
Shedding Light on the Dark Ages: Sketching Potential Trade Relationships in Early Medieval Romania through Mitochondrial DNA Analysis of Sheep Remains

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection, Archaeological Context, and Radiocarbon Dating
2.2. DNA Extraction, PCR Amplification and mtDNA Sequencing
2.3. Sequence and Population Genetics Analyses
- In order to investigate the genetic relationship between the Capidava cohort and other ancient sequences representative for different geographic regions and historical periods, as well as to estimate the degree of admixture and determine potential trade routes and migration patterns, a Median Joining Network [51] was constructed and pairwise genetic distances were calculated using the Ovis_Cap sequences and 41 sequences available in the international databases (GenBank Acc. No. available in Table S2). All sequences were aligned and trimmed a 304 bp region between the nucleotide positions 16099 to 16402 (NC_001941) to allow for a homogenous set.
- To estimate the degree of genetic continuity and similarity to modern-day breeds, a Median Joining Network [51] was constructed, and pairwise genetic distances were calculated on the Ovis_Cap sequences and 37 and 59 sequences, respectively, representing a variety of breeds originating from different countries (GenBank Acc. No. available in Table S2). All sequences were aligned and trimmed to a 474 bp region, between the nucleotide positions 16027 to 16500 (NC_001941).
- As a means to analyze whether the Capidava cohort shared a higher percentage of haplotypes with the ancient or modern data set, a shared haplotype analysis (SHA) was performed by counting the absolute and relative number of shared haplotypes between the Ovis_Cap sequences and 88 other sequences, combining the previously defined ancient and modern data sets (GenBank Acc. No. available in Table S2). All sequences were trimmed to a 304 bp region, between the nucleotide positions 16092 to 16396 (NC_001941).
3. Results
3.1. Radiocarbon Dating, PCR Amplification and Identified SNPs
3.2. Summary Statistics Analysis
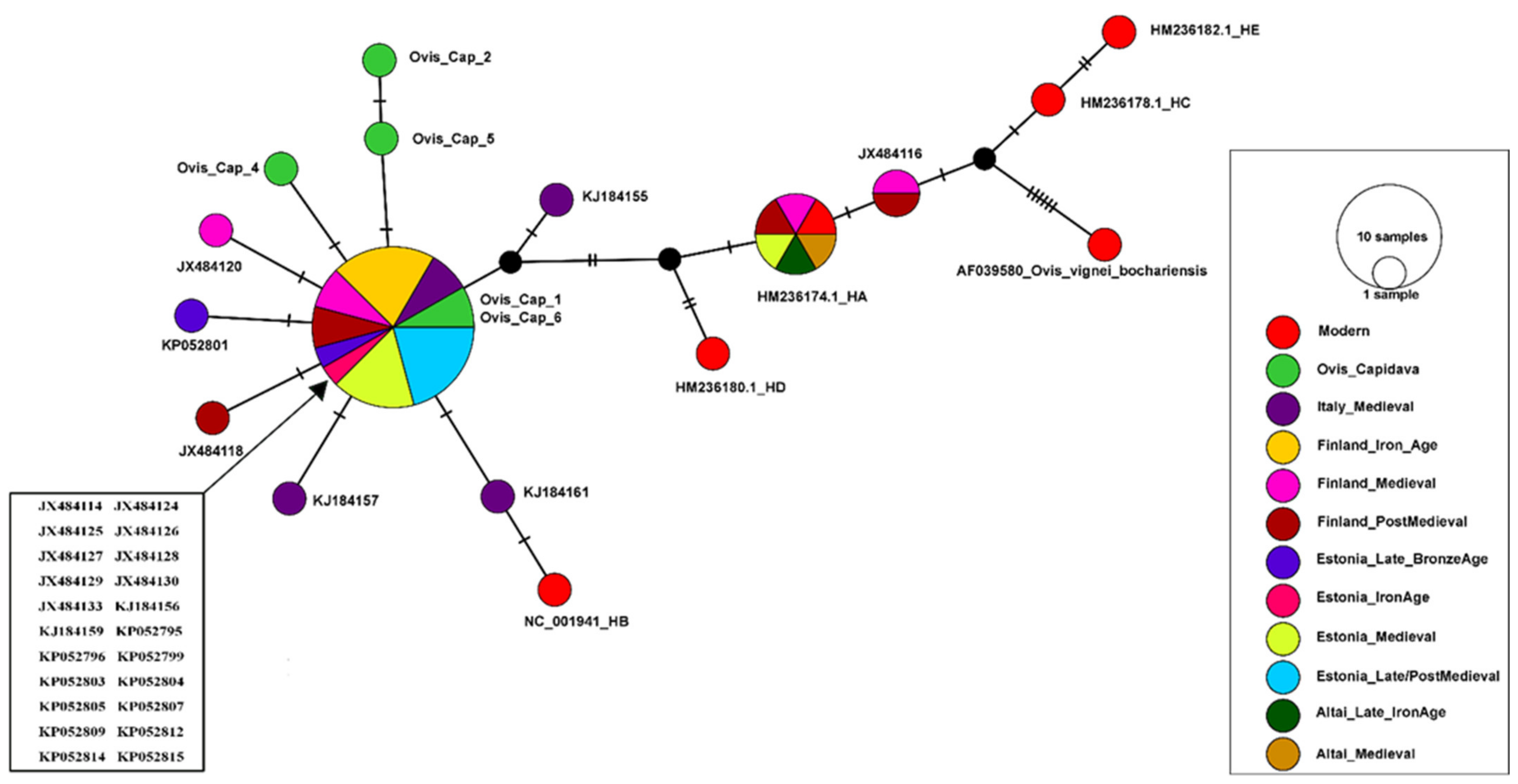
3.3. Capidava Sheep Compared to Ancient Cohorts
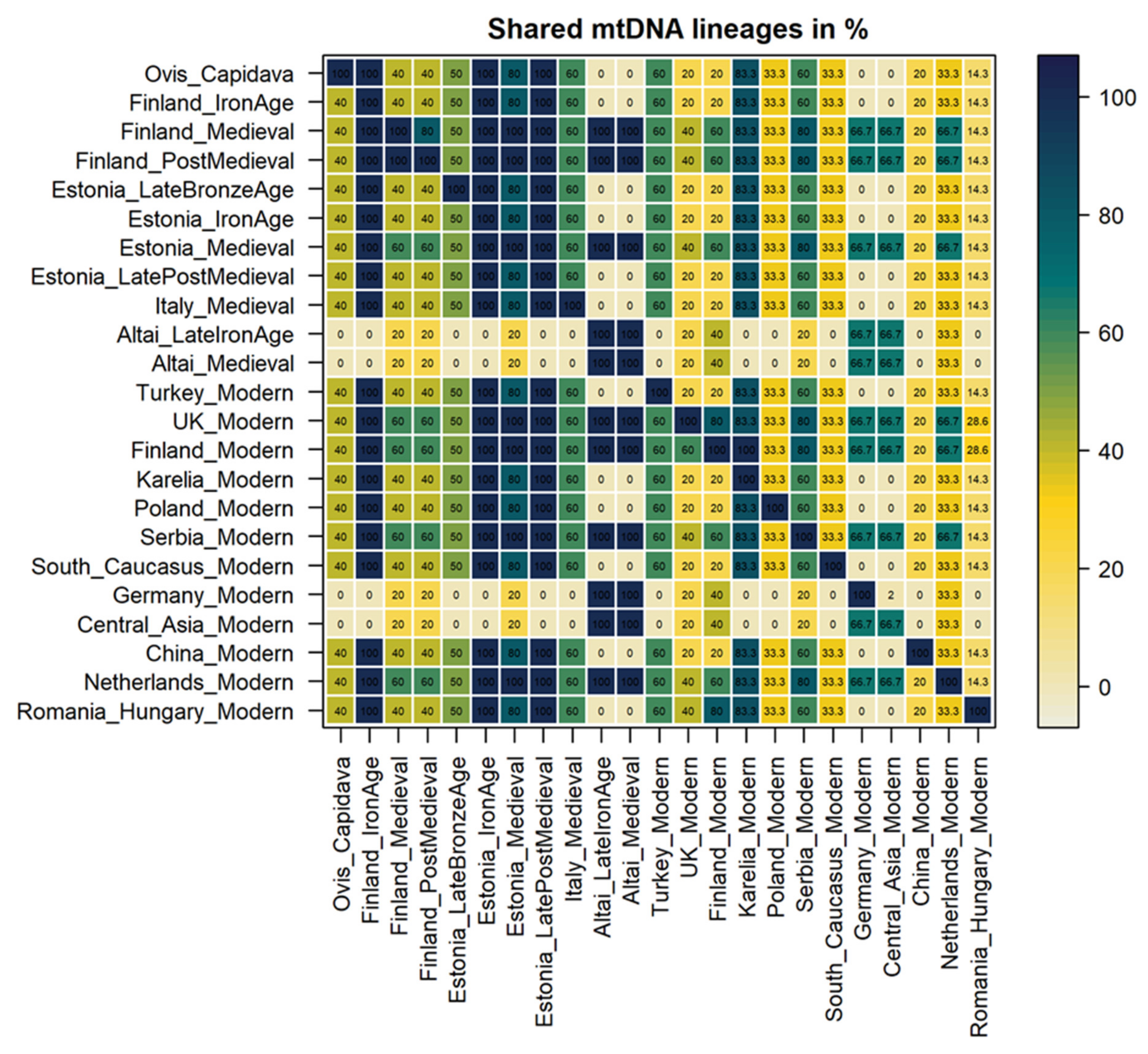
3.3.1. Haplotype Similarity Analysis
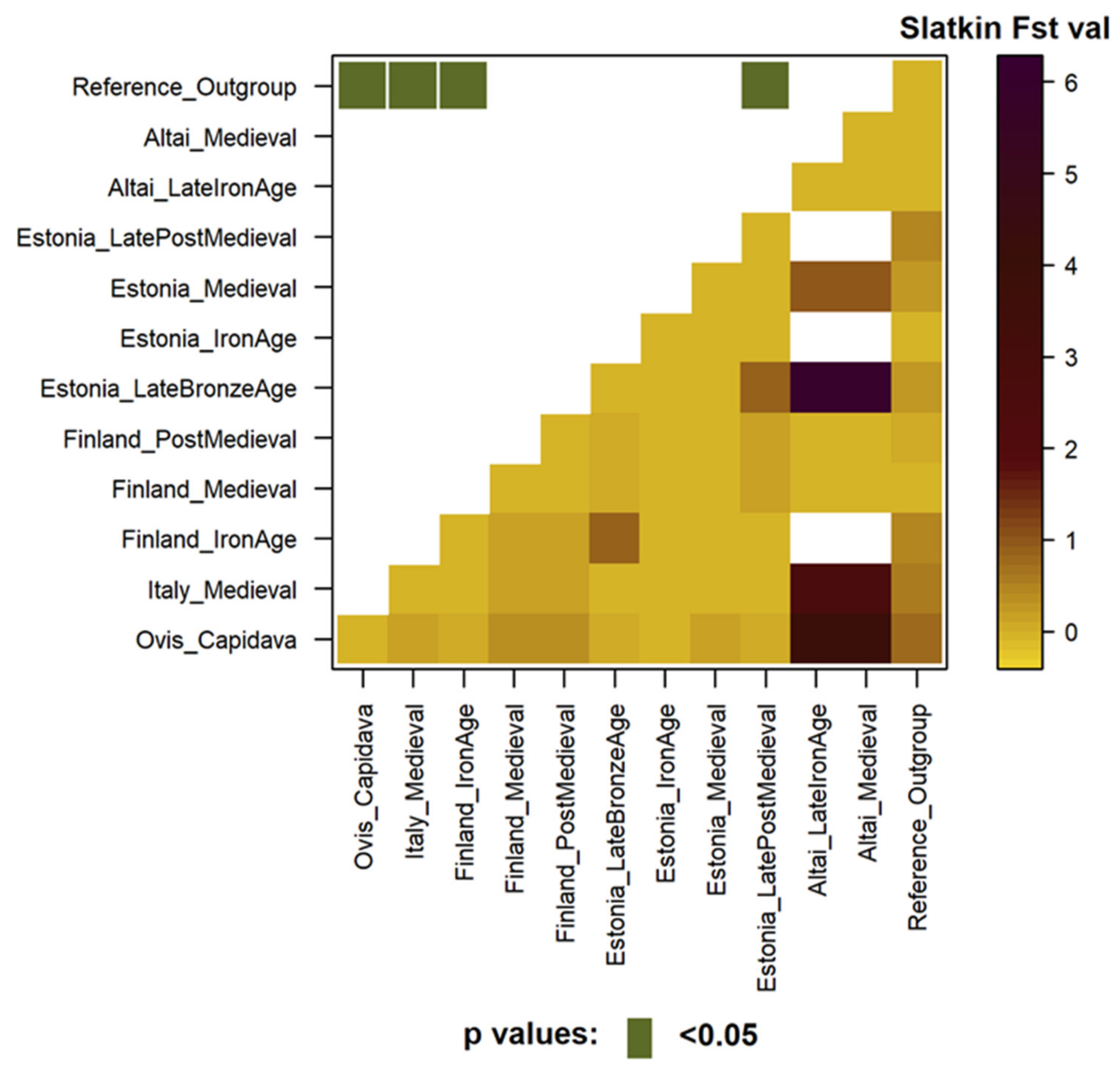
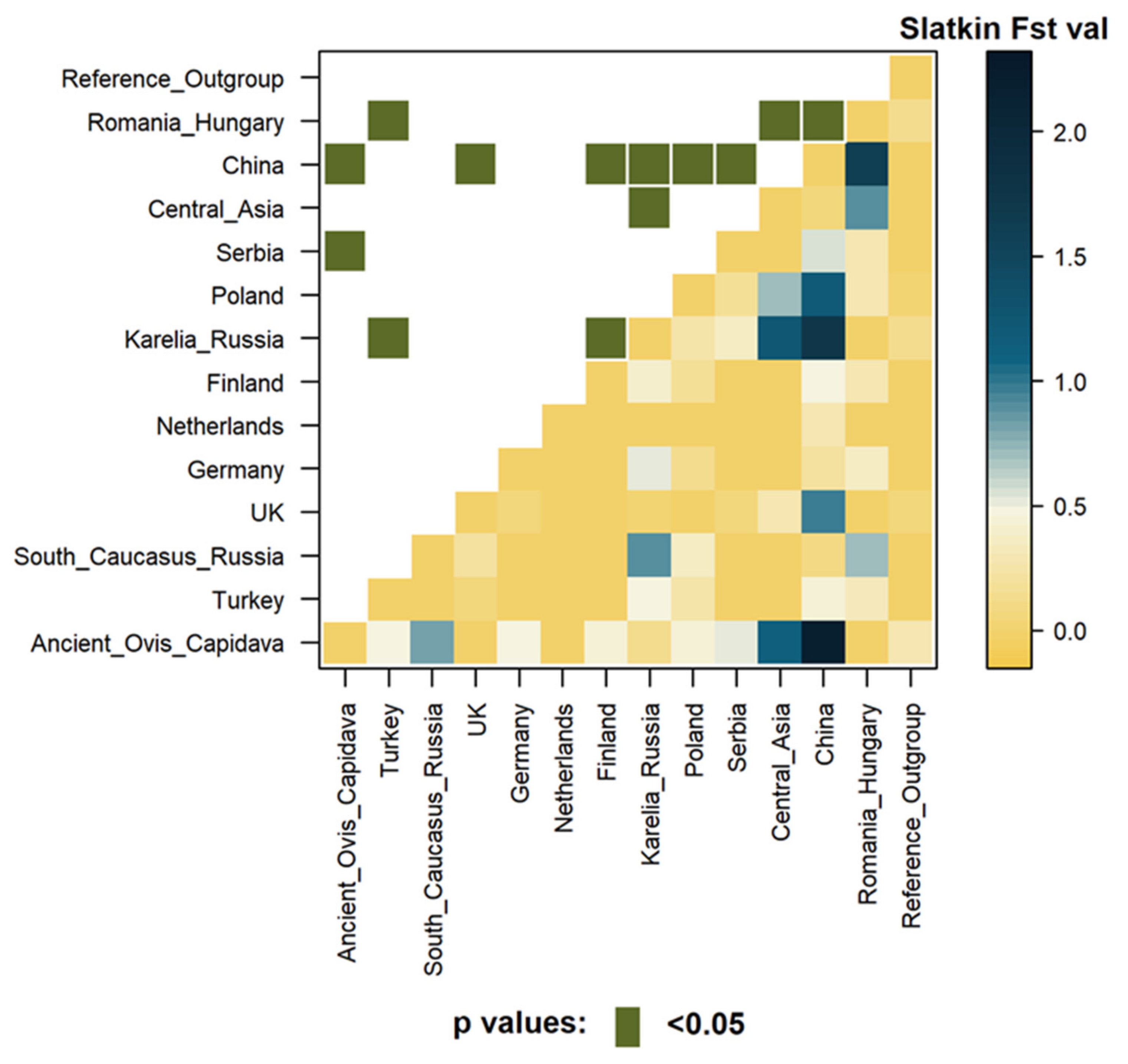
3.3.2. Genetic Distance Estimates
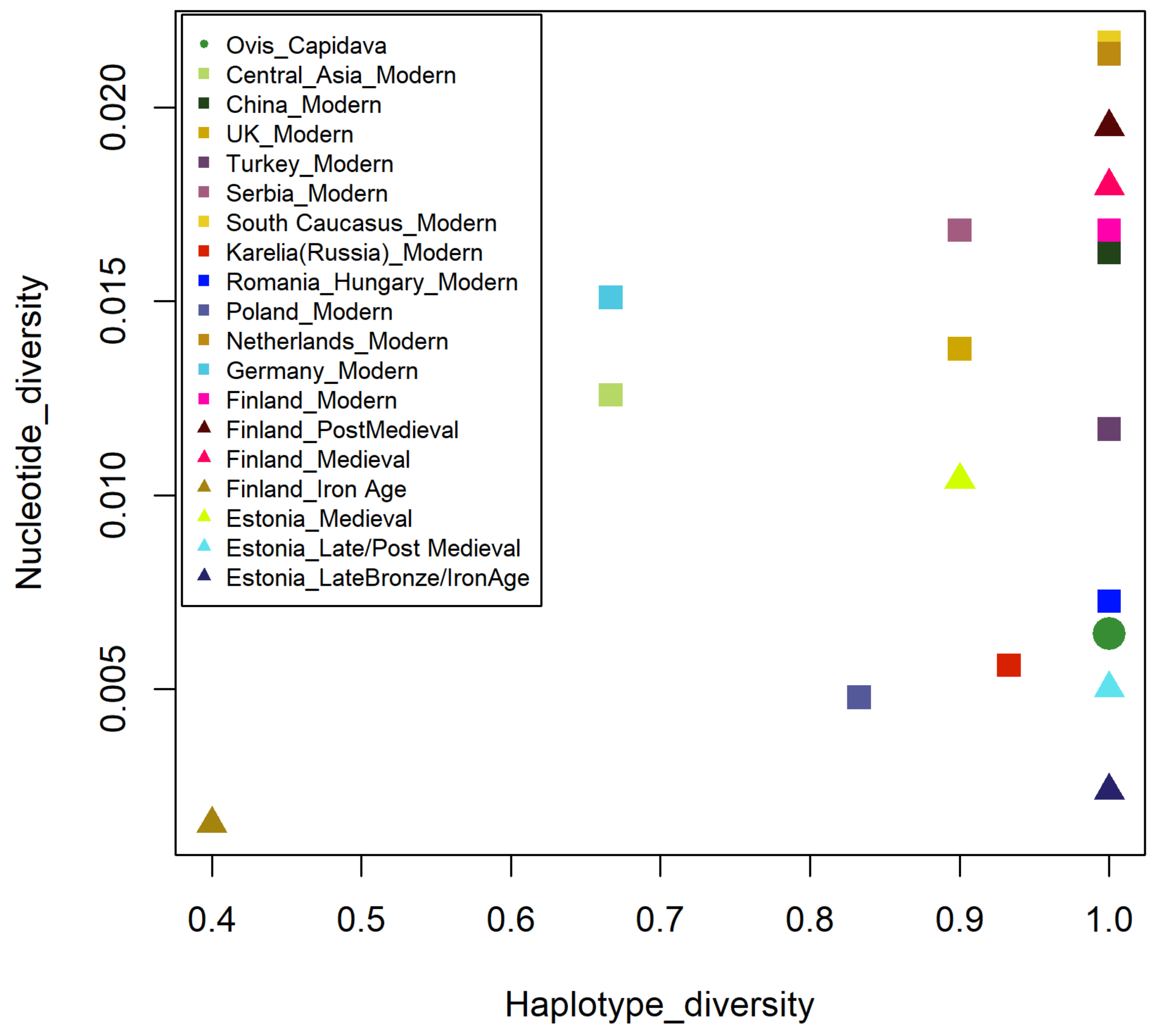
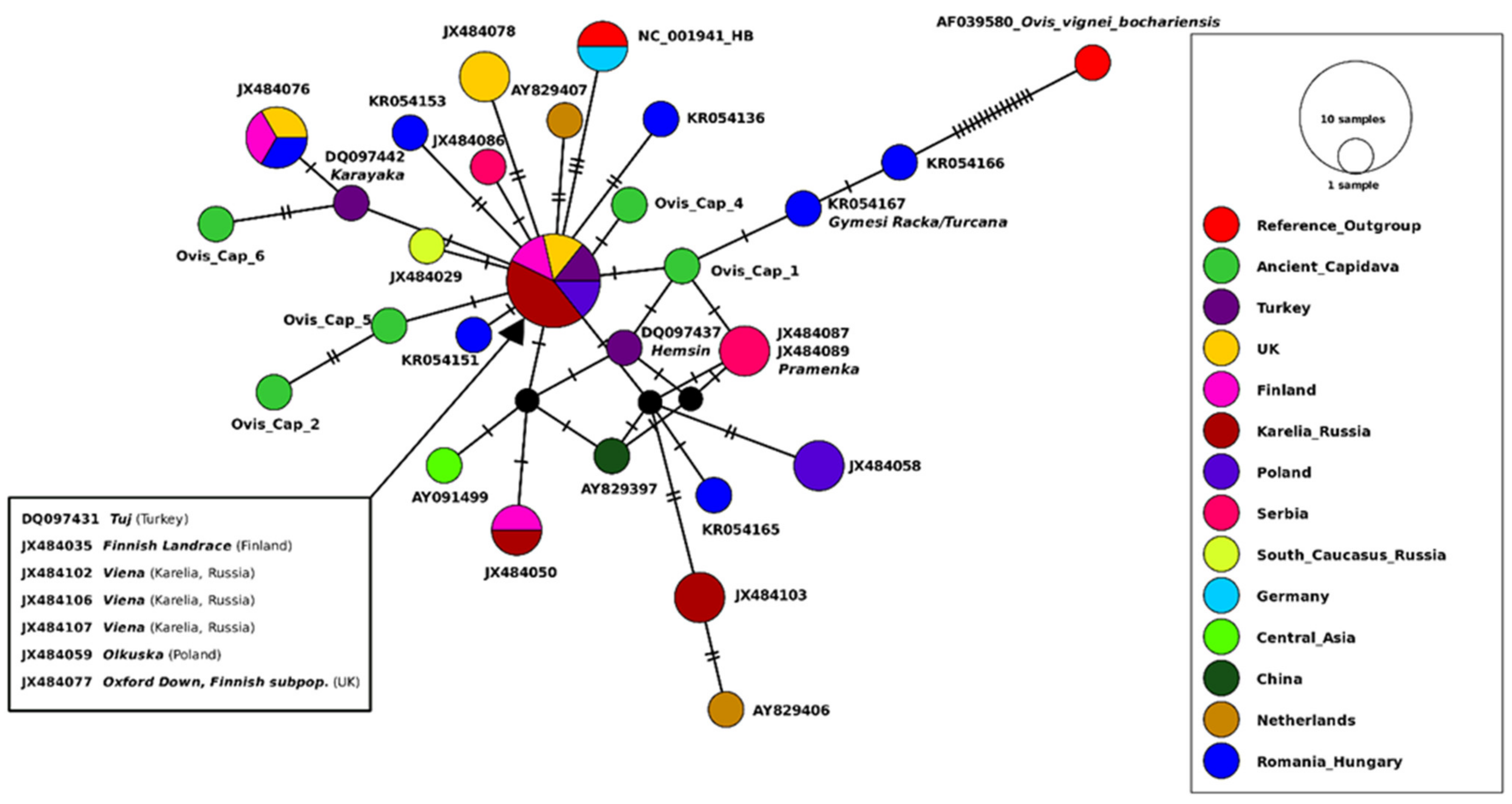
3.4. Capidava Sheep Compared to Modern Populations
3.4.1. Haplotype Similarity Analysis
3.4.2. Genetic Distance Estimates with Focus on Different Geographic Regions
3.5. Analysis of Shared Haplotypes between the Capidava, Ancient, and Modern Datasets
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zeder, M.A. Domestication and early agriculture in the Mediterranean Basin: Origins, diffusion, and impact. Proc. Natl. Acad. Sci. USA 2008, 105, 11597–11604. [Google Scholar] [CrossRef] [Green Version]
- Meadows, J.R.; Cemal, I.; Karaca, O.; Gootwine, E.; Kijas, J.W. Five ovine mitochondrial lineages identified from sheep breeds of the near east. Genetics 2007, 175, 1371–1379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hiendleder, S.; Kaupe, B.; Wassmuth, R.; Janke, A. Molecular analysis of wild and domestic sheep questions current nomenclature and provides evidence for domestication from two different subspecies. Proc. R. Soc. B Biol. Sci. 2002, 269, 893–904. [Google Scholar] [CrossRef] [Green Version]
- Hiendleder, S.; Lewalski, H.; Wassmuth, R.; Janke, A. The complete mitochondrial DNA sequence of the domestic sheep (Ovis aries) and comparison with the other major ovine haplotype. J. Mol. Evol. 1998, 47, 441–448. [Google Scholar] [CrossRef]
- Chessa, B.; Pereira, F.; Arnaud, F.; Amorim, A.; Goyache, F.; Mainland, I.; Kao, R.R.; Pemberton, J.M.; Beraldi, D.; Stear, M.J.; et al. Revealing the history of sheep domestication using retrovirus integrations. Science 2009, 324, 532–536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zilhão, J. Radiocarbon evidence for maritime pioneer colonization at the origins of farming in west Mediterranean Europe. Proc. Natl. Acad. Sci. USA 2001, 98, 14180–14185. [Google Scholar] [CrossRef] [Green Version]
- Ryder, M. Sheep. In Evolution of Domesticated Animals; Mason, I.L., Ed.; Longman: London, UK; New York, NY, USA, 1984; Volume 1, pp. 63–85. [Google Scholar]
- Niemi, M.; Bläuer, A.; Iso-Touru, T.; Nyström, V.; Harjula, J.; Taavitsainen, J.P.; Storå, J.; Lidén, K.; Kantanen, J. Mitochondrial DNA and Y-chromosomal diversity in ancient populations of domestic sheep (Ovis aries) in Finland: Comparison with contemporary sheep breeds. Genet. Sel. Evol. 2013, 45, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tapio, M.; Marzanov, N.; Ozerov, M.; Ćinkulov, M.; Gonzarenko, G.; Kiselyova, T.; Murawski, M.; Viinalass, H.; Kantanen, J. Sheep mitochondrial DNA variation in European, Caucasian, and Central Asian areas. Mol. Biol. Evol. 2006, 23, 1776–1783. [Google Scholar] [CrossRef] [PubMed]
- Ćinkulov, M.; Popovski, Z.; Porcu, K.; Tanaskovska, B.; Hodžić, A.; Bytyqi, H.; Mehmeti, H.; Margeta, V.; Djedović, R.; Hoda, A.; et al. Genetic diversity and structure of the West Balkan Pramenka sheep types as revealed by microsatellite and mitochondrial DNA analysis. J. Anim. Breed. Genet. 2008, 215, 417–426. [Google Scholar] [CrossRef]
- Tapio, M.; Ozerov, M.; Tapio, I.; Toro, M.A.; Marzanov, N.; Ćinkulov, M.; Goncharenko, G.; Kiselyova, T.; Murawski, M.; Kantanen, J. Microsatellite-based genetic diversity and population structure of domestic sheep in northern Eurasia. BMC Genet. 2010, 11, 76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Handley, L.L.; Byrne, K.; Santucci, F.; Townsend, S.; Taylor, M.; Bruford, M.W.; Hewitt, G.M. Genetic structure of European sheep breeds. Heredity 2007, 99, 620–631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pariset, L.; Mariotti, M.; Gargani, M.; Joost, S.; Negrini, R.; Perez, T.; Bruford, M.; Ajmone Marsan, P.; Valentini, A. Genetic diversity of sheep breeds from Albania, Greece, and Italy assessed by mitochondrial DNA and nuclear polymorphisms (SNPs). Sci. World J. 2011, 11, 41–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peter, C.; Bruford, M.; Perez, T.; Dalamitra, S.; Hewitt, G.; Erhardt, G. Genetic diversity and subdivision of 57 European and Middle-Eastern sheep breeds. Anim. Genet. 2007, 38, 37–44. [Google Scholar] [CrossRef]
- Ciani, E.; Mastrangelo, S.; Da Silva, A.; Marroni, F.; Ferenčaković, M.; Ajmone-Marsan, P.; Baird, H.; Barbato, M.; Colli, L.; Delvento, C.; et al. On the origin of European sheep as revealed by the diversity of the Balkan breeds and by optimizing population-genetic analysis tools. Genet. Sel. Evol. 2020, 52, 1–14. [Google Scholar] [CrossRef]
- Bruford, M.W.; Bradley, D.G.; Luikart, G. DNA markers reveal the complexity of livestock domestication. Nat. Rev. Genet. 2003, 4, 900–910. [Google Scholar] [CrossRef] [PubMed]
- Taberlet, P.; Valentini, A.; Rezaei, H.R.; Naderi, S.; Pompanon, F.; Negrini, R.; Ajmone-Marsan, P. Are cattle, sheep, and goats endangered species? Mol. Ecol. 2008, 17, 275–284. [Google Scholar] [CrossRef]
- Chen, S.Y.; Duan, Z.Y.; Sha, T.; Xiangyu, J.; Wu, S.F.; Zhang, Y.P. Origin, genetic diversity, and population structure of Chinese domestic sheep. Gene 2006, 376, 216–223. [Google Scholar] [CrossRef] [PubMed]
- Zhao, E.; Yu, Q.; Zhang, N.; Kong, D.; Zhao, Y. Mitochondrial DNA diversity and the origin of Chinese indigenous sheep. Trop. Anim. Health Prod. 2013, 45, 1715–1722. [Google Scholar] [CrossRef] [PubMed]
- Kijas, J.W.; Lenstra, J.A.; Hayes, B.; Boitard, S.; Neto, L.R.P.; San Cristobal, M.; Servin, B.; McCulloch, R.; Whan, V.; Gietzen, K.; et al. Genome-wide analysis of the world’s sheep breeds reveals high levels of historic mixture and strong recent selection. PLoS Biol. 2012, 10, e1001258. [Google Scholar] [CrossRef] [Green Version]
- Meadows, J.R.S.; Li, K.; Kantanen, J.; Tapio, M.; Sipos, W.; Pardeshi, V.; Gupta, V.; Calvo, J.H.; Whan, V.; Norris, B.; et al. Mitochondrial sequence reveals high levels of gene flow between breeds of domestic sheep from Asia and Europe. J. Hered. 2005, 96, 494–501. [Google Scholar] [CrossRef] [Green Version]
- Niemi, M.; Sajantila, A.; Ahola, V.; Vilkki, J. Sheep and cattle population dynamics based on ancient and modern DNA reflects key events in the human history of the North-East Baltic Sea Region. J. Archaeol. Sci. Rep. 2018, 18, 169–173. [Google Scholar] [CrossRef]
- Olivieri, C.; Ermini, L.; Rizzi, E.; Corti, G.; Luciani, S.; Marota, I.; De Bellis, G.; Rollo, F. Phylogenetic position of a copper age sheep (Ovis aries) mitochondrial DNA. PLoS ONE 2012, 7, e33792. [Google Scholar] [CrossRef] [Green Version]
- Cai, D.; Tang, Z.; Yu, H.; Han, L.; Ren, X.; Zhao, X.; Zhu, H.; Zhou, H. Early history of Chinese domestic sheep indicated by ancient DNA analysis of Bronze Age individuals. J. Archaeol. Sci. 2011, 38, 896–902. [Google Scholar] [CrossRef]
- Cai, D.W.; Han, L.; Zhang, X.L.; Zhou, H.; Zhu, H. DNA analysis of archaeological sheep remains from China. J. Archaeol. Sci. 2007, 34, 1347–1355. [Google Scholar] [CrossRef]
- Demirci, S.; Baştanlar, E.K.; Dağtaş, N.D.; Pişkin, E.; Engin, A.; Özer, F.; Yüncü, E.; Doğan, Ş.A.; Togan, I. Mitochondrial DNA diversity of modern, ancient and wild sheep (Ovis gmelinii anatolica) from Turkey: New insights on the evolutionary history of sheep. PLoS ONE 2013, 8, e81952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dymova, M.A.; Zadorozhny, A.V.; Mishukova, O.V.; Khrapov, E.A.; Druzhkova, A.S.; Trifonov, V.A.; Kichigin, I.G.; Tishkin, A.A.; Grushin, S.P.; Filipenko, M.L. Mitochondrial DNA analysis of ancient sheep from Altai. Anim Genet. 2017, 48, 615–618. [Google Scholar] [CrossRef] [PubMed]
- Rannamäe, E.; Lõugas, L.; Speller, C.F.; Valk, H.; Maldre, L.; Wilczyński, J.; Mikhailov, A.; Saarma, U. Three thousand years of continuity in the maternal lineages of ancient sheep (Ovis aries) in Estonia. PLoS ONE 2016, 11, e0163676. [Google Scholar] [CrossRef]
- Rannamäe, E.; Lõugas, L.; Niemi, M.; Kantanen, J.; Maldre, L.; Kadõrova, N.; Saarma, U. Maternal and paternal genetic diversity of ancient sheep in Estonia from the Late Bronze Age to the post-medieval period and comparison with other regions in Eurasia. Anim. Genet. 2015, 47, 208–218. [Google Scholar] [CrossRef] [PubMed]
- Gabbianelli, F.; Gargani, M.; Pariset, L.; Mariotti, M.; Alhaique, F.; De Minicis, E.; Barelli, L.; Ciammetti, E.; Redi, F.; Valentini, A. Mitochondrial DNA analysis of medieval sheep (Ovis aries) in central Italy reveals the predominance of haplogroup B already in the Middle Ages. Anim. Genet. 2015, 46, 329–332. [Google Scholar] [CrossRef]
- Curta, F.; Stephenson, P. Southeastern Europe in the Middle Ages 500–1250; Cambridge University Press: Cambridge, UK, 2006. [Google Scholar]
- Rusu, I.; Modi, A.; Vai, S.; Pilli, E.; Mircea, C.; Radu, C.; Urduzia, C.; Pinter, Z.K.; Bodolică, V.; Dobrinescu, C.; et al. Maternal DNA lineages at the gate of Europe in the 10th century AD. PLoS ONE 2018, 13, e0193578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinter, Z.K.; Dobrinescu, C.I.; Dragotă, A.; Kelemen, B. Preliminary Research in Capidava Medieval Necropolis (Topalu com., Constanţa County). Pontica 2011, 44, 387–400. [Google Scholar]
- Harvey, V.L.; Egerton, V.M.; Chamberlain, A.T.; Manning, P.L.; Buckley, M. Collagen Fingerprinting: A new screening technique for radiocarbon dating ancient bone. PLoS ONE 2016, 11, e0150650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brock, F.; Geoghegan, V.; Thomas, B.; Jurkschat, K.; Higham, T.F.G. Analysis of bone “collagen” extraction products for radiocarbon dating. Radiocarbon 2013, 55, 445–463. [Google Scholar] [CrossRef] [Green Version]
- Yizhaq, M.; Mintz, G.; Cohen, I.; Khalaily, H.; Weiner, S.; Boaretto, E. Quality controlled radiocarbon dating of bones and charcoal from the early pre-pottery neolithic β (ppnb) of Motza (Israel). Radiocarbon 2005, 47, 193–206. [Google Scholar] [CrossRef] [Green Version]
- Stan-Sion, C.; Enachescu, M.; Ghita, D.G.; Calinescu, C.I.; Petre, A.; Mosu, D.V.; Klein, M. A new AMS facility based on a Cockcroft-Walton type 1 MV tandetron at IFIN-HH Magurele, Romania. Nuclear Instrum. Methods Phys. Res. Sect. B Beam Interact. Mater. Atoms. 2014, 319, 117–122. [Google Scholar] [CrossRef]
- Wacker, L.; Němec, M.; Bourquin, J. A revolutionary graphitisation system: Fully automated, compact and simple. Nuclear Instrum. Methods Phys. Res. Sect. B Beam Interact. Mater. Atoms. 2010, 268, 931–934. [Google Scholar] [CrossRef]
- 4990C NS. International Standard Reference Material for Contemporary Carbon-14, NIST: USA.
- Wacker, L.; Christl, M.; Synal, H.A. Bats: A new tool for AMS data reduction. Nuclear Instrum. Methods Phys. Res. B. 2010, 268, 976–979. [Google Scholar] [CrossRef]
- Bronk Ramsey, C. Analysis of chronological information and radiocarbon calibration: The program OxCal. Archaeol. Comput. Newsl. 1994, 41, 11–16. [Google Scholar]
- Reimer, P.J.; Austin, W.E.; Bard, E.; Bayliss, A.; Blackwell, P.G.; Ramsey, C.B.; Butzin, M.; Cheng, H.; Edwards, R.L.; Friedrich, M.; et al. The IntCal20 Northern Hemisphere Radiocarbon Age Calibration Curve (0–55 cal kBP). Radiocarbon 2020, 62, 725–757. [Google Scholar] [CrossRef]
- Yang, D.Y.; Eng, B.; Waye, J.S.; Dudar, J.C.; Saunders, S.R. Technical Note: Improved DNA Extraction from Ancient Bones Using Silica-Based Spin Columns. Am. J. Phys. Anthropol. 1998, 105, 539–543. [Google Scholar] [CrossRef]
- Hervella, M.; Izagirre, N.; Alonso, S.; Fregel, R.; Alonso, A.; Cabrera, V.M.; de la Rúa, C. Ancient DNA from hunter-gatherer and farmer groups from Northern Spain supports a random dispersion model for the Neolithic expansion into Europe. PLoS ONE 2012, 7, e34417. [Google Scholar] [CrossRef]
- Rohland, N.; Glocke, I.; Aximu-Petri, A.; Meyer, M. Extraction of highly degraded DNA from ancient bones, teeth and sediments for high-throughput sequencing. Nat. Protoc. 2018, 13, 2447–2461. [Google Scholar] [CrossRef]
- Dabney, J.; Knapp, M.; Glocke, I.; Gansauge, M.T.; Weihmann, A.; Nickel, B.; Valdiosera, C.; García, N.; Pääbo, S.; Arsuaga, J.L.; et al. Complete mitochondrial genome sequence of a Middle Pleistocene cave bear reconstructed from ultrashort DNA fragments. Proc. Natl. Acad. Sci. USA 2013, 110, 15758–15763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kusza, S.; Zakar, E.; Budai, C.; Cziszter, L.T.; Padeanu, I.; Gavojdian, D. Mitochondrial DNA variability in Gyimesi Racka and Turcana sheep breeds. Acta Biochim. Pol. 2015, 62, 273–280. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Du, L.X.; Ma, Y.H.; Guan, W.J.; Li, H.B.; Zhao, Q.J.; Li, X.; Rao, S.Q. A novel maternal lineage revealed in sheep (Ovis aries). Anim. Genet. 2005, 36, 331–336. [Google Scholar] [CrossRef]
- Meadows, J.R.S.; Hiendleder, S.; Kijas, J.W. Haplogroup relationships between domestic and wild sheep resolved using a mitogenome panel. Heredity 2011, 106, 700–706. [Google Scholar] [CrossRef] [Green Version]
- Pedrosa, S.; Uzun, M.; Arranz, J.J.; Gutiérrez-Gil, B.; San Primitivo, F.; Bayón, Y. Evidence of three maternal lineages in near eastern sheep supporting multiple domestication events. Proc. R. Soc. B Biol. Sci. 2005, 272, 2211–2217. [Google Scholar] [CrossRef] [Green Version]
- Bandelt, H.J.; Forster, P.; Röhl, A. Median-joining networks for inferring intraspecific phylogenies. Mol. Biol. Evol. 1999, 16, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Leigh, J.; Bryant, D. PopART: Full-feature software for haplotype network construction. Methods Ecol. Evol. 2015, 6, 1110–1116. [Google Scholar] [CrossRef]
- Excoffier, L.; Lischer, H.E. Arlequin suite ver 3.5: A new series of programs to perform population genetics analyses under Linux and Windows. Mol. Ecol. Resour. 2010, 10, 564–567. [Google Scholar] [CrossRef]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D.J. jModelTest 2: More models, new heuristics and high-performance computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [Green Version]
- Guindon, S.; Gascuel, O. A Simple, Fast, and Accurate Algorithm to Estimate Large Phylogenies by Maximum Likelihood. Syst. Biol. 2003, 52, 696–704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef]
- R.C. Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Association. Q. QGIS Geographic Information System. QGIS.org. 2020. Available online: http://www.qgis.org (accessed on 15 December 2020).
- Fulton, T.L. Setting up an Ancient DNA Laboratory. In Ancient DNA Methods and Protocols; Shapiro, B., Hofreiter, M., Eds.; Humana Press: Totowa, NJ, USA, 2009. [Google Scholar]
- Oner, Y.; Calvo, J.H.; Elmaci, C. Investigation of the genetic diversity among native Turkish sheep breeds using mtDNA polymorphisms. Trop. Anim. Health Prod. 2013, 45, 947–951. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.Z.; Batsuuri, L.; Badamdorj, D.; Olivier, H.; Han, J.L. Origin and genetic diversity of Mongolian and Chinese sheep using mitochondrial DNA D-loop sequences. Acta Genet. Sin. 2005, 32, 1256–1265. [Google Scholar] [PubMed]
- Zhao, Y.; Zhao, E.; Zhang, N.; Duan, C. Mitochondrial DNA diversity, origin, and phylogenic relationships of three Chinese large-fat-tailed sheep breeds. Trop. Anim. Health Prod. 2011, 43, 1405–1410. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Age (Cal. AD) | Amplified Fragment Length | Variable Position | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 15993 | 16042 | 16048 | 16128 | 16208 | 16293 | 16343 | 16343.1 | 16347 | 16440 | 16472 | 16484 | 16496 | ||||
| NC_001941 | C | T | T | C | A | A | T | - | C | T | T | C | T | |||
| Ovis_Cap_1 | 771–899 AD | 598 bp | T | C | T | T | A | A | C | C | C | T | - | C | T | |
| Ovis_Cap_2 | 418–550 AD | 529 bp | T | T | T | G | G | C | C | C | T | - | T | T | ||
| Ovis_Cap_3 | 424–564 AD | |||||||||||||||
| Ovis_Cap_4 | 771–885 AD | 529 bp | T | T | T | A | A | C | C | T | T | - | C | T | ||
| Ovis_Cap_5 | 870–991 AD | 529 bp | T | T | T | G | A | C | C | C | T | - | C | T | ||
| Ovis_Cap_6 | 771–884 AD | 529 bp | T | C | T | A | A | C | C | C | C | - | C | C | ||
| Ovis_Cap_7 | 271–351 AD | |||||||||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Acatrinei, A.; Rusu, I.; Mircea, C.; Zagrean-Tuza, C.; Gál, E.; Păceșilă, D.; Gâza, O.; Urduzia, C.; Pinter, Z.K.; Dobrinescu, C.; et al. Shedding Light on the Dark Ages: Sketching Potential Trade Relationships in Early Medieval Romania through Mitochondrial DNA Analysis of Sheep Remains. Diversity 2021, 13, 208. https://doi.org/10.3390/d13050208
Acatrinei A, Rusu I, Mircea C, Zagrean-Tuza C, Gál E, Păceșilă D, Gâza O, Urduzia C, Pinter ZK, Dobrinescu C, et al. Shedding Light on the Dark Ages: Sketching Potential Trade Relationships in Early Medieval Romania through Mitochondrial DNA Analysis of Sheep Remains. Diversity. 2021; 13(5):208. https://doi.org/10.3390/d13050208
Chicago/Turabian StyleAcatrinei, Arina, Ioana Rusu, Cristina Mircea, Cezara Zagrean-Tuza, Emese Gál, Doru Păceșilă, Oana Gâza, Claudia Urduzia, Zeno Karl Pinter, Cătălin Dobrinescu, and et al. 2021. "Shedding Light on the Dark Ages: Sketching Potential Trade Relationships in Early Medieval Romania through Mitochondrial DNA Analysis of Sheep Remains" Diversity 13, no. 5: 208. https://doi.org/10.3390/d13050208