
Tracking Adaptive Pathways of Invasive Insects: Novel Insight from Genomics

Abstract
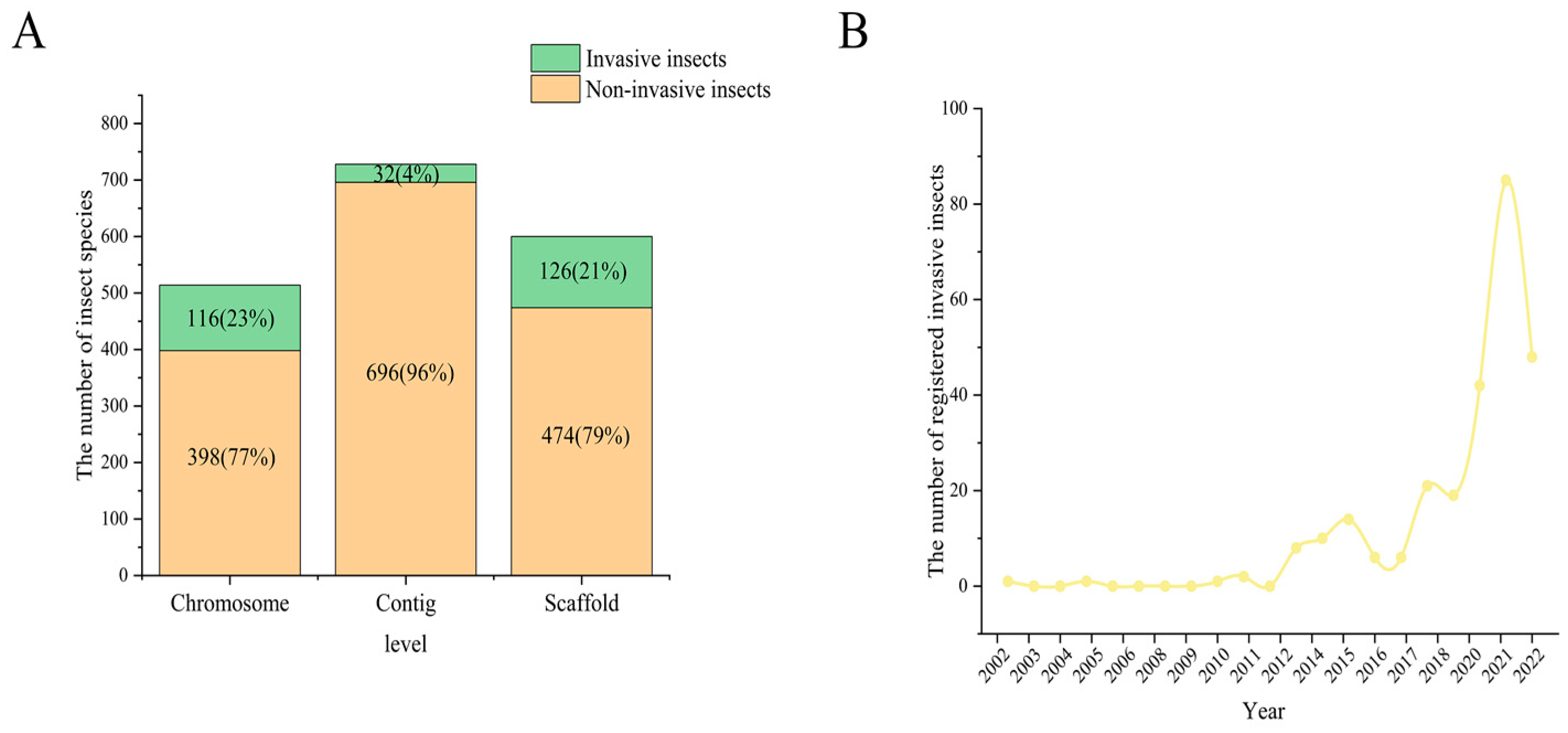
:1. Introduction
2. Whole-Genome Sequencing of Invasive Insects
3. Whole-Genome Sequencing Reveals Insect Invasion Mechanisms
3.1. Rapid Genetic Variation and Evolution of Invasive Populations
3.2. Invasion History and Dispersal Path
3.3. Rapid Adaptation to Different Host Plant Ranges
3.4. Strong Environmental Adaptation
3.5. Development of Insecticide Resistance
3.6. Synergistic Harm Caused by Invasive Insects and Endosymbiotic Bacteria
4. Using Genome Sequencing for Developing New Strategies to Control Invasive Insects
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ielmini, M.R.; Sankaran, K. Invasive Alien Species: A Prodigious Global Threat in the Anthropocene. Invasive Alien Species Obs. Issues Around World 2021, 1, 1–79. [Google Scholar]
- Early, R.; Bradley, B.A.; Dukes, J.S.; Lawler, J.J.; Olden, J.D.; Blumenthal, D.M.; Gonzalez, P.; Grosholz, E.D.; Ibañez, I.; Miller, L.P. Global threats from invasive alien species in the twenty-first century and national response capacities. Nat. Commun. 2016, 7, 12485. [Google Scholar] [CrossRef] [PubMed]
- Paini, D.R.; Sheppard, A.W.; Cook, D.C.; De Barro, P.J.; Worner, S.P.; Thomas, M.B. Global threat to agriculture from invasive species. Proc. Natl. Acad. Sci. USA 2016, 113, 7575–7579. [Google Scholar] [CrossRef] [PubMed]
- Venette, R.C.; Hutchison, W.D. Invasive insect species: Global challenges, strategies & opportunities. Front. Insect Sci. 2021, 1, 650520. [Google Scholar]
- Seebens, H.; Blackburn, T.M.; Dyer, E.E.; Genovesi, P.; Hulme, P.E.; Jeschke, J.M.; Pagad, S.; Pyšek, P.; Winter, M.; Arianoutsou, M. No saturation in the accumulation of alien species worldwide. Nat. Commun. 2017, 8, 14435. [Google Scholar] [CrossRef]
- Bradshaw, C.J.; Leroy, B.; Bellard, C.; Roiz, D.; Albert, C.; Fournier, A.; Barbet-Massin, M.; Salles, J.-M.; Simard, F.; Courchamp, F. Massive yet grossly underestimated global costs of invasive insects. Nat. Commun. 2016, 7, 12986. [Google Scholar] [CrossRef] [PubMed]
- Hinds, W.; Dew, J. The grass worm or fall army worm. Bulletin 1915, 186, 59–92. [Google Scholar]
- Mallapur, C.P.; Naik, A.K.; Hagari, S.; Prabhu, S.T.; Patil, R.K. Status of alien pest fall armyworm, Spodoptera frugiperda (J E Smith) on maize in Northern Karnataka. AkiNik Publ. 2018, 6, 432–436. [Google Scholar]
- Li, X.J.; Wu, M.F.; Ma, J.; Gao, B.Y.; Wu, Q.L.; Chen, A.D.; Liu, J.; Jiang, Y.Y.; Zhai, B.P.; Early, R. Prediction of migratory routes of the invasive fall armyworm in eastern China using a trajectory analytical approach. Pest Manag. Sci. 2020, 76, 454–463. [Google Scholar] [CrossRef]
- Han, P.; Bayram, Y.; Shaltiel-Harpaz, L.; Sohrabi, F.; Saji, A.; Esenali, U.T.; Jalilov, A.; Ali, A.; Shashank, P.R.; Ismoilov, K. Tuta absoluta continues to disperse in Asia: Damage, ongoing management and future challenges. J. Pest Sci. 2019, 92, 1317–1327. [Google Scholar] [CrossRef]
- Opoku, J.; Kleczewski, N.M.; Hamby, K.A.; Herbert, D.A.; Malone, S.; Mehl, H.L. Relationship between invasive Brown Marmorated Stink Bug (Halyomorpha halys) and fumonisin contamination of field corn in the mid-Atlantic US. Plant Dis. 2019, 103, 1189–1195. [Google Scholar] [CrossRef]
- Rezende, H.R.; Romano, C.M.; Claro, I.M.; Caleiro, G.S.; Vicente, C.R. First report of Aedes albopictus infected by Dengue and Zika virus in a rural outbreak in Brazil. PLoS ONE 2020, 15, e0229847. [Google Scholar] [CrossRef]
- Comeault, A.A.; Wang, J.; Tittes, S.; Isbell, K.; Ingley, S.; Hurlbert, A.H.; Matute, D.R. Genetic diversity and thermal performance in invasive and native populations of African fig flies. Mol. Biol. Evol. 2020, 37, 1893–1906. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Xing, Z.; Ma, T.; Xu, D.; Li, Y.; Lei, Z.; Gao, Y. Competitive interaction between Frankliniella occidentalis and locally present thrips species: A global review. J. Pest Sci. 2021, 94, 5–16. [Google Scholar] [CrossRef]
- Xiao, H.; Ye, X.; Xu, H.; Mei, Y.; Yang, Y.; Chen, X.; Yang, Y.; Liu, T.; Yu, Y.; Yang, W. The genetic adaptations of fall armyworm Spodoptera frugiperda facilitated its rapid global dispersal and invasion. Mol. Ecol. Resour. 2020, 20, 1050–1068. [Google Scholar] [CrossRef]
- Schoville, S.D.; Chen, Y.H.; Andersson, M.N.; Benoit, J.B.; Bhandari, A.; Bowsher, J.H.; Brevik, K.; Cappelle, K.; Chen, M.-J.M.; Childers, A.K. A model species for agricultural pest genomics: The genome of the Colorado potato beetle, Leptinotarsa decemlineata (Coleoptera: Chrysomelidae). Sci. Rep. 2018, 8, 1931. [Google Scholar] [CrossRef]
- Wan, F.; Yin, C.; Tang, R.; Chen, M.; Wu, Q.; Huang, C.; Qian, W.; Rota-Stabelli, O.; Yang, N.; Wang, S. A chromosome-level genome assembly of Cydia pomonella provides insights into chemical ecology and insecticide resistance. Nat. Commun. 2019, 10, 4237. [Google Scholar] [CrossRef] [PubMed]
- Mcgeoch, M.A.; Lythe, M.J.; Henriksen, M.V.; Mcgrannachan, C.M. Environmental impact classification for alien insects: A review of mechanisms and their biodiversity outcomes. Curr. Opin. Insect Sci. 2015, 12, 46–53. [Google Scholar] [CrossRef]
- Harrison, M.C.; Jongepier, E.; Robertson, H.M.; Arning, N.; Bitard-Feildel, T.; Chao, H.; Childers, C.P.; Dinh, H.; Doddapaneni, H.; Dugan, S. Hemimetabolous genomes reveal molecular basis of termite eusociality. Nat. Ecol. Evol. 2018, 2, 557–566. [Google Scholar] [CrossRef]
- Zheng, X.; Wang, L.; Liu, Y.; Yang, Z.; Li, F.; Yan, L.; Shen, Y.; Yue, B.; Zhou, C. Improved genome assembly provides new insights into the environmental adaptation of the American cockroach, Periplaneta americana. Arch. Insect Biochem. Physiol. 2022, 111, e21956. [Google Scholar] [CrossRef]
- Liu, Y.; Henkel, J.; Beaurepaire, A.; Evans, J.D.; Neumann, P.; Huang, Q. Comparative genomics suggests local adaptations in the invasive small hive beetle. Ecol. Evol. 2021, 11, 15780–15791. [Google Scholar] [CrossRef]
- Evans, J.D.; McKenna, D.; Scully, E.; Cook, S.C.; Dainat, B.; Egekwu, N.; Grubbs, N.; Lopez, D.; Lorenzen, M.D.; Reyna, S.M. Genome of the small hive beetle (Aethina tumida, Coleoptera: Nitidulidae), a worldwide parasite of social bee colonies, provides insights into detoxification and herbivory. Gigascience 2018, 7, giy138. [Google Scholar] [CrossRef]
- Xie, W.; Yang, X.; Chen, C.; Yang, Z.; Guo, L.; Wang, D.; Huang, J.; Zhang, H.; Wen, Y.; Zhao, J. The invasive MED/Q Bemisia tabaci genome: A tale of gene loss and gene gain. BMC Genom. 2018, 19, 68. [Google Scholar] [CrossRef]
- Filipović, I.; Rašić, G.; Hereward, J.; Gharuka, M.; Devine, G.J.; Furlong, M.J.; Etebari, K. A high-quality de novo genome assembly based on nanopore sequencing of a wild-caught coconut rhinoceros beetle (Oryctes rhinoceros). BMC Genom. 2022, 23, 426. [Google Scholar] [CrossRef] [PubMed]
- Andersson, M.N.; Keeling, C.I.; Mitchell, R.F. Genomic content of chemosensory genes correlates with host range in wood-boring beetles (Dendroctonus ponderosae, Agrilus planipennis, and Anoplophora glabripennis). BMC Genom. 2019, 20, 690. [Google Scholar] [CrossRef] [PubMed]
- Cui, M.; Wu, Y.; Javal, M.; Giguère, I.; Roux, G.; Andres, J.A.; Keena, M.; Shi, J.; Wang, B.; Braswell, E. Genome-scale phylogeography resolves the native population structure of the Asian longhorned beetle, Anoplophora glabripennis (Motschulsky). Evol. Appl. 2022, 15, 934. [Google Scholar] [CrossRef]
- McKenna, D.D.; Scully, E.D.; Pauchet, Y.; Hoover, K.; Kirsch, R.; Geib, S.M.; Mitchell, R.F.; Waterhouse, R.M.; Ahn, S.-J.; Arsala, D. Genome of the Asian longhorned beetle (Anoplophora glabripennis), a globally significant invasive species, reveals key functional and evolutionary innovations at the beetle–plant interface. Genome Biol. 2016, 17, 227. [Google Scholar] [CrossRef]
- Liu, Z.; Xing, L.; Huang, W.; Liu, B.; Wan, F.; Raffa, K.F.; Hofstetter, R.W.; Qian, W.; Sun, J. Chromosome-level genome assembly and population genomic analyses provide insights into adaptive evolution of the red turpentine beetle, Dendroctonus valens. BMC Biol. 2022, 20, 190. [Google Scholar] [CrossRef] [PubMed]
- Coates, B.S.; Alves, A.P.; Wang, H.; Walden, K.K.; French, B.W.; Miller, N.J.; Abel, C.A.; Robertson, H.M.; Sappington, T.W.; Siegfried, B.D. Distribution of genes and repetitive elements in the Diabrotica virgifera virgifera genome estimated using BAC sequencing. J. Biomed. Biotechnol. 2012, 2012, 604076. [Google Scholar] [CrossRef]
- Chen, M.; Mei, Y.; Chen, X.; Chen, X.; Xiao, D.; He, K.; Li, Q.; Wu, M.; Wang, S.; Zhang, F. A chromosome-level assembly of the harlequin ladybird Harmonia axyridis as a genomic resource to study beetle and invasion biology. Mol. Ecol. Resour. 2021, 21, 1318–1332. [Google Scholar] [CrossRef]
- Vega, F.E.; Brown, S.M.; Chen, H.; Shen, E.; Nair, M.B.; Ceja-Navarro, J.A.; Brodie, E.L.; Infante, F.; Dowd, P.F.; Pain, A. Draft genome of the most devastating insect pest of coffee worldwide: The coffee berry borer, Hypothenemus hampei. Sci. Rep. 2015, 5, 12525. [Google Scholar] [CrossRef] [PubMed]
- Matthews, B.J.; Dudchenko, O.; Kingan, S.B.; Koren, S.; Antoshechkin, I.; Crawford, J.E.; Glassford, W.J.; Herre, M.; Redmond, S.N.; Rose, N.H. Improved reference genome of Aedes aegypti informs arbovirus vector control. Nature 2018, 563, 501–507. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.-G.; Jiang, X.; Gu, J.; Xu, M.; Wu, Y.; Deng, Y.; Zhang, C.; Bonizzoni, M.; Dermauw, W.; Vontas, J. Genome sequence of the Asian Tiger mosquito, Aedes albopictus, reveals insights into its biology, genetics, and evolution. Proc. Natl. Acad. Sci. USA 2015, 112, E5907–E5915. [Google Scholar] [CrossRef]
- Dritsou, V.; Topalis, P.; Windbichler, N.; Simoni, A.; Hall, A.; Lawson, D.; Hinsley, M.; Hughes, D.; Napolioni, V.; Crucianelli, F. A draft genome sequence of an invasive mosquito: An Italian Aedes albopictus. Pathog. Glob. Health 2015, 109, 207–220. [Google Scholar] [CrossRef]
- Miller, J.R.; Koren, S.; Dilley, K.A.; Puri, V.; Brown, D.M.; Harkins, D.M.; Thibaud-Nissen, F.; Rosen, B.; Chen, X.-G.; Tu, Z. Analysis of the Aedes albopictus C6/36 genome provides insight into cell line utility for viral propagation. Gigascience 2018, 7, gix135. [Google Scholar] [CrossRef]
- Chakraborty, M.; Ramaiah, A.; Adolfi, A.; Halas, P.; Kaduskar, B.; Ngo, L.T.; Jayaprasad, S.; Paul, K.; Whadgar, S.; Srinivasan, S. Hidden genomic features of an invasive malaria vector, Anopheles stephensi, revealed by a chromosome-level genome assembly. BMC Biol. 2021, 19, 28. [Google Scholar] [CrossRef] [PubMed]
- Sim, S.B.; Geib, S.M. A chromosome-scale assembly of the Bactrocera cucurbitae genome provides insight to the genetic basis of white pupae. G3 Genes Genomes Genet. 2017, 7, 1927–1940. [Google Scholar] [CrossRef]
- Jiang, F.; Liang, L.; Wang, J.; Zhu, S. Chromosome-level genome assembly of Bactrocera dorsalis reveals its adaptation and invasion mechanisms. Commun. Biol. 2022, 5, 25. [Google Scholar] [CrossRef]
- Bayega, A.; Djambazian, H.; Tsoumani, K.T.; Gregoriou, M.-E.; Sagri, E.; Drosopoulou, E.; Mavragani-Tsipidou, P.; Giorda, K.; Tsiamis, G.; Bourtzis, K. De novo assembly of the olive fruit fly (Bactrocera oleae) genome with linked-reads and long-read technologies minimizes gaps and provides exceptional Y chromosome assembly. BMC Genom. 2020, 21, 259. [Google Scholar] [CrossRef]
- Gilchrist, A.S.; Shearman, D.C.; Frommer, M.; Raphael, K.A.; Deshpande, N.P.; Wilkins, M.R.; Sherwin, W.B.; Sved, J.A. The draft genome of the pest tephritid fruit fly Bactrocera tryoni: Resources for the genomic analysis of hybridising species. BMC Genom. 2014, 15, 1153. [Google Scholar] [CrossRef]
- Papanicolaou, A.; Schetelig, M.F.; Arensburger, P.; Atkinson, P.W.; Benoit, J.B.; Bourtzis, K.; Castanera, P.; Cavanaugh, J.P.; Chao, H.; Childers, C. The whole genome sequence of the Mediterranean fruit fly, Ceratitis capitata (Wiedemann), reveals insights into the biology and adaptive evolution of a highly invasive pest species. Genome Biol. 2016, 17, 192. [Google Scholar] [CrossRef] [PubMed]
- Olazcuaga, L.; Loiseau, A.; Parrinello, H.; Paris, M.; Fraimout, A.; Guedot, C.; Diepenbrock, L.M.; Kenis, M.; Zhang, J.; Chen, X. A whole-genome scan for association with invasion success in the fruit fly Drosophila suzukii using contrasts of allele frequencies corrected for population structure. Mol. Biol. Evol. 2020, 37, 2369–2385. [Google Scholar] [CrossRef]
- Lewald, K.M.; Abrieux, A.; Wilson, D.A.; Lee, Y.; Conner, W.R.; Andreazza, F.; Beers, E.H.; Burrack, H.J.; Daane, K.M.; Diepenbrock, L. Population genomics of Drosophila suzukii reveal longitudinal population structure and signals of migrations in and out of the continental United States. G3 Genes Genomes Genet. 2021, 11, jkab343. [Google Scholar] [CrossRef] [PubMed]
- Vicoso, B.; Bachtrog, D. Numerous transitions of sex chromosomes in Diptera. PLoS Biol. 2015, 13, e1002078. [Google Scholar] [CrossRef] [PubMed]
- Romine, M.G.; Knutie, S.A.; Crow, C.M.; Vaziri, G.J.; Chaves, J.A.; Koop, J.A.; Lamichhaney, S. The genome sequence of the avian vampire fly (Philornis downsi), an invasive nest parasite of Darwin’s finches in Galápagos. G3 Genes Genomes Genet. 2022, 12, jkab414. [Google Scholar] [CrossRef] [PubMed]
- Consortium, I.A.G. Genome sequence of the pea aphid Acyrthosiphon pisum. PLoS Biol. 2010, 8, e1000313. [Google Scholar]
- Giordano, R.; Donthu, R.K.; Zimin, A.V.; Chavez, I.C.J.; Gabaldon, T.; van Munster, M.; Hon, L.; Hall, R.; Badger, J.H.; Nguyen, M. Soybean aphid biotype 1 genome: Insights into the invasive biology and adaptive evolution of a major agricultural pest. Insect Biochem. Mol. Biol. 2020, 120, 103334. [Google Scholar] [CrossRef] [PubMed]
- Wenger, J.A.; Cassone, B.J.; Legeai, F.; Johnston, J.S.; Bansal, R.; Yates, A.D.; Coates, B.S.; Pavinato, V.A.; Michel, A. Whole genome sequence of the soybean aphid, Aphis glycines. Insect Biochem. Mol. Biol. 2020, 123, 102917. [Google Scholar] [CrossRef] [PubMed]
- Hussain, S.; Farooq, M.; Malik, H.J.; Amin, I.; Scheffler, B.E.; Scheffler, J.A.; Liu, S.-S.; Mansoor, S. Whole genome sequencing of Asia II 1 species of whitefly reveals that genes involved in virus transmission and insecticide resistance have genetic variances between Asia II 1 and MEAM1 species. BMC Genom. 2019, 20, 507. [Google Scholar] [CrossRef]
- Chen, W.; Hasegawa, D.K.; Kaur, N.; Kliot, A.; Pinheiro, P.V.; Luan, J.; Stensmyr, M.C.; Zheng, Y.; Liu, W.; Sun, H. The draft genome of whitefly Bemisia tabaci MEAM1, a global crop pest, provides novel insights into virus transmission, host adaptation, and insecticide resistance. BMC Biol. 2016, 14, 110. [Google Scholar] [CrossRef]
- Rosenfeld, J.A.; Reeves, D.; Brugler, M.R.; Narechania, A.; Simon, S.; Durrett, R.; Foox, J.; Shianna, K.; Schatz, M.C.; Gandara, J. Genome assembly and geospatial phylogenomics of the bed bug Cimex lectularius. Nat. Commun. 2016, 7, 10164. [Google Scholar] [CrossRef] [PubMed]
- Rispe, C.; Legeai, F.; Nabity, P.D.; Fernández, R.; Arora, A.K.; Baa-Puyoulet, P.; Banfill, C.R.; Bao, L.; Barberà, M.; Bouallegue, M. The genome sequence of the grape phylloxera provides insights into the evolution, adaptation, and invasion routes of an iconic pest. BMC Biol. 2020, 18, 90. [Google Scholar]
- Arp, A.P.; Hunter, W.B.; Pelz-Stelinski, K.S. Annotation of the Asian citrus psyllid genome reveals a reduced innate immune system. Front. Physiol. 2016, 7, 570. [Google Scholar] [CrossRef] [PubMed]
- Hosmani, P.S.; Flores-Gonzalez, M.; Shippy, T.; Vosburg, C.; Massimino, C.; Tank, W.; Reynolds, M.; Tamayo, B.; Miller, S.; Norus, J. Chromosomal length reference assembly for Diaphorina citri using single-molecule sequencing and Hi-C proximity ligation with manually curated genes in developmental, structural and immune pathways. Biorxiv 2019, 12, 869685. [Google Scholar] [CrossRef]
- Nicholson, S.J.; Nickerson, M.L.; Dean, M.; Song, Y.; Hoyt, P.R.; Rhee, H.; Kim, C.; Puterka, G.J. The genome of Diuraphis noxia, a global aphid pest of small grains. BMC Genom. 2015, 16, 429. [Google Scholar] [CrossRef] [PubMed]
- Burger, N.F.V.; Botha, A.-M. Genome of Russian wheat aphid an economically important cereal aphid. Stand. Genom. Sci. 2017, 12, 90. [Google Scholar] [CrossRef]
- Sparks, M.E.; Bansal, R.; Benoit, J.B.; Blackburn, M.B.; Chao, H.; Chen, M.; Cheng, S.; Childers, C.; Dinh, H.; Doddapaneni, H.V. Brown marmorated stink bug, Halyomorpha halys (Stål), genome: Putative underpinnings of polyphagy, insecticide resistance potential and biology of a top worldwide pest. BMC Genom. 2020, 21, 227. [Google Scholar] [CrossRef]
- Ettinger, C.L.; Byrne, F.J.; Collin, M.A.; Carter-House, D.; Walling, L.L.; Atkinson, P.W.; Redak, R.A.; Stajich, J.E. Improved draft reference genome for the Glassy-winged Sharpshooter (Homalodisca vitripennis), a vector for Pierce’s disease. G3 Genes Genomes Genet. 2021, 11, jkab255. [Google Scholar] [CrossRef]
- Kohli, S.; Gulati, P.; Narang, A.; Maini, J.; Shamsudheen, K.; Pandey, R.; Scaria, V.; Sivasubbu, S.; Brahmachari, V. Genome and transcriptome analysis of the mealybug Maconellicoccus hirsutus: Correlation with its unique phenotypes. Genomics 2021, 113, 2483–2494. [Google Scholar] [CrossRef]
- Singh, K.S.; Cordeiro, E.M.; Troczka, B.J.; Pym, A.; Mackisack, J.; Mathers, T.C.; Duarte, A.; Legeai, F.; Robin, S.; Bielza, P. Global patterns in genomic diversity underpinning the evolution of insecticide resistance in the aphid crop pest Myzus persicae. Commun. Biol. 2021, 4, 847. [Google Scholar] [CrossRef]
- Ma, L.; Cao, L.J.; Gong, Y.J.; Hoffmann, A.A.; Zeng, A.P.; Wei, S.J.; Zhou, Z.S. Development of novel microsatellites for population genetic analysis of Phenacoccus solenopsis Tinsley (Hemipeta: Pseudoccoccidae) based on genomic analysis. Int. J. Biol. Macromol. 2019, 121, 1135–1144. [Google Scholar] [CrossRef]
- Villarroel, C.A.; González-González, A.; Alvarez-Baca, J.K.; Villarreal, P.; Ballesteros, G.I.; Figueroa, C.C.; Cubillos, F.A.; Ramírez, C.C. Genome sequencing of a predominant clonal lineage of the grain aphid Sitobion avenae. Insect Biochem. Mol. Biol. 2022, 143, 103742. [Google Scholar] [CrossRef]
- Byrne, S.; Schughart, M.; Carolan, J.C.; Gaffney, M.; Thorpe, P.; Malloch, G.; Wilkinson, T.; McNamara, L. Genome sequence of the English grain aphid, Sitobion avenae and its endosymbiont Buchnera aphidicola. G3 Genes Genomes Genet. 2022, 12, jkab418. [Google Scholar] [CrossRef]
- Xie, W.; He, C.; Fei, Z.; Zhang, Y. Chromosome-level genome assembly of the greenhouse whitefly (Trialeurodes vaporariorum Westwood). Mol. Ecol. Resour. 2020, 20, 995–1006. [Google Scholar] [CrossRef] [PubMed]
- Cridland, J.M.; Tsutsui, N.D.; Ramírez, S.R. The complex demographic history and evolutionary origin of the western honey bee, Apis mellifera. Genome Biol. Evol. 2017, 9, 457–472. [Google Scholar] [CrossRef]
- Consortium, H.G.S. Insights into social insects from the genome of the honeybee Apis mellifera. Nature 2006, 443, 931. [Google Scholar]
- Schrader, L.; Kim, J.W.; Ence, D.; Zimin, A.; Klein, A.; Wyschetzki, K.; Weichselgartner, T.; Kemena, C.; Stökl, J.; Schultner, E. Transposable element islands facilitate adaptation to novel environments in an invasive species. Nat. Commun. 2014, 5, 5495. [Google Scholar] [CrossRef]
- Robertson, H.M.; Waterhouse, R.M.; Walden, K.K.; Ruzzante, L.; Reijnders, M.J.; Coates, B.S.; Legeai, F.; Gress, J.C.; Biyiklioglu, S.; Weaver, D.K. Genome sequence of the wheat stem sawfly, Cephus cinctus, representing an early-branching lineage of the Hymenoptera, illuminates evolution of hymenopteran chemoreceptors. Genome Biol. Evol. 2018, 10, 2997–3011. [Google Scholar] [CrossRef] [PubMed]
- Smith, C.D.; Zimin, A.; Holt, C.; Abouheif, E.; Benton, R.; Cash, E.; Croset, V.; Currie, C.R.; Elhaik, E.; Elsik, C.G. Draft genome of the globally widespread and invasive Argentine ant (Linepithema humile). Proc. Natl. Acad. Sci. USA 2011, 108, 5673–5678. [Google Scholar] [CrossRef]
- Rubin, B.E.; Moreau, C.S. Comparative genomics reveals convergent rates of evolution in ant–plant mutualisms. Nat. Commun. 2016, 7, 12679. [Google Scholar] [CrossRef]
- Wurm, Y.; Wang, J.; Riba-Grognuz, O.; Corona, M.; Nygaard, S.; Hunt, B.G.; Ingram, K.K.; Falquet, L.; Nipitwattanaphon, M.; Gotzek, D. The genome of the fire ant Solenopsis invicta. Proc. Natl. Acad. Sci. USA 2011, 108, 5679–5684. [Google Scholar] [CrossRef]
- Harrop, T.W.; Guhlin, J.; McLaughlin, G.M.; Permina, E.; Stockwell, P.; Gilligan, J.; Le Lec, M.F.; Gruber, M.A.; Quinn, O.; Lovegrove, M. High-quality assemblies for three invasive social wasps from the Vespula genus. G3 Genes Genomes Genet. 2020, 10, 3479–3488. [Google Scholar] [CrossRef] [PubMed]
- Calla, B.; Demkovich, M.; Siegel, J.P.; Viana, J.P.G.; Walden, K.K.; Robertson, H.M.; Berenbaum, M.R. Selective sweeps in a nutshell: The genomic footprint of rapid insecticide resistance evolution in the almond agroecosystem. Genome Biol. Evol. 2021, 13, 234. [Google Scholar] [CrossRef] [PubMed]
- Cao, L.J.; Song, W.; Yue, L.; Guo, S.K.; Chen, J.C.; Gong, Y.J.; Hoffmann, A.A.; Wei, S.J. Chromosome-level genome of the peach fruit moth Carposina sasakii (Lepidoptera: Carposinidae) provides a resource for evolutionary studies on moths. Mol. Ecol. Resour. 2021, 21, 834–848. [Google Scholar] [CrossRef] [PubMed]
- Thrimawithana, A.H.; Wu, C.; Christeller, J.T.; Simpson, R.M.; Hilario, E.; Tooman, L.K.; Begum, D.; Jordan, M.D.; Crowhurst, R.; Newcomb, R.D. The genomics and population genomics of the light brown apple moth, Epiphyas postvittana, an invasive tortricid pest of horticulture. Insects 2022, 13, 264. [Google Scholar] [CrossRef]
- Cao, L.-J.; Song, W.; Chen, J.-C.; Fan, X.-L.; Hoffmann, A.A.; Wei, S.-J. Population genomic signatures of the oriental fruit moth related to the Pleistocene climates. Commun. Biol. 2022, 5, 142. [Google Scholar] [CrossRef]
- Wu, N.; Zhang, S.; Li, X.; Cao, Y.; Liu, X.; Wang, Q.; Liu, Q.; Liu, H.; Hu, X.; Zhou, X.J. Fall webworm genomes yield insights into rapid adaptation of invasive species. Nat. Ecol. Evol. 2019, 3, 105–115. [Google Scholar] [CrossRef]
- Chen, Q.; Zhao, H.; Wen, M.; Li, J.; Zhou, H.; Wang, J.; Zhou, Y.; Liu, Y.; Du, L.; Kang, H. Genome of the webworm Hyphantria cunea unveils genetic adaptations supporting its rapid invasion and spread. BMC Genom. 2020, 21, 242. [Google Scholar] [CrossRef]
- Sparks, M.E.; Hebert, F.O.; Johnston, J.S.; Hamelin, R.C.; Cusson, M.; Levesque, R.C.; Gundersen-Rindal, D.E. Sequencing, assembly and annotation of the whole-insect genome of Lymantria dispar dispar, the European gypsy moth. G3 Genes Genomes Genet. 2021, 11, jkab150. [Google Scholar] [CrossRef]
- Derks, M.F.; Smit, S.; Salis, L.; Schijlen, E.; Bossers, A.; Mateman, C.; Pijl, A.S.; de Ridder, D.; Groenen, M.A.; Visser, M.E. The genome of winter moth (Operophtera brumata) provides a genomic perspective on sexual dimorphism and phenology. Genome Biol. Evol. 2015, 7, 2321–2332. [Google Scholar] [CrossRef]
- Ryan, S.F.; Lombaert, E.; Espeset, A.; Vila, R.; Talavera, G.; Dincă, V.; Doellman, M.M.; Renshaw, M.A.; Eng, M.W.; Hornett, E.A. Global invasion history of the agricultural pest butterfly Pieris rapae revealed with genomics and citizen science. Proc. Natl. Acad. Sci. USA 2019, 116, 20015–20024. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; Cong, Q.; Kinch, L.N.; Borek, D.; Otwinowski, Z.; Grishin, N.V. Complete genome of Pieris rapae, a resilient alien, a cabbage pest, and a source of anti-cancer proteins. F1000Research 2016, 5, 2631. [Google Scholar] [CrossRef] [PubMed]
- You, M.; Ke, F.; You, S.; Wu, Z.; Liu, Q.; He, W.; Baxter, S.W.; Yuchi, Z.; Vasseur, L.; Gurr, G.M. Variation among 532 genomes unveils the origin and evolutionary history of a global insect herbivore. Nat. Commun. 2020, 11, 2321. [Google Scholar] [CrossRef] [PubMed]
- You, M.; Yue, Z.; He, W.; Yang, X.; Yang, G.; Xie, M.; Zhan, D.; Baxter, S.W.; Vasseur, L.; Gurr, G.M. A heterozygous moth genome provides insights into herbivory and detoxification. Nat. Genet. 2013, 45, 220–225. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Liu, Z.; Régnière, J.; Vasseur, L.; Lin, J.; Huang, S.; Ke, F.; Chen, S.; Li, J.; Huang, J. Large-scale genome-wide study reveals climate adaptive variability in a cosmopolitan pest. Nat. Commun. 2021, 12, 7206. [Google Scholar] [CrossRef] [PubMed]
- Gui, F.; Lan, T.; Zhao, Y.; Guo, W.; Dong, Y.; Fang, D.; Liu, H.; Li, H.; Wang, H.; Hao, R. Genomic and transcriptomic analysis unveils population evolution and development of pesticide resistance in fall armyworm Spodoptera frugiperda. Protein Cell 2022, 13, 513–531. [Google Scholar] [CrossRef] [PubMed]
- Gouin, A.; Bretaudeau, A.; Nam, K.; Gimenez, S.; Aury, J.-M.; Duvic, B.; Hilliou, F.; Durand, N.; Montagné, N.; Darboux, I. Two genomes of highly polyphagous lepidopteran pests (Spodoptera frugiperda, Noctuidae) with different host-plant ranges. Sci. Rep. 2017, 7, 11816. [Google Scholar] [CrossRef]
- Gimenez, S.; Abdelgaffar, H.; Goff, G.L.; Hilliou, F.; Blanco, C.A.; Hänniger, S.; Bretaudeau, A.; Legeai, F.; Nègre, N.; Jurat-Fuentes, J.L. Adaptation by copy number variation increases insecticide resistance in the fall armyworm. Commun. Biol. 2020, 3, 664. [Google Scholar] [CrossRef]
- Zhang, L.; Liu, B.; Zheng, W.; Liu, C.; Zhang, D.; Zhao, S.; Li, Z.; Xu, P.; Wilson, K.; Withers, A. Genetic structure and insecticide resistance characteristics of fall armyworm populations invading China. Mol. Ecol. Resour. 2020, 20, 1682–1696. [Google Scholar] [CrossRef] [PubMed]
- Brand, P.; Lin, W.; Johnson, B.R. The draft genome of the invasive walking stick, Medauroidea extradendata, reveals extensive lineage-specific gene family expansions of cell wall degrading enzymes in phasmatodea. G3 Genes Genomes Genet. 2018, 8, 1403–1408. [Google Scholar] [CrossRef] [PubMed]
- Rotenberg, D.; Baumann, A.A.; Ben-Mahmoud, S.; Christiaens, O.; Dermauw, W.; Ioannidis, P.; Jacobs, C.G.; Vargas Jentzsch, I.M.; Oliver, J.E.; Poelchau, M.F. Genome-enabled insights into the biology of thrips as crop pests. BMC Biol. 2020, 18, 142. [Google Scholar] [CrossRef]
- Talla, V.; Suh, A.; Kalsoom, F.; Dincă, V.; Vila, R.; Friberg, M.; Wiklund, C.; Backström, N. Rapid increase in genome size as a consequence of transposable element hyperactivity in wood-white (Leptidea) butterflies. Genome Biol. Evol. 2017, 9, 2491–2505. [Google Scholar] [CrossRef]
- Lavergne, S.; Muenke, N.J.; Molofsky, J. Genome size reduction can trigger rapid phenotypic evolution in invasive plants. Ann. Bot. 2010, 105, 109–116. [Google Scholar] [CrossRef] [PubMed]
- Specchia, V.; Janzen, S.; Marini, G.; Pinna, M. The potential link between mobile DNA and the invasiveness of the Species. J. RNAi Gene Silenc. 2017, 13. [Google Scholar]
- Singh, S.; Pandher, S.; Mishra, S.K.; Rathore, P. Weather Based Models for Forewarning of Pest Population in Cotton at Faridkot District of Punjab State. In Proceedings of the New-Dimensions in Agrometeorology for Sustainable Agriculture (NASA-2014), Pantnagar, India, 16–18 October 2014. [Google Scholar]
- Pitt, J. Modelling the Spread of Invasive Species across Heterogeneous Landscapes. Ph.D. Thesis, Lincoln University, Lincoln, New Zealand, 2008. [Google Scholar]
- Hlasny, V. Invasion of non-indigenous insects: The impact of quarantine laws. Int. J. Agric. Resour. Gov. Ecol. 2012, 9, 147. [Google Scholar] [CrossRef]
- Anila, P.; Enilda, S.; Odeta, L.; Etleva, H.; Bledar, P.; Erjola, K.; Eltjon, H. Geographical distribution of invasive insect species in albania. Sci. Ann. Danub. Delta Inst. 2012, 18, 211–218. [Google Scholar]
- Wan, X.W.; Liu, Y.H.; Luo, L.M.; Feng, C.H.; Ma, L. Inference on the invasion history of invasive alien species based on molecular methods. Acta Ecol. Sin. 2015, 35, 1296–1309. [Google Scholar]
- Guo, S.K.; Cao, L.J.; Song, W.; Shi, P.; Gao, Y.F.; Gong, Y.J.; Chen, J.C.; Hoffmann, A.A.; Wei, S.J. Chromosome-level assembly of the melon thrips genome yields insights into evolution of a sap-sucking lifestyle and pesticide resistance. Mol. Ecol. Resour. 2020, 20, 1110–1125. [Google Scholar] [CrossRef]
- Qiao, H.; Keesey, I.W.; Hansson, B.S.; Knaden, M. Gut microbiota affects development and olfactory behavior in Drosophila melanogaster. J. Exp. Biol. 2019, 222, 192500. [Google Scholar] [CrossRef]
- Zheng, H.; Powell, J.E.; Steele, M.I.; Dietrich, C.; Moran, N.A. Honeybee gut microbiota promotes host weight gain via bacterial metabolism and hormonal signaling. Proc. Natl. Acad. Sci. USA 2017, 114, 4775–4780. [Google Scholar] [CrossRef] [PubMed]
- Husnik, F.; McCutcheon, J.P. Repeated replacement of an intrabacterial symbiont in the tripartite nested mealybug symbiosis. Proc. Natl. Acad. Sci. USA 2016, 113, E5416–E5424. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Order | Scientific Name | Level | Total Length (Mb) | Gene Number | Release Date | References |
|---|---|---|---|---|---|---|
| Blattaria | Blattella germanica | Scaffold | 2037.3 | 29,581 | 2018 | [19] |
| Cryptotermes secundus | Scaffold | 1018.93 | 29,285 | 2018 | [19] | |
| Periplaneta americana | Scaffold | 3374.84 | NA | 2018 | [20] | |
| Zootermopsis nevadensis | Scaffold | 485.025 | 30,187 | 2014 | ||
| Coleoptera | Aethina tumida | Chromosome | 259.984 | 21,401 | 2022 | [21,22,23,24] |
| Agrilus planipennis | Scaffold | 353.073 | 22,159 | 2014 | [25] | |
| Anoplophora glabripennis | Scaffold | 706.969 | 20,632 | 2013 | [26,27] | |
| Dendroctonus valens | Scaffold | 322.407 | NA | 2022 | [28] | |
| Diabrotica virgifera | Scaffold | 2418.07 | 28,061 | 2018 | [29] | |
| Harmonia axyridis | Chromosome | 425.525 | 23,861 | 2021 | [30] | |
| Hypothenemus hampei | Scaffold | 162.571 | NA | 2020 | [31] | |
| Leptinotarsa decemlineata | Scaffold | 641.993 | 19,038 | 2013 | [16] | |
| Oryctes rhinoceros | Scaffold | 377.356 | NA | 2021 | [24] | |
| Rhynchophorus ferrugineus | Scaffold | 782.098 | NA | 2020 | ||
| Diptera | Aedes aegypti | Chromosome | 1278.73 | 28,317 | 2017 | [32] |
| Aedes albopictus | Scaffold | 2538.39 | 40,086 | 2019 | [33,34,35] | |
| Anopheles stephensi | Chromosome | 243.46 | 29,660 | 2020 | [36] | |
| Bactrocera cucurbitae | Chromosome | 244.9 | 13,286 | NA | [37] | |
| Bactrocera dorsalis | Chromosome | 530.327 | 30,102 | 2022 | [38] | |
| Bactrocera oleae | Scaffold | 484.947 | 33,727 | 2015 | [39] | |
| Bactrocera tryoni | Chromosome | 570.659 | 23,950 | 2021 | [40] | |
| Ceratitis capitata | Scaffold | 436.491 | 22,949 | 2013 | [41] | |
| Culex quinquefasciatus | Chromosome | 573.23 | 24,531 | 2020 | ||
| Drosophila suzukii | Chromosome | 268.012 | 25,540 | 2020 | [42,43] | |
| Liriomyza trifolii | Scaffold | 69.6986 | NA | 2015 | [44] | |
| Mayetiola destructor | Scaffold | 185.828 | NA | 2010 | ||
| Philornis downsi | Scaffold | 954.765 | NA | 2021 | [45] | |
| Zaprionus indianus | Scaffold | 197.261 | NA | 2021 | [13] | |
| Hemiptera | Acyrthosiphon pisum | Chromosome | 533.649 | 27,913 | 2019 | [46] |
| Aphis glycines | Scaffold | 308.064 | 18,358 | 2019 | [47,48] | |
| Bemisia tabaci | Scaffold | 615.017 | 22,737 | 2016 | [23,49,50] | |
| Cimex lectularius | Scaffold | 510.849 | 24,194 | 2014 | [51] | |
| Daktulosphaira vitifoliae | Scaffold | 282.6 | NA | 2022 | [52] | |
| Diaphorina citri | Scaffold | 485.705 | 24,730 | 2013 | [53,54] | |
| Diuraphis noxia | Scaffold | 395.074 | 17,476 | 2015 | [55,56] | |
| Euschistus heros | Contig | 1325.16 | NA | 2018 | ||
| Halyomorpha halys | Scaffold | 998.241 | 25,026 | 2014 | [57] | |
| Homalodisca vitripennis | Chromosome | 2305 | 31,163 | 2021 | [58] | |
| Lycorma delicatula | Scaffold | 2252 | 10,652 | NA | ||
| Maconellicoccus hirsutus | Contig | 189.239 | NA | 2018 | [59] | |
| Myzus persicae | Scaffold | 347.313 | 23,910 | 2016 | [60] | |
| Paracoccus marginatus | Scaffold | 191.208 | NA | 2016 | ||
| Phenacoccus solenopsis | Chromosome | 292.538 | NA | 2019 | [61] | |
| Piezodorus guildinii | Chromosome | 1205.42 | NA | 2022 | ||
| Sitobion avenae | Scaffold | 393.019 | NA | 2021 | [62,63] | |
| Trialeurodes vaporariorum | Scaffold | 787.484 | NA | 2020 | [64] | |
| Hymenoptera | Apis mellifera | Chromosome | 225.251 | 23,471 | 2018 | [65,66] |
| Cardiocondyla obscurior | Scaffold | 193.051 | NA | 2021 | [67] | |
| Cephus cinctus | Scaffold | 160.751 | 31,793 | 2014 | [68] | |
| Linepithema humile | Scaffold | 219.501 | 21,674 | 2011 | [69] | |
| Polistes dominula | Scaffold | 208.026 | 20,855 | 2015 | ||
| Pseudomyrmex gracilis | Scaffold | 282.776 | 23,473 | 2017 | [70] | |
| Solenopsis invicta | Chromosome | 378.102 | 30,910 | 2021 | [71] | |
| Vespula germanica | Chromosome | 205.789 | NA | 2021 | [72] | |
| Vespula pensylvanica | Chromosome | 179.37 | 23,842 | 2020 | [72] | |
| Vespula vulgaris | Chromosome | 188.205 | NA | 2021 | [72] | |
| Lepidoptera | Amyelois transitella | Scaffold | 406.468 | 18,472 | 2015 | [73] |
| Carposina sasakii | Chromosome | 399.04 | NA | 2020 | [74] | |
| Cydia pomonella | Scaffold | 772.892 | NA | 2018 | [17] | |
| Diatraea saccharalis | Scaffold | 453.235 | NA | 2020 | ||
| Epiphyas postvittana | Scaffold | 598.1 | 31,389 | NA | [75] | |
| Grapholita molesta | Chromosome | 517.71 | NA | 2022 | [76] | |
| Helicoverpa armigera | Chromosome | 356.686 | 28,010 | 2022 | ||
| Helicoverpa zea | Chromosome | 375.181 | 23,696 | 2022 | ||
| Hyphantria cunea | Scaffold | 510.512 | NA | 2018 | [77,78] | |
| Lymantria dispar dispar | Scaffold | 864.963 | NA | 2019 | [79] | |
| Operophtera brumata | Chromosome | 619.179 | NA | 2022 | [80] | |
| Pieris rapae | Chromosome | 256.37 | 20,497 | 2021 | [81,82] | |
| Plutella xylostella | Chromosome | 323.303 | 22,256 | 2022 | [83,84,85] | |
| Spodoptera frugiperda | Chromosome | 485.97 | 30,806 | 2020 | [15,86,87,88,89] | |
| Phasmida | Medauroidea extradentata | Scaffold | 2593.36 | NA | 2018 | [90] |
| Thysanoptera |
Frankliniella occidentalis
Thrips palmi | Scaffold Scaffold | 274.99 237.845 | 23,356 27,217 | 2014 2020 | [91] [37] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, Z.; Chen, Y.; Chen, Y.; Lu, Z.; Gui, F. Tracking Adaptive Pathways of Invasive Insects: Novel Insight from Genomics. Int. J. Mol. Sci. 2023, 24, 8004. https://doi.org/10.3390/ijms24098004
Sun Z, Chen Y, Chen Y, Lu Z, Gui F. Tracking Adaptive Pathways of Invasive Insects: Novel Insight from Genomics. International Journal of Molecular Sciences. 2023; 24(9):8004. https://doi.org/10.3390/ijms24098004
Chicago/Turabian StyleSun, Zhongxiang, Yao Chen, Yaping Chen, Zhihui Lu, and Furong Gui. 2023. "Tracking Adaptive Pathways of Invasive Insects: Novel Insight from Genomics" International Journal of Molecular Sciences 24, no. 9: 8004. https://doi.org/10.3390/ijms24098004