
Why Is the Alpine Flora Comparatively Robust against Climatic Warming?


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
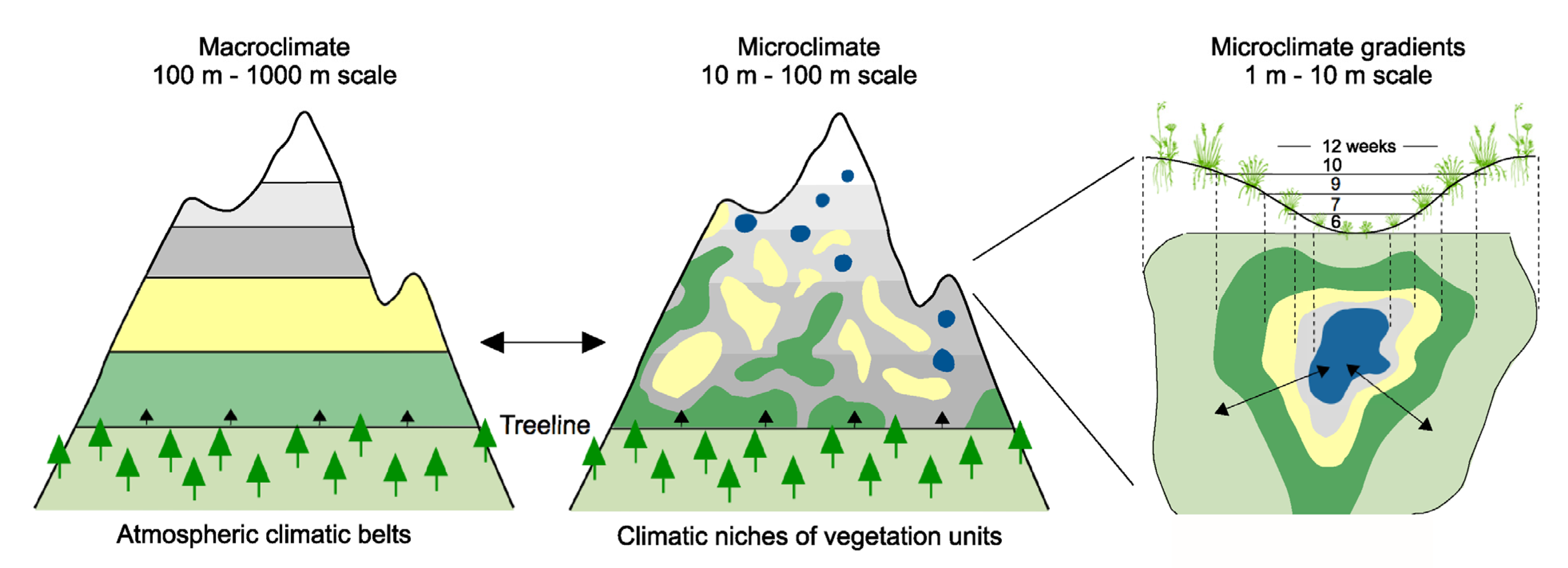
2. Geodiversity Drives Biodiversity
3. Alpine Plant Life Strategies
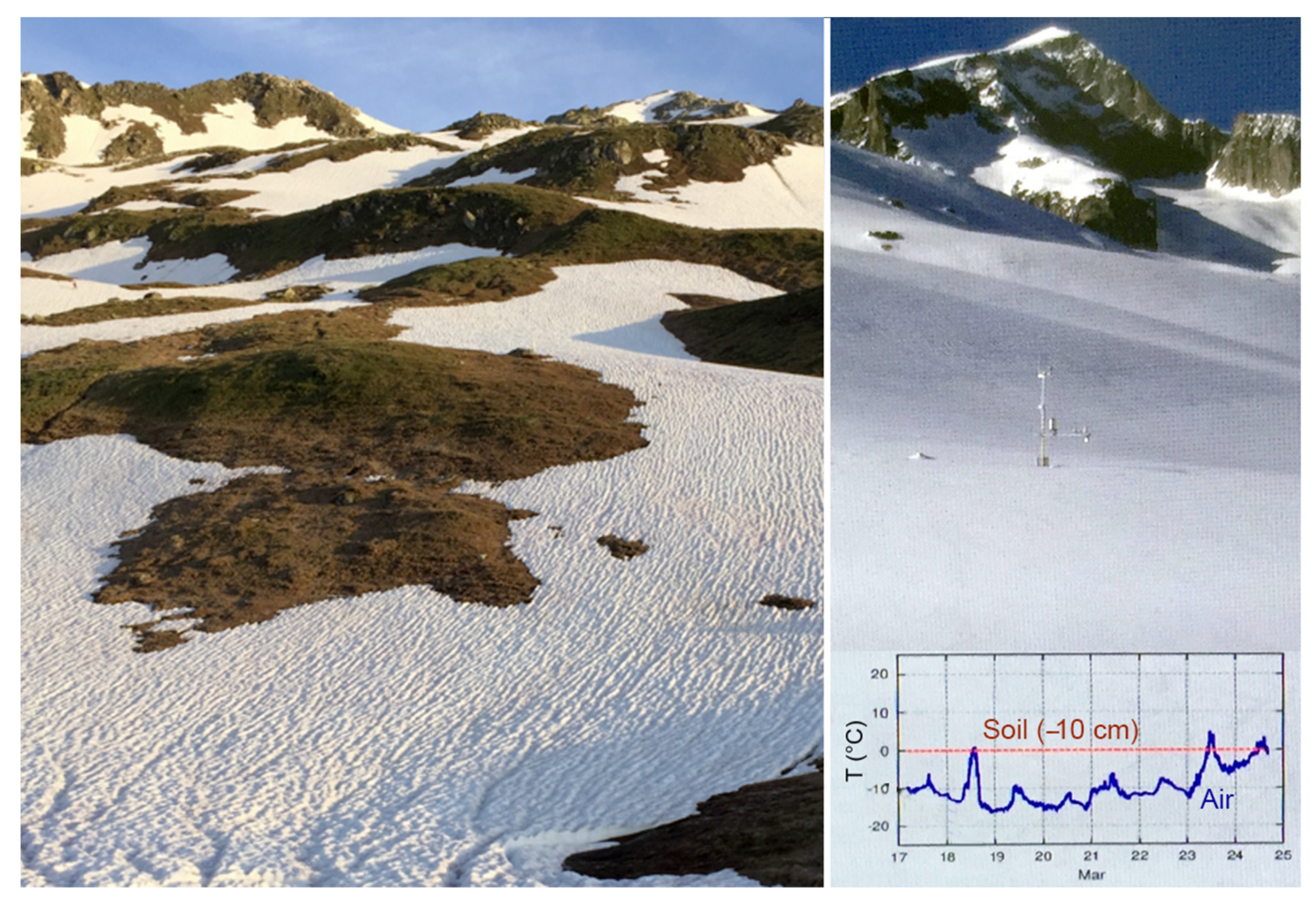
4. The Critical Role of Snow
5. Species Range Limits
6. Change in Alpine Land Area Due to Climatic Warming
7. Viewing Alpine Plant Life over Geological Time Scales
8. Alpine Plant Diversity under Global Change
9. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Körner, C. The use of ‘altitude’ in ecological research. Trends Ecol. Evol. 2007, 22, 569–574. [Google Scholar] [CrossRef] [PubMed]
- Körner, C.; Paulsen, J.; Spehn, E.M. A definition of mountains and their bioclimatic belts for global comparison of biodiversity data. Alp. Bot. 2011, 121, 73–78. [Google Scholar] [CrossRef] [Green Version]
- Körner, C.; Jetz, W.; Paulsen, J.; Payne, D.; Rudmann-Maurer, K.; Spehn, E.M. A global inventory of mountains for bio-geographical applications. Alp. Bot. 2017, 127, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Körner, C. Mountain ecosystems in a changing environment. J. Prot. Mt. Areas Res. Manag. 2014, 6, 71–77. [Google Scholar] [CrossRef] [Green Version]
- Körner, C. Mountain biodiversity, its causes and function. Ambio Spec. Rep. 2004, 13, 11–17. [Google Scholar] [CrossRef]
- Körner, C. Alpine Plant Life, 3rd ed.; Springer: Cham, Switzerland, 2021. [Google Scholar]
- Testolin, R.; Attorre, F.; Borchardt, P.; Brand, R.F.; Bruelheide, H.; Chytrý, M.; De Sanctis, M.; Dolezal, J.; Finckh, M.; Haider, S.; et al. Global patterns and drivers of alpine plant species richness. Glob. Ecol. Biogeogr. 2021, 30, 1218–1231. [Google Scholar] [CrossRef]
- Körner, C. The cold range limit of trees. Trends Ecol. Evol. 2021. [Google Scholar] [CrossRef]
- Paulsen, J.; Körner, C. A climate-based model to predict potential treeline position around the globe. Alp. Bot. 2014, 124, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Scherrer, D.; Körner, C. Infra-red thermometry of alpine landscapes challenges climatic warming projections. Glob. Chang. Biol. 2009, 16, 2602–2613. [Google Scholar] [CrossRef]
- Scherrer, D.; Körner, C. Topographically controlled thermal-habitat differentiation buffers alpine plant diversity against climate warming. J. Biogeogr. 2011, 38, 406–416. [Google Scholar] [CrossRef]
- Winkler, M.; Lamprecht, A.; Steinbauer, K.; Hülber, K.; Theurillat, J.-P.; Breiner, F.; Choler, P.; Ertl, S.; Girón, A.G.; Rossi, G.; et al. The rich sides of mountain summits-a pan-European view on aspect preferences of alpine plants. J. Biogeogr. 2016, 43, 2261–2273. [Google Scholar] [CrossRef]
- Körner, C.; Diemer, M. In situ photosynthetic responses to light, temperature and carbon dioxide in herbaceous plants from low and high altitude. Funct. Ecol. 1987, 1, 179–194. [Google Scholar] [CrossRef]
- Körner, C.; Hiltbrunner, E. The 90 ways to describe plant temperature. Perspect. Plant Ecol. Evol. Syst. 2018, 30, 16–21. [Google Scholar] [CrossRef]
- Oldfather, M.F.; Ackerly, D.D. Microclimate and demography interact to shape stable population dynamics across the range of an alpine plant. New Phytol. 2019, 222, 193–205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cernusca, A. Bestandesstruktur, Bioklima und Energiehaushalt von alpinen Zwergstrauchbeständen. Oecol. Plant 1976, 11, 71–102. [Google Scholar]
- Buri, A.; Grand, S.; Yashiro, E.; Adatte, T.; Spangenberg, J.E.; Pinto-Figeroa, E.; Varrecchia, E.; Guisan, A. What are the most crucial soil variables for predicting the distribution of mountain plant species? A comprehensive study in the Swiss. Alps. J. Biogeogr. 2019, 47, 1143–1153. [Google Scholar]
- Steinger, T.; Körner, C.; Schmid, B. Long-term persistence in a changing climate: DNA analysis suggests very old ages of clones of alpine Carex curvula. Oecologia 1996, 105, 94–99. [Google Scholar] [CrossRef]
- De Witte, L.C.; Armbruster, G.F.J.; Gielly, L.; Taberlet, P.; Stöcklin, J. AFLP markers reveal high clonal diversity and extreme longevity in four key arctic-alpine species. Mol. Ecol. 2012, 21, 1081–1097. [Google Scholar] [CrossRef]
- Mauracher, S.; Wagner, J. Flower preformation in the nival plant Ranunculus glacialis L.: Shoot architecture and impact of growing season length on floral morphogenesis and developmental dynamics. Alp. Bot. 2021, 131, 1–12. [Google Scholar] [CrossRef]
- Hiltbrunner, E.; Arnaiz, J.; Körner, C. Biomass allocation and seasonal non-structural carbohydrate dynamics do not explain the success of tall forbs in short alpine grassland. Oecologia 2021, 1–15. [Google Scholar] [CrossRef]
- Körner, C.; Riedl, S.; Keplinger, T.; Richter, A.; Wiesenbauer, J.; Schweingruber, F.; Hiltbrunner, E. Life at 0 °C: The biology of the alpine snowbed plant. Alp. Bot. 2019, 129, 63–80. [Google Scholar] [CrossRef]
- Vorkauf, M.; Kahmen, A.; Körner, C.; Hiltbrunner, E. Flowering phenology in alpine grassland strongly responds to shifts in snowmelt but weakly to summer drought. Alp. Bot. 2021, 131, 73–88. [Google Scholar] [CrossRef]
- Scholz, K.; Hammerle, A.; Hiltbrunner, E.; Wohlfahrt, G. Analyzing the effects of growing season length on the net ecosystem production of an alpine grassland using model-data fusion. Ecosystems 2018, 21, 982–999. [Google Scholar] [CrossRef] [Green Version]
- Sedlacek, J.; Wheeler, J.A.; Cortes, A.J.; Bossdorf, O.; Hoch, G.; Lexer, C.; Wipf, S.; Karrenberg, S.; Van Kleunen, M.; Rixen, C. The response of the alpine dwarf shrub Salix herbacea to altered snowmelt timing: Lessons from a multi-site transplant experiment. PLoS ONE 2015, 10, e0122395. [Google Scholar] [CrossRef]
- Oberbauer, S.F.; Elmendorf, S.C.; Troxler, T.G.; Hollister, R.D.; Rocha, A.V.; Bret-Harte, M.S.; Dawes, M.A.; Fosaa, A.M.; Henry, G.H.R.; Hoye, T.T.; et al. Phenological response of tundra plants to background climate variation tested using the International Tundra Experiment. Philos. Trans. R. Soc. B Biol. Sci. 2013, 368, 20120481. [Google Scholar] [CrossRef]
- Klein, G.; Rebetez, M.; Rixen, C.; Vitasse, Y. Unchanged risk of frost exposure in subalpine and alpine plants after snowmelt in Switzerland despite climate warming. Int. J. Biomet. 2018, 62, 1755–1762. [Google Scholar] [CrossRef] [PubMed]
- Keller, F.; Körner, C. The role of photoperiodism in alpine plant development. Arct. Antarct. Alp. Res. 2003, 35, 361–368. [Google Scholar] [CrossRef]
- Randin, C.F.; Engler, R.; Normand, S.; Zappa, M.; Zimmermann, N.E.; Pearman, P.B.; Vittoz, P.; Thuiller, W.; Guisan, A. Climate change and plant distribution: Local models predict high-elevation persistence. Glob. Chang. Biol. 2009, 15, 1557–1569. [Google Scholar] [CrossRef] [Green Version]
- Scherrer, D.; Schmid, S.; Körner, C. Elevational species shifts in a warmer climate are overestimated when based on weather station data. Int. J. Biometeorol. 2011, 55, 645–654. [Google Scholar] [CrossRef] [PubMed]
- Feldmeier, S.; Schmidt, B.R.; Zimmermann, N.E.; Veith, M.; Ficetola, G.F.; Lötters, S. Shifting aspect or elevation? The climate change response of ectotherms in a complex mountain topography. Divers. Distrib. 2020, 26, 1483–1495. [Google Scholar] [CrossRef]
- Saetersdal, M.; Birks, H.J.B. A comparative ecological study of Norwegian mountain plants in relation to possible future climatic change. J. Biogeogr. 1997, 24, 127–152. [Google Scholar] [CrossRef]
- Guisan, A.; Theurillat, J.P.; Kienast, F. Predicting the potential distribution of plant species in an Alpine environment. J. Veg. Sci. 1998, 9, 65–74. [Google Scholar] [CrossRef]
- Randin, C.F.; Dirnbock, T.; Dullinger, S.; Zimmermann, N.E.; Zappa, M.; Guisan, A. Are niche-based species distribution models transferable in space? J. Biogeogr. 2006, 33, 1689–1703. [Google Scholar] [CrossRef]
- Löffler, J.; Pape, R. Thermal niche predictors of alpine plant species. Ecology 2020, 101, e02891. [Google Scholar] [CrossRef] [Green Version]
- Qian, H.; Ricklefs, R.E.; Thuiller, W. Evolutionary assembly of flowering plants into sky islands. Nat. Ecol. Evol. 2021, 5, 640–646. [Google Scholar] [CrossRef]
- Körner, C.; Diemer, M.; Schäppi, B.; Niklaus, P.; Arnone, J. The responses of alpine grassland to four seasons of CO2 enrichment: A synthesis. Acta Oecologica 1997, 18, 165–175. [Google Scholar] [CrossRef]
- Inauen, N.; Körner, C.; Hiltbrunner, E. No growth stimulation by CO2 enrichment in alpine glacier forefield plants. Glob. Chang. Biol. 2012, 18, 985–999. [Google Scholar] [CrossRef]
- Hiltbrunner, E.; Schwikowski, M.; Körner, C. Inorganic nitrogen storage in alpine snow pack in the Central Alps (Switzerland). Atmos. Environ. 2005, 39, 2249–2259. [Google Scholar] [CrossRef]
- Kosonen, Z.; Schnyder, E.; Hiltbrunner, E.; Thimonier, A.; Schmitt, M.; Seitler, E.; Thöni, L. Current atmospheric nitrogen deposition still exceeds critical loads for sensitive, semi-natural ecosystems in Switzerland. Atmos. Environ. 2019, 211, 214–225. [Google Scholar] [CrossRef]
- Vorkauf, M.; Marty, C.; Kahmen, A.; Hiltbrunner, E. Past and future snowmelt trends in the Swiss Alps: The role of temperature and snowpack. Clim. Chang. 2021, 165, 1–19. [Google Scholar] [CrossRef]
- Bürli, S.; Theurillat, J.-P.; Winkler, M.; Lamprecht, A.; Pauli, H.; Rixen, C.; Steinbauer, K.; Wipf, S.; Abdaladze, O.; Andrews, C.; et al. A common soil temperature threshold for the upper limit of alpine glasslands in European mountains. Alp. Bot. 2021, 131, 41–52. [Google Scholar] [CrossRef]
- Körner, C. Alpine Treelines; Springer: Basel, Switzerland, 2012. [Google Scholar]
- Larcher, W.; Kainmüller, C.; Wagner, J. Survival types of high mountain plants under extreme temperatures. Flora Morphol. Distrib. Funct. Ecol. Plants 2010, 205, 3–18. [Google Scholar] [CrossRef]
- Paulsen, J.; Weber, U.M.; Körner, C. Tree growth near treeline: Abrupt or gradual reduction with altitude? Arct. Antarct. Alp. Res. 2000, 32, 14–20. [Google Scholar] [CrossRef]
- Dolezal, J.; Kurnotova, M.; Sastna, P.; Klimesova, J. Alpine plant growth and reproduction dynamics in a warmer world. New Phytol. 2020, 228, 1295–1305. [Google Scholar] [CrossRef] [PubMed]
- Hock, R.; Rasul, G.; Adler, C.; Cáceres, B.; Gruber, S.; Hirabayashi, Y.; Jackson, M.; Kääb, A.; Kang, S.; Kutuzov, S.; et al. High mountain areas (Chap. 3). In Special Report on Ocean and Cryosphere in a Changing Climate; Pörtner, H.O., Roberts, D.C., Masson-Delmotte, V., Zhai, P., Tignor, M., Poloczanska, E., Mintenbeck, K., Alegría, A., Nicolai, M., Okem, A., et al., Eds.; IPCC: Nairobi, Kenya, 2020. [Google Scholar]
- Birks, H.J.B. Statistical approaches to interpreting diversity patterns in the Norwegian mountain flora. Ecography 1996, 19, 332–340. [Google Scholar] [CrossRef]
- Schönswetter, P.; Tribsch, A.; Stehlik, I.; Niklfeld, H. Glacial history of high alpine Ranunculus glacialis (Ranunculaceae) in the European Alps in a comparative phylogeographical context. Biol. J. Linn. Soc. 2004, 81, 183–195. [Google Scholar] [CrossRef] [Green Version]
- Kuss, P.; Armbruster, G.F.J.; Aegisdóttir, H.H.; Scheepens, J.F.; Stöcklin, J. Spatial genetic structure of Campanula thyrsoides across the European Alps: Indications for glaciation-driven allopatric subspeciation. Perspect. Plant Ecol. Evol. Syst. 2011, 13, 101–110. [Google Scholar] [CrossRef]
- Scheepens, J.F.; Stöcklin, J. Glacial history and local adaptation explain differentiation in phenotypic traits in the Alpine grassland herb Campanula barbata. Plant Ecol. Divers. 2011, 4, 403–413. [Google Scholar] [CrossRef]
- Jiménez-Alfaro, B.; Abdulhak, S.; Attorre, F.; Bergamini, A.; Carranza, M.L.; Chiarucci, A.; Ćušterevska, R.; Dullinger, S.; Gavilán, R.G.; del Galdo, G.G.; et al. Post-glacial determinants of regional species pools in alpine grasslands. Glob. Ecol. Biogeogr. 2021, 30, 1101–1115. [Google Scholar] [CrossRef]
- Parisod, C. Postglacial recolonisation of plants in the western Alps of Switzerland. Bot. Helv. 2008, 118, 1–12. [Google Scholar] [CrossRef]
- De Vos, J.M.; Joppa, L.N.; Gittleman, J.L.; Stephens, P.R.; Pimm, S.L. Estimating the normal background rate of species extinction. Cons. Biol. 2015, 29, 452–462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heiri, O.; Wick, L.; van Leeuwen, J.F.N.; van der Knaap, W.O.; Lotter, A.F. Holocene tree immigration and the chironomid fauna of a small Swiss subalpine lake (Hinterburgsee, 1515 m asl). Palaeogeogr. Palaeoclimatol. Palaeoecol. 2003, 189, 35–53. [Google Scholar] [CrossRef] [Green Version]
- Muellner-Riehl, A.N.; Schnitzler, J.; Kissling, W.D.; Mosbrugger, V.; Rijsdijk, K.F.; Seijmonsbergen, A.C.; Versteegh, H.; Favre, A. Origins of global mountain plant biodiversity: Testing the ‘mountain-geobiodiversity hypothesis’. J. Biogeogr. 2019, 46, 2826–2838. [Google Scholar] [CrossRef] [Green Version]
- Tinner, W.; Kaltenrieder, P. Rapid responses of high-mountain vegetation to early Holocene environmental changes in the Swiss Alps. J. Ecol. 2005, 93, 936–947. [Google Scholar] [CrossRef]
- Kammer, P.M.; Schob, C.; Choler, P. Increasing species richness on mountain summits: Upward migration due to anthropogenic climate change or re-colonisation? J. Veg. Sci. 2007, 18, 301–306. [Google Scholar] [CrossRef]
- Stöckli, V.; Wipf, S.; Nilsson, C.; Rixen, C. Using historical plant surveys to track biodiversity on mountain summits. Plant Ecol. Divers. 2012, 4, 415–425. [Google Scholar] [CrossRef]
- Wipf, S.; Stöckli, V.; Herz, K.; Rixen, C. The oldest monitoring site of the Alps revisited: Accelerated increase in plant species richness on Piz Linard summit since 1835. Plant Ecol. Divers. 2013, 6, 447–455. [Google Scholar] [CrossRef]
- Ren, Y.Y.; Ren, G.Y.; Sun, X.B.; Shresta, A.B.; You, Q.L.; Zhan, Y.L.; Rajbhandari, R.; Zhang, P.-F.; Wen, K.-M. Observed changes in surface air temperature and precipitation in the Hindukush Himalaya region during 1901–2014. Adv. Clim. Chang. Res. 2017, 8, 148–156. [Google Scholar] [CrossRef]
- Pauli, H.; Gottfried, M.; Dullinger, S.; Abdaladze, O.; Akhalkatsi, M.; Alonso, J.L.B.; Coldea, G.; Dick, J.; Erschbamer, B.; Fernández Calzado, R.; et al. Recent plant diversity changes on Europe’s mountain summits. Science 2012, 336, 353–355. [Google Scholar] [CrossRef] [Green Version]
- Lamprecht, A.; Semenchuk, P.R.; Steinbauer, K.; Winkler, M.; Pauli, H. Climate change leads to accelerated transformation of high-elevation vegetation in the central Alps. New Phytol. 2018, 220, 447–459. [Google Scholar] [CrossRef]
- Steinbauer, M.J.; Grytnes, J.-A.; Jurasinski, G.; Kulonen, A.; Lenoir, J.; Pauli, H.; Rixen, C.; Winkler, M.; Bardy-Durchhalter, M.; Barni, E.; et al. Accelerated increase in plant species richness on mountain summits is linked to warming. Nature 2018, 556, 231–234. [Google Scholar] [CrossRef] [PubMed]
- Steinbauer, K.; Lamprecht, A.; Semenchuk, P.R.; Winkler, M.; Pauli, H. Dieback and expansions: Species-specific responses during 20 years of amplified warming in the high Alps. Alp. Bot. 2019, 130, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Roth, T.; Plattner, M.; Amrhein, V. Plants, Birds and Butterflies: Short-Term Responses of Species Communities to Climate Warming Vary by Taxon and with Altitude. PLoS ONE 2014, 9, e82490. [Google Scholar] [CrossRef] [PubMed]
- Winkler, M.; Illmer, P.; Querner, P.; Fischer, B.M.; Hofmann, K.; Lamprecht, A.; Praeg, N.; Schied, J.; Steinbauer, K.; Pauli, H. Side by side? Vascular plant, invertebrate, and microorganism distribution patterns along an alpine to nival elevation gradient. Arctic, Antarct. Alp. Res. 2018, 50, e1475951. [Google Scholar]
- Vitasse, Y.; Ursenbacher, S.; Klein, G.; Bohnenstengel, T.; Chittaro, Y.; Delestrade, A.; Monnerat, C.; Rebetez, M.; Rixen, C.; Strebel, N.; et al. Phenological and elevational shifts of plants, animals and fungi under climate change in the European Alps. Nat. Sci. 2021. [Google Scholar] [CrossRef]
- Vittoz, P.; Randin, C.; Dutoit, A.; Bonnet, F.; Hegg, O. Low impact of climate change on subalpine grasslands in the Swiss Northern Alps. Glob. Chang. Biol. 2009, 15, 209–220. [Google Scholar] [CrossRef]
- Rumpf, S.B.; Hülber, K.; Klonner, G.; Moser, D.; Schütz, M.; Wessely, J.; Willner, W.; Zimmermann, N.E.; Dullinger, S. Range dynamics of mountain plants decrease with elevation. PNAS 2018, 115, 18481853. [Google Scholar] [CrossRef] [Green Version]
- Körner, C. Comparative, long-term ecosystem monitoring across the Alps: Austrian Hohe Tauern National Park, South-Tyrol and Swiss central Alps. In 6th Symposium for Research in Protected Areas; Bauch, K., Ed.; Salzburger Nationalparkfonds, Mittersill, and Austrian Acad Sci: Vienna, Austria, 2018; pp. 331–337. [Google Scholar]
- Little, C.J.; Wheeler, J.A.; Sedlacek, J.; Cortes, A.J.; Rixen, C. Small-scale drivers: The importance of nutrient availability and snowmelt timing on performance of the alpine shrub Salix herbacea. Oecologia 2016, 180, 1015–1024. [Google Scholar] [CrossRef] [Green Version]
- Dullinger, S.; Gattringer, A.; Thuiller, W.; Moser, D.; Zimmermann, N.; Guisan, A.; Willner, W.; Plutzar, C.; Leitner, M.; Mang, T.; et al. Extinction debt of high-mountain plants under twenty-first-century climate change. Nat. Clim. Chang. 2012, 2, 619–622. [Google Scholar] [CrossRef]
- Svenning, J.C.; Sandel, B. Disequilibrium vegetation dynamics under future climate change. Am. J. Bot. 2013, 100, 1266–1286. [Google Scholar] [CrossRef]
- Rumpf, S.B.; Hülber, K.; Wessely, J.; Willner, W.; Moser, D.; Gattringer, A.; Klonner, G.; Zimmermann, N.E.; Dullinger, S. Extinction debts and colonization credits of non-forest plants in the European Alps. Nat. Commun. 2019, 10, 4293. [Google Scholar] [CrossRef] [Green Version]
- Usinowicz, J.; Levine, J.M. Climate-driven range shifts reduce persistence of competitors in a perennial plant community. Glob. Chang. Biol. 2021, 27, 1890–1903. [Google Scholar] [CrossRef]
- Nomoto, H.A.; Alexander, J.M. Drivers of local extinction risk in alpine plants under warming climate. Ecol. Lett. 2021, 24, 1157–1166. [Google Scholar] [CrossRef]
- Weppler, T.; Stoll, P.; Stöcklin, J. The relative importance of sexual and clonal reproduction for population growth in the long-lived alpine plant Geum reptans. J. Ecol. 2006, 94, 869–879. [Google Scholar] [CrossRef]
- Scherrer, D.; Stoll, P.; Stöcklin, J. Colonization dynamics of a clonal pioneer plant on a glacier foreland inferred from spatially explicit and size structured matrix models. Folia Geobot. Phytotaxon. 2017, 52, 353–366. [Google Scholar] [CrossRef]
- Körner, C. Ecology of populations and vegetation. In Strasburger’s Plant Sciences; Bresinsky, A., Kadereit, J.W., Körner, C., Neuhaus, G., Sonnewald, U., Eds.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 1167–1215. [Google Scholar]
- Aegisdottir, H.H.; Kuss, P.; Stocklin, J. Isolated populations of a rare alpine plant show high genetic diversity and considerable population differentiation. Ann. Bot. 2009, 104, 1313–1322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stöcklin, J.; Kuss, P.; Pluess, A.R. Genetic diversity, phenotypic variation and local adaption in the alpine landscape: Case studies with alpine plant species. Bot. Helv. 2009, 119, 125–133. [Google Scholar] [CrossRef]
- Körner, C. Fading of the temperature-growth coupling in treeline trees reflects a conceptual bias. Glob. Chang. Biol. 2021, 27, 3951–3952. [Google Scholar] [CrossRef]
- Knaus, P.; Antoniazza, S.; Wechsler, S.; Guélat, J.; Kéry, M.; Strebel, N.; Sattler, T. Schweizer Brutvogelatlas 2013–2016. Verbreitung und Bestandsentwicklung der Vögel in der Schweiz und im Fürstentum Liechtenstein; Schweizer Vogelwarte: Sempach, Switzerland, 2018. [Google Scholar]
- Descombes, P.; Pitteloud, C.; Glausen, G.; Defossez, E.; Kergunteuil, A.; Allard, P.M.; Rasmann, S.; Pellisier, L. Novel trophic interactions under climate change promote alpine plant coexistance. Science 2020, 370, 1469–1473. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Körner, C.; Hiltbrunner, E. Why Is the Alpine Flora Comparatively Robust against Climatic Warming? Diversity 2021, 13, 383. https://doi.org/10.3390/d13080383
Körner C, Hiltbrunner E. Why Is the Alpine Flora Comparatively Robust against Climatic Warming? Diversity. 2021; 13(8):383. https://doi.org/10.3390/d13080383
Chicago/Turabian StyleKörner, Christian, and Erika Hiltbrunner. 2021. "Why Is the Alpine Flora Comparatively Robust against Climatic Warming?" Diversity 13, no. 8: 383. https://doi.org/10.3390/d13080383