1. Introduction
Pork is the most consumed meat worldwide and accounts for 37% of all meat, more than beef or chicken [
1]. In recent years, the demand for high-quality meat has been increasing dramatically. As a result, growing studies are dedicated to producing delicious meat with higher nutritional values in order to meet consumer demand. Skeletal muscle accounts for about 40−60% of the mammalian body weight (BW) [
2]. The skeletal muscle is a highly heterogeneous tissue type, which is closely related to economic traits, such as muscle growth and development and meat quality in livestock and poultry production [
2]. Pork quality can be affected by numerous factors, and the type and composition of muscle fiber is one of the crucial factors. Muscle-fiber characteristics are associated with numerous meat quality traits, including pH, drip loss, meat color, tenderness, and intramuscular fat content [
3]. In addition, the size, number, and type of muscle fibers are closely interrelated and implicated in muscle-fiber characteristics [
4]. The primary myofibers develop between 25 and 50 days of gestation and act as a template for forming secondary fibers that are completed between 80 to 90 days of gestation [
5]. Furthermore, muscle development undergoes significant changes during ontogenesis [
6]. Hence, improving the growth and development of skeletal muscle is a vital strategy to improve the productivity and quality of pork.
During gestation and lactation, the nutritional and health status of the sow has pivotal effects on their offspring’s pre- and post-natal growth and development. The placenta and milk are the determining factors that could affect intrauterine fetal development and postpartum neonatal growth and development. Moreover, the fetal and neonatal are the most vital periods for skeletal muscle development. Furthermore, the development of post-natal muscle fibers is determined by the composition and types of fetal and neonatal muscle fibers [
7]. Therefore, regulation of maternal nutrition and health plays a pivotal role in the skeletal muscle performance of offspring pigs.
Feed additives are always used to improve productivity and meat quality in swine production. For example, prebiotics, probiotics, and synbiotics have been studied extensively as feed additives in swine production, which have shown significant impacts on microbial community composition in the short and long term [
8]. In the past few years, numerous studies have been conducted on the beneficial effects of probiotics and prebiotics in animals. Shin et al. [
9] revealed that probiotics supplementation has beneficial effects on pigs, including improving growth performance, promoting nutrient digestion, absorption, and utilization, modulating intestinal microbiota, as well as ameliorating gut health complications. Additionally, a previous study illustrated that oral administration of β-glucan could improve the duodenal villi dimensions, splenic lymphoid diameter, muscular fiber diameter, and muscular glycogen areas in New Zealand white and APRI rabbits [
10]. Dietary supplementation of
Saccharomyces cerevisiae provided beneficial effects on growth performance and profitability in rabbits [
11]. Abd EI-Aziz et al. [
12] highlighted that fructo-oligosaccharide supplementation in drinking water enhances growth and carcass traits by improving the hematobiochemical parameters and antioxidant status and reducing cecal pathogenic bacteria in two different rabbit breeds. Moreover, our previous study indicated that sows and their offspring’s (sow-offspring) diets supplemented with probiotics or synbiotics could alter microbiota composition by increasing the abundance of beneficial bacteria (i.e.,
Bifidobacterium and
Lactobacillus), decreasing potentially harmful bacteria (i.e.,
E.coli), and enhancing the immune and antioxidant capacity in offspring pigs [
13]. In addition, these feed additives have also been found to be an effective strategy for improving pork quality by improving meat redness and tenderness, increasing meat protein, and decreasing meat drip loss [
14]. However, a few studies focused on whether sow-offspring diets supplemented with probiotics or synbiotics could improve the growth performance and meat quality in offspring pigs.
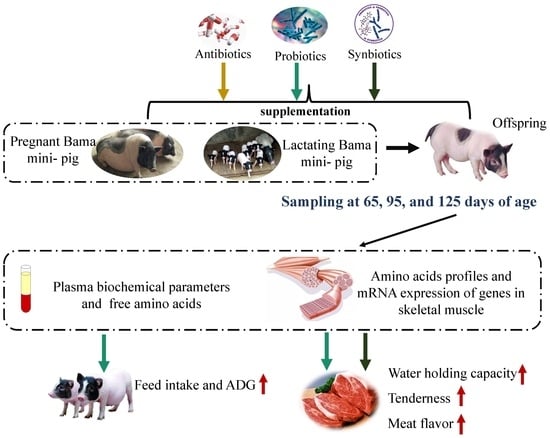
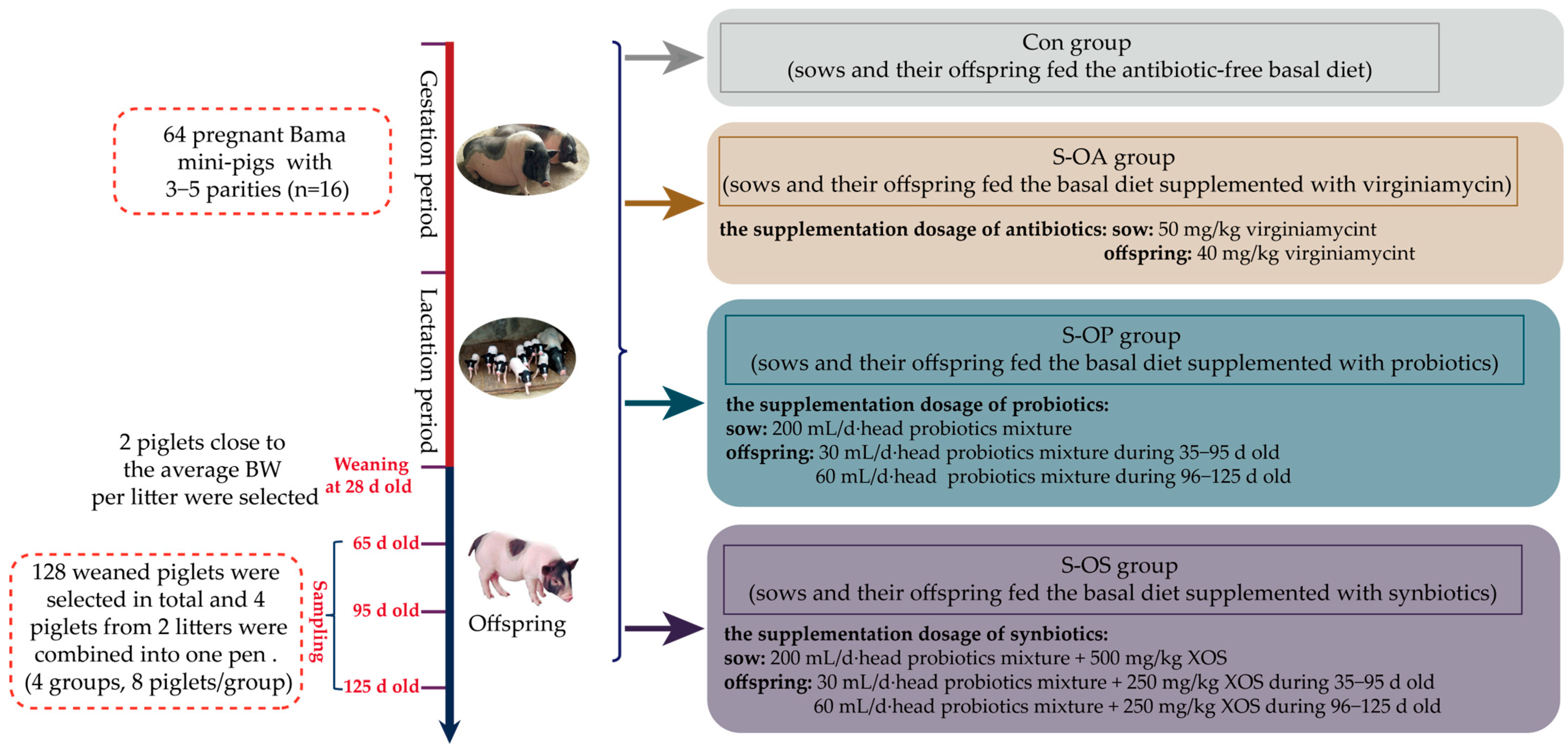
Thus, we hypothesized that sow-offspring probiotics and synbiotics supplementation could improve offspring pigs’ growth performance and meat quality. Therefore, the present study was conducted to explore the effects of dietary probiotics and synbiotics supplementation in sow-offspring diets on growth performance and meat quality in offspring pigs at different time points (65, 95, and 125 d old) after weaning. Thus, we examined related indicators and revealed the possible mechanism from the aspects of the metabolism of the body and gene expression related to the muscle-fiber type and muscle growth. Moreover, this study could also provide the theoretical basis for improving pork quality.
3. Discussion
Growing interest has been given to the positive effects of probiotics and synbiotics, including promoting growth and development, improving gut health, boosting the immune system, and preventing diseases in humans and animals. Our previous study demonstrated that maternal diets supplemented with probiotics and synbiotics have beneficial roles in improving piglet performance [
15]. However, it is poorly known whether sow-offspring diets supplemented with probiotics and synbiotics could have positive effects on the growth performance and meat quality of the offspring pigs. In the present study, our findings demonstrated that sow-offspring diets supplemented with probiotics and synbiotics have beneficial effects on meat quality, manifested in the improvement of sensory indexes (including tenderness, water-holding capacity, and meat flavor), alteration of plasma metabolites related to nitrogen metabolism, and gene expression associated with muscle-fiber types and muscle growth. In addition, probiotics supplementation in sow-offspring diets have better effects on improving growth performance and the meat quality of offspring pigs than antibiotics.
Generally, probiotics are live or dynamic microbes that are beneficial to the host’s health. Previous studies have demonstrated that probiotics and synbiotics play a vital role in improving the growth performance of weaned piglets [
16] and growing-finishing pigs [
17] and enhancing weaning BW gain [
18]. In the present study, the BW, ADG, and ADFI of offspring pigs were significantly elevated in the S-OP group during 95–125 d old; however, these indicators had opposite changes in the S-OS group. These findings suggest that probiotics supplementation in sow-offspring diets could promote piglet feed intake and thus influence the growth of offspring pigs during 95–125 d old, which may be related to the improving effects of probiotics on the growth performance of offspring pigs. Several studies have shown that probiotics have positive effects on feed efficiency and BW gain [
19,
20,
21]. However, synbiotics supplementation had an adverse effect on the growth performance of offspring pigs during 95–125 d old in this study. These differences may be related to the types of probiotics and prebiotics and their combination. A previous study also reported that there was no interaction between dietary supplementation of probiotics and xylo-oligosaccharides (XOS) concerning growth performance in weanling piglets [
22]. However, fructo-oligosaccharide combined with
Lactobacillus plantarum ZLP001 as a potential synbiotics showed synergistic effects [
23].
Backfat thickness is positively correlated with the eating quality traits of meat, such as flavor, juiciness, and overall acceptability [
24]. Our results showed that sow-offspring diets supplemented with three additives (antibiotics, probiotics, and synbiotics) significantly decreased the backfat thickness, and dietary antibiotics supplementation significantly decreased the fat percentage and leaf-fat ratio at 95 d old. However, dietary probiotics supplementation significantly increased the loin-eye muscle area and muscle percentage of offspring pigs at 95 d old. Moreover, our results showed that the loin-eye muscle area and muscle percentage of offspring pigs were significantly decreased in the S-OS group at 125 d old. These findings indicate that dietary antibiotics, probiotics, and synbiotics have a negative effect on fat deposition at 95 d old. Furthermore, probiotics and synbiotics supplementation significantly increase muscle mass at 125 d old. Liu et al. [
25] also indicated that combining dietary multi-strain probiotics and
Castanea crenata shell extract can increase the loin-eye muscle area of finishing pigs. In addition, sow-offspring diets supplemented with probiotics significantly increased the fat percentage, backfat thickness, and leaf-fat ratio of offspring pigs at 125 d old in the present study. These results suggest that dietary probiotics could enhance fat deposition, and then improve pork quality.
The physicochemical characteristics of meat affect its appearance and sensory quality [
26]. Drip loss, color, pH, tenderness, and taste of pork are the most critical parameters for consumers [
27]. Drip loss could change the chemical composition and affect the tenderness of meat [
28]. Shear force is particularly important when assessing the value of meat intended for culinary purposes and is also an indicator of meat tenderness [
29]. The pH value is a factor that has a direct influence on the technological properties of meat. A previous study indicated that the eating quality of meat is most desirable at intermediate pH (5.4–6.0) [
30]. In the present study, the pH
24h value ranged from 5.46 to 5.62 in the LDM, suggesting normal glycolysis and glycogenolysis progress in the four treatment groups [
31]. The declining rate of muscle pH is highly relevant to the drip loss and shear force of meat. In the present study, dietary probiotics supplementation significantly increased pH
24h at 65 d old, while synbiotics significantly increased pH
45min at 125 d old. However, probiotics and synbiotics supplementation significantly decreased the drip loss in the LDM at three different days of age in comparison with the Con group. These findings suggest that sow-offspring diets supplemented with probiotics and synbiotics could decrease the risk for PSE meat (pale, soft, and exudative). A recent study also showed that dietary multi-strain probiotics and
Castanea crenata shell extract can reduce drip loss in finishing pigs [
25]. In addition, our results also demonstrated a significant decrease in the shear force of LDM at 125 d old when supplemented with probiotics and synbiotics. These findings suggest that probiotics and synbiotics supplementation in sow-offspring diets may attenuate pH decline and positively affect the postmortem water-holding capacity and tenderness of the LDM.
Meat color is a critical indicator in evaluating muscular appearance, influencing consumers’ purchase decisions as it is deemed a visual measure of freshness and quality [
32]. Cooking yield reflects the water loss during cooking. In the present study, the LDM lightness value of offspring pigs was significantly decreased at 65 d old when sow-offspring diets were supplemented with antibiotics, probiotics, and synbiotics, as well as at 95 d old when supplemented with antibiotics and probiotics; moreover, antibiotics supplementation significantly increased redness value and cooking yield at 95 d old, and synbiotics supplementation significantly increased cooking yield at 65 d old. These findings indicate that antibiotics, probiotics, and synbiotics could improve meat color to some extent when those were supplemented in sow-offspring diets. A previous study reported that
Bacillus subtilis and
Clostridium butyricum supplementation in sow diets increased meat color scores and redness [
33]. Moreover, antibiotics and synbiotics supplementation could decrease the cooking loss of muscle.
Pork is the major animal protein source for humans, and amino acids composition determines muscle protein quality [
34]. We found that sow-offspring diets supplemented with synbiotics significantly decreased the CP content of the PMM at 65 d old, suggesting that synbiotics have a negative effect on the protein accumulation of PMM. Meat with high-quality protein contains all EAA required by the mammalian body [
35]. Several studies demonstrated that Lys, Arg, Asp, and Glu in meat are considered precursors for flavoring substances formation [
36]. Our findings clarified that probiotics and synbiotics supplementation in sow-offspring diets significantly increased the Asp, Arg, and Glu content in the LDM at different stages of age. Histidine is a kind of EAA for infants and children [
37]. In our study, at 125 d old, synbiotics supplementation significantly increased the His content in the LDM, as well as that in the PMM when probiotics and synbiotics were supplemented in sow-offspring diets. A previous study showed that Glu and Asp present a pleasant fresh taste, and Gly, Ala, and Ser present a sweet taste [
38]. In the present study, dietary probiotics and synbiotics supplementation significantly enhanced the Ala content in the LDM at 95 and 125 d old, as well as Pro and Phe at 65 d old and Pro and Tyr at 125 d old in the PMM of offspring pigs. These findings suggest that sow-offspring diets supplemented with probiotics and synbiotics could alter the amino acid composition and then improve the meat flavor. A previous study demonstrated that diets supplemented with
Clostridium butyricum could improve duck meat flavor by changing the FAA content [
39]. Furthermore, significantly higher content of Glu, Gly, Ser, Arg, Val, NEAA, FAA, and TAA were noted in the PMM when sow-offspring diets were supplemented with antibiotics, indicating that antibiotics supplementation could improve meat flavor and nutritional value at 65 d old.
Free amino acids in plasma participate in muscle protein synthesis [
40] and are essential in reflecting muscle nutrition metabolism. Branched-chain amino acids, including Leu, Ile, and Val, play important roles in energy homeostasis and nutritional metabolism [
41]. Furthermore, Tau plays an important role in energy metabolism in the skeletal muscle, heart, liver, and adipose tissues [
42]. The plasma concentrations of Leu, Ile, and Tau were significantly decreased at 65 d old when sow-offspring diets were supplemented with probiotics, as well as at 95 d old when sow-offspring diets were supplemented with probiotics and synbiotics in the present study. These findings suggest that dietary probiotics and synbiotics supplementation in sow-offspring diets are beneficial to muscle energy homeostasis and metabolism. A recent study demonstrated that Ser and Gly are essential for skeletal muscle regeneration [
43]. Our findings showed that sow-offspring diets supplemented with probiotics and synbiotics significantly reduced the plasma Orn and Ser concentrations at 125 d old, indicating that these feed additives may be beneficial for skeletal muscle regeneration. In addition, sow-offspring diets supplemented with probiotics and synbiotics significantly reduced the plasma concentrations of γ-ABA, α-AAA, α-ABA, and β-Ala at 95 d old. Overall, the findings suggest that dietary probiotics and synbiotics supplementation in sow-offspring diets could promote AA deposition in the muscle of offspring pigs. However, further investigations are needed to reveal the underlying mechanism.
The nutritional status of animals could be reflected by the plasma biochemical parameters [
44]. The ALP activity reflects the growth performance and is positively correlated with ADG and calcium and phosphorus utilization rate [
45]. Our study demonstrated that sow-offspring diets supplemented with probiotics and synbiotics significantly increased the plasma ALP activity at 65 d old. However, the ADG of offspring pigs did not significantly change, which may be due to the supplementation of probiotics and synbiotics enhancing calcium and phosphorus utilization, which improved the bone development of offspring pigs. Moreover, the maximum period of bone growth for pigs is in the post-natal 12 weeks [
46]. Increased plasma ALT and AST activities are associated with improved amino acid metabolism [
45]. The deamination of amino acids could produce AMM, which is involved in the synthesis and breakdown of animal proteins or amino acids [
47]. The UN is the main nitrogenous product of protein and amino acid catabolism. Therefore, a decrease in the plasma UN concentration indicates an increase in the ability of amino acids in the blood to synthesize proteins. Furthermore, plasma AMM and UN concentrations could reflect protein metabolism and amino acid balance. Our results showed that sow-offspring diets supplemented with probiotics and synbiotics significantly decreased the plasma UN concentration at 95 d old and AMM concentration at 125 d old, probiotics supplementation significantly decreased the plasma AMM concentration at 95 d old, whereas synbiotics supplementation significantly increased the plasma ALT activity at 95 d old. These results indicate that sow-offspring diets supplemented with probiotics and synbiotics could improve the balance of amino acids and protein utilization.
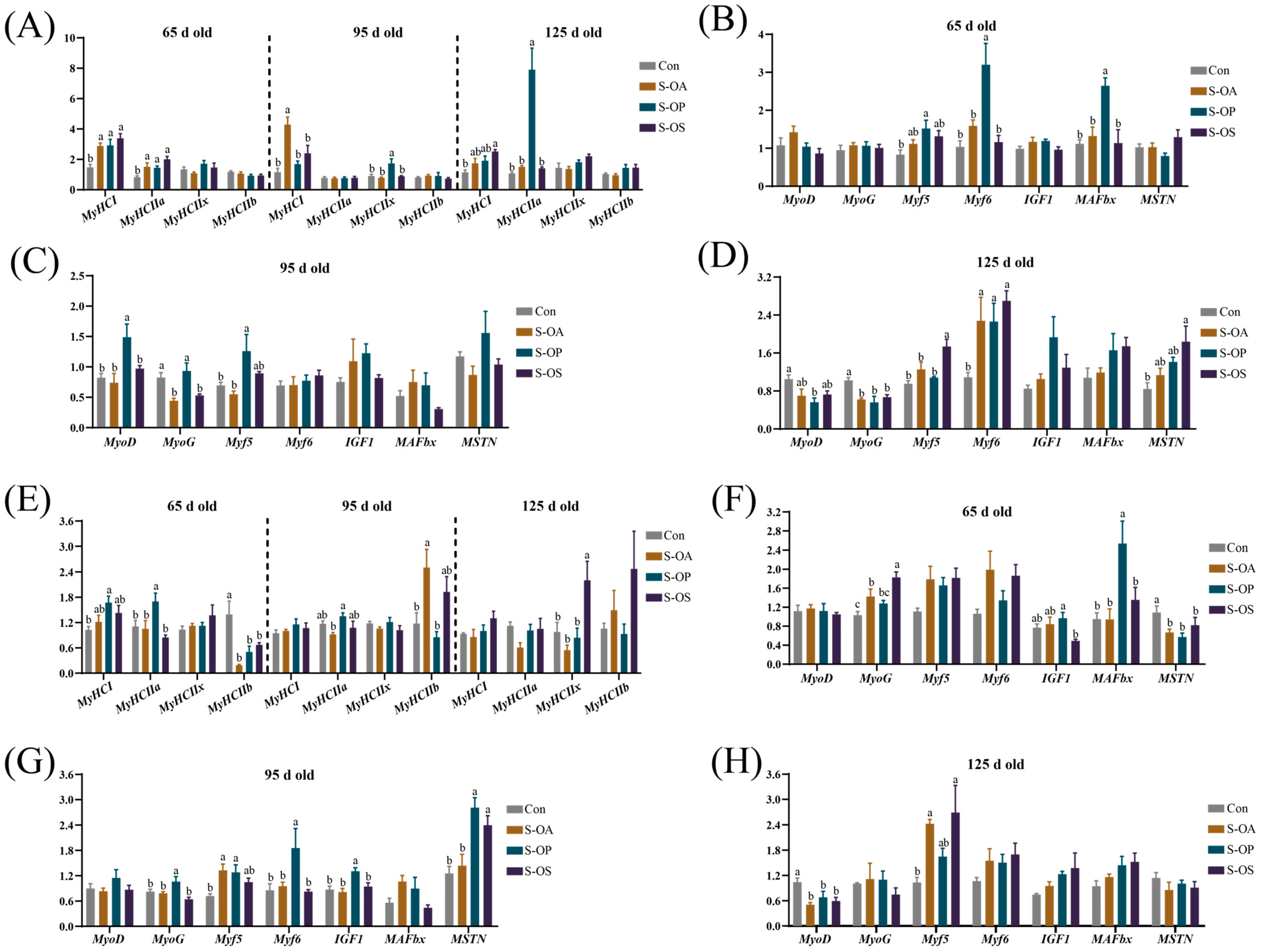
The composition and proportion of muscle-fiber types are closely related to meat quality [
48]. Among the four muscle-fiber types,
MyHCI and
MyHCIIa are positively correlated with high-quality meat, and
MyHCIIa and
MyHCIIx are the critical factors affecting the meat quality [
49]. In our study, at 65 d old, the
MyHCI and
MyHCIIa expressions in the LDM were significantly upregulated by sow-offspring diets supplemented with probiotics and synbiotics, as well as in the PMM by probiotics supplementation, while significantly downregulated
MyHCIIb expression; moreover, at 125 d old,
MyHCIIa expression in the LDM and
MyHCIIx expression in the PMM were significantly upregulated, and synbiotics supplementation significantly upregulated the
MyHCI expression in the LDM. Thus, sow-offspring diets supplemented with probiotics and synbiotics could alter the composition of muscle-fiber types and then improve meat quality. Similarly, Tian et al. [
50] also indicated that dietary
Lactobacillus reuteri 1 supplementation could upregulate the
MyHCI expression and downregulate
MyHCIIb expression in the muscles of pigs. In addition, antibiotics also had positive effects on the muscle-fiber type to some extent.
Muscle regulatory factors play important roles in the proliferation and differentiation of muscle cells, formation and function of muscle fibers, and the muscle maturation [
51].
MyoD and
Myf5 are involved in muscle formation,
MyoG plays a pivotal role in the differentiation of myocytes into muscle fibers, and
Myf6 is essential for muscle maintenance [
52]. In the present study, sow-offspring diets supplemented with probiotics and synbiotics significantly upregulated the
Myf6 expression in the LDM at 125 d old; probiotics supplementation significantly upregulated
Myf5 and
Myf6 expressions in the LDM at 65 d old, and
MyoD and
Myf5 expressions in the LDM and
IGF1,
Myf5,
Myf6, and
MyoG expressions in the PMM at 95 d old; synbiotics supplementation significantly upregulated
MyoG expression in the PMM and
Myf5 expression in the LDM and PMM at 65 d old. These results suggest that probiotics and synbiotics supplementation in sow-offspring diets could promote skeletal muscle growth in offspring pigs.
MSTN is a negative regulator of skeletal muscle mass [
53] and is negatively correlated with muscle growth and development [
54]. Sow-offspring diets supplemented with probiotics and synbiotics significantly downregulated the
MSTN expression in the PMM at 65 d old in the present study. The results suggest that the expressions of genes that suppress muscle growth were downregulated by probiotics and synbiotics supplementation in sow-offspring diets, thereby promoting muscle growth and development in offspring pigs.



 ,
, 



{kind=link}
{kind=link}
{kind=link}