
Endangered Apes—Can Their Behaviors Be Used to Index Fear and Disturbance in Anthropogenic Landscapes?

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Study Animals
2.3. Defining Exposure and Accessibility
2.4. Data Collection
2.5. Analysis
3. Results
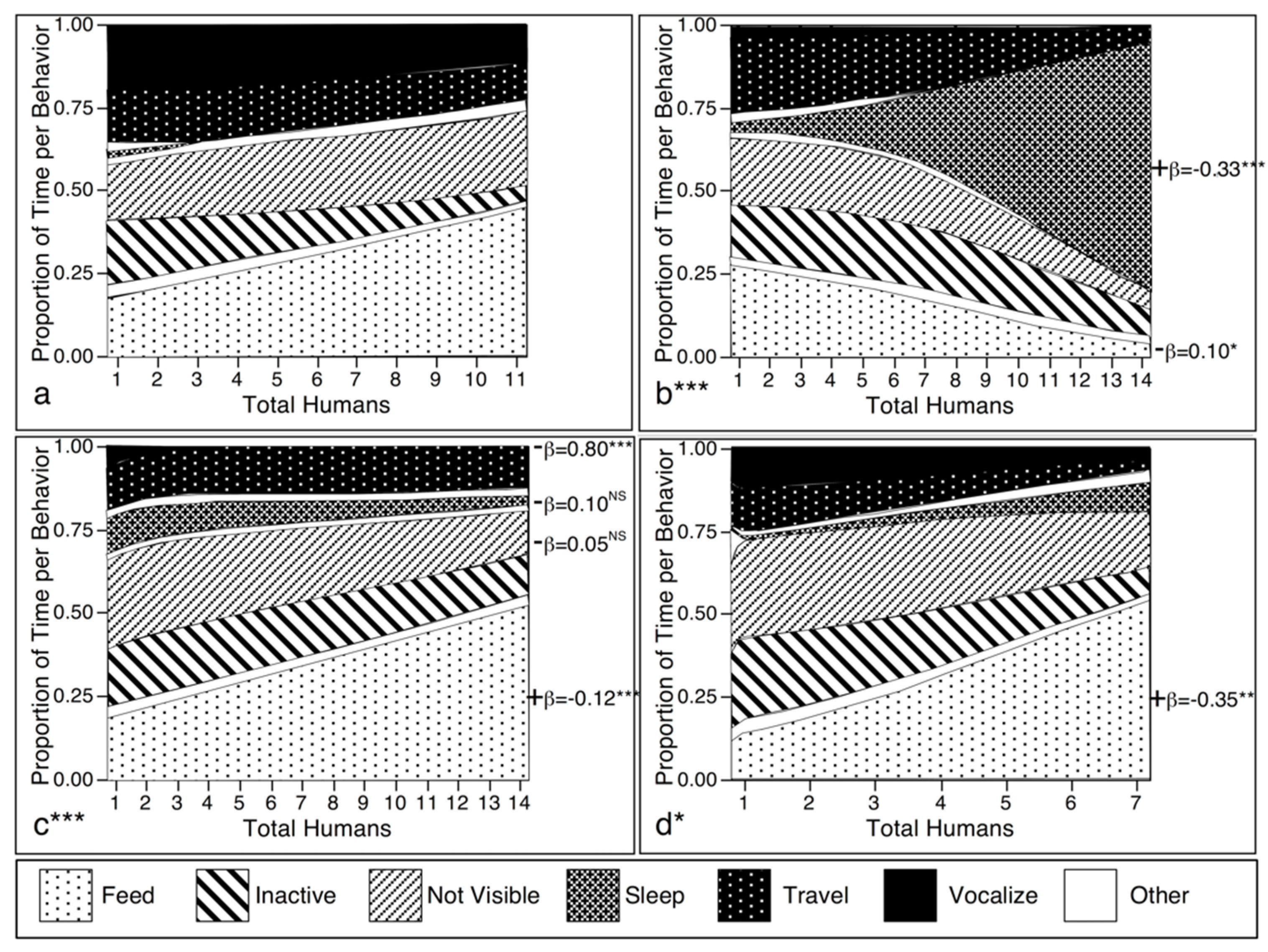
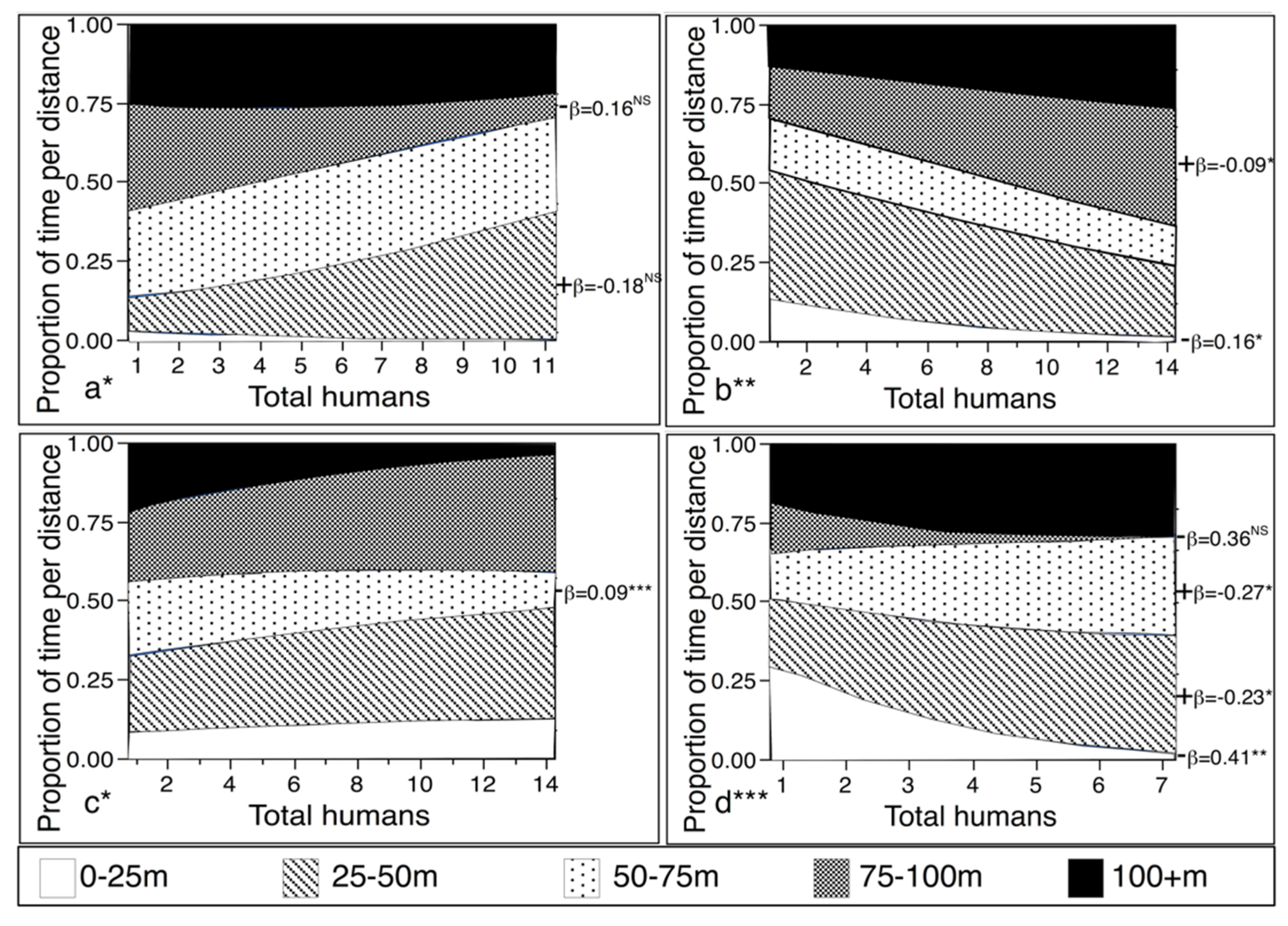
3.1. Test of Prediction I: If Javan Gibbons Treat Humans as Predators, Gibbon Groups Will Decrease Time Spent on Conspicuous Behavior as the Number of Humans Increase
3.1.1. Behavior by Number of Humans
3.1.2. Distance by Number of Humans
3.2. Prediction II: If Javan Gibbons Treat Humans as Predators, Gibbon Groups will Decrease Time Spent on Conspicuous Behavior as the Proximity to Humans Decreases
3.3. Prediction III: If Javan Gibbons Treat Humans as Predators, Gibbon Groups Will Respond to Humans More Similarly to How They Respond Other Predators Than How They Respond to Other Non-Predator Primate Species (Conspecifics, Monkey Species)
4. Discussion
4.1. Response to Human Group Size
4.2. Response to Distance to Nearest Human
4.3. Conservation Implications
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Duchesne, M.; Cote, S.D.; Barrette, C. Responses of woodland caribou to winter ecotourism in the Charlevoix Biosphere Reserve, Canada. Biol. Conserv. 2000, 96, 311–317. [Google Scholar] [CrossRef]
- Lott, D.F.; McCoy, M. Asian rhinos Rhinoceros unicornis on the run? Impact of tourist visits on one population. Biol. Conserv. 1995, 73, 23–26. [Google Scholar] [CrossRef]
- Rode, K.D.; Farley, S.D.; Fortin, J.; Robbins, C.T. Nutritional consequences of experimentally introduced tourism in brown bears. J. Wildl. Manag. 2007, 71, 929–939. [Google Scholar] [CrossRef]
- Constantine, R. Increased avoidance of swimmers by wild bottlenose dolphins (Tursiops truncatus) due to long-term exposure to swim-with-dolphin tourism. Mar. Mammal Sci. 2001, 17, 689–702. [Google Scholar] [CrossRef]
- Lusseau, D.; Higham, J.E.S. Managing the impacts of dolphin-based tourism through the definition of critical habitats: The case of bottlenose dolphins (Tursiops spp.) in Doubtful Sound, New Zealand. Tour. Manag. 2004, 25, 657–667. [Google Scholar] [CrossRef]
- Müllner, A.; Eduard Linsenmair, K.; Wikelski, M. Exposure to ecotourism reduces survival and affects stress response in hoatzin chicks (Opisthocomus hoazin). Biol. Conserv. 2004, 118, 549–558. [Google Scholar] [CrossRef]
- Skagen, S.K.; Knight, R.L.; Orians, G.H. Human disturbance of an avian scavenging guild. Ecol. Appl. 1991, 1, 215–225. [Google Scholar] [CrossRef] [PubMed]
- Gerrodette, T.; Gilmartin, W.G. Demographic consequences of changing pupping and hauling sites of the Hawaiian monk seal. Conserv. Biol. 1990, 4, 423–430. [Google Scholar] [CrossRef] [Green Version]
- Kappeler, M. The Javan Silvery Gibbon (Hylobates lar moloch). Ph.D. Thesis, Universitat Basel, Basel, Switzerland, 1981. [Google Scholar]
- Bshary, R. Diana monkeys, Cercopithecus diana, adjust their anti-predator response behaviour to human hunting strategies. Behav. Ecol. Sociobiol. 2001, 50, 251–256. [Google Scholar] [CrossRef] [Green Version]
- Nijman, V. Forest (and) Primates: Conservation and Ecology of the Endemic Primates of Java and Borneo; Tropenbos International: Amsterdam, The Netherlands, 2001. [Google Scholar]
- Frid, A.; Dill, L.M. Human-caused disturbance stimuli as a form of predation risk. Conserv. Ecol. 2002, 6, 11–26. [Google Scholar] [CrossRef]
- Stanford, C.B. Avoiding predators: Expectations and evidence in primate antipredator behavior. Int. J. Primatol. 2002, 23, 741–757. [Google Scholar] [CrossRef]
- Croes, B.M.; Laurance, W.F.; Lahm, S.A.; Tchignoumba, L.; Alonso, A.; Lee, M.E.; Campbell, P.; Buij, R. The Influence of hunting on antipredator behavior in central African monkeys and duikers. Biotropica 2007, 39, 257–263. [Google Scholar] [CrossRef]
- Miller, L.E.; Treves, A. Predation on primates: Past studies, current challenges, and directions for the future. In Primates in Perspective; Campbell, C.J., Fuentes, A., Mackinnon, K.C., Panger, M.A., Eds.; Oxford University Press: Oxford, UK, 2007; pp. 525–543. [Google Scholar]
- Grossberg, R.; Treves, A.; Naughton-Treves, L. The incidental ecotourist: Measuring visitor impacts on endangered howler monkeys at a Belizean archaeological site. Environ. Conserv. 2003, 30, 40–51. [Google Scholar] [CrossRef] [Green Version]
- Treves, A.; Brandon, K. Tourist impacts on the behavior of black howling monkeys (Alouatta pigra) at Lamanai, Belize. In Commensalism and Conflict: The Human-Primate Interface; Pattterson, J.D., Wallis, J., Eds.; The American Society of Primatologists: Norman, OK, USA, 2005; pp. 146–166. [Google Scholar]
- de la Torre, S.; Snowdon, C.T.; Bejarano, M. Effects of human activities on wild pygmy marmosets in Ecuadorian Amazonia. Biol. Conserv. 2000, 94, 153–163. [Google Scholar] [CrossRef]
- Klailova, M.; Hodgkinson, C.; Lee, P.C. Behavioral responses of one western lowland gorilla (Gorilla gorilla gorilla) group at Bai Hokou, Central African Republic, to tourists, researchers and trackers. Am. J. Primatol. 2010, 72, 897–906. [Google Scholar] [CrossRef] [PubMed]
- Johns, A.D. Behavioral responses of two Malaysian primates (Hylobates lar and Presbytis melalophos) to selective logging: Vocal behavior, territoriality, and nonemigration. Int. J. Primatol. 1985, 6, 423–433. [Google Scholar] [CrossRef]
- Johns, A.D. Effects of selective logging on the behavioral ecology of West Malaysian primates. Ecology 1986, 67, 684–694. [Google Scholar] [CrossRef]
- Brown, J.S.; Morgan, R.A. Effects of foraging behavior and spatial scale on diet selectivity: A test with fox squirrels. Oikos 1995, 74, 122–136. [Google Scholar] [CrossRef]
- Sinclair, A.R.E.; Arcese, P. Population consequences of predation sensitive foraging: The Serengeti wildebeest. Ecology 1995, 76, 882–891. [Google Scholar] [CrossRef]
- Ruxton, G.D.; Lima, S.L. Predator-induced breeding suppression and its consequences for predator-prey population dynamics. Proc. R. Soc. B Biol. Sci. 1997, 264, 409–415. [Google Scholar] [CrossRef] [Green Version]
- Schmitz, O.J.; Beckerman, A.P.; O’Brien, K.M. Behaviorally mediated trophic cascades: Effects of predation risk on food webs. Ecology 1997, 78, 1388–1399. [Google Scholar] [CrossRef]
- Brown, J.H.; Ernest, S.K.M. Rain and rodents: Complex dynamics of desert consumers. BioScience 2002, 52, 979–987. [Google Scholar] [CrossRef]
- Ripple, W.J.; Beschta, R.L. Wolves and the ecology of fear: Can predation risk restructure ecosystems. Bioscience 2004, 54, 755–766. [Google Scholar] [CrossRef] [Green Version]
- Berger, J. Fear, human shields, and the redistribution of prey and predators in protected areas. Biol. Lett. 2007, 3, 620–623. [Google Scholar] [CrossRef] [Green Version]
- Berger, J. The Better to Eat You with: Fear in the Animal World; University of Chicago Press: Chicago, IL, USA, 2008. [Google Scholar]
- Brown, J.S.; Kotler, B.P. Foraging and the ecology of fear. In Foraging: Behavior and Ecology; Stephens, D.W., Brown, J.S., Ydenberg, R.C., Eds.; University of Chicago Press: Chicago, IL, USA, 2007; pp. 437–482. [Google Scholar]
- Creel, S.; Christianson, D.; Liley, S.; Winnie, J.A. Predation risk affects reproductive physiology and demography of elk. Science 2007, 315, 960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Middleton, A.D.; Kauffman, M.J.; McWhirter, D.E.; Jimenez, M.D.; Cook, R.C.; Cook, J.G.; Albeke, S.E.; Sawyer, H.; White, P.J. Linking anti-predator behavior to prey demography reveals limited risk effects of an actively hunting large carnivore. Ecol. Lett. 2013, 16, 1023–1030. [Google Scholar] [CrossRef]
- Brown, J.S.; Alkon, P.U. Testing values of crested porcupine habitats by experimental food patches. Oecologia 1990, 83, 512–518. [Google Scholar] [CrossRef]
- Creel, S.; Christianson, D. Relationships between direct predation and risk effects. Trends Ecol. Evol. 2008, 23, 194–201. [Google Scholar] [CrossRef]
- Bejder, L.; Samuels, A.; Whitehead, H.; Gales, N. Interpreting short-term behavioural responses to disturbance within a longitudinal perspective. Anim. Behav. 2006, 72, 1149–1158. [Google Scholar] [CrossRef] [Green Version]
- Strier, K.B. Demography and conservation of an endangered primate, Brachyteles arachnoids. Conserv. Biol. 1991, 5, 214–218. [Google Scholar] [CrossRef]
- Strier, K.B. Population viabilities and conservation implications for muriquis (Brachyteles arachnoids) in Brazil’s Atlantic forest. Biotropica 2000, 32, 903–913. [Google Scholar]
- Strier, K.B.; Boubli, J.P.; Possamai, C.B.; Mendes, S.L. Population demography of northern muriquis (Brachyteles hypoxanthus) at the Estação Biológica de Caratinga/Reserva particular do Patrimônio Natural-Felìciano Miguel Abdala, Minas Gerais, Brazil. Am. J. Phys. Anthropol. 2006, 130, 227–237. [Google Scholar] [CrossRef]
- Cowlishaw, G.; Dunbar, R. Primate Conservation Biology; University of Chicago Press: Chicago, IL, USA, 2000. [Google Scholar]
- Embassy of Indonesia, Ottawa. Available online: http://www.indonesia-ottawa.org/ (accessed on 1 January 2008).
- Wessing, R. A change in the forest: Myth and history in West Java. J. Southeast Asian Stud. 1993, 24, 1–17. [Google Scholar] [CrossRef]
- Geertz, C. Agricultural Involution: The Process of Ecological Change in Indonesia; University of California Press: Berkeley, CA, USA, 1963. [Google Scholar]
- Nijman, V. Conservation of the Javan gibbon Hylobates moloch: Population estimates, local extinctions, and conservation priorities. Raffles Bull. Zool. 2004, 52, 271–280. [Google Scholar]
- Malone, N.M. The Socioecology of the Critically Endangered Javan Gibbon (Hylobates moloch): Assessing the Impact of Anthropogenic Disturbance on Primate Social Systems. Ph.D. Thesis, University of Oregon, Eugene, OR, USA, 2007. [Google Scholar]
- Malone, N.M.; Fuentes, A.; Purnama, A.R.; Wedana, I. Displaced hylobatids: Biological, cultural, and economic aspects of the primate trade in Jawa and Bali, Indonesia. Trop. Biodivers. 2003, 8, 41–50. [Google Scholar]
- Andayani, N.; Brockelman, W.; Geissmann, T.; Nijman, V.; Supriatna, J. Hylobates moloch. IUCN 2012. IUCN Red List of Threatened Species. Version 2012.2. Available online: www.iucnredlist.org (accessed on 18 January 2013).
- Supriatna, J.; Mootnick, A.; Andayani, N. Javan gibbon (Hylobates moloch): Population and conservation. In Indonesian Primates; Gursky, S., Supriatna, J., Eds.; Springer: New York, NY, USA, 2010; pp. 57–72. [Google Scholar]
- Supriatna, J. Conservation programs for the endangered Javan gibbon (Hylobates moloch). Primate Conserv. 2006, 21, 155–162. [Google Scholar] [CrossRef]
- Nijman, V. In Full Swing. An Assessment of the Trade in Gibbons and Orangutans on Java and Bali, Indonesia; TRAFFIC South-east Asia: Kuala Lumpur, Malaysia, 2005. [Google Scholar]
- Wedana, A.P.I.M.; Jeffery, S. Reinforcing the Javan silvery gibbon population in the Mount Tilu Nature Reserve, West Java, Indonesia. In Proceedings of the Programme of the XXVIth Congress of the International Primatological Society, Chicago, IL, USA, 21–27 August 2016. [Google Scholar]
- Lebbie, A.R.; Freudenberger, M. Sacred groves in Africa: Forest patches in transition. In Forest Patches in Tropical Landscapes; Shelhas, J., Greenberg, R., Eds.; Island Press: Washington, DC, USA, 1996; pp. 300–324. [Google Scholar]
- Reisland, M.A. Conservation in a Sacred Forest: An Integrated Approach to Assessing the Management of a Community-Based Conservation Site. Ph.D. Thesis, University of Wisconsin, Madison, WI, USA, 2013. [Google Scholar]
- Malone, N.; Wade, A.; Putra, M.W.A.; Reisland, M.; Selby, M. Calibrating a conservation strategy for silvery gibbons (Hylobates moloch). Am. J. Phys. Anthropol. 2012, 147, 202. [Google Scholar]
- Malone, N.; Wedana, M. Struggling for socio-ecological resilience: A long-term study of silvery gibbons (Hylobates moloch) in the fragmented Sancang Forest Nature Reserve, West Java, Indonesia. In Primate Research and Conservation in the Anthropocene; Behie, A., Teichroeb, J., Malone, N., Eds.; Cambridge University Press: Cambridge, UK, 2019; pp. 17–32. [Google Scholar]
- Cybertracker2010. Available online: http://www.cybertracker.org (accessed on 15 June 2010).
- Martin, P.; Bateson, P. Measuring Behavior: An Introductory Guide, 2nd ed.; Cambridge University Press: Cambridge, UK, 1993. [Google Scholar]
- Holm, S. A simple sequentially rejective multiple test procedure. Scand. J. Stat. 1979, 6, 65–70. [Google Scholar]
- Bejder, L.; Samuels, A.; Whitehead, H.; Finn, H.; Allen, S. Impact assessment research: Use and misuse of habituation, sensitisation and tolerance in describing wildlife responses to anthropogenic stimuli. Mar. Ecol. Prog. Ser. 2009, 395, 177–185. [Google Scholar] [CrossRef]
- Gill, J.A.; Sutherland, W.J. Predicting the consequences of human disturbance from behavioural decisions. In Behaviour and Conservation; Gosling, L.M., Sutherland, W.J., Eds.; Cambridge University Press: Cambridge, UK, 2000; pp. 51–64. [Google Scholar]
- Kappeler, M. Vocal bouts and territorial maintenance in the moloch gibbon. In The Lesser Apes. Evolutionary and Behavioural Biology; Preuschoft, H., Chivers, D.J., Brockelman, W.Y., Creel, N., Eds.; Edinburgh University Press: Edinburgh, UK, 1984; pp. 376–389. [Google Scholar]
- Geissmann, T.; Nijman, V. Calling in wild silvery gibbons (Hylobates moloch) in Java (Indonesia): Behavior, phylogeny, and conservation. Am. J. Primatol. 2006, 68, 1–19. [Google Scholar] [CrossRef]
- Mitani, J.C. The behavioral regulation of monogamy in gibbons (Hylobates muelleri). Behav. Ecol. Sociobiol. 1984, 15, 225–229. [Google Scholar] [CrossRef]
- Mitani, J.C. Gibbon song duets and intergroup spacing. Behaviour 1985, 92, 59–96. [Google Scholar] [CrossRef]
- Mitani, J.C. Location-specific responses of gibbons (Hylobates muelleri) to male songs. Z. Tierpsychol. 1985, 70, 219–224. [Google Scholar] [CrossRef]
- Mitani, J.C. Territoriality and monogamy among agile gibbons (Hylobates agilis). Behav. Ecol. Sociobiol. 1987, 20, 265–269. [Google Scholar] [CrossRef]
- Raemaekers, J.J.; Raemaekers, P.M. Field playback of loud calls to gibbons (Hylobates lar): Territorial, sex-specific and species-specific responses. Animal Behav. 1985, 33, 481–493. [Google Scholar] [CrossRef]
- Raemaekers, P.M.; Raemaekers, J.J. Long-range vocal interactions between groups of gibbons (Hylobates lar). Behaviour 1985, 95, 26–44. [Google Scholar] [CrossRef]
- Leighton, D.R. Gibbons: Territoriality and monogamy. In Primate Societies; Smuts, B.B., Cheney, D.L., Seyfarth, R.M., Wrangham, R.W., Struhsaker, T.T., Eds.; University of Chicago Press: Chicago, IL, USA, 1987; pp. 135–145. [Google Scholar]
- Cowlishaw, G. Song function in gibbons. Behaviour 1992, 121, 131–153. [Google Scholar] [CrossRef]
- Geissmann, T. Duet songs of the siamang, Hylobates syndactylus: II. Testing the pair-bonding hypothesis during a partner exchange. Behaviour 1999, 136, 1005–1039. [Google Scholar] [CrossRef] [Green Version]
- Geissmann, T.; Orgeldinger, M. The relationship between duet songs and pair bonds in siamangs, Hylobates syndactylus. Anim. Behav. 2000, 60, 805–809. [Google Scholar] [CrossRef] [Green Version]
- Krüger, O. The role of ecotourism in conservation: Panacea or Pandora’s box? Biodivers. Conserv. 2005, 14, 579–600. [Google Scholar] [CrossRef]
- Ferguson, M.A.D.; Keith, L.B. Influence of Nordic skiing on distribution of moose and elk in Elk Island National Park, Alberta. Can. Field-Nat. 1982, 96, 69–78. [Google Scholar]
- Lippold, L.K. Primate population decline at Cabo Blanco Absolute Nature Reserve, Costa Rica. Brenesia 1990, 34, 145–152. [Google Scholar]
- Cassirer, E.F.; Freddy, D.J.; Ables, E.D. Elk responses to disturbance by cross-country skiers in Yellowstone National Park. Wildl. Soc. Bull. 1992, 20, 375–381. [Google Scholar]
- Beier, P. Dispersal of juvenile cougars in fragmented habitat. J. Wildl. Manag. 1995, 59, 228–237. [Google Scholar] [CrossRef]
- McNeilage, A. Ecotourism and mountain gorillas in the Virunga volcanoes. In The Exploitation of Mammal Populations; Taylor, V.J., Dunstone, N., Eds.; Chapman & Hall: London, UK, 1996; pp. 334–344. [Google Scholar]
- Gander, H.; Ingold, P. Reactions of male alpine chamois Rupicapra r. rupicapra to hikers, joggers, and mountainbikers. Biol. Conserv. 1997, 79, 107–109. [Google Scholar] [CrossRef]
- Buckley, R. Impacts of ecotourism on terrestrial wildlife. In Environmental Impacts of Ecotourism; Buckley, R., Ed.; CABI Publishing: Cambridge, UK, 2004; pp. 211–228. [Google Scholar]
- Reisland, M.A.; Lambert, J.E. Sympatric apes in sacred forests: Shared space and habitat use by humans and endangered Javan gibbons (Hylobates moloch). PLoS ONE 2016, 11, e0146891. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | # Hum Rank 1 | Access. Rank 2 | Gibbon | Age/Sex | # Scans | Obs. Hrs. | Hum/scan 3 | SD |
|---|---|---|---|---|---|---|---|---|
| A | 3 | 2 | Amelia | Adult Female | 1280 | 213.33 | 1.46 a | 1.0 |
| B | 2 | 1 | Tono | Adult Male | 1468 | 244.67 | 1.73 b | 1.66 |
| Tini | Adult Female | 1592 | 265.33 | 1.82 b | 1.7 | |||
| Udian (dis. 11/10) | Juvenile | 194 | 32.33 | 2.07 b | 1.77 | |||
| C | 1 | 3 | Jay | Adult Male | 569 | 94.83 | 3.24 c | 3.1 |
| Ann | Adult Female | 663 | 110.5 | 2.94 c | 2.88 | |||
| Cika | Juvenile Male | 591 | 98.5 | 3.10 c | 2.83 | |||
| Wana (born 5/11) | Infant | N/A | N/A | N/A | N/A | |||
| D | 3 | 4 | Sastro | Adult Male | 311 | 51.83 | 1.25 a | 0.66 |
| Dian | Adult Female | 477 | 79.5 | 1.32 a | 0.78 | |||
| Wardini | Juvenile Female | 372 | 62 | 1.25 a | 0.66 | |||
| Bayi (dis. 3/11) | Infant | N/A | N/A | N/A | N/A |
| Group | Behavior | Distance | Difference from Expected a | N | Cell χ2 |
|---|---|---|---|---|---|
| A N = 692 | Feed | 0–25 m | + | 14 | 12.50 |
| 50–75 m | + | 80 | 7.09 | ||
| 75–100 m | – | 40 | 16.27 | ||
| Vocalize | 25–50 m | – | 3 | 6.45 | |
| 75–100 m | + | 62 | 12.43 | ||
| B N = 1713 | Feed | 0–25 m | + | 121 | 39.95 |
| 50–75 m | – | 52 | 5.92 | ||
| 75–100 m | – | 41 | 10.14 | ||
| 100+ m | – | 14 | 13.99 | ||
| Not Visible | 0–25 m | – | 20 | 22.11 | |
| 25–50 m | – | 129 | 5.07 | ||
| 50–75 m | + | 77 | 6.14 | ||
| 100+ m | + | 64 | 41.24 | ||
| Sleep | 100+ m | – | 0 | 4.89 | |
| Inactive | 100+ m | – | 8 | 10.19 | |
| Travel | 0–25 m | – | 42 | 6.43 | |
| Social | 50–75 m | – | 1 | 4.10 | |
| Vocalize | 25–50 m | – | 15 | 8.73 | |
| 100+ m | + | 19 | 26.98 | ||
| Other | 0–25 | + | 8 | 3.89 | |
| C N = 536 | Feed | 100+ m | – | 0 | 11.57 |
| Not Visible | 0–25 m | – | 17 | 6.73 | |
| 100+ m | + | 32 | 7.55 | ||
| Sleep | 50–75 m | + | 17 | 7.46 | |
| 100+ m | – | 0 | 4.16 | ||
| Inactive | 0–25 m | + | 39 | 25.51 | |
| 100+ m | – | 0 | 11.10 | ||
| Vocalize | 100+ m | + | 17 | 45.53 | |
| D N = 790 | Feed | 0–25 m | + | 57 | 13.53 |
| 50–75 m | – | 11 | 4.64 | ||
| Not Visible | 0–25 m | – | 45 | 12.76 | |
| 50–75 m | + | 67 | 10.62 | ||
| Sleep | 50–75 m | + | 3 | 6.54 | |
| Inactive | 0–25 m | + | 85 | 10.06 | |
| 50–75 m | – | 21 | 6.08 | ||
| 100+ m | – | 1 | 17.22 | ||
| Travel | 0–25 m | – | 24 | 3.90 | |
| Vocalize | 0–25 m | – | 17 | 6.77 | |
| 100+ m | + | 34 | 57.09 | ||
| Social | 0–25 m | + | 6 | 8.52 | |
| Other | 0–25 m | + | 7 | 9.94 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reisland, M.A.; Malone, N.; Lambert, J.E. Endangered Apes—Can Their Behaviors Be Used to Index Fear and Disturbance in Anthropogenic Landscapes? Diversity 2021, 13, 660. https://doi.org/10.3390/d13120660
Reisland MA, Malone N, Lambert JE. Endangered Apes—Can Their Behaviors Be Used to Index Fear and Disturbance in Anthropogenic Landscapes? Diversity. 2021; 13(12):660. https://doi.org/10.3390/d13120660
Chicago/Turabian StyleReisland, Melissa A., Nicholas Malone, and Joanna E. Lambert. 2021. "Endangered Apes—Can Their Behaviors Be Used to Index Fear and Disturbance in Anthropogenic Landscapes?" Diversity 13, no. 12: 660. https://doi.org/10.3390/d13120660