.; Back, J. Two New Species of the Family Canuellidae Lang, 1944 (Copepoda: Polyarthra), from Korea, with a Key to Species of the Genus Scottolana Huys, 2009. Diversity 2022, 14, 967. https://doi.org/10.3390/d14110967
AMA Style
Bang HW, Moon H, Back J. Two New Species of the Family Canuellidae Lang, 1944 (Copepoda: Polyarthra), from Korea, with a Key to Species of the Genus Scottolana Huys, 2009. Diversity. 2022; 14(11):967. https://doi.org/10.3390/d14110967
Chicago/Turabian StyleBang, Hyun Woo, Heejin Moon, and Jinwook Back. 2022. "Two New Species of the Family Canuellidae Lang, 1944 (Copepoda: Polyarthra), from Korea, with a Key to Species of the Genus Scottolana Huys, 2009" Diversity 14, no. 11: 967. https://doi.org/10.3390/d14110967
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.