
Domain Localization by Graphene Oxide in Supported Lipid Bilayers

Abstract
:1. Introduction
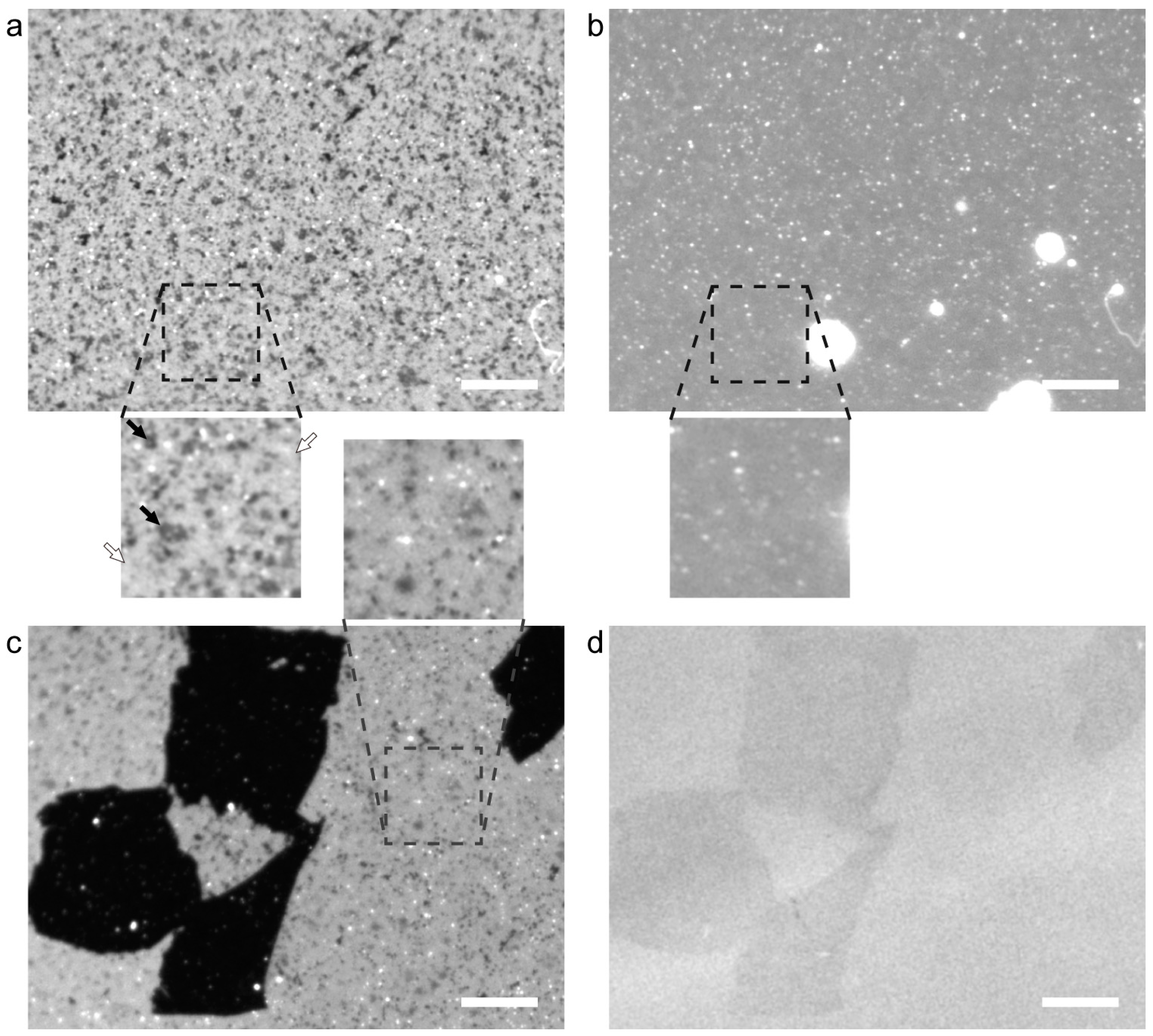
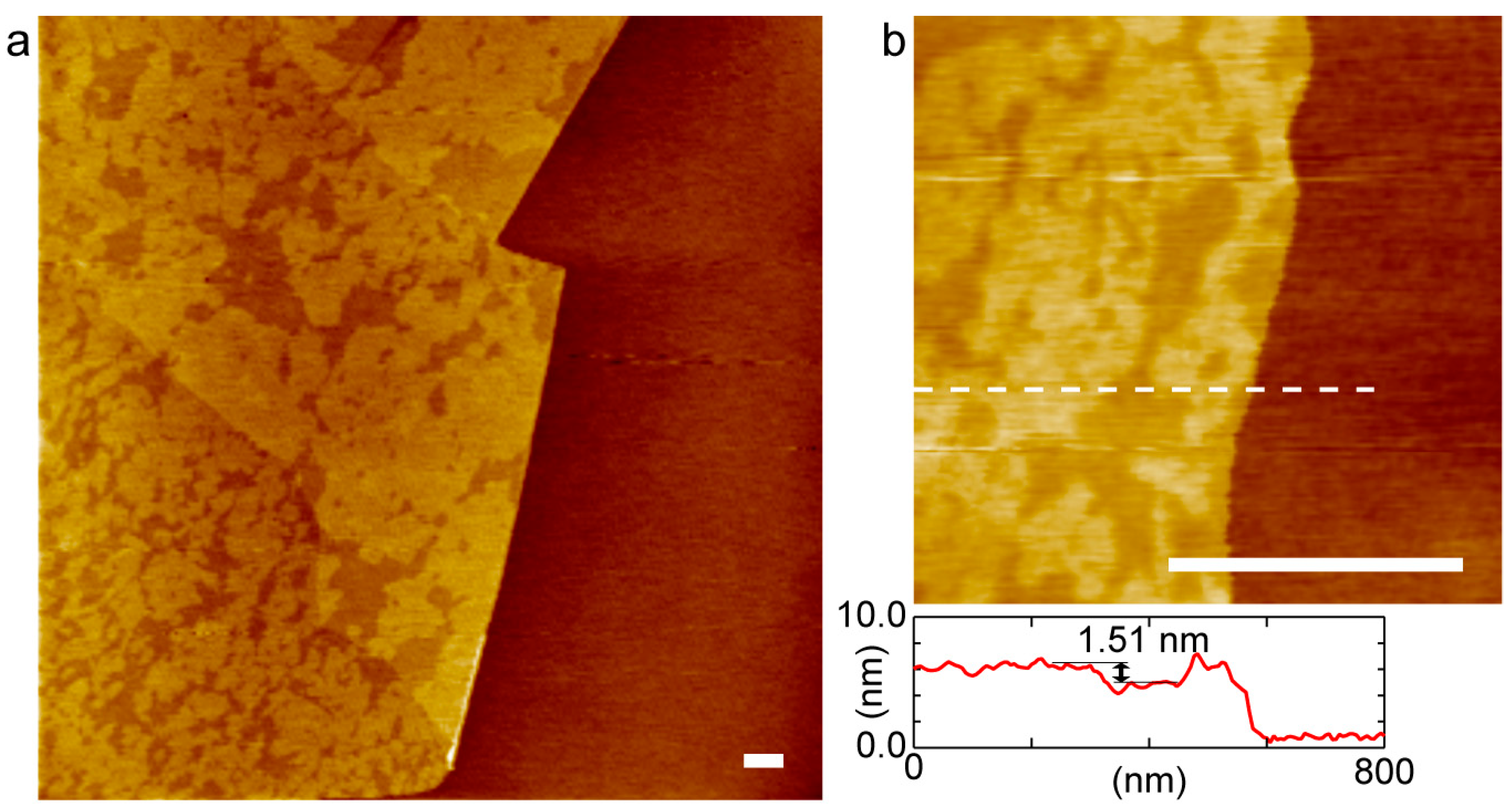
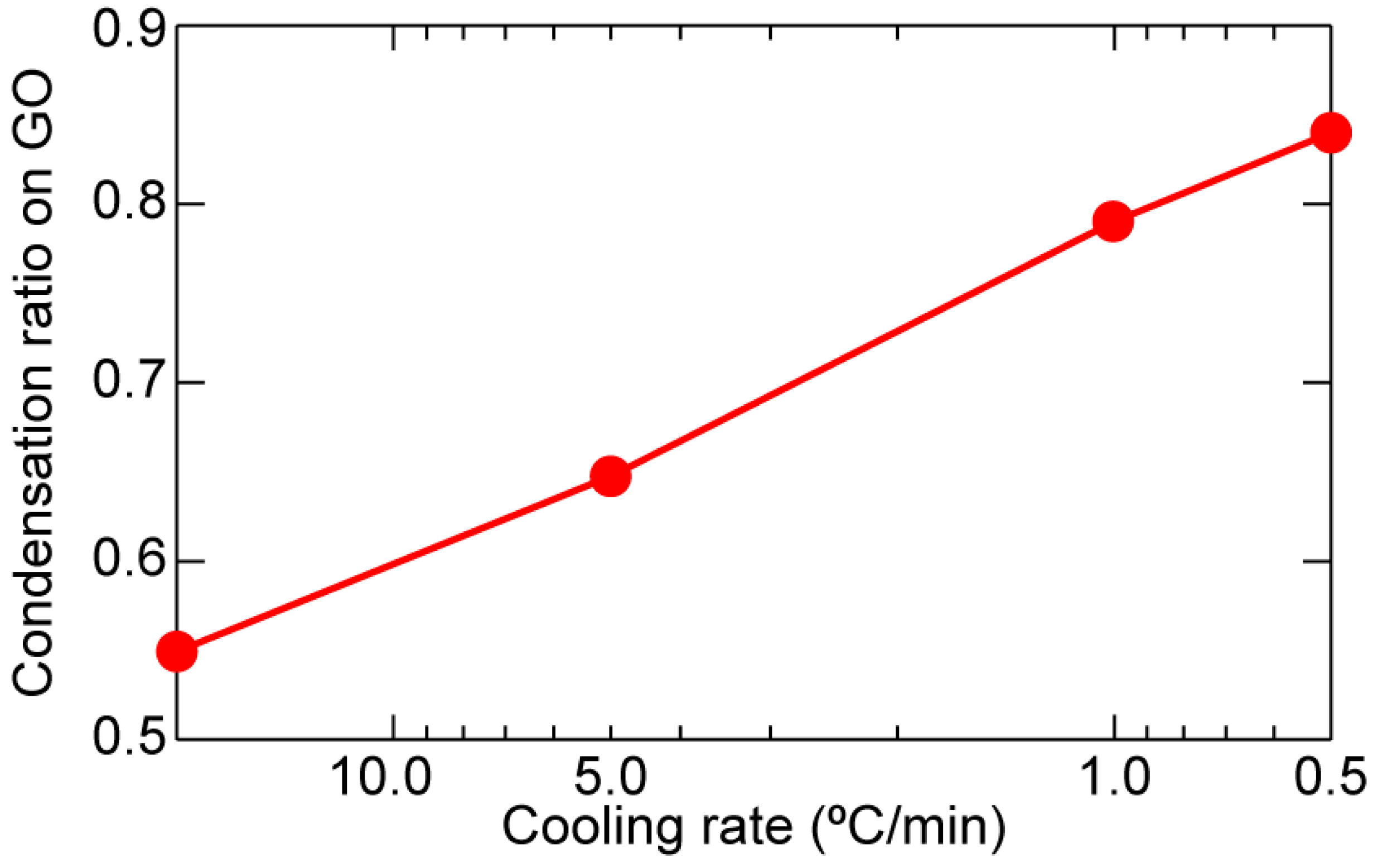
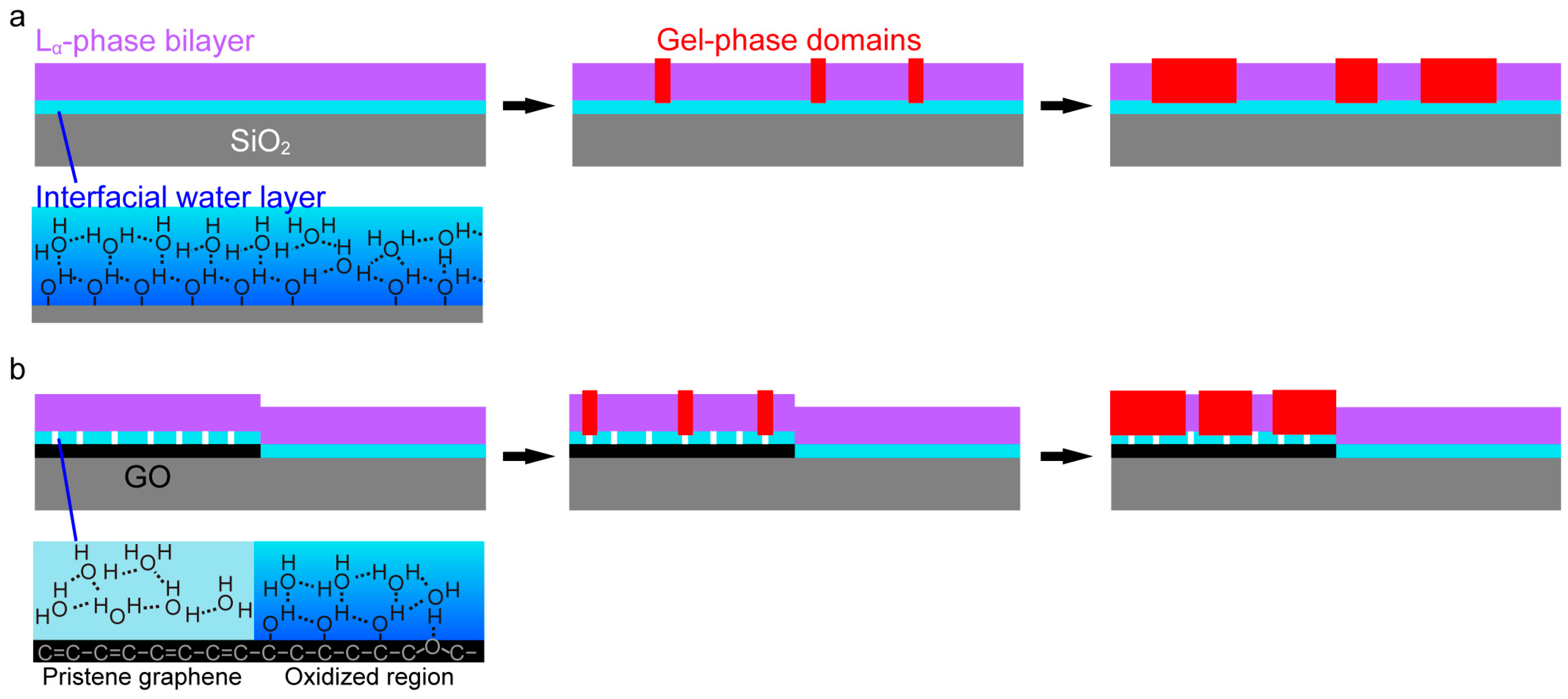
2. Results and Discussion
3. Materials and Methods
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Binder, W.H.; Barragan, V.; Menger, F.M. Domains and rafts in lipid membranes. Angew. Chem. Int. Ed. Engl. 2003, 42, 5802–5827. [Google Scholar] [CrossRef]
- van Meer, G.; Voelker, D.R.; Feigenson, G.W. Membrane lipids: Where they are and how they behave. Nat. Rev. Mol. Cell Biol. 2008, 9, 112–124. [Google Scholar] [CrossRef]
- Lingwood, D.; Simons, K. Lipid rafts as a membrane-organizing principle. Science 2010, 327, 46–50. [Google Scholar] [CrossRef] [Green Version]
- Spira, F.; Mueller, N.S.; Beck, G.; von Olshausen, P.; Beig, J.; Wedlich-Söldner, R. Patchwork organization of the yeast plasma membrane into numerous coexisting domains. Nat. Cell Biol. 2012, 14, 640–648. [Google Scholar] [CrossRef]
- Castellana, E.T.; Cremer, P.S. Solid supported lipid bilayers: From biophysical studies to sensor design. Surf. Sci. Rep. 2006, 61, 429–444. [Google Scholar] [CrossRef]
- Tero, R. Substrate Effects on the Formation Process, Structure and Physicochemical Properties of Supported Lipid Bilayers. Materials 2012, 5, 2658–2680. [Google Scholar] [CrossRef] [Green Version]
- Fromherz, P.; Kiessling, V.; Kottig, K.; Zeck, G. Membrane transistor with giant lipid vesicle touching a silicon chip. Appl. Phys. A Mater. Sci. Process. 1999, 69, 571–576. [Google Scholar] [CrossRef]
- Ajo-Franklin, C.M.; Yoshina-Ishii, C.; Boxer, S.G. Probing the structure of supported membranes and tethered oligonucleotides by fluorescence interference contrast microscopy. Langmuir 2005, 21, 4976–4983. [Google Scholar] [CrossRef]
- Yang, J.; Appleyard, J. The Main Phase Transition of Mica-Supported Phosphatidylcholine Membranes. J. Phys. Chem. B 2000, 104, 8097–8100. [Google Scholar] [CrossRef]
- Keller, D.; Larsen, N.; Møller, I.; Mouritsen, O. Decoupled Phase Transitions and Grain-Boundary Melting in Supported Phospholipid Bilayers. Phys. Rev. Lett. 2005, 94, 025701. [Google Scholar] [CrossRef]
- Lin, W.C.; Blanchette, C.D.; Ratto, T.V.; Longo, M.L. Lipid asymmetry in DLPC/DSPC-supported lipid bilayers: A combined AFM and fluorescence microscopy study. Biophys. J. 2006, 90, 228–237. [Google Scholar] [CrossRef] [Green Version]
- Motegi, T.; Yamazaki, K.; Ogino, T.; Tero, R. Substrate-Induced Structure and Molecular Dynamics in a Lipid Bilayer Membrane. Langmuir 2017, 33, 14748–14755. [Google Scholar] [CrossRef]
- Groves, J.T.; Boxer, S.G. Micropattern Formation in Supported Lipid Membranes. Acc. Chem. Res. 2002, 35, 149–157. [Google Scholar] [CrossRef]
- Jackson, B.L.; Groves, J.T. Hybrid Protein−Lipid Patterns from Aluminum Templates. Langmuir 2007, 23, 2052–2057. [Google Scholar] [CrossRef]
- Tanimoto, Y.; Okada, K.; Hayashi, F.; Morigaki, K. Evaluating the Raftophilicity of Rhodopsin Photoreceptor in a Patterned Model Membrane. Biophys. J. 2015, 109, 2307–2316. [Google Scholar] [CrossRef] [Green Version]
- Yoon, T.-Y.; Jeong, C.; Lee, S.-W.; Kim, J.H.; Choi, M.C.; Kim, S.-J.; Kim, M.W.; Lee, S.-D. Topographic control of lipid-raft reconstitution in model membranes. Nat. Mater. 2006, 5, 281–285. [Google Scholar] [CrossRef]
- Parthasarathy, R.; Yu, C.; Groves, J.T. Curvature-modulated phase separation in lipid bilayer membranes. Langmuir 2006, 22, 5095–5099. [Google Scholar] [CrossRef]
- Kaizuka, Y.; Groves, J.T. Bending-mediated superstructural organizations in phase-separated lipid membranes. New J. Phys. 2010, 12, 095001. [Google Scholar] [CrossRef]
- Sumitomo, K.; Oshima, A. Liquid-Ordered/Liquid-Crystalline Phase Separation at a Lipid Bilayer Suspended over Microwells. Langmuir 2017, 33, 13277–13283. [Google Scholar] [CrossRef]
- Belling, J.N.; Cheung, K.M.; Jackman, J.A.; Sut, T.N.; Allen, M.; Park, J.H.; Jonas, S.J.; Cho, N.J.; Weiss, P.S. Lipid Bicelle Micropatterning Using Chemical Lift-Off Lithography. ACS Appl. Mater. Interfaces 2020, 12, 13447–13455. [Google Scholar] [CrossRef]
- Rossetti, F.F.; Bally, M.; Michel, R.; Textor, M.; Reviakine, I. Interactions between titanium dioxide and phosphatidyl serine-containing liposomes: Formation and patterning of supported phospholipid bilayers on the surface of a medically relevant material. Langmuir 2005, 21, 6443–6450. [Google Scholar] [CrossRef]
- Tero, R.; Watanabe, H.; Urisu, T. Supported phospholipid bilayer formation on hydrophilicity-controlled silicon dioxide surfaces. Phys. Chem. Chem. Phys. 2006, 8, 3885–3894. [Google Scholar] [CrossRef]
- Sibold, J.; Tewaag, V.E.; Vagedes, T.; Mey, I.; Steinem, C. Phase separation in pore-spanning membranes induced by differences in surface adhesion. Phys. Chem. Chem. Phys. 2020, 22, 9308–9315. [Google Scholar] [CrossRef] [Green Version]
- Gao, W.; Alemany, L.B.; Ci, L.; Ajayan, P.M. New insights into the structure and reduction of graphite oxide. Nat. Chem. 2009, 1, 403–408. [Google Scholar] [CrossRef]
- Kim, J.; Kim, F.; Huang, J. Seeing graphene-based sheets. Mater. Today 2010, 13, 28–38. [Google Scholar] [CrossRef]
- Chen, D.; Feng, H.; Li, J. Graphene Oxide: Preparation, Functionalization, and Electrochemical Applications. Chem. Rev. 2012, 112, 6027–6053. [Google Scholar] [CrossRef]
- Erickson, K.; Erni, R.; Lee, Z.; Alem, N.; Gannett, W.; Zettl, A. Determination of the Local Chemical Structure of Graphene Oxide and Reduced Graphene Oxide. Adv. Mater. 2010, 22, 4467–4472. [Google Scholar] [CrossRef]
- Cote, L.J.; Kim, J.; Zhang, Z.; Sun, C.; Huang, J. Tunable assembly of graphene oxide surfactant sheets: Wrinkles, overlaps and impacts on thin film properties. Soft Matter 2010, 6, 6096. [Google Scholar] [CrossRef]
- Okamoto, Y.; Tsuzuki, K.; Iwasa, S.; Ishikawa, R.; Sandhu, A.; Tero, R. Fabrication of Supported Lipid Bilayer on Graphene Oxide. J. Phys. Conf. Ser. 2012, 352, 012017. [Google Scholar] [CrossRef]
- Okamoto, Y.; Motegi, T.; Iwasa, S.; Sandhu, A.; Tero, R. Fluidity evaluation of cell membrane model formed on graphene oxide with single particle tracking using quantum dot. Jpn. J. Appl. Phys. 2015, 54, 04DL09. [Google Scholar] [CrossRef]
- Okamoto, Y.; Iwasa, S.; Tero, R. Quenching Efficiency of Quantum Dots Conjugated to Lipid Bilayers on Graphene Oxide Evaluated by Fluorescence Single Particle Tracking. Appl. Sci. 2022, 12, 3733. [Google Scholar] [CrossRef]
- Furuya, K.; Mitsui, T. Phase Transitions in Bilayer Membranes of Dioleoyl-Phosphatidylcholine/Dipalmitoyl-Phosphatidylcholine. J. Phys. Soc. Japan 1979, 46, 611–616. [Google Scholar] [CrossRef]
- Marsh, D. Handbook of Lipid Bilayers, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2013; ISBN 9781420088328. [Google Scholar]
- Maekawa, T.; Chin, H.; Nyu, T.; Sut, T.N.; Ferhan, A.R.; Hayashi, T.; Cho, N.J. Molecular diffusion and nano-mechanical properties of multi-phase supported lipid bilayers. Phys. Chem. Chem. Phys. 2019, 21, 16686–16693. [Google Scholar] [CrossRef] [PubMed]
- Himeno, H.; Shimokawa, N.; Komura, S.; Andelman, D.; Hamada, T.; Takagi, M. Charge-induced phase separation in lipid membranes. Soft Matter 2014, 10, 7959–7967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ujihara, T.; Suzuki, S.; Yamauchi, Y.; Tero, R.; Takeda, Y. Local concentration of gel phase domains in supported lipid bilayers under light irradiation in binary mixture of phospholipids doped with dyes for photoinduced activation. Langmuir 2008, 24, 10974–10980. [Google Scholar] [CrossRef]
- Goh, M.W.S.; Tero, R. Non-raft submicron domain formation in cholesterol-containing lipid bilayers induced by polyunsaturated phosphatidylethanolamine. Colloids Surf. B Biointerfaces 2022, 210, 112235. [Google Scholar] [CrossRef]
- Tero, R.; Ujihara, T.; Urisu, T. Lipid bilayer membrane with atomic step structure: Supported bilayer on a step-and-terrace TiO2(100) surface. Langmuir 2008, 24, 11567–11576. [Google Scholar] [CrossRef]
- Tero, R.; Lau, J.M.Y.; Kanomata, K.; Hirose, F. Controlling fluorescence quenching efficiency by graphene oxide in supported lipid bilayers using SiO2 layer fabricated by atomic layer deposition. Jpn. J. Appl. Phys. 2023, 62, SC1041. [Google Scholar] [CrossRef]
- Nagashio, K.; Nishimura, T.; Kita, K.; Toriumi, A. Mobility Variations in Mono- and Multi-Layer Graphene Films. Appl. Phys. Express 2009, 2, 025003. [Google Scholar] [CrossRef] [Green Version]
- Attwood, S.J.; Choi, Y.; Leonenko, Z. Preparation of DOPC and DPPC supported planar lipid bilayers for atomic force microscopy and atomic force spectroscopy. Int. J. Mol. Sci. 2013, 14, 3514–3539. [Google Scholar] [CrossRef] [Green Version]
- Nagle, J.F.; Tristram-Nagle, S. Structure of lipid bilayers. Biochim. Biophys. Acta—Rev. Biomembr. 2000, 1469, 159–195. [Google Scholar] [CrossRef] [Green Version]
- Giocondi, M.C.; Vié, V.; Lesniewska, E.; Milhiet, P.E.; Zinke-Allmang, M.; Le Grimellec, C. Phase topology and growth of single domains in lipid bilayers. Langmuir 2001, 17, 1653–1659. [Google Scholar] [CrossRef]
- Blanchette, C.D.; Lin, W.C.; Orme, C.A.; Ratto, T.V.; Longo, M.L. Using nucleation rates to determine the interfacial line tension of symmetric and asymmetric lipid bilayer domains. Langmuir 2007, 23, 5875–5877. [Google Scholar] [CrossRef] [PubMed]
- Kakimoto, Y.; Tachihara, Y.; Okamoto, Y.; Miyazawa, K.; Fukuma, T.; Tero, R. Morphology and Physical Properties of Hydrophilic-Polymer-Modified Lipids in Supported Lipid Bilayers. Langmuir 2018, 34, 7201–7209. [Google Scholar] [CrossRef]
- Kaufmann, S.; Borisov, O.; Textor, M.; Reimhult, E. Mechanical properties of mushroom and brush poly(ethylene glycol)-phospholipid membranes. Soft Matter 2011, 7, 9267–9275. [Google Scholar] [CrossRef]
- Tsuzuki, K.; Okamoto, Y.; Iwasa, S.; Ishikawa, R.; Sandhu, A.; Tero, R. Reduced Graphene Oxide as the Support for Lipid Bilayer Membrane. J. Phys. Conf. Ser. 2012, 352, 012016. [Google Scholar] [CrossRef] [Green Version]
- Yamazaki, K.; Kunii, S.; Ogino, T. Characterization of Interfaces between Graphene Films and Support Substrates by Observation of Lipid Membrane Formation. J. Phys. Chem. C 2013, 117, 18913–18918. [Google Scholar] [CrossRef]
- Lee, J.S.; Kim, N.H.; Kang, M.S.; Yu, H.; Lee, D.R.; Oh, J.H.; Chang, S.T.; Cho, J.H. Wafer-Scale Patterning of Reduced Graphene Oxide Electrodes by Transfer-and-Reverse Stamping for High Performance OFETs. Small 2013, 9, 2817–2825. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, R.D.; Murastov, G.V.; Lipovka, A.; Fatkullin, M.I.; Nozdrina, O.; Pavlov, S.K.; Postnikov, P.S.; Chehimi, M.M.; Chen, J.J.; Sheremet, E. High-power laser-patterning graphene oxide: A new approach to making arbitrarily-shaped self-aligned electrodes. Carbon N. Y. 2019, 151, 148–155. [Google Scholar] [CrossRef]
- Shi, Y.; Osada, M.; Ebina, Y.; Sasaki, T. Single Droplet Assembly for Two-Dimensional Nanosheet Tiling. ACS Nano 2020, 14, 15216–15226. [Google Scholar] [CrossRef]
- Takami, T.; Ito, T.; Ogino, T. Self-Assembly of a Monolayer Graphene Oxide Film Based on Surface Modification of Substrates and its Vapor-Phase Reduction. J. Phys. Chem. C 2014, 118, 9009–9017. [Google Scholar] [CrossRef]
- Hummers, W.S., Jr.; Offeman, R.E. Preparation of graphitic oxide. J. Am. Chem. Soc. 1958, 80, 1339. [Google Scholar] [CrossRef]
- Tung, V.C.; Allen, M.J.; Yang, Y.; Kaner, R.B. High-throughput solution processing of large-scale graphene. Nat. Nanotechnol. 2009, 4, 25–29. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cooling Rate (°C/min) | 20.0 | 5.0 | 1.0 | 0.5 |
|---|---|---|---|---|
| GO/SiO2/Si (%) | 6.4 | 4.9 | 3.0 | 2.4 |
| SiO2/Si without GO (%) | 14.2 | 13.9 | 14.3 | 15.0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tero, R.; Hagiwara, Y.; Saito, S. Domain Localization by Graphene Oxide in Supported Lipid Bilayers. Int. J. Mol. Sci. 2023, 24, 7999. https://doi.org/10.3390/ijms24097999
Tero R, Hagiwara Y, Saito S. Domain Localization by Graphene Oxide in Supported Lipid Bilayers. International Journal of Molecular Sciences. 2023; 24(9):7999. https://doi.org/10.3390/ijms24097999
Chicago/Turabian StyleTero, Ryugo, Yoshi Hagiwara, and Shun Saito. 2023. "Domain Localization by Graphene Oxide in Supported Lipid Bilayers" International Journal of Molecular Sciences 24, no. 9: 7999. https://doi.org/10.3390/ijms24097999