Dasybranchus geojedoensis sp. nov. (Annelida: Capitellidae), a New Capitellid Species from Southern Korea


Abstract
:1. Introduction
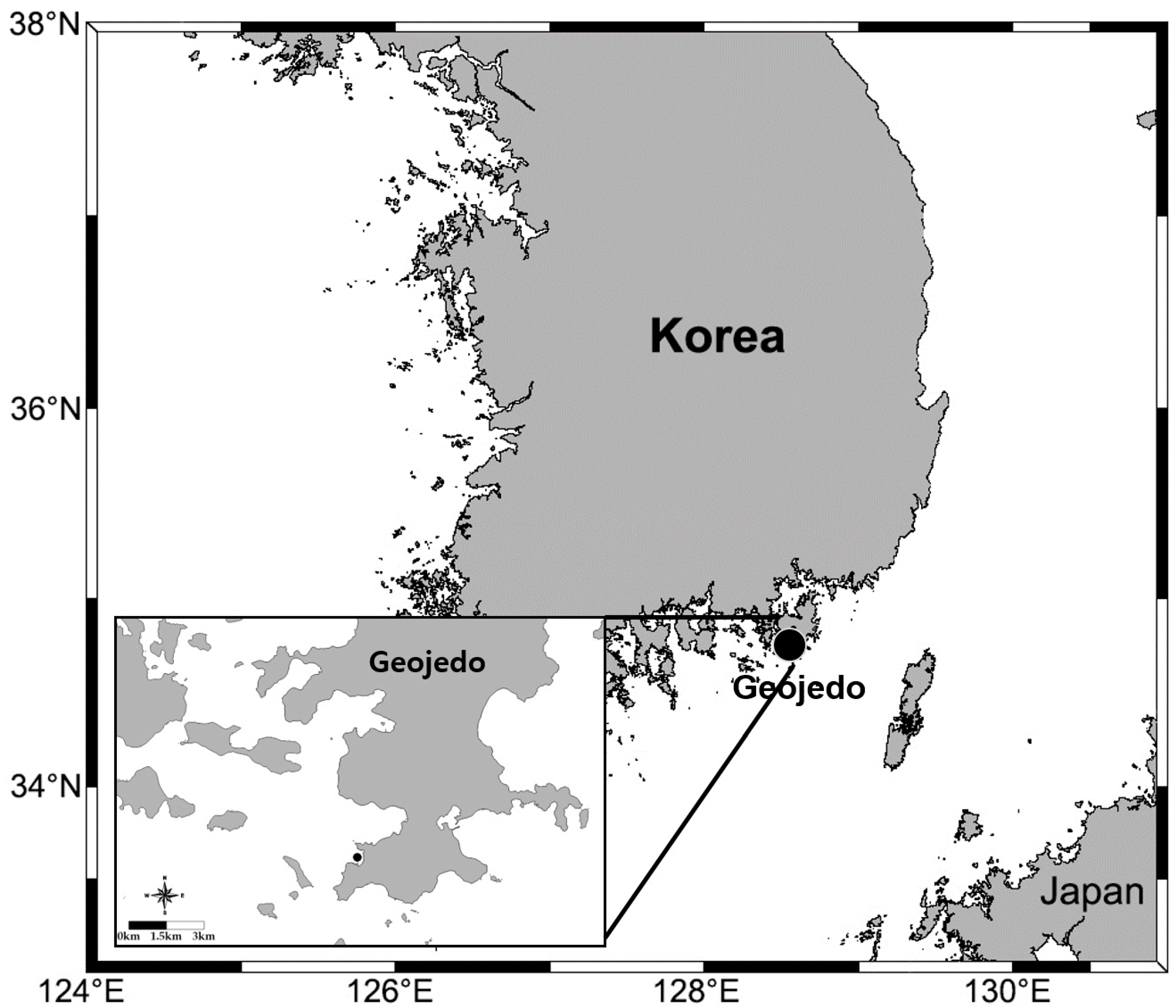
2. Materials and Methods
3. Results
3.1. Systematics
3.2. Generic Diagnosis
3.3. Material Examined
3.4. Species Diagnosis
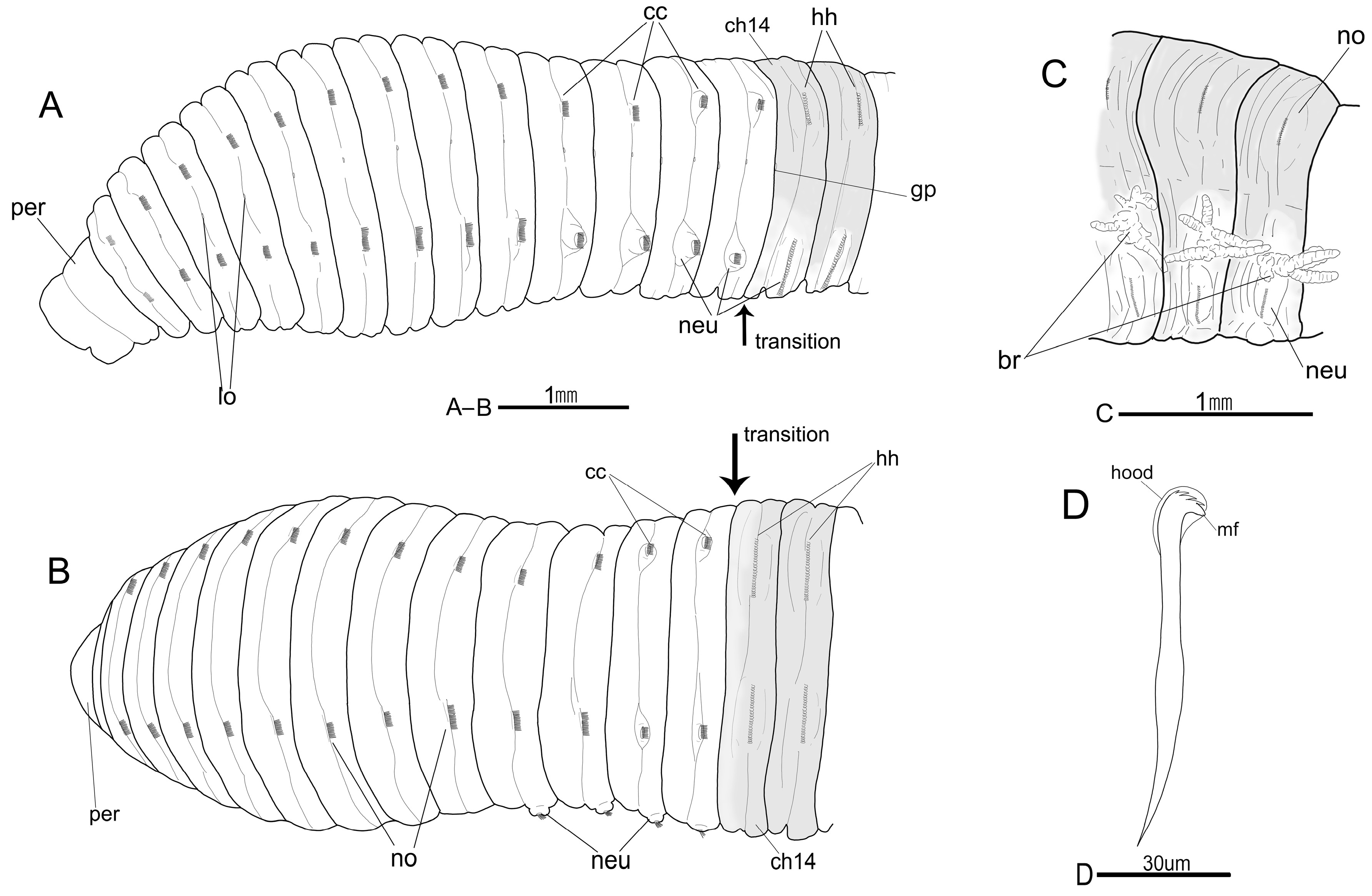
3.5. Description
3.6. Etymology
3.7. Distribution and Ecology
3.8. Remarks
3.9. Molecular Comparison
4. Discussion
| 1 Thorax with 12 chaetigers | 2 |
| - Thorax with 13 chaetigers | 3 |
| 2 Prostomium without eyespots; all thoracic segments tessellated | |
| D. carneus Grube, 1870 | |
| - Prostomium with eyespots; anterior thoracic segments tessellated | |
| D. rectus Treadwell, 1901 | |
| 3 Prostomium with eyespots; all thoracic segments smooth, | |
| D. lunulatus Ehlers, 1887 [26] | |
| - Prostomium without eyespots | 4 |
| 4 Anterior abdominal notopodia dorsally far apart from each other | 5 |
| - Anterior abdominal notopodia dorsally close to each other | 8 |
| 5 Branchiae beginning from near abdominal chaetigers 10–20 | 6 |
| - Branchiae beginning from near abdominal chaetigers 40–50 | 7 |
| 6 Each branchial tuft with 15–20 filaments; genital pores between chaetigers 12–15 | |
| D. caducus (Grube, 1846) | |
| - Each branchial tuft with 5–30 filaments; genital pores between chaetigers 9–25 | |
| D. platyceps Hartman, 1947 | |
| 7 Anterior thoracic segments weakly tessellated; genital pores between chaetigers 10–14 | |
| D. geojedoensis sp. nov. | |
| - All thoracic segments tessellated; genital pores between chaetigers 9–15 | |
| D. bipartitus (Schmarda, 1861) | |
| 8 All thoracic segments tessellated | 9 |
| - All thoracic segments smooth | 10 |
| 9 Abdominal hooded hooks with teeth in three to four rows above main fang; each branchial tuft with 10–20 filaments | |
| D. gajolae Eisig, 1887 | |
| - Abdominal hooded hooks with six teeth in two rows above main fang; each branchial tuft with 2–10 filaments | |
| D. parplatyceps Kudenov, 1975 | |
| - Abdominal hooded hooks with three teeth in one row above main fang; each branchial tuft with 30 filaments | |
| D. lumbricoides Grube, 1878 [27] | |
| 10 Abdominal hooded hooks with seven teeth in two rows above main fang | |
| D. eisigi Green, 2002 | |
| - Abdominal hooded hooks with three teeth in two to three rows above main fang; each branchial tuft with 2–3 filaments | |
| D. glabrus Moore, 1909 [28]. | |
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Grube, A.E. Die Familien der Anneliden.; Archiv für Naturgeschichte: Berlin, Germany, 1850; pp. 249–364. [Google Scholar]
- Day, J.H. A Monograph on the Polychaeta of Southern Africa; Part II; British Museum (Natural History): London, UK, 1967; pp. 1–450. [Google Scholar]
- Green, K. Capitellidae (Polychaeta) from the Andaman Sea. Phuket Mar. Biol. Cent. Spec. Publ. 2002, 24, 249–343. [Google Scholar]
- Grube, E. Beschreibungen neuer oder weniger bekannter von Hrn. Ehrenberg gesammelter Anneliden des rothen Meeres. Mon. K. Preuss. Akad. Wiss. Berl. 1870, 1869, 484–521. [Google Scholar]
- Hartman, O. Polychaetous annelids. Part VII. Capitellidae. Allan Hancock Pac. Exped. 1947, 10, 391–481. [Google Scholar]
- Armstrong, M. The predator-prey relationships of Irish Sea poor-cod (Trisopterus minutus L.), pouting (Trisopterus luscus L.) and cod (Gadus morhua L.). Ices J. Mar. Sci. 1982, 40, 135–152. [Google Scholar] [CrossRef]
- Fauchald, K.; Jumars, P.A. The diet of worms: A study of polychaete feeding guilds. Oceanogr. Mar. Biol. Annu. Rev. 1979, 17, 193–284. [Google Scholar]
- Stoner, A.W.; Buchanan, B.A. Ontogeny and overlap in the diets of four tropical Callinectes species. Bull. Mar. Sci. 1990, 46, 3–12. [Google Scholar]
- Grube, A.E. Beschreibung neuer oder wenig bekannter Anneliden. Beitrag: Canephorus, Ammochares, Dasymallus, Scalis-Arten. Arch. Nat. Berl. 1846, 12, 161–171. [Google Scholar]
- Eisig, H. Monographie der Capitelliden des Golfes von Neapel. Fauna Flora Golf. Neapel 1887, 16, 1–906. [Google Scholar]
- Treadwell, A.L. The polychaetous annelids of Porto Rico. Bull. U. S. Fish Comm. 1901, 20, 181–210. [Google Scholar]
- Schmarda, L.K. Neue Wirbellose Thiere Beobachtet und Gesammelt auf einer Reise um die Erde 1853 bis 1857; Wilh. Engelmann: Leipzig, Germany, 1861; Volume 3, pp. 1–164. [Google Scholar]
- Paik, E.I. New records of five polychaetous annelids species in Korea. Korean J. Fish. Aquat. Sci. 1979, 12, 35–39. [Google Scholar]
- Jeong, M.-K.; Wi, J.H.; Suh, H.-L. A reassessment of Capitella species (Polychaeta: Capitellidae) from Korean coastal waters, with morphological and molecular evidence. Mar. Biodivers. 2017, 48, 1969–1978. [Google Scholar] [CrossRef]
- Carr, C.M.; Hardy, S.M.; Brown, T.M.; Macdonald, T.A.; Hebert, P.D. A tri-oceanic perspective: DNA barcoding reveals geographic structure and cryptic diversity in Canadian polychaetes. PLoS ONE 2011, 6, e22232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colgan, D.; McLauchlan, A.; Wilson, G.; Livingston, S.; Edgecombe, G.; Macaranas, J.; Cassis, G.; Gray, M. Histone H3 and U2 snRNA DNA sequences and arthropod molecular evolution. Aust. J. Zool. 1998, 46, 419–437. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef]
- Grube, A.E. Noch ein Wort über die Capitellen und ihre Stelle im Systeme der Anneliden; Archiv fur Naturgeschichte: Berlin, Germany, 1862; pp. 366–378. [Google Scholar]
- Day, J.H. The Polychaeta of South Africa. Part 3. Sedentary species from Cape shores and estuaries. Zool. J. Linn. Soc. 1955, 42, 407–452. [Google Scholar] [CrossRef]
- Kudenov, J.D. Sedentary polychaetes from the Gulf of California, Mexico. J. Nat. Hist. 1975, 9, 205–231. [Google Scholar] [CrossRef]
- Jeong, M.K.; Soh, H.Y.; Suh, H.L. Three new species of Heteromastus (Annelida, Capitellidae) from Korean waters, with genetic evidence based on two gene markers. Zookeys 2019, 869, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Magalhães, W.F.; Bailey-Brock, J.H. Capitellidae Grube, 1862 (Annelida: Polychaeta) from the Hawaiian Islands with description of two new species. Zootaxa 2012, 3581, 1–52. [Google Scholar] [CrossRef]
- Silva, C.; Amaral, A.C.Z. Scyphoproctus Gravier, 1904 (Annelida, Capitellidae): Description of three new species and relocation of Heteromastides Augener, 1914 in Scyphoproctus. Zootaxa 2019, 4560, 95–120. [Google Scholar] [CrossRef]
- Silva, C.F.; Shimabukuro, M.; Alfaro-Lucas, J.M.; Fujiwara, Y.; Sumida, P.Y.; Amaral, A.C. A new Capitella polychaete worm (Annelida: Capitellidae) living inside whale bones in the abyssal South Atlantic. Deep Sea Res. Oceanogr. Res. Pap. 2016, 108, 23–31. [Google Scholar] [CrossRef]
- Ehlers, E. Reports on the Results of Dredging, Under the Direction of L.F. Pourtalès, During the Years 1868–1870, and of Alexander Agassiz in the Gulf of Mexico (1877–78) and in the Caribbean Sea (1878–79), in the U.S. Coast Survey Steamer "Blake", Lieut.-Com. C.D. Sigsbee, U.S.N., and Commander J.R. Bartlett, U.S.N., Commanding: Report on the Annelids; Memoirs of the Museum of Comparative Zoology at Harvard College: Cambridge, MA, USA, 1887; pp. 1–335. [Google Scholar]
- Grube, A.E. Annulata Semperiana. Beiträge zur Kenntnis der Annelidenfauna der Philippine; Kessinger Publishing LLC: Whitefish, MT, USA, 1878; pp. 1–300. [Google Scholar]
- Moore, J.P. Polychaetous annelids from Monterey Bay and San Diego, California. Proc. Acad. Natl. Sci. Phila. 1909, 61, 235–295. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Location | Latitude/Longitude | Species | Type | Collection Numbers | Accession Numbers | ||
|---|---|---|---|---|---|---|---|
| Nation | District | mtCOI | Histone H3 | ||||
| Korea | Geojedo | 34°43.08′ N 128°35.13′ E | D. geojedoensis sp. nov. | Holotype | NA00141494 | ||
| Paratype | NA00141495 | MT513740 | MT513742 | ||||
| Paratype | NA00141496 | MT513741 | MT513743 | ||||
| Species | Tessellated Segments | Genital Pores | Denticulation of Hooded Hooks | Branchiae (Number) | MGSP | Habitat (Type Locality) | References |
|---|---|---|---|---|---|---|---|
| D. geojedoensis sp. nov. | Ch 1–5 | Between Ch 10–14 | >26 teeth in 4 rows (3/6/6/ > 11) | from 47th abdominal chaetiger (7–9) | Dorsum of abdomen stained light blue | Subtidal, 13 m depth, sandy mud (southern Korea) | This study |
| D. bipartitus | Ch 1–13 | Between Ch 9–15 | >17 teeth in 3 rows (1–4/4–8/?) | from 40th abdominal chaetiger (2–13) | Not examined | Unknown (South Africa) | [9,12,20] |
| D. caducus | Ch 1–9 | Between Ch 12–15 | 3–24 teeth in 1–3 row(s); reexamination required | from 20th abdominal chaetiger (>20) | Not examined | Sandy mud (Mediterranean Sea) | [2,3] |
| D. eisigi | Absent | Between Ch 11–15 | 7 teeth in 2 rows (4/3) | Not examined | Ch 1–14 stained green, Ch 15–18 and venter of Ch 19–21 stained dark green | Subtidal, 21 m depth, sand with shell fragments (Thailand) | [3] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jeong, M.-K.; Soh, H.Y. Dasybranchus geojedoensis sp. nov. (Annelida: Capitellidae), a New Capitellid Species from Southern Korea. Diversity 2020, 12, 290. https://doi.org/10.3390/d12080290
Jeong M-K, Soh HY. Dasybranchus geojedoensis sp. nov. (Annelida: Capitellidae), a New Capitellid Species from Southern Korea. Diversity. 2020; 12(8):290. https://doi.org/10.3390/d12080290
Chicago/Turabian StyleJeong, Man-Ki, and Ho Young Soh. 2020. "Dasybranchus geojedoensis sp. nov. (Annelida: Capitellidae), a New Capitellid Species from Southern Korea" Diversity 12, no. 8: 290. https://doi.org/10.3390/d12080290