
The Role of Extracellular Vesicles (EVs) in Chronic Graft vs. Host Disease, and the Potential Function of Placental Cell-Derived EVs as a Therapeutic Tool
Abstract
:1. Introduction
2. Results
2.1. Characteristics of cGVHD Patient EVs Compared to EVs of Healthy Controls
2.1.1. cGVHD Patient Characteristics
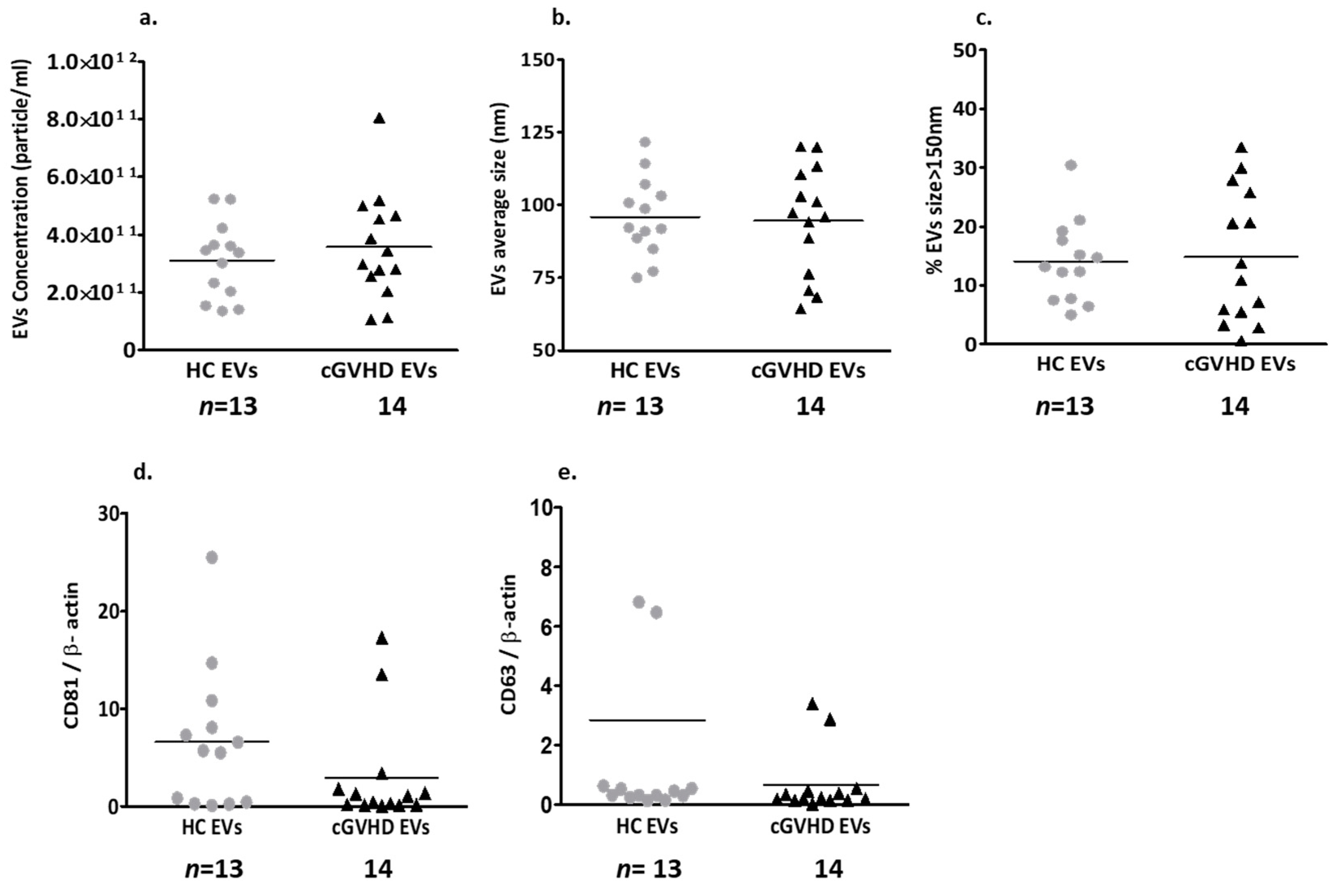
2.1.2. Patient EV Characteristics
Patient EV Size and Exosome Markers
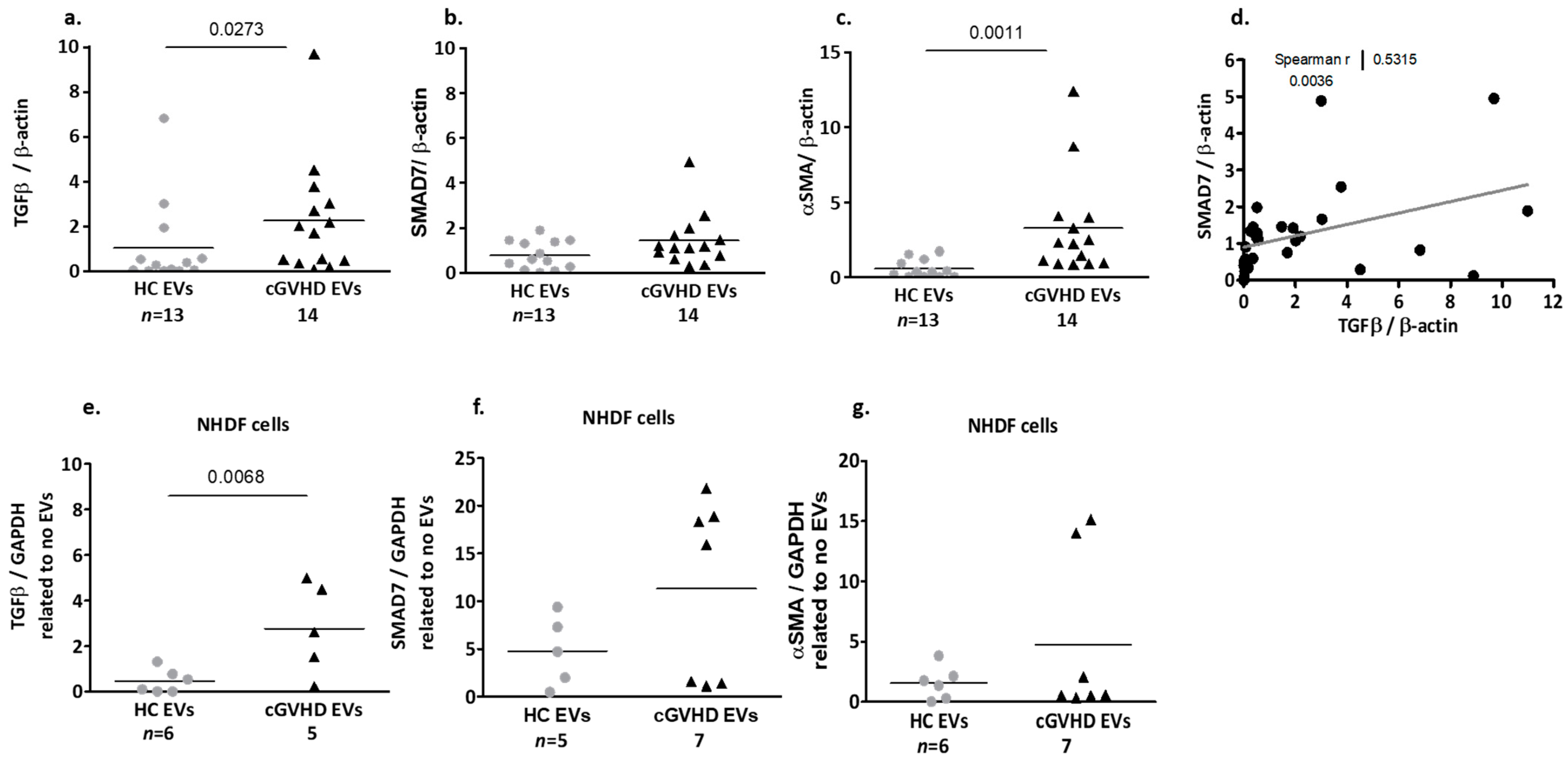
Fibrosis-Related Proteins Cargo of cGVHD Patient EVs
2.1.3. Patient EV Effects on Dermal Fibroblasts (NHDF)
2.2. The Therapeutic Properties of Placental EVs
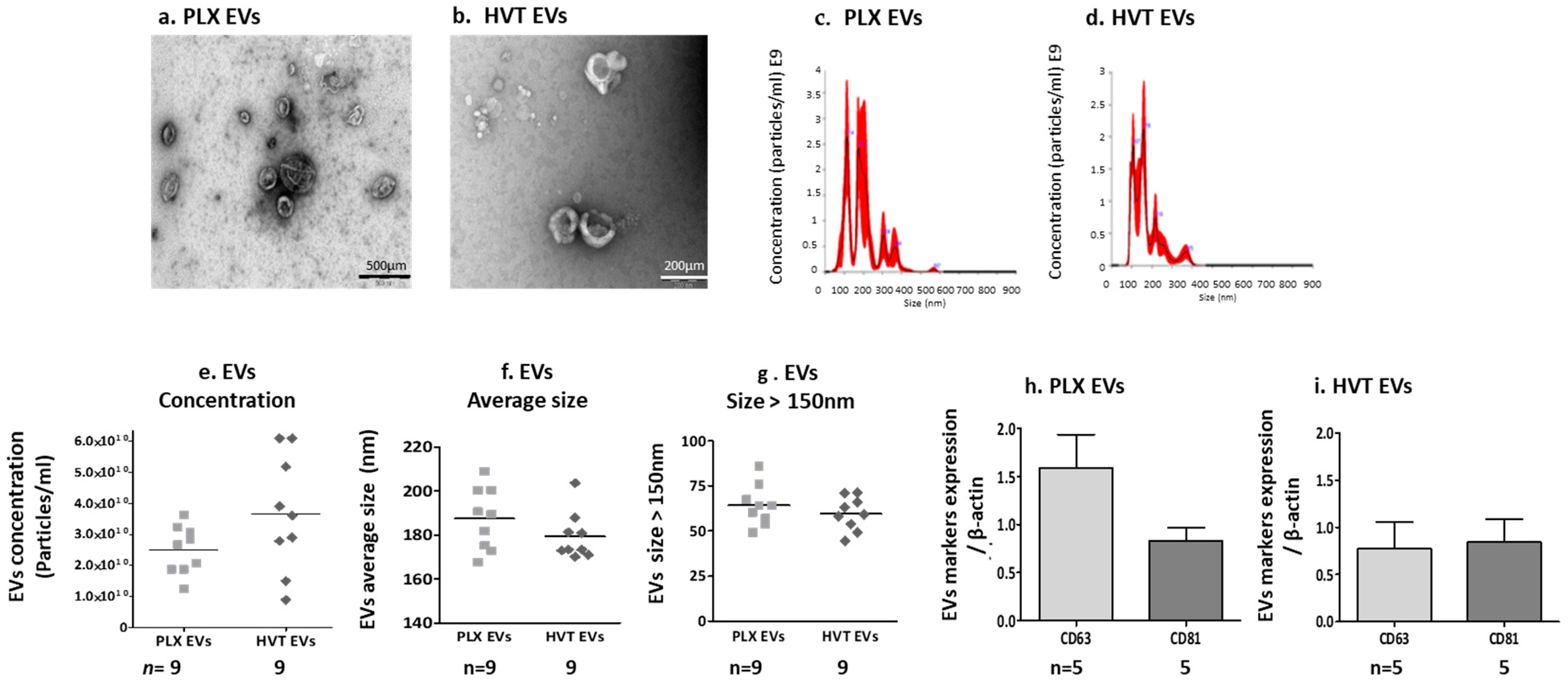
2.2.1. Placental EV Characteristics and Function
Placental EV Size and Exosome Markers
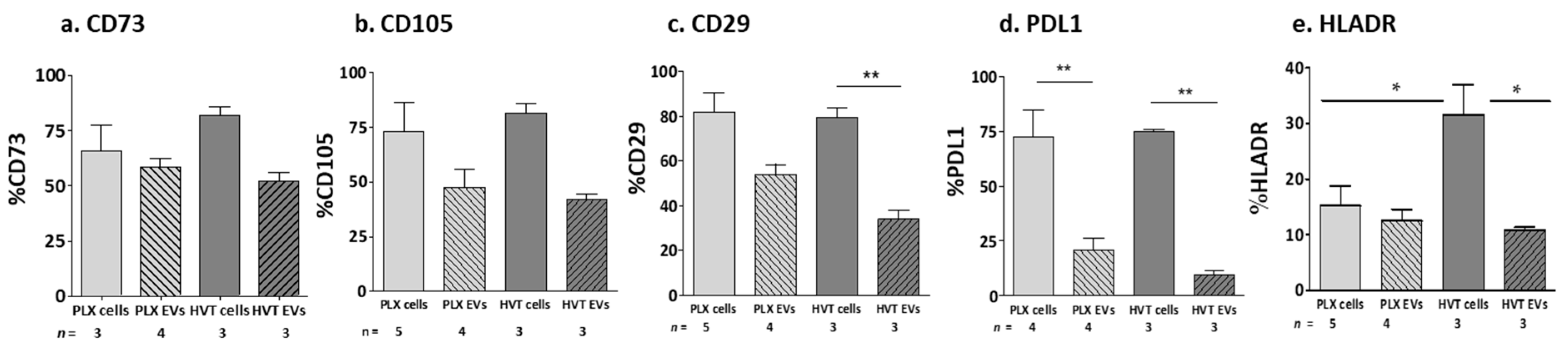
MSC Marker Expression on Placental Cells and Their EVs
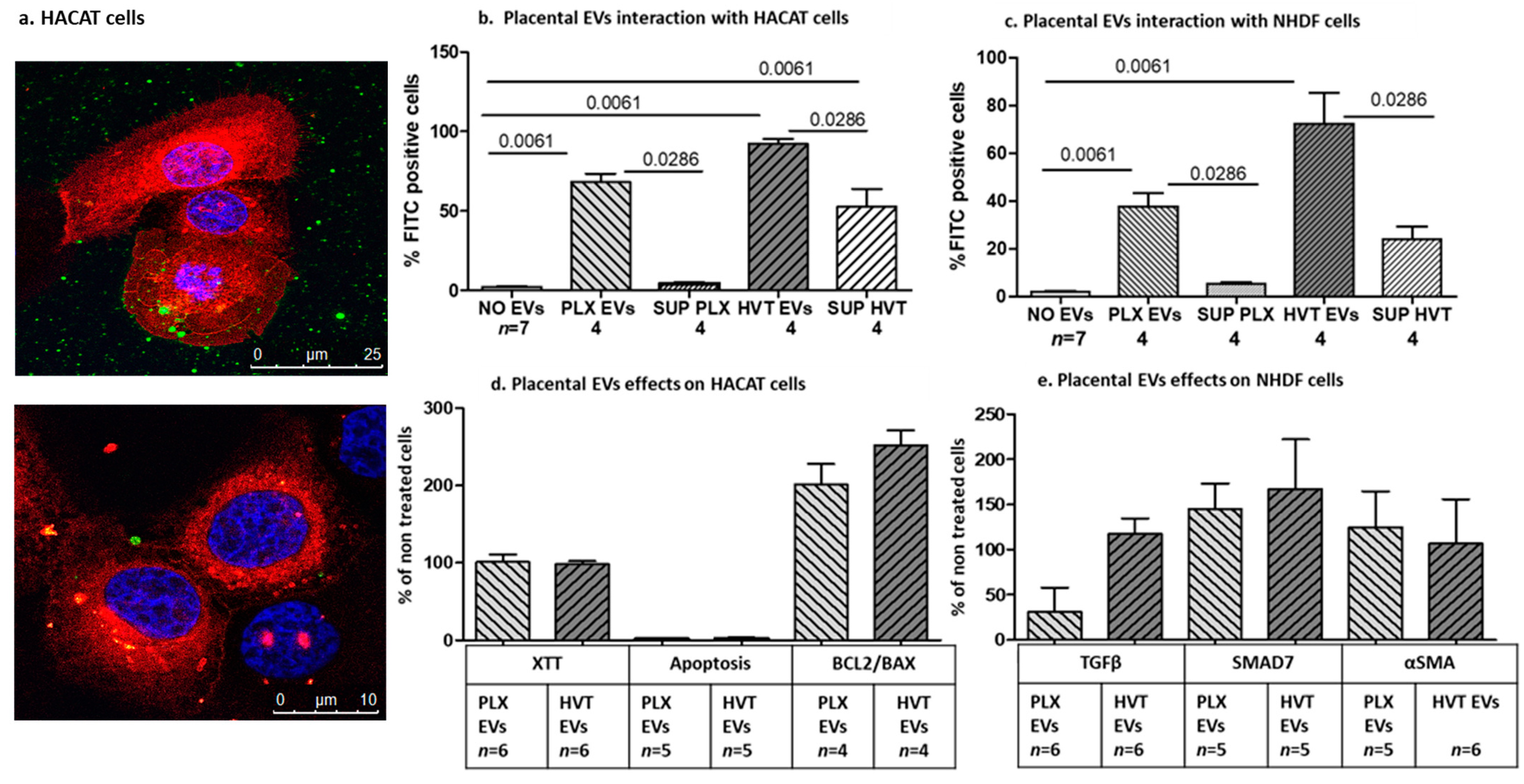
2.2.2. Interaction of Placental EVs with Dermal Cells
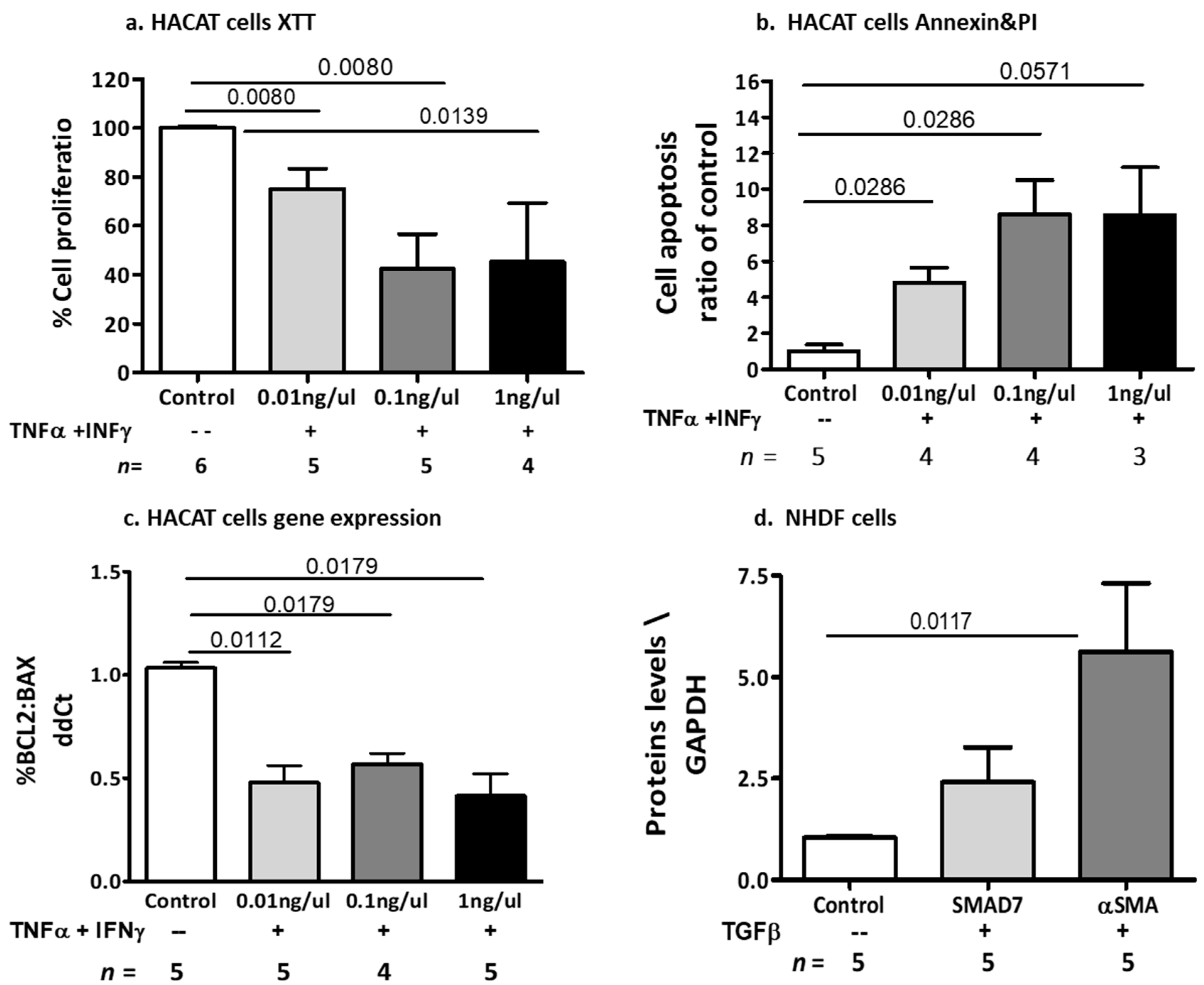
2.2.3. The Effects of Cytokine-Induced Inflammation and Fibrosis on Dermal Cell Models
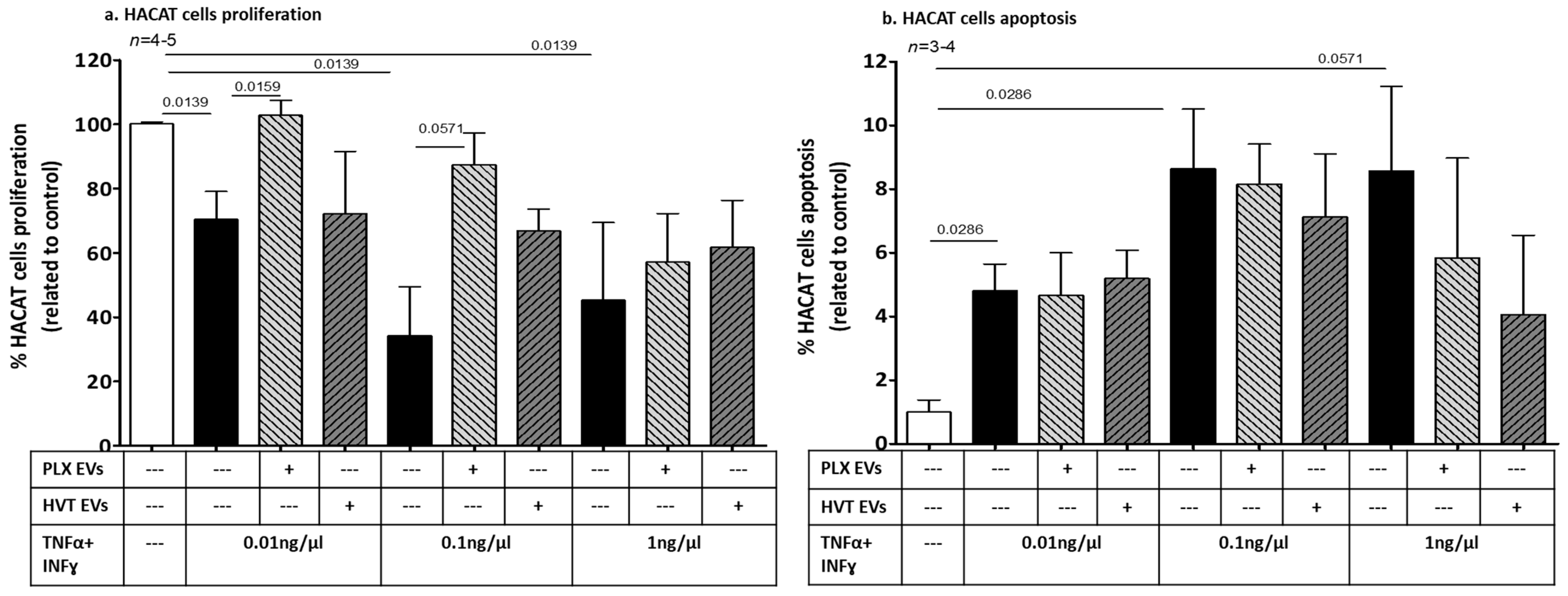
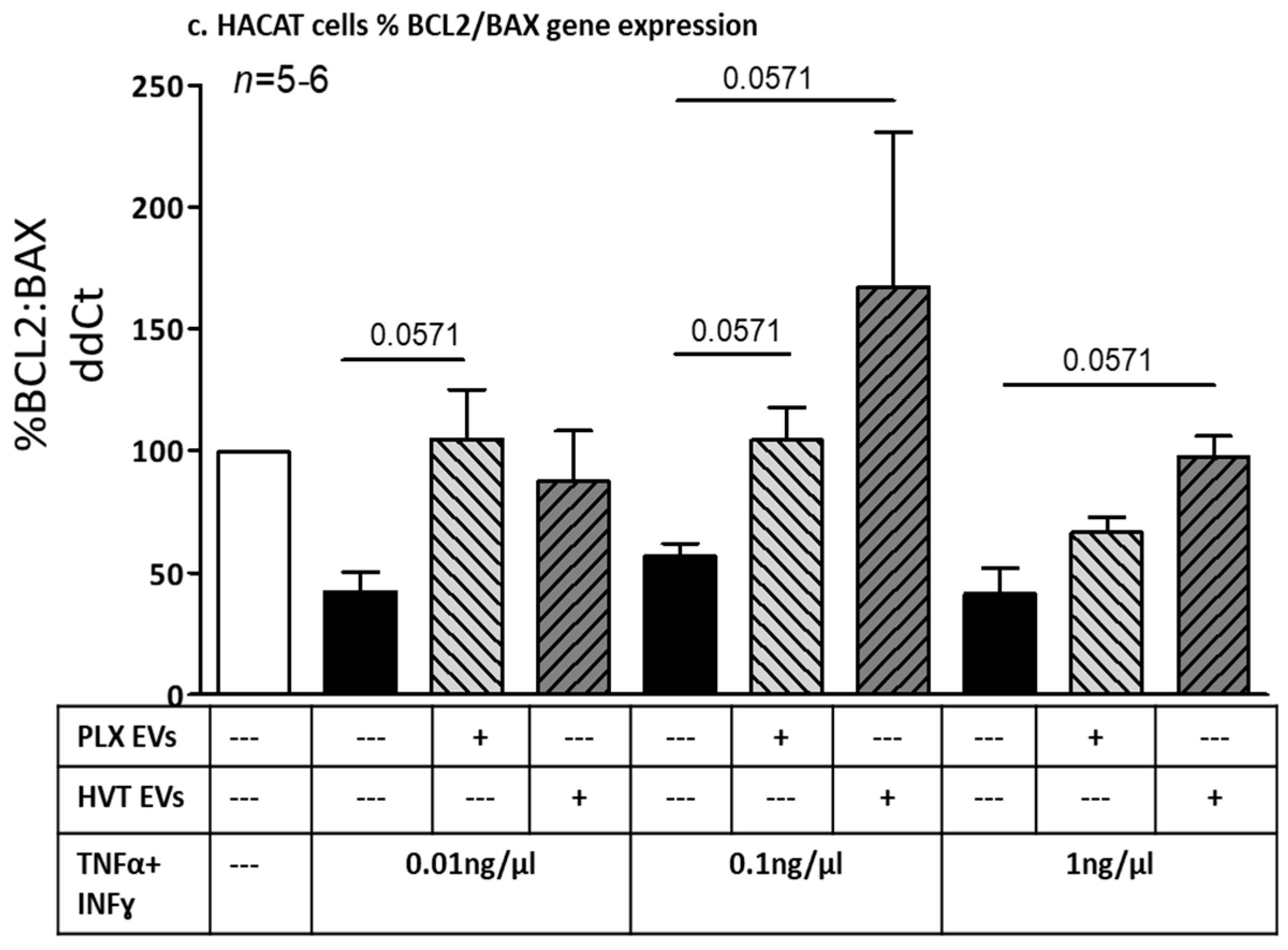
2.2.4. Effects of Placental EVs on Cytokine-Induced Keratinocyte Cell Functions
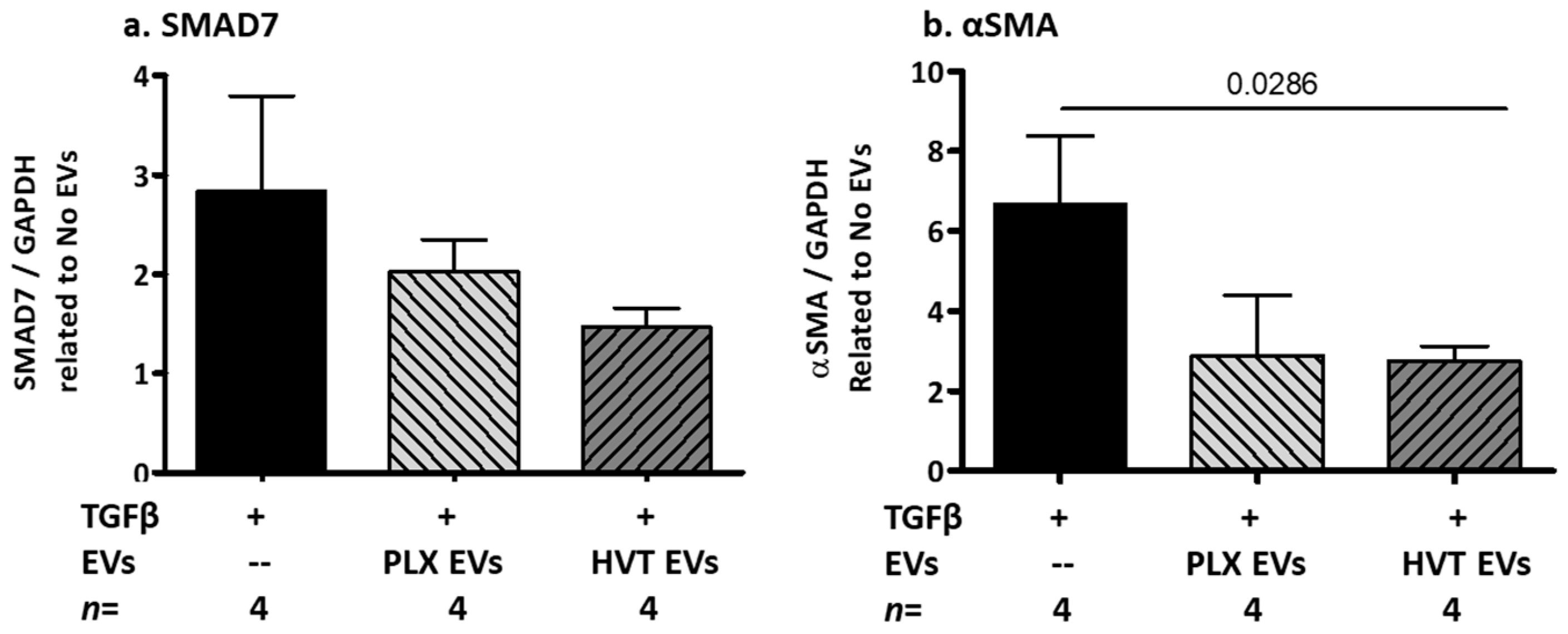
2.2.5. Effects of Placental EVs on TGF-β-Induced Fibrosis-Related Proteins in NHDF Cells
3. Discussion
4. Materials and Methods
4.1. Patient Acquisition
4.2. Patients and Healthy Controls EV Isolation and Characterization
4.2.1. Patients and Healthy Controls EV Isolation
4.2.2. EV Characterization
4.3. Cell Culture
4.3.1. Human Early-Stage Trophoblast (HVT) Cells
4.3.2. Placental-Expanded (PLX) Cells
4.3.3. Placental EV Isolation from Cell Medium
4.3.4. Primary Human Keratinocytes (HaCaT)
4.3.5. Normal Human Dermal Fibroblasts (NHDF) Cells
4.4. Placental EV and Cell Interaction
4.5. Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zeiser, R.; Blazar, B.R. Pathophysiology of Chronic Graft-versus-Host Disease and Therapeutic Targets. N. Engl. J. Med. 2017, 377, 2565–2579. [Google Scholar] [CrossRef] [PubMed]
- Socie, G. Treating chronic GVHD-induced fibrosis? Blood 2018, 131, 1396–1397. [Google Scholar] [CrossRef] [PubMed]
- Pidala, J.; Sarwal, M.; Roedder, S.; Lee, S.J. Biologic markers of chronic GVHD. Bone Marrow Transplant. 2014, 49, 324–331. [Google Scholar] [CrossRef] [PubMed]
- Hill, G.R.; Koyama, M. Cytokines and costimulation in acute graft-versus-host disease. Blood 2020, 136, 418–428. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.H.; Chen, D.Q.; Wang, Y.N.; Feng, Y.L.; Cao, G.; Vaziri, N.D.; Zhao, Y.Y. New insights into TGF-beta/Smad signaling in tissue fibrosis. Chem.-Biol. Interact. 2018, 292, 76–83. [Google Scholar] [CrossRef] [PubMed]
- Juhl, P.; Bondesen, S.; Hawkins, C.L.; Karsdal, M.A.; Bay-Jensen, A.C.; Davies, M.J.; Siebuhr, A.S. Dermal fibroblasts have different extracellular matrix profiles induced by TGF-beta, PDGF and IL-6 in a model for skin fibrosis. Sci. Rep. 2020, 10, 17300. [Google Scholar] [CrossRef]
- Shi, X.; Young, C.D.; Zhou, H.; Wang, X. Transforming Growth Factor-beta Signaling in Fibrotic Diseases and Cancer-Associated Fibroblasts. Biomolecules 2020, 10, 1666. [Google Scholar] [CrossRef]
- MacDonald, K.P.; Hill, G.R.; Blazar, B.R. Chronic graft-versus-host disease: Biological insights from preclinical and clinical studies. Blood 2017, 129, 13–21. [Google Scholar] [CrossRef]
- Socie, G.; Ritz, J. Current issues in chronic graft-versus-host disease. Blood 2014, 124, 374–384. [Google Scholar] [CrossRef]
- Sehmi, R.; Denburg, J.A. Haemopoietic mechanisms in allergic rhinitis. Clin. Exp. Allergy J. Br. Soc. Allergy Clin. Immunol. 2005, 35, 1–3. [Google Scholar] [CrossRef]
- Aly, R.M. Current state of stem cell-based therapies: An overview. Stem Cell Investig. 2020, 7, 8. [Google Scholar] [CrossRef] [PubMed]
- Talwadekar, M.D.; Kale, V.P.; Limaye, L.S. Placenta-derived mesenchymal stem cells possess better immunoregulatory properties compared to their cord-derived counterparts-a paired sample study. Sci. Rep. 2015, 5, 15784. [Google Scholar] [CrossRef] [PubMed]
- Lublin, F.D.; Bowen, J.D.; Huddlestone, J.; Kremenchutzky, M.; Carpenter, A.; Corboy, J.R.; Freedman, M.S.; Krupp, L.; Paulo, C.; Hariri, R.J.; et al. Human placenta-derived cells (PDA-001) for the treatment of adults with multiple sclerosis: A randomized, placebo-controlled, multiple-dose study. Mult. Scler. Relat. Disord. 2014, 3, 696–704. [Google Scholar] [CrossRef]
- Barzegar, M.; Wang, Y.; Eshaq, R.S.; Yun, J.W.; Boyer, C.J.; Cananzi, S.G.; White, L.A.; Chernyshev, O.; Kelley, R.E.; Minagar, A.; et al. Human placental mesenchymal stem cells improve stroke outcomes via extracellular vesicles-mediated preservation of cerebral blood flow. EBioMedicine 2021, 63, 103161. [Google Scholar] [CrossRef] [PubMed]
- Alberro, A.; Iparraguirre, L.; Fernandes, A.; Otaegui, D. Extracellular Vesicles in Blood: Sources, Effects, and Applications. Int. J. Mol. Sci. 2021, 22, 8163. [Google Scholar] [CrossRef]
- Simons, M.; Raposo, G. Exosomes–vesicular carriers for intercellular communication. Curr. Opin. Cell Biol. 2009, 21, 575–581. [Google Scholar] [CrossRef]
- Zocco, D.; Ferruzzi, P.; Cappello, F.; Kuo, W.P.; Fais, S. Extracellular vesicles as shuttles of tumor biomarkers and anti-tumor drugs. Front. Oncol. 2014, 4, 267. [Google Scholar] [CrossRef]
- Tzoran, I.; Rebibo-Sabbah, A.; Brenner, B.; Aharon, A. Disease dynamics in patients with acute myeloid leukemia: New biomarkers. Exp. Hematol. 2015, 43, 936–943. [Google Scholar] [CrossRef]
- Levin, C.; Koren, A.; Rebibo-Sabbah, A.; Koifman, N.; Brenner, B.; Aharon, A. Extracellular Vesicle Characteristics in beta-thalassemia as Potential Biomarkers for Spleen Functional Status and Ineffective Erythropoiesis. Front. Physiol. 2018, 9, 1214. [Google Scholar] [CrossRef]
- Lener, T.; Gimona, M.; Aigner, L.; Borger, V.; Buzas, E.; Camussi, G.; Chaput, N.; Chatterjee, D.; Court, F.A.; Del Portillo, H.A.; et al. Applying extracellular vesicles based therapeutics in clinical trials—An ISEV position paper. J. Extracell. Vesicles 2015, 4, 30087. [Google Scholar] [CrossRef]
- Clark, K.; Zhang, S.; Barthe, S.; Kumar, P.; Pivetti, C.; Kreutzberg, N.; Reed, C.; Wang, Y.; Paxton, Z.; Farmer, D.; et al. Placental Mesenchymal Stem Cell-Derived Extracellular Vesicles Promote Myelin Regeneration in an Animal Model of Multiple Sclerosis. Cells 2019, 8, 1497. [Google Scholar] [CrossRef] [PubMed]
- Chernoff, G. The Utilization of Human Placental Mesenchymal Stem Cell Derived Exosomes in Aging Skin: An Investigational Pilot Study. J. Surg. 2021, 6, 1388. [Google Scholar] [CrossRef]
- Shomer, E.; Katzenell, S.; Zipori, Y.; Sammour, R.N.; Isermann, B.; Brenner, B.; Aharon, A. Microvesicles of women with gestational hypertension and preeclampsia affect human trophoblast fate and endothelial function. Hypertension 2013, 62, 893–898. [Google Scholar] [CrossRef] [PubMed]
- Lahiani, A.; Zahavi, E.; Netzer, N.; Ofir, R.; Pinzur, L.; Raveh, S.; Arien-Zakay, H.; Yavin, E.; Lazarovici, P. Human placental eXpanded (PLX) mesenchymal-like adherent stromal cells confer neuroprotection to nerve growth factor (NGF)-differentiated PC12 cells exposed to ischemia by secretion of IL-6 and VEGF. Biochim. Biophys. Acta 2015, 1853, 422–430. [Google Scholar] [CrossRef] [PubMed]
- Jagasia, M.H.; Greinix, H.T.; Arora, M.; Williams, K.M.; Wolff, D.; Cowen, E.W.; Palmer, J.; Weisdorf, D.; Treister, N.S.; Cheng, G.S.; et al. National Institutes of Health Consensus Development Project on Criteria for Clinical Trials in Chronic Graft-versus-Host Disease: I. The 2014 Diagnosis and Staging Working Group report. Biol. Blood Marrow Transplant. J. Am. Soc. Blood Marrow Transplant. 2015, 21, 389–401.e1. [Google Scholar] [CrossRef]
- Crossland, R.E.; Perutelli, F.; Bogunia-Kubik, K.; Mooney, N.; Milutin Gasperov, N.; Pucic-Bakovic, M.; Greinix, H.; Weber, D.; Holler, E.; Pulanic, D.; et al. Potential Novel Biomarkers in Chronic Graft-Versus-Host Disease. Front. Immunol. 2020, 11, 602547. [Google Scholar] [CrossRef]
- Perez-Simon, J.A.; Sanchez-Abarca, I.; Diez-Campelo, M.; Caballero, D.; San Miguel, J. Chronic graft-versus-host disease: Pathogenesis and clinical management. Drugs 2006, 66, 1041–1057. [Google Scholar] [CrossRef]
- Banovic, T.; MacDonald, K.P.; Morris, E.S.; Rowe, V.; Kuns, R.; Don, A.; Kelly, J.; Ledbetter, S.; Clouston, A.D.; Hill, G.R. TGF-beta in allogeneic stem cell transplantation: Friend or foe? Blood 2005, 106, 2206–2214. [Google Scholar] [CrossRef]
- Marinelli Busilacchi, E.; Costantini, A.; Mancini, G.; Tossetta, G.; Olivieri, J.; Poloni, A.; Viola, N.; Butini, L.; Campanati, A.; Goteri, G.; et al. Nilotinib Treatment of Patients Affected by Chronic Graft-versus-Host Disease Reduces Collagen Production and Skin Fibrosis by Downmodulating the TGF-beta and p-SMAD Pathway. Biol. Blood Marrow Transplant. J. Am. Soc. Blood Marrow Transplant. 2020, 26, 823–834. [Google Scholar] [CrossRef]
- Hu, Y.; He, J.; He, L.; Xu, B.; Wang, Q. Expression and function of Smad7 in autoimmune and inflammatory diseases. J. Mol. Med. 2021, 99, 1209–1220. [Google Scholar] [CrossRef]
- Dadashzadeh, E.; Saghaeian Jazi, M.; Abdolahi, N.; Mohammadi, S.; Saeidi, M. Comparison of a Suggested Model of Fibrosis in Human Dermal Fibroblasts by Serum from Systemic Sclerosis Patients with Transforming Growth Factor beta Induced in vitro Model. Int. J. Mol. Cell. Med. 2022, 11, 31–40. [Google Scholar] [CrossRef]
- Wen, F.Q.; Kohyama, T.; Skold, C.M.; Zhu, Y.K.; Liu, X.; Romberger, D.J.; Stoner, J.; Rennard, S.I. Glucocorticoids modulate TGF-beta production. Inflammation 2002, 26, 279–290. [Google Scholar] [CrossRef]
- Ding, D.C.; Shyu, W.C.; Lin, S.Z. Mesenchymal stem cells. Cell Transplant. 2011, 20, 5–14. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Chen, S.; Yang, P.; Cao, H.; Li, L. The role of mesenchymal stem cells in hematopoietic stem cell transplantation: Prevention and treatment of graft-versus-host disease. Stem Cell Res. Ther. 2019, 10, 182. [Google Scholar] [CrossRef]
- Sher, N.; Ofir, R. Placenta-Derived Adherent Stromal Cell Therapy for Hematopoietic Disorders: A Case Study of PLX-R18. Cell Transplant. 2018, 27, 140–150. [Google Scholar] [CrossRef] [PubMed]
- Malek, A.; Bersinger, N.A. Human placental stem cells: Biomedical potential and clinical relevance. J. Stem Cells 2011, 6, 75–92. [Google Scholar] [PubMed]
- Lalu, M.M.; McIntyre, L.; Pugliese, C.; Fergusson, D.; Winston, B.W.; Marshall, J.C.; Granton, J.; Stewart, D.J.; Canadian Critical Care Trials, G. Safety of cell therapy with mesenchymal stromal cells (SafeCell): A systematic review and meta-analysis of clinical trials. PLoS ONE 2012, 7, e47559. [Google Scholar] [CrossRef]
- Zhu, X.; Badawi, M.; Pomeroy, S.; Sutaria, D.S.; Xie, Z.; Baek, A.; Jiang, J.; Elgamal, O.A.; Mo, X.; Perle, K.; et al. Comprehensive toxicity and immunogenicity studies reveal minimal effects in mice following sustained dosing of extracellular vesicles derived from HEK293T cells. J. Extracell. Vesicles 2017, 6, 1324730. [Google Scholar] [CrossRef]
- Elsharkasy, O.M.; Nordin, J.Z.; Hagey, D.W.; de Jong, O.G.; Schiffelers, R.M.; Andaloussi, S.E.; Vader, P. Extracellular vesicles as drug delivery systems: Why and how? Adv. Drug Deliv. Rev. 2020, 159, 332–343. [Google Scholar] [CrossRef]
- Contini, P.; Murdaca, G.; Puppo, F.; Negrini, S. HLA-G Expressing Immune Cells in Immune Mediated Diseases. Front. Immunol. 2020, 11, 1613. [Google Scholar] [CrossRef]
- Stamou, P.; Marioli, D.; Patmanidi, A.L.; Sgourou, A.; Vittoraki, A.; Theofani, E.; Pierides, C.; Taraviras, S.; Costeas, P.A.; Spyridonidis, A. Simple in vitro generation of human leukocyte antigen-G-expressing T-regulatory cells through pharmacological hypomethylation for adoptive cellular immunotherapy against graft-versus-host disease. Cytotherapy 2017, 19, 521–530. [Google Scholar] [CrossRef] [PubMed]
- Herishanu, Y.; Avivi, I.; Aharon, A.; Shefer, G.; Levi, S.; Bronstein, Y.; Morales, M.; Ziv, T.; Shorer Arbel, Y.; Scarfo, L.; et al. Efficacy of the BNT162b2 mRNA COVID-19 vaccine in patients with chronic lymphocytic leukemia. Blood 2021, 137, 3165–3173. [Google Scholar] [CrossRef] [PubMed]
- Aharon, A.; Horn, G.; Bar-Lev, T.H.; Zagagi Yohay, E.; Waks, T.; Levin, M.; Deshet Unger, N.; Avivi, I.; Globerson Levin, A. Extracellular Vesicles Derived from Chimeric Antigen Receptor-T Cells: A Potential Therapy for Cancer. Hum. Gene Ther. 2021, 32, 1224–1241. [Google Scholar] [CrossRef]
- Aharon, A.; Spector, P.; Ahmad, R.S.; Horrany, N.; Sabbach, A.; Brenner, B.; Aharon-Peretz, J. Extracellular Vesicles of Alzheimer’s Disease Patients as a Biomarker for Disease Progression. Mol. Neurobiol. 2020, 57, 4156–4169. [Google Scholar] [CrossRef] [PubMed]
- Thery, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): A position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J. Extracell. Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef]
- Aharon, A.; Rebibo-Sabbah, A.; Ahmad, R.S.; Dangot, A.; Bar-Lev, T.H.; Brenner, B.; Cohen, A.H.; David, C.B.; Weiner, Z.; Solt, I. Associations of maternal and placental extracellular vesicle miRNA with preeclampsia. Front. Cell Dev. Biol. 2023, 11, 1080419. [Google Scholar] [CrossRef] [PubMed]
- Nunez Lopez, Y.O.; Coen, P.M.; Goodpaster, B.H.; Seyhan, A.A. Gastric bypass surgery with exercise alters plasma microRNAs that predict improvements in cardiometabolic risk. Int. J. Obes. 2017, 41, 1121–1130. [Google Scholar] [CrossRef]
- Peng, G.; Tang, Y.; Cowan, T.M.; Zhao, H.; Scharfe, C. Timing of Newborn Blood Collection Alters Metabolic Disease Screening Performance. Front. Pediatr. 2020, 8, 623184. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Domain | cGVHD Patients (n = 14) |
|---|---|
| Age, median (range) | 56 (23–72) |
| Sex, n female (%) | 7 (44%) |
| Ethnicity, n (%) | |
| Ashkenazi Jewish | 8 (50%) |
| Sephardic Jewish | 6 (38%) |
| Arab | 2 (12%) |
| Time from AlloHCT (months), median (range) | |
| Time from onset of chronic GVHD (months), median (range) | 60 (34–134) |
| Chronic GVHD severity, n severe (%) | 16 (100%) |
| Previous lines of treatment, median, range | 5 (3–5) |
| Organs involved, n (%) | |
| Mouth | 13 (81%) |
| Eyes | 15 (94%) |
| Skin | 16 (100%) |
| Fascia | 11 (69%) |
| Joints | 9 (56%) |
| Lungs | 11 (69%) |
| Gastro-intestinal | 5 (31%) |
| Liver | 4 (25%) |
| Medications at the time of plasma sampling | |
| Prednisone | 10 (71%) |
| Tacrolimus, Cyclosporin | 2 (14%) |
| JAK-STAT inhibitor (Ruxolitinib) | 6 (43%) |
| Tyrosine kinase inhibitor (Imatinib) | 2 (14%) |
| Extra corporal photopheresis | 1 (7%) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zavaro, M.; Dangot, A.; Bar-Lev, T.H.; Amit, O.; Avivi, I.; Ram, R.; Aharon, A. The Role of Extracellular Vesicles (EVs) in Chronic Graft vs. Host Disease, and the Potential Function of Placental Cell-Derived EVs as a Therapeutic Tool. Int. J. Mol. Sci. 2023, 24, 8126. https://doi.org/10.3390/ijms24098126
Zavaro M, Dangot A, Bar-Lev TH, Amit O, Avivi I, Ram R, Aharon A. The Role of Extracellular Vesicles (EVs) in Chronic Graft vs. Host Disease, and the Potential Function of Placental Cell-Derived EVs as a Therapeutic Tool. International Journal of Molecular Sciences. 2023; 24(9):8126. https://doi.org/10.3390/ijms24098126
Chicago/Turabian StyleZavaro, Mor, Ayelet Dangot, Tali Hana Bar-Lev, Odelia Amit, Irit Avivi, Ron Ram, and Anat Aharon. 2023. "The Role of Extracellular Vesicles (EVs) in Chronic Graft vs. Host Disease, and the Potential Function of Placental Cell-Derived EVs as a Therapeutic Tool" International Journal of Molecular Sciences 24, no. 9: 8126. https://doi.org/10.3390/ijms24098126