
The Diversity and Community Assembly Process of Wetland Plants from Lakeshores on the Qinghai-Tibetan Plateau

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
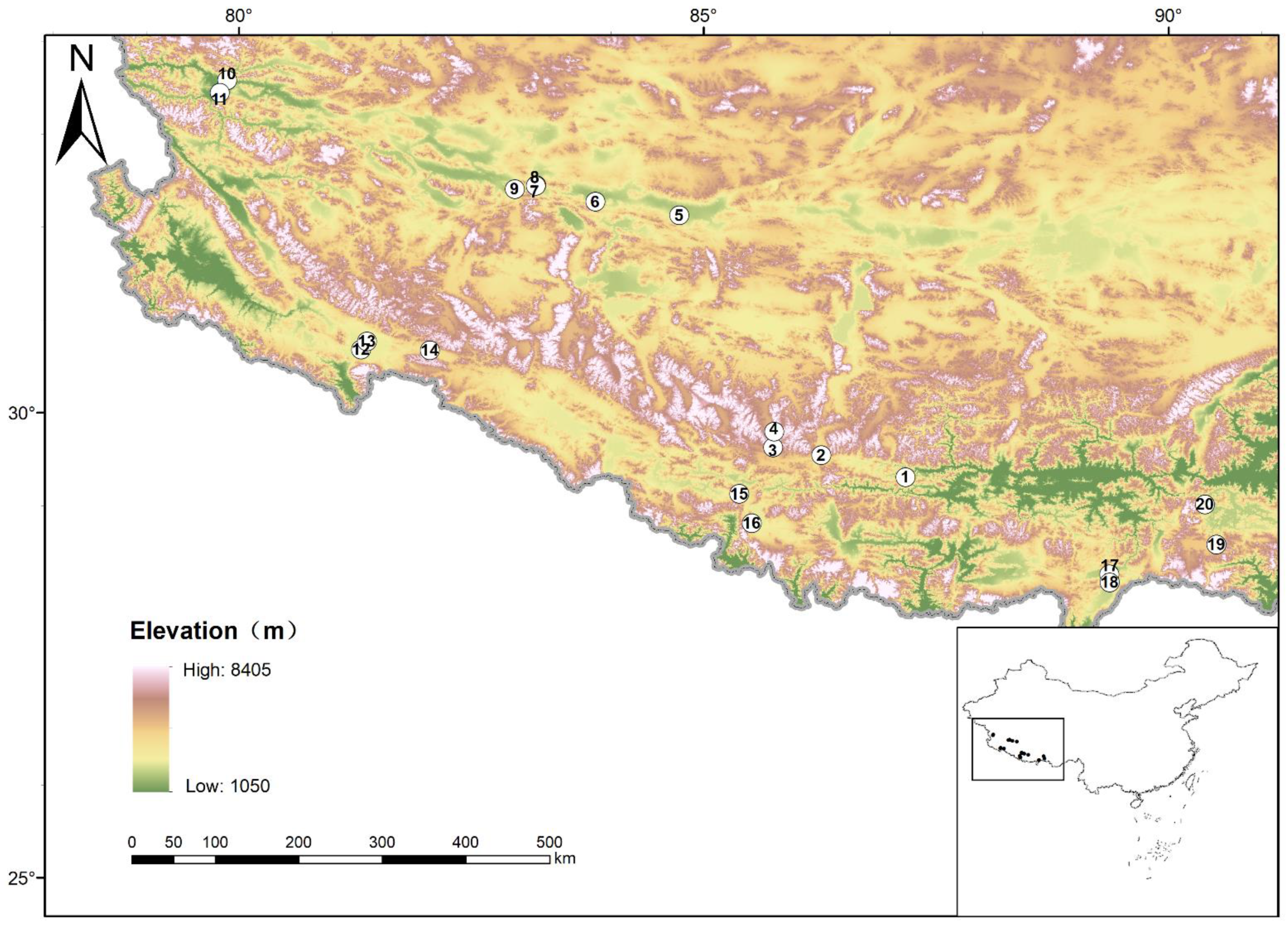
2.1. Vegetation Survey and Sample Collection
2.2. Measurements of Environmental Factors
2.3. Data Analysis
3. Results
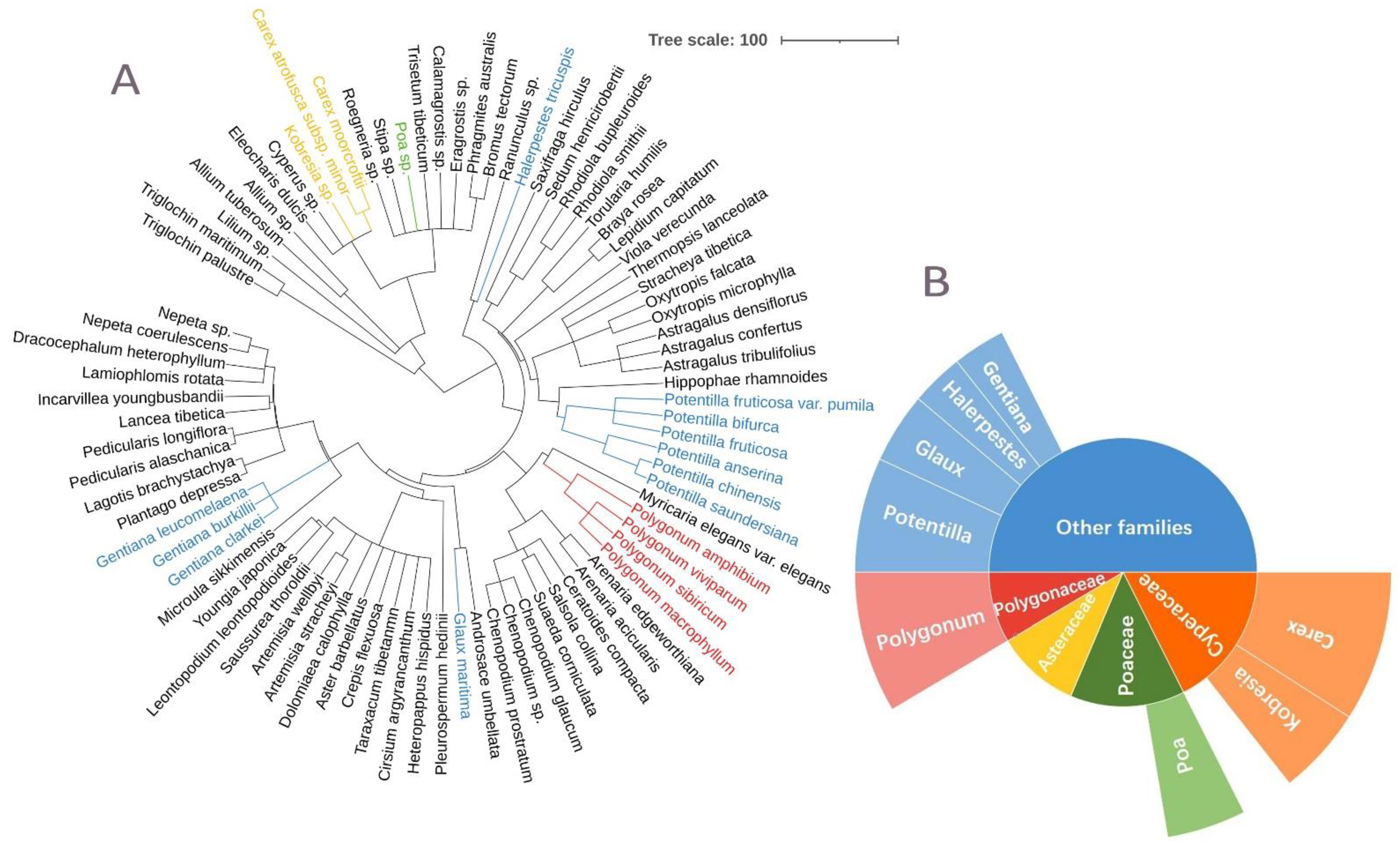
3.1. Wetland Plants of the Lakeshore across the QTP
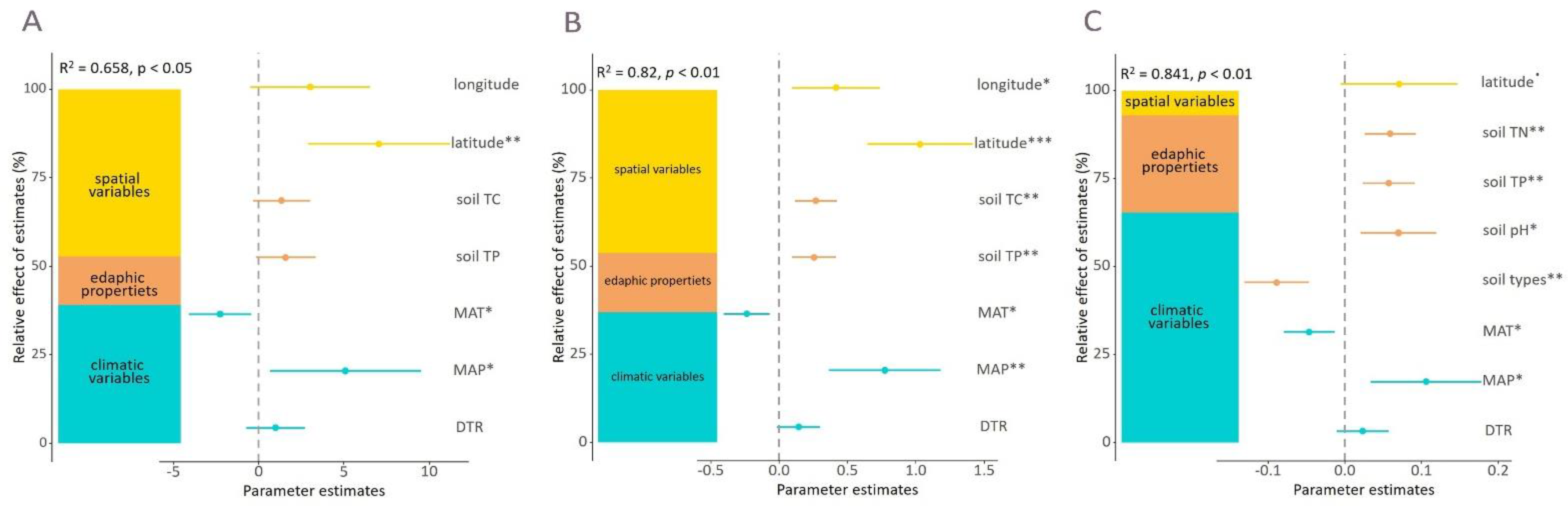
3.2. Geospatial and Environmental Drivers of the Taxonomic and Phylogenetic Alpha Diversity
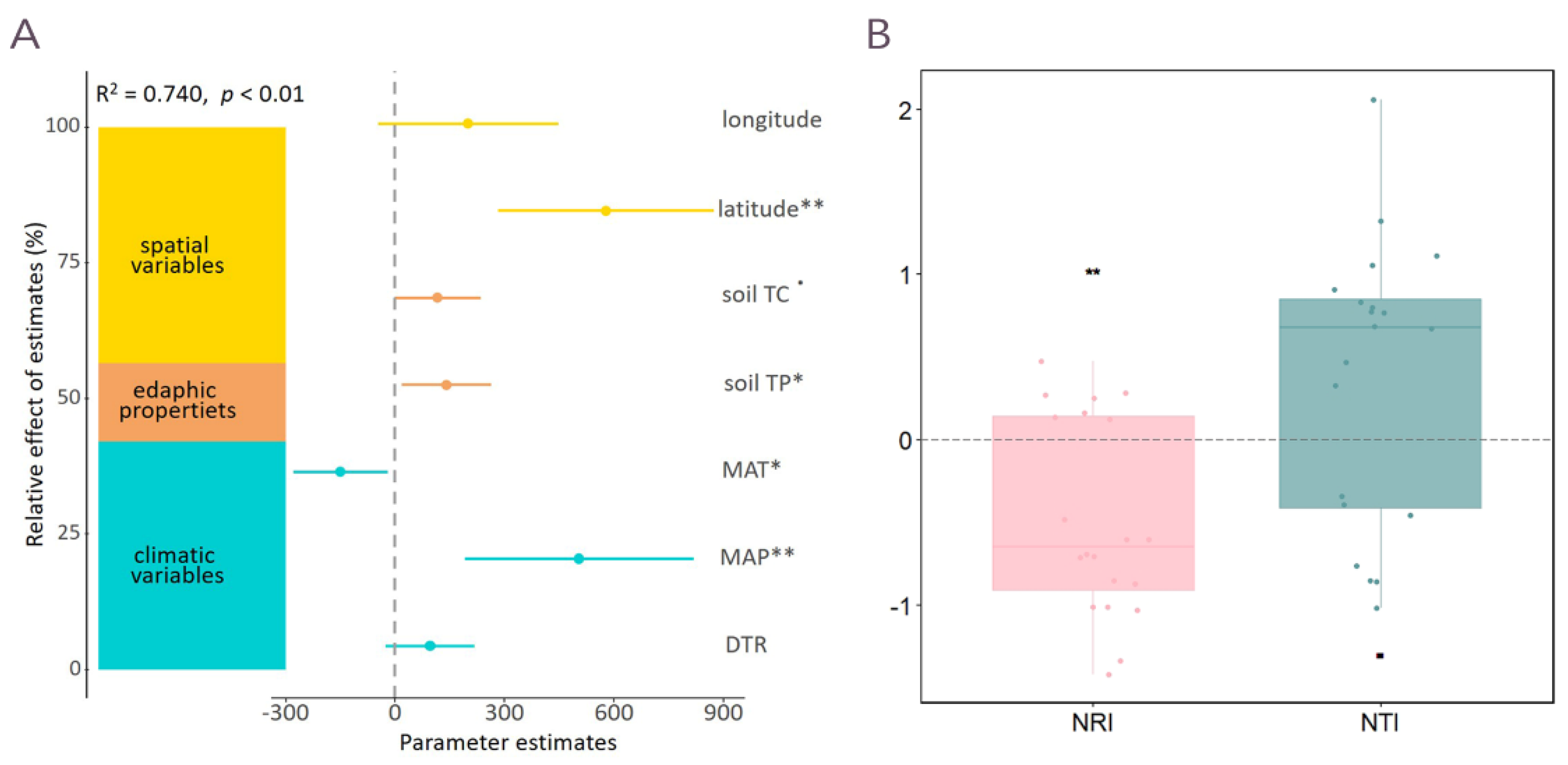
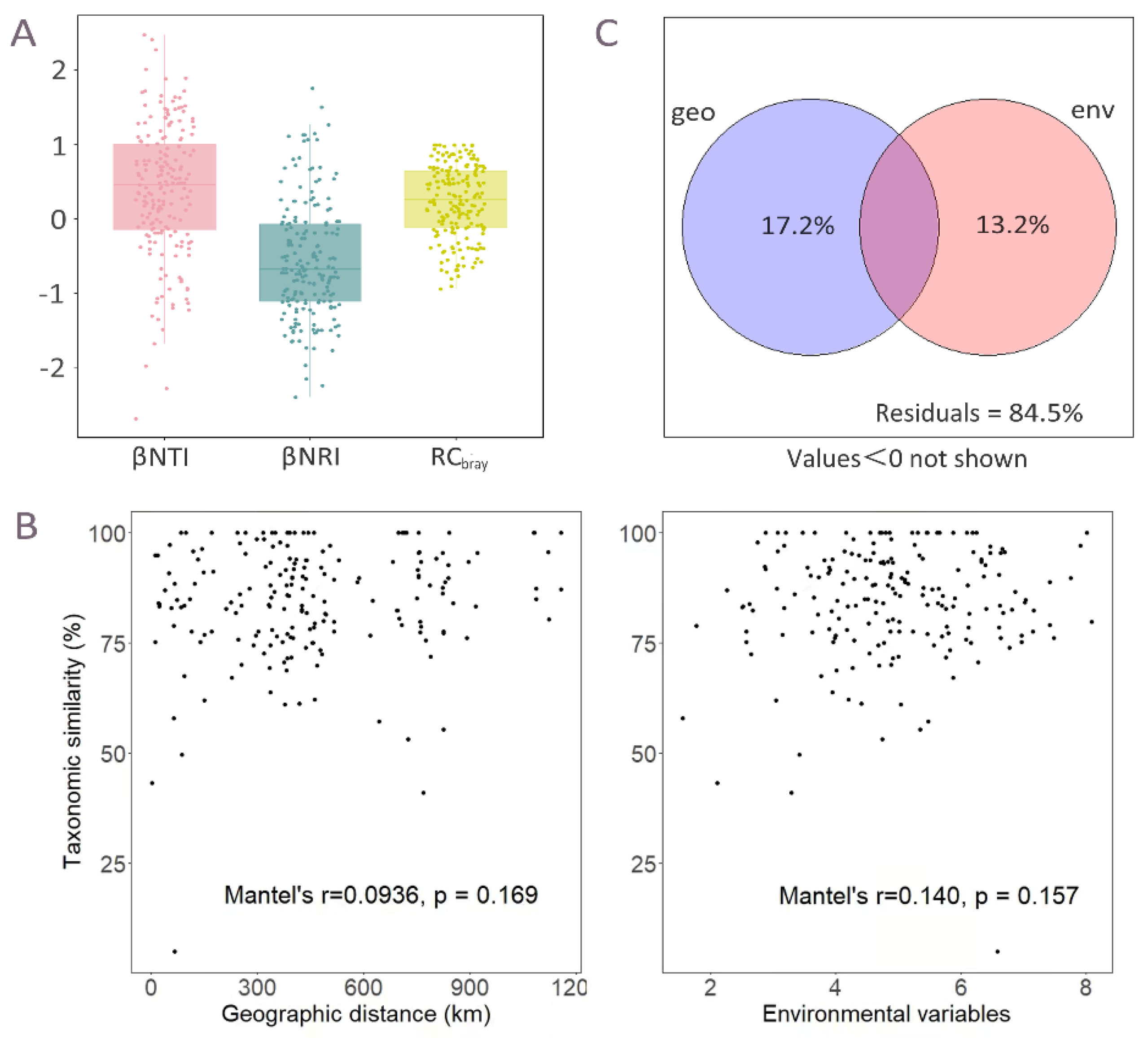
3.3. Potential Community Assembly Process of the Wetland Plants
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gaston, K.J. Global patterns in biodiversity. Nature 2000, 405, 220–227. [Google Scholar] [CrossRef]
- Lyashevska, O.; Farnsworth, K.D. How many dimensions of biodiversity do we need? Ecol. Indic. 2012, 18, 485–492. [Google Scholar] [CrossRef]
- Cardinale, B.J.; Duffy, J.E.; Gonzalez, A.; Hooper, D.U.; Perrings, C.; Venail, P.; Narwani, A.; Mace, G.M.; Tilman, D.; Wardle, D.A.; et al. Biodiversity loss and its impact on humanity. Nature 2012, 486, 59–67. [Google Scholar] [CrossRef]
- Pavoine, S.; Bonsall, M. Measuring biodiversity to explain community assembly: A unified approach. Biol. Rev. 2011, 86, 792–812. [Google Scholar] [CrossRef] [PubMed]
- Swenson, N.G. The role of evolutionary processes in producing biodiversity patterns, and the interrelationships between taxonomic, functional and phylogenetic biodiversity. Am. J. Bot. 2011, 98, 472–480. [Google Scholar] [CrossRef] [Green Version]
- Srivastava, D.S.; Cadotte, M.W.; MacDonald, A.A.M.; Marushia, R.G.; Mirotchnick, N. Phylogenetic diversity and the functioning of ecosystems. Ecol. Lett. 2012, 15, 637–648. [Google Scholar] [CrossRef] [PubMed]
- Peralta, G.; Frost, C.M.; Didham, R.K.; Varsani, A.; Tylianakis, J. Phylogenetic diversity and co-evolutionary signals among trophic levels change across a habitat edge. J. Anim. Ecol. 2015, 84, 364–372. [Google Scholar] [CrossRef]
- Rands, M.R.W.; Adams, W.M.; Bennun, L.; Butchart, S.H.M.; Clements, A.; Coomes, D.; Entwistle, A.; Hodge, I.; Kapos, V.; Scharlemann, J.P.W.; et al. Biodiversity conservation: Challenges beyond 2010. Science 2010, 329, 1298–1303. [Google Scholar] [CrossRef] [Green Version]
- Urban, M.C.; Leibold, M.A.; Amarasekare, P.; De Meester, L.; Gomulkiewicz, R.; Hochberg, M.E.; Klausmeier, C.A.; Loeuille, N.; de Mazancourt, C.; Norberg, J. The evolutionary ecology of metacommunities. Trends Ecol. Evol. 2008, 23, 311–317. [Google Scholar] [CrossRef] [PubMed]
- Vanoverbeke, J.; Urban, M.C.; De Meester, L. Community assembly is a race between immigration and adaptation: Eco-evolutionary interactions across spatial scales. Ecography 2015, 39, 858–870. [Google Scholar] [CrossRef]
- Hanson, C.A.; Fuhrman, J.; Horner-Devine, C.; Martiny, J. Beyond biogeographic patterns: Processes shaping the microbial landscape. Nat. Rev. Microbiol. 2012, 10, 497–506. [Google Scholar] [CrossRef] [PubMed]
- Vellend, M. Conceptual Synthesis in Community Ecology. Q. Rev. Biol. 2010, 85, 183–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, J.; Ning, D. Stochastic community assembly: Does it matter in microbial ecology? Microbiol. Mol. Biol. Rev. 2017, 81, e00002–e00017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ellwood, M.D.; Manica, A.; Foster, W.A. Stochastic and deterministic processes jointly structure tropical arthropod communities. Ecol. Lett. 2009, 12, 277–284. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Guo, H.; Zhang, J.; Du, G. Stochastic and deterministic processes together determine alpine meadow plant community composition on the Tibetan Plateau. Oecologia 2013, 171, 495–504. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Jiang, X.; Ouyang, J.; Lu, B.; Liu, W.; Liu, G. Environmental factors, more than spatial distance, explain community structure of soil ammonia-oxidizers in wetlands on the Qinghai—Tibetan Plateau. Microorganisms 2020, 8, 933–949. [Google Scholar]
- Möls, T.; Vellak, K.; Vellak, A.; Ingerpuu, N. Global gradients in moss and vascular plant diversity. Biodivers. Conserv. 2013, 22, 1537–1551. [Google Scholar] [CrossRef]
- Qian, H.; Song, J.-S.; Krestov, P.; Guo, Q.; Wu, Z.; Shen, X.; Guo, X. Large-scale phytogeographical patterns in East Asia in relation to latitudinal and climatic gradients. J. Biogeogr. 2003, 30, 129–141. [Google Scholar] [CrossRef] [Green Version]
- Field, R.; Hawkins, B.A.; Cornell, H.V.; Currie, D.J.; Diniz-Filho, J.A.; Guégan, J.-F.; Kaufman, D.M.; Kerr, J.T.; Mittelbach, G.G.; Oberdorff, T.; et al. Spatial species-richness gradients across scales: A meta-analysis. J. Biogeogr. 2009, 36, 132–147. [Google Scholar] [CrossRef]
- Hawkins, B.A.; Field, R.; Cornell, H.V.; Currie, D.J.; Guégan, J.-F.; Kaufman, D.M.; Kerr, J.T.; Mittelbach, G.G.; Oberdorff, T.; O’Brien, E.M.; et al. Energy, water, and broad-scale geographic patterns of species richness. Ecology 2003, 84, 3105–3117. [Google Scholar] [CrossRef] [Green Version]
- Baudena, M.; Sánchez, A.; Georg, C.-P.; Ruiz-Benito, P.; Rodríguez, M.Á.; Zavala, M.A.; Rietkerk, M. Revealing patterns of local species richness along environmental gradients with a novel network tool. Sci. Rep. 2015, 5, 11561–11575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whittaker, R.J.; Nogués-Bravo, D.; Araújo, M.B. Geographical gradients of species richness: A test of the water-energy conjecture of Hawkins et al. (2003) using European data for five taxa. Glob. Ecol. Biogeogr. 2007, 16, 76–89. [Google Scholar] [CrossRef]
- Allen, P.A.; Brown, J.H.; Gillooly, J.F. Global biodiversity, biochemical kinetics, and the energetic-equivalence rule. Science 2002, 297, 1545–1548. [Google Scholar] [CrossRef]
- Kreft, H.; Jetz, W. Global patterns and determinants of vascular plant diversity. Proc. Natl. Acad. Sci. USA 2007, 104, 5925–5930. [Google Scholar] [CrossRef] [Green Version]
- Chappuis, E.; Ballesteros, E.; Gacia, E. Distribution and richness of aquatic plants across Europe and Mediterranean countries: Patterns, environmental driving factors and comparison with total plant richness. J. Veg. Sci. 2012, 23, 985–997. [Google Scholar] [CrossRef]
- Crow, G.E. Species diversity in aquatic angiosperms: Latitudinal patterns. Aquat. Bot. 1993, 44, 229–258. [Google Scholar] [CrossRef]
- Yao, T.; Thompson, L.; Yang, W.; Yu, W.; Gao, Y.; Guo, X.; Yang, X.; Duan, K.; Zhao, H.; Xu, B.; et al. Different glacier status with atmospheric circulations in Tibetan Plateau and surroundings. Nat. Clim. Chang. 2012, 2, 663–667. [Google Scholar] [CrossRef]
- Yan, Y.; Yang, X.; Tang, Z. Patterns of species diversity and phylogenetic structure of vascular plants on the Qinghai-Tibetan Plateau. Ecol. Evol. 2013, 3, 4584–4595. [Google Scholar] [CrossRef] [PubMed]
- Ma, W.H.; He, J.S.; Yang, Y.H.; Wang, X.P.; Liang, C.Z.; Anwar, M.; Zeng, H.; Fang, J.Y.; Schmid, B. Environmental factors covary with plant diversity-productivity relationships among Chinese grassland sites. Glob. Ecol. Biogeogr. 2010, 19, 233–243. [Google Scholar] [CrossRef]
- Hijmans, R.J.; Cameron, S.E.; Parra, J.L.; Jones, P.G.; Jarvis, A. Very high resolution interpolated climate surfaces for global land areas. Int. J. Climatol. 2005, 25, 1965–1978. [Google Scholar] [CrossRef]
- The Angiosperm Phylogeny Group. An update of the angiosperm phylogeny group classification for the orders and families of flowering plants: APG iii. Bot. J. Linn. Soc. 2009, 161, 105–121. [Google Scholar] [CrossRef] [Green Version]
- Webb, C.O.; Ackerly, D.D.; Kembel, S.W. Phylocom: Software for the analysis of phylogenetic community structure and trait evolution. Bioinformatics 2008, 24, 2098–2100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zanne, A.E.; Tank, D.C.; Cornwell, W.; Eastman, J.M.; Smith, S.; Fitzjohn, R.G.; McGlinn, D.J.; O’Meara, B.; Moles, A.; Reich, P.; et al. Three keys to the radiation of angiosperms into freezing environments. Nature 2014, 506, 89–92. [Google Scholar] [CrossRef] [PubMed]
- Gorelick, R. Combining richness and abundance into a single diversity index using matrix analogues of Shannon’s and Simpson’s indices. Ecography 2006, 29, 525–530. [Google Scholar] [CrossRef]
- Faith, D.P. Conservation evaluation and phylogenetic diversity. Biol. Conserv. 1992, 61, 1–10. [Google Scholar] [CrossRef]
- Kling, M.M.; Mishler, B.D.; Thornhill, A.; Baldwin, B.G.; Ackerly, D. Facets of phylodiversity: Evolutionary diversification, divergence and survival as conservation targets. Philos. Trans. R. Soc. B Biol. Sci. 2019, 374, 20170397. [Google Scholar] [CrossRef] [Green Version]
- Webb, C.O.; Ackerly, D.D.; McPeek, M.A.; Donoghue, M.J. Phylogenies and Community Ecology. Annu. Rev. Ecol. Syst. 2002, 33, 475–505. [Google Scholar] [CrossRef] [Green Version]
- Cavender-Bares, J.; Ackerly, D.D.; Baum, D.A.; Bazzaz, F.A. Phylogenetic Overdispersion in Floridian Oak Communities. Am. Nat. 2004, 163, 823–843. [Google Scholar] [CrossRef] [PubMed]
- Brunbjerg, A.K.; Cavender-Bares, J.; Eiserhardt, W.; Ejrnæs, R.; Aarssen, L.W.; Buckley, H.; Forey, E.; Jansen, F.; Kattge, J.; Lane, C.; et al. Multi-scale phylogenetic structure in coastal dune plant communities across the globe. J. Plant Ecol. 2014, 7, 101–114. [Google Scholar] [CrossRef]
- Huang, M.K.; Chai, L.W.; Jiang, D.L.; Zhang, M.J.; Jia, W.Q.; Huang, Y.; Zhou, J.Z. Dissovled organic matter (DOM) quality drives biogeographic patterns of soil bacterial communities and their association networks in semi-arid regions. FEMS Microbiol. Ecol. 2021, 97, fiab083. [Google Scholar] [CrossRef]
- Stegen, J.; Lin, X.; Fredrickson, J.K.; Chen, X.; Kennedy, D.; Murray, C.J.; Rockhold, M.L.; Konopka, A. Quantifying community assembly processes and identifying features that impose them. ISME J. 2013, 7, 2069–2079. [Google Scholar] [CrossRef]
- Stegen, J.C.; Lin, X.; Fredrickson, J.K.; Konopka, A.E. Estimating and mapping ecological processes influencing microbial community assembly. Front. Microbiol. 2015, 6, 370–384. [Google Scholar] [CrossRef] [Green Version]
- Feng, Y.; Chen, R.; Stegen, J.C.; Guo, Z.; Zhang, J.; Li, Z.; Lin, X. Two key features influencing community assembly processes at regional scale: Initial state and degree of change in environmental conditions. Mol. Ecol. 2018, 27, 5238–5251. [Google Scholar] [CrossRef]
- van Breugel, M.; Craven, D.; Lai, H.R.; Baillon, M.; Turner, B.L.; Hall, J.S. Soil nutrients and dispersal limitation shape compositional variation in secondary tropical forests across multiple scales. J. Ecol. 2019, 107, 566–581. [Google Scholar] [CrossRef]
- Gad, M.; Hou, L.; Li, J.; Wu, Y.; Rashid, A.; Chen, N.; Hu, A. Distinct mechanisms underlying the assembly of microeukaryotic generalists and specialists in an anthropogenically impacted river. Sci. Total Environ. 2020, 748, 141434. [Google Scholar] [CrossRef] [PubMed]
- Shi, W.; Wang, Y.; Xiang, W.; Li, X.; Cao, K. Environmental filtering and dispersal limitation jointly shaped the taxonomic and phylogenetic beta diversity of natural forests in southern China. Ecol. Evol. 2021, 11, 8783–8794. [Google Scholar] [CrossRef] [PubMed]
- Khan, H.; Baig, S. High altitude wetlands of the HKH region of northern Pakistan—Status of current knowledge, challenges and research opportunities. Wetlands 2017, 37, 371–380. [Google Scholar] [CrossRef]
- Cai, H.; Lyu, L.; Shrestha, N.; Tang, Z.; Su, X.; Xu, X.; Dimitrov, D.; Wang, Z. Geographical patterns in phylogenetic diversity of Chinese woody plants and its application for conservation planning. Divers. Distrib. 2021, 27, 179–194. [Google Scholar] [CrossRef]
- Geedicke, I.; Schultz, M.; Rudolph, B.; Oldeland, J. Phylogenetic clustering found in lichen but not in plant communities in European heathlands. Community Ecol. 2016, 17, 216–224. [Google Scholar] [CrossRef] [Green Version]
- Morlon, H.; Schwilk, D.W.; Bryant, J.A.; Marquet, P.A.; Rebelo, A.G.; Tauss, C.; Bohannan, B.J.M.; Green, J.L. Spatial patterns of phylogenetic diversity. Ecol. Lett. 2011, 14, 141–149. [Google Scholar] [CrossRef] [Green Version]
- Tucker, C.M.; Cadotte, M.W. Unifying measures of biodiversity: Understanding when richness and phylogenetic diversity should be congruent. Divers. Distrib. 2013, 19, 845–854. [Google Scholar] [CrossRef]
- Qian, H.; Sandel, B. Phylogenetic relatedness of native and exotic plants along climate gradients in California, USA. Divers. Distrib. 2017, 23, 1323–1333. [Google Scholar] [CrossRef] [Green Version]
- Kraft, N.J.; Cornwell, W.K.; Webb, C.O.; Ackerly, D.D. Trait evolution, community assembly, and the phylogenetic structure of ecological communities. Am. Nat. 2007, 170, 271–283. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.H.; Huang, J.H.; Lu, X.H.; Ma, K.P. Diversity distribution patterns of Chinese endemic seed plant species and their implications for conservation planning. Sci. Rep. 2016, 6, 33913. [Google Scholar] [CrossRef] [Green Version]
- Ricklefs, R.E. Evolutionary diversification and the origin of the diversity—Environment relationship. Ecology 2006, 87, S3–S13. [Google Scholar] [CrossRef]
- Ning, D.; Deng, Y.; Tiedje, J.M.; Zhou, J. A general framework for quantitatively assessing ecological stochasticity. Proc. Natl. Acad. Sci. USA 2019, 116, 16892–16898. [Google Scholar] [CrossRef] [Green Version]
- Jiang, L.; Lv, G.; Gong, Y.; Li, Y.; Wang, H.; Wu, D. Characteristics and driving mechanisms of species beta diversity in desert plant communities. PLoS ONE 2021, 16, e0245249. [Google Scholar] [CrossRef] [PubMed]
- Legendre, P.; Mi, X.; Ren, H.; Ma, K.; Yu, M.; Sun, I.-F.; He, F. Partitioning beta diversity in a subtropical broad-leaved forest of China. Ecology 2009, 90, 663–674. [Google Scholar] [CrossRef] [Green Version]
- Vellend, M.; Srivastava, D.S.; Anderson, K.M.; Brown, C.D.; Jankowski, J.E.; Kleynhans, E.J.; Kraft, N.; Letaw, A.D.; Macdonald, A.A.M.; Maclean, J.E.; et al. Assessing the relative importance of neutral stochasticity in ecological communities. Oikos 2014, 123, 1420–1430. [Google Scholar] [CrossRef]
- Lowe, W.H.; McPeek, M.A. Is dispersal neutral? Trends Ecol. Evol. 2014, 29, 444–450. [Google Scholar] [CrossRef]
- Nemergut, D.R.; Schmidt, S.K.; Fukami, T.; O’Neill, S.P.; Bilinski, T.M.; Stanish, L.F.; Knelman, J.E.; Darcy, J.L.; Lynch, R.C.; Wickey, P.; et al. Patterns and Processes of Microbial Community Assembly. Microbiol. Mol. Biol. Rev. 2013, 77, 342–356. [Google Scholar] [CrossRef] [Green Version]
- Finch-Savage, W.E.; Leubner-Metzger, G. Seed dormancy and the control of germination. New Phytol. 2006, 171, 501–523. [Google Scholar] [CrossRef]
- Jabot, F.; Chave, J. Analyzing Tropical Forest Tree Species Abundance Distributions Using a Nonneutral Model and through Approximate Bayesian Inference. Am. Nat. 2011, 178, E37–E47. [Google Scholar] [CrossRef]
- Gralka, M.; Hallatschek, O. Environmental heterogeneity can tip the population genetics of range expansions. eLife 2019, 8, e44359. [Google Scholar] [CrossRef]
- Sun, H.; Niu, Y.; Chen, Y.-S.; Song, B.; Liu, C.-Q.; Peng, D.-L.; Chen, J.-G.; Yang, Y. Survival and reproduction of plant species in the Qinghai-Tibet Plateau. J. Syst. Evol. 2014, 52, 378–396. [Google Scholar] [CrossRef] [Green Version]
- Little, L.; Eidesen, P.B.; Müller, E.; Dickinson, K.J.M.; Lord, J.M. Leaf and floral heating in cold climates: Do sub-antarctic megaherbs resemble tropical alpine giants? Polar Res. 2016, 35, 26030. [Google Scholar] [CrossRef] [Green Version]
- Morlon, H. Phylogenetic approaches for studying diversification. Ecol. Lett. 2014, 17, 508–525. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, B.; Levine, J.M. Ecological drift and the distribution of species diversity. Proc. R. Soc. B Boil. Sci. 2017, 284, 20170507. [Google Scholar] [CrossRef]
- Siqueira, T.; Saito, V.; Bini, L.M.; Melo, A.S.; Petsch, D.K.; Landeiro, V.L.; Tolonen, K.T.; Jyrkänkallio-Mikkola, J.; Soininen, J.; Heino, J. Community size can affect the signals of ecological drift and niche selection on biodiversity. Ecology 2020, 101, e03014. [Google Scholar] [CrossRef] [Green Version]
- Mouillot, D.; Leprêtre, A. A comparison of species diversity estimators. Res. Popul. Ecol. 1999, 41, 203–215. [Google Scholar]
- Wang, W.; Wang, Q.; Li, S.; Wang, G. Distribution and Species Diversity of Plant Communities along Transect on the Northeastern Tibetan Plateau. Biodivers. Conserv. 2006, 15, 1811–1828. [Google Scholar] [CrossRef]
- Pauli, H.; Gottfried, M.; Dullinger, S.; Abdaladze, O.; Akhalkatsi, M.; Alonso, J.L.B.; Coldea, G.; Dick, J.; Erschbamer, B.; Calzado, R.F.; et al. Recent Plant Diversity Changes on Europe’s Mountain Summits. Science 2012, 336, 353–355. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Zhou, H.; Ma, Z. Phylogenetic structure of alpine steppe plant communities along a precipitation and temperature gradient on the Tibetan Plateau. Glob. Ecol. Conserv. 2020, 24, e01379. [Google Scholar] [CrossRef]
- Cuesta, F.; Muriel, P.; Llambí, L.D.; Halloy, S.; Aguirre, N.; Beck, S.; Carilla, J.; Meneses, R.I.; Cuello, S.; Grau, A.; et al. Latitudinal and altitudinal patterns of plant community diversity on mountain summits across the tropical Andes. Ecography 2017, 40, 1381–1394. [Google Scholar] [CrossRef]
- Lamprecht, A.; Pauli, H.; Calzado, M.R.F.; Lorite, J.; Mesa, J.M.; Steinbauer, K.; Winkler, M. Changes in plant diversity in a water-limited and isolated high-mountain range (Sierra Nevada, Spain). Alp. Bot. 2021, 131, 27–39. [Google Scholar] [CrossRef]
- Liu, Q.; Buyantuev, A.; Wu, J.; Niu, J.; Yu, D.; Zhang, Q. Intensive land-use drives regional-scale homogenization of plant communities. Sci. Total Environ. 2018, 644, 806–814. [Google Scholar] [CrossRef]
- Zheng, P.R.; Li, C.H.; Ye, C.; Wang, H.; Wei, W.W.; Zheng, Y.; Zheng, X.Y. Factors Driving Wetland Herbs Distribution Range of Lake-Terrestrial Ecotone in Tibet, China. Paper Presented at Research Square. Available online: https://www.researchsquare.com/article/rs-1078363/v1 (accessed on 13 November 2021).
- Wang, R.; He, M.; Niu, Z. Responses of Alpine Wetlands to Climate Changes on the Qinghai-Tibetan Plateau Based on Remote Sensing. Chin. Geogr. Sci. 2020, 30, 189–201. [Google Scholar] [CrossRef]
- Cavieres, L.A.; Brooker, R.W.; Butterfield, B.J.; Cook, B.J.; Kikvidze, Z.; Lortie, C.; Michalet, R.; Pugnaire, F.I.; Schöb, C.; Xiao, S.; et al. Facilitative plant interactions and climate simultaneously drive alpine plant diversity. Ecol. Lett. 2014, 17, 193–202. [Google Scholar] [CrossRef]
- Hulshof, C.M.; Spasojevic, M.J. The edaphic control of plant diversity. Glob. Ecol. Biogeogr. 2020, 29, 1634–1650. [Google Scholar] [CrossRef]
- Davidson, E.A.; Janssens, I.A. Temperature sensitivity of soil carbon decomposition and feedbacks to climate change. Nature 2006, 440, 165–173. [Google Scholar] [CrossRef]
- Mipam, T.D.; Chen, S.; Liu, J.; Miehe, G.; Tian, L. Short-term yak-grazing alters plant-soil stoichiometric relations in an alpine meadow on the eastern Tibetan Plateau. Plant Soil 2019, 458, 125–137. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NMDS1 | NMDS2 | |||
|---|---|---|---|---|
| Adjust R2 | p | Adjust R2 | p | |
| lat | −0.052 | 0.821 | 0.267 | 0.012 |
| lon | −0.046 | 0.682 | 0.139 | 0.059 |
| TC | −0.054 | 0.881 | 0.063 | 0.148 |
| TN | −0.041 | 0.618 | −0.048 | 0.726 |
| TP | −0.024 | 0.466 | −0.045 | 0.669 |
| pH | −0.044 | 0.664 | 0.045 | 0.186 |
| Sty | −0.045 | 0.678 | −0.056 | 0.999 |
| MAT | −0.048 | 0.727 | 0.025 | 0.239 |
| MAP | 0.034 | 0.212 | 0.162 | 0.044 |
| DTR | −0.032 | 0.533 | 0.098 | 0.097 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cui, S.; Ouyang, J.; Lu, Y.; Liu, W.; Li, W.; Liu, G.; Zhou, W. The Diversity and Community Assembly Process of Wetland Plants from Lakeshores on the Qinghai-Tibetan Plateau. Diversity 2021, 13, 685. https://doi.org/10.3390/d13120685
Cui S, Ouyang J, Lu Y, Liu W, Li W, Liu G, Zhou W. The Diversity and Community Assembly Process of Wetland Plants from Lakeshores on the Qinghai-Tibetan Plateau. Diversity. 2021; 13(12):685. https://doi.org/10.3390/d13120685
Chicago/Turabian StyleCui, Shasha, Jian Ouyang, Yu Lu, Wenzhi Liu, Wenyang Li, Guihua Liu, and Wen Zhou. 2021. "The Diversity and Community Assembly Process of Wetland Plants from Lakeshores on the Qinghai-Tibetan Plateau" Diversity 13, no. 12: 685. https://doi.org/10.3390/d13120685