
Ribosome Specialization in Protozoa Parasites
 , and
, and
Abstract
:1. Introduction
1.1. Types of Ribosome Heterogeneity
1.2. Ribosome Heterogeneity and Specialization in Non-Parasitic Protozoa

2. Ribosome Specialization in Plasmodium spp.
Organellar Ribosomes of Plasmodium
3. Ribosome Specialization in Trypanosomatids
3.1. Trypanosoma spp.

3.2. Leishmania spp.



3.3. Trypanosomatid Mitoribosomes
4. Role of Ribosome Specialization in Parasitism
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| aSD | Anti-Shine-Dalgarno region |
| BSF | Bloodstream forms |
| CryoEM | Cryo-Electron Microscopy |
| COI | Cytochrome c oxidase subunit I |
| Cyb | Cytochrome b |
| eIF4E | Eukaryotic initiation factor 4E |
| FMRP | Fragile X Mental Retardation Protein |
| H69 | Helix 69 |
| LSU | Large subunit |
| LiAlba1 | Leishmania infantum Acetylation lowers binding affinity protein 1 |
| LiAlba3 | Leishmania infantum Acetylation lowers binding affinity protein 3 |
| MDa | Mega Dalton |
| mESCs | Mouse embryonic stem cells |
| mRNA | Messenger RNA |
| MRP | Mitochondrial ribosomal protein |
| mtDNA | Mitochondrial DNA |
| mtETC | Mitochondrial electron transport chain |
| mTORC1 | Mammalian target of rapamycin complex 1 |
| ncRNA | Non-coding RNA |
| OOD | Oocyst development defect |
| PCF | Polycyclic forms |
| PDB | Protein database |
| PPR | Pentatricopeptides repeats |
| PTC | Peptidyl transferase center |
| RACK1 | Receptor for activated C kinase 1 |
| rancRNA | Ribosome associated non-coding RNA |
| rDNA | Ribosomal DNA |
| RNP | Ribonucleoprotein |
| RP | Ribosomal protein |
| RPL | Ribosomal protein L |
| RPS | Ribosomal protein S |
| rRNA | Ribosomal RNA |
| SAR | Social amoebae ribosomes |
| scaRNAs | Small Cajal body-specific RNAs |
| snoRNA | Small nucleolar RNA |
| SSU | Small subunit |
| TA | Toxin-Antitoxin |
| TbRACK1 | Trypanosoma brucei Receptor of activated protein C kinase 1 |
| VAR | Vegetative amoebae ribosomes |
| Ψ | Pseudouridylation |
References
- Melnikov, S.; Ben-Shem, A.; Garreau de Loubresse, N.; Jenner, L.; Yusupova, G.; Yusupov, M. One core, two shells: Bacterial and eukaryotic ribosomes. Nat. Struct. Mol. Biol. 2012, 19, 560–567. [Google Scholar] [CrossRef] [PubMed]
- Ben-Shem, A.; Garreau de Loubresse, N.; Melnikov, S.; Jenner, L.; Yusupova, G.; Yusupov, M. The structure of the eukaryotic ribosome at 3.0 A resolution. Science 2011, 334, 1524–1529. [Google Scholar] [CrossRef] [PubMed]
- Field, K.G.; Olsen, G.J.; Lane, D.J.; Giovannoni, S.J.; Ghiselin, M.T.; Raff, E.C.; Pace, N.R.; Raff, R.A. Molecular phylogeny of the animal kingdom. Science 1988, 239, 748–753. [Google Scholar] [CrossRef] [PubMed]
- Xue, S.; Barna, M. Specialized ribosomes: A new frontier in gene regulation and organismal biology. Nat. Rev. Mol. Cell Biol. 2012, 13, 355–369. [Google Scholar] [CrossRef]
- Van Duin, J.; Kurland, C.G. Functional heterogeneity of the 30S ribosomal subunit of E. coli. Mol. Gen. Genet. 1970, 109, 169–176. [Google Scholar] [CrossRef]
- Deusser, E. Heterogeneity of ribosomal populations in Escherichia coli cells grown in different media. Mol. Gen. Genet. 1972, 119, 249–258. [Google Scholar] [CrossRef]
- Leipold, R.J.; Dhurjati, P. Specialized ribosomes in Escherichia coli. Biotechnol. Prog. 1993, 9, 443–449. [Google Scholar] [CrossRef]
- Ramagopal, S. Are eukaryotic ribosomes heterogeneous? Affirmations on the horizon. Biochem. Cell Biol. 1992, 70, 269–272. [Google Scholar] [CrossRef]
- Hui, A.; de Boer, H.A. Specialized ribosome system: Preferential translation of a single mRNA species by a subpopulation of mutated ribosomes in Escherichia coli. Proc. Natl. Acad. Sci. USA 1987, 84, 4762–4766. [Google Scholar] [CrossRef]
- Mauro, V.P.; Edelman, G.M. The ribosome filter hypothesis. Proc. Natl. Acad. Sci. USA 2002, 99, 12031–12036. [Google Scholar] [CrossRef]
- Mauro, V.P.; Edelman, G.M. The ribosome filter redux. Cell Cycle 2007, 6, 2246–2251. [Google Scholar] [CrossRef]
- Shi, Z.; Fujii, K.; Kovary, K.M.; Genuth, N.R.; Rost, H.L.; Teruel, M.N.; Barna, M. Heterogeneous Ribosomes Preferentially Translate Distinct Subpools of mRNAs Genome-wide. Mol. Cell 2017, 67, 71–83.e77. [Google Scholar] [CrossRef]
- Bauer, J.W.; Brandl, C.; Haubenreisser, O.; Wimmer, B.; Weber, M.; Karl, T.; Klausegger, A.; Breitenbach, M.; Hintner, H.; von der Haar, T.; et al. Specialized yeast ribosomes: A customized tool for selective mRNA translation. PLoS ONE 2013, 8, e67609. [Google Scholar] [CrossRef]
- Xue, S.; Tian, S.; Fujii, K.; Kladwang, W.; Das, R.; Barna, M. RNA regulons in Hox 5′ UTRs confer ribosome specificity to gene regulation. Nature 2015, 517, 33–38. [Google Scholar] [CrossRef]
- Simsek, D.; Tiu, G.C.; Flynn, R.A.; Byeon, G.W.; Leppek, K.; Xu, A.F.; Chang, H.Y.; Barna, M. The Mammalian Ribo-interactome Reveals Ribosome Functional Diversity and Heterogeneity. Cell 2017, 169, 1051–1065.e1018. [Google Scholar] [CrossRef]
- Ferretti, M.B.; Ghalei, H.; Ward, E.A.; Potts, E.L.; Karbstein, K. Rps26 directs mRNA-specific translation by recognition of Kozak sequence elements. Nat. Struct. Mol. Biol. 2017, 24, 700–707. [Google Scholar] [CrossRef]
- Miller, S.C.; MacDonald, C.C.; Kellogg, M.K.; Karamysheva, Z.N.; Karamyshev, A.L. Specialized Ribosomes in Health and Disease. Int. J. Mol. Sci. 2023, 24, 6334. [Google Scholar] [CrossRef]
- Genuth, N.R.; Shi, Z.; Kunimoto, K.; Hung, V.; Xu, A.F.; Kerr, C.H.; Tiu, G.C.; Oses-Prieto, J.A.; Salomon-Shulman, R.E.A.; Axelrod, J.D.; et al. A stem cell roadmap of ribosome heterogeneity reveals a function for RPL10A in mesoderm production. Nat. Commun. 2022, 13, 5491. [Google Scholar] [CrossRef]
- Kondrashov, N.; Pusic, A.; Stumpf, C.R.; Shimizu, K.; Hsieh, A.C.; Ishijima, J.; Shiroishi, T.; Barna, M. Ribosome-mediated specificity in Hox mRNA translation and vertebrate tissue patterning. Cell 2011, 145, 383–397. [Google Scholar] [CrossRef]
- Sugihara, Y.; Honda, H.; Iida, T.; Morinaga, T.; Hino, S.; Okajima, T.; Matsuda, T.; Nadano, D. Proteomic analysis of rodent ribosomes revealed heterogeneity including ribosomal proteins L10-like, L22-like 1, and L39-like. J. Proteome Res. 2010, 9, 1351–1366. [Google Scholar] [CrossRef]
- Sugihara, Y.; Sadohara, E.; Yonezawa, K.; Kugo, M.; Oshima, K.; Matsuda, T.; Nadano, D. Identification and expression of an autosomal paralogue of ribosomal protein S4, X-linked, in mice: Potential involvement of testis-specific ribosomal proteins in translation and spermatogenesis. Gene 2013, 521, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Lopes, A.M.; Miguel, R.N.; Sargent, C.A.; Ellis, P.J.; Amorim, A.; Affara, N.A. The human RPS4 paralogue on Yq11.223 encodes a structurally conserved ribosomal protein and is preferentially expressed during spermatogenesis. BMC Mol. Biol. 2010, 11, 33. [Google Scholar] [CrossRef] [PubMed]
- Zou, Q.; Yang, L.; Shi, R.; Qi, Y.; Zhang, X.; Qi, H. Proteostasis regulated by testis-specific ribosomal protein RPL39L maintains mouse spermatogenesis. iScience 2021, 24, 103396. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Zong, Q.; He, Z.; Liu, C.; Wang, Y.F. Knockdown of RpL36 in testes impairs spermatogenesis in Drosophila melanogaster. J. Exp. Zool. Part B Mol. Dev. Evol. 2021, 336, 417–430. [Google Scholar] [CrossRef]
- Mageeney, C.M.; Ware, V.C. Specialized eRpL22 paralogue-specific ribosomes regulate specific mRNA translation in spermatogenesis in Drosophila melanogaster. Mol. Biol. Cell 2019, 30, 2240–2253. [Google Scholar] [CrossRef]
- Li, H.; Huo, Y.; He, X.; Yao, L.; Zhang, H.; Cui, Y.; Xiao, H.; Xie, W.; Zhang, D.; Wang, Y.; et al. A male germ-cell-specific ribosome controls male fertility. Nature 2022, 612, 725–731. [Google Scholar] [CrossRef]
- Ghulam, M.M.; Catala, M.; Abou Elela, S. Differential expression of duplicated ribosomal protein genes modifies ribosome composition in response to stress. Nucleic Acids Res. 2020, 48, 1954–1968. [Google Scholar] [CrossRef]
- Ghulam, M.M.; Catala, M.; Reulet, G.; Scott, M.S.; Abou Elela, S. Duplicated ribosomal protein paralogs promote alternative translation and drug resistance. Nat. Commun. 2022, 13, 4938. [Google Scholar] [CrossRef]
- Guo, H. Specialized ribosomes and the control of translation. Biochem. Soc. Trans. 2018, 46, 855–869. [Google Scholar] [CrossRef]
- Filipovska, A.; Rackham, O. Specialization from synthesis: How ribosome diversity can customize protein function. FEBS Lett. 2013, 587, 1189–1197. [Google Scholar] [CrossRef]
- Sauert, M.; Temmel, H.; Moll, I. Heterogeneity of the translational machinery: Variations on a common theme. Biochimie 2015, 114, 39–47. [Google Scholar] [CrossRef]
- Shi, Z.; Barna, M. Translating the genome in time and space: Specialized ribosomes, RNA regulons, and RNA-binding proteins. Annu. Rev. Cell Dev. Biol. 2015, 31, 31–54. [Google Scholar] [CrossRef]
- Genuth, N.R.; Barna, M. Heterogeneity and specialized functions of translation machinery: From genes to organisms. Nat. Rev. Genet. 2018, 19, 431–452. [Google Scholar] [CrossRef]
- Li, D.; Wang, J. Ribosome heterogeneity in stem cells and development. J. Cell Biol. 2020, 219, e202001108. [Google Scholar] [CrossRef]
- Gay, D.M.; Lund, A.H.; Jansson, M.D. Translational control through ribosome heterogeneity and functional specialization. Trends Biochem. Sci. 2022, 47, 66–81. [Google Scholar] [CrossRef]
- Joo, M.; Yeom, J.H.; Choi, Y.; Jun, H.; Song, W.; Kim, H.L.; Lee, K.; Shin, E. Specialised ribosomes as versatile regulators of gene expression. RNA Biol. 2022, 19, 1103–1114. [Google Scholar] [CrossRef]
- Komili, S.; Farny, N.G.; Roth, F.P.; Silver, P.A. Functional specificity among ribosomal proteins regulates gene expression. Cell 2007, 131, 557–571. [Google Scholar] [CrossRef]
- Imami, K.; Milek, M.; Bogdanow, B.; Yasuda, T.; Kastelic, N.; Zauber, H.; Ishihama, Y.; Landthaler, M.; Selbach, M. Phosphorylation of the Ribosomal Protein RPL12/uL11 Affects Translation during Mitosis. Mol. Cell 2018, 72, 84–98.e89. [Google Scholar] [CrossRef]
- Ladror, D.T.; Frey, B.L.; Scalf, M.; Levenstein, M.E.; Artymiuk, J.M.; Smith, L.M. Methylation of yeast ribosomal protein S2 is elevated during stationary phase growth conditions. Biochem. Biophys. Res. Commun. 2014, 445, 535–541. [Google Scholar] [CrossRef]
- Parks, M.M.; Kurylo, C.M.; Dass, R.A.; Bojmar, L.; Lyden, D.; Vincent, C.T.; Blanchard, S.C. Variant ribosomal RNA alleles are conserved and exhibit tissue-specific expression. Sci. Adv. 2018, 4, eaao0665. [Google Scholar] [CrossRef]
- Decatur, W.A.; Fournier, M.J. rRNA modifications and ribosome function. Trends Biochem. Sci. 2002, 27, 344–351. [Google Scholar] [CrossRef] [PubMed]
- Charette, M.; Gray, M.W. Pseudouridine in RNA: What, where, how, and why. IUBMB Life 2000, 49, 341–351. [Google Scholar] [CrossRef] [PubMed]
- Ofengand, J.; Fournier, M.J. The Pseudouridine Residues of rRNA: Number, Location, Biosynthesis, and Function. Modif. Ed. RNA 2014, 229–253. [Google Scholar] [CrossRef]
- Maden, B.E. The numerous modified nucleotides in eukaryotic ribosomal RNA. Prog. Nucleic Acid Res. Mol. Biol. 1990, 39, 241–303. [Google Scholar] [CrossRef] [PubMed]
- Lane, B.G.; Ofengand, J.; Gray, M.W. Pseudouridine and O2′-methylated nucleosides. Significance of their selective occurrence in rRNA domains that function in ribosome-catalyzed synthesis of the peptide bonds in proteins. Biochimie 1995, 77, 7–15. [Google Scholar] [CrossRef]
- Smith, J.E.; Cooperman, B.S.; Mitchell, P. Methylation sites in Escherichia coli ribosomal RNA: Localization and identification of four new sites of methylation in 23S rRNA. Biochemistry 1992, 31, 10825–10834. [Google Scholar] [CrossRef]
- Jack, K.; Bellodi, C.; Landry, D.M.; Niederer, R.O.; Meskauskas, A.; Musalgaonkar, S.; Kopmar, N.; Krasnykh, O.; Dean, A.M.; Thompson, S.R.; et al. rRNA pseudouridylation defects affect ribosomal ligand binding and translational fidelity from yeast to human cells. Mol. Cell 2011, 44, 660–666. [Google Scholar] [CrossRef]
- Sloan, K.E.; Warda, A.S.; Sharma, S.; Entian, K.D.; Lafontaine, D.L.J.; Bohnsack, M.T. Tuning the ribosome: The influence of rRNA modification on eukaryotic ribosome biogenesis and function. RNA Biol. 2017, 14, 1138–1152. [Google Scholar] [CrossRef]
- Adams, D.R.; Ron, D.; Kiely, P.A. RACK1, A multifaceted scaffolding protein: Structure and function. Cell Commun. Signal. 2011, 9, 22. [Google Scholar] [CrossRef]
- Gallo, S.; Ricciardi, S.; Manfrini, N.; Pesce, E.; Oliveto, S.; Calamita, P.; Mancino, M.; Maffioli, E.; Moro, M.; Crosti, M.; et al. RACK1 Specifically Regulates Translation through Its Binding to Ribosomes. Mol. Cell. Biol. 2018, 38, e00230-18. [Google Scholar] [CrossRef]
- Darnell, J.C.; Van Driesche, S.J.; Zhang, C.; Hung, K.Y.; Mele, A.; Fraser, C.E.; Stone, E.F.; Chen, C.; Fak, J.J.; Chi, S.W.; et al. FMRP stalls ribosomal translocation on mRNAs linked to synaptic function and autism. Cell 2011, 146, 247–261. [Google Scholar] [CrossRef]
- O’Brien, T.W. Evolution of a protein-rich mitochondrial ribosome: Implications for human genetic disease. Gene 2002, 286, 73–79. [Google Scholar] [CrossRef]
- Sharma, M.R.; Koc, E.C.; Datta, P.P.; Booth, T.M.; Spremulli, L.L.; Agrawal, R.K. Structure of the mammalian mitochondrial ribosome reveals an expanded functional role for its component proteins. Cell 2003, 115, 97–108. [Google Scholar] [CrossRef]
- O’Brien, T.W. Properties of human mitochondrial ribosomes. IUBMB Life 2003, 55, 505–513. [Google Scholar] [CrossRef]
- Koc, E.C.; Burkhart, W.; Blackburn, K.; Moseley, A.; Spremulli, L.L. The small subunit of the mammalian mitochondrial ribosome. Identification of the full complement of ribosomal proteins present. J. Biol. Chem. 2001, 276, 19363–19374. [Google Scholar] [CrossRef]
- Koc, E.C.; Burkhart, W.; Blackburn, K.; Moyer, M.B.; Schlatzer, D.M.; Moseley, A.; Spremulli, L.L. The large subunit of the mammalian mitochondrial ribosome. Analysis of the complement of ribosomal proteins present. J. Biol. Chem. 2001, 276, 43958–43969. [Google Scholar] [CrossRef]
- Koc, E.C.; Cimen, H.; Kumcuoglu, B.; Abu, N.; Akpinar, G.; Haque, M.E.; Spremulli, L.L.; Koc, H. Identification and characterization of CHCHD1, AURKAIP1, and CRIF1 as new members of the mammalian mitochondrial ribosome. Front. Physiol. 2013, 4, 183. [Google Scholar] [CrossRef]
- Suzuki, T.; Terasaki, M.; Takemoto-Hori, C.; Hanada, T.; Ueda, T.; Wada, A.; Watanabe, K. Structural compensation for the deficit of rRNA with proteins in the mammalian mitochondrial ribosome. Systematic analysis of protein components of the large ribosomal subunit from mammalian mitochondria. J. Biol. Chem. 2001, 276, 21724–21736. [Google Scholar] [CrossRef]
- Suzuki, T.; Terasaki, M.; Takemoto-Hori, C.; Hanada, T.; Ueda, T.; Wada, A.; Watanabe, K. Proteomic analysis of the mammalian mitochondrial ribosome. Identification of protein components in the 28 S small subunit. J. Biol. Chem. 2001, 276, 33181–33195. [Google Scholar] [CrossRef]
- Ott, M.; Herrmann, J.M. Co-translational membrane insertion of mitochondrially encoded proteins. Biochim. Biophys. Acta 2010, 1803, 767–775. [Google Scholar] [CrossRef]
- Yusupova, G.; Yusupov, M. High-resolution structure of the eukaryotic 80S ribosome. Annu. Rev. Biochem. 2014, 83, 467–486. [Google Scholar] [CrossRef] [PubMed]
- Desai, N.; Brown, A.; Amunts, A.; Ramakrishnan, V. The structure of the yeast mitochondrial ribosome. Science 2017, 355, 528–531. [Google Scholar] [CrossRef]
- Ramrath, D.J.F.; Niemann, M.; Leibundgut, M.; Bieri, P.; Prange, C.; Horn, E.K.; Leitner, A.; Boehringer, D.; Schneider, A.; Ban, N. Evolutionary shift toward protein-based architecture in trypanosomal mitochondrial ribosomes. Science 2018, 362, eaau7735. [Google Scholar] [CrossRef] [PubMed]
- Amunts, A.; Brown, A.; Toots, J.; Scheres, S.H.W.; Ramakrishnan, V. Ribosome. The structure of the human mitochondrial ribosome. Science 2015, 348, 95–98. [Google Scholar] [CrossRef] [PubMed]
- Zikova, A.; Panigrahi, A.K.; Dalley, R.A.; Acestor, N.; Anupama, A.; Ogata, Y.; Myler, P.J.; Stuart, K. Trypanosoma brucei mitochondrial ribosomes: Affinity purification and component identification by mass spectrometry. Mol. Cell. Proteom. 2008, 7, 1286–1296. [Google Scholar] [CrossRef]
- Maslov, D.A.; Sharma, M.R.; Butler, E.; Falick, A.M.; Gingery, M.; Agrawal, R.K.; Spremulli, L.L.; Simpson, L. Isolation and characterization of mitochondrial ribosomes and ribosomal subunits from Leishmania tarentolae. Mol. Biochem. Parasitol. 2006, 148, 69–78. [Google Scholar] [CrossRef]
- Kessin, R.H. Dictyostelium: Evolution, Cell Biology, and the Development of Multicellularity; Cambridge University Press: Cambridge, UK, 2010. [Google Scholar]
- Dunn, J.D.; Bosmani, C.; Barisch, C.; Raykov, L.; Lefrancois, L.H.; Cardenal-Munoz, E.; Lopez-Jimenez, A.T.; Soldati, T. Eat Prey, Live: Dictyostelium discoideum As a Model for Cell-Autonomous Defenses. Front. Immunol. 2017, 8, 1906. [Google Scholar] [CrossRef]
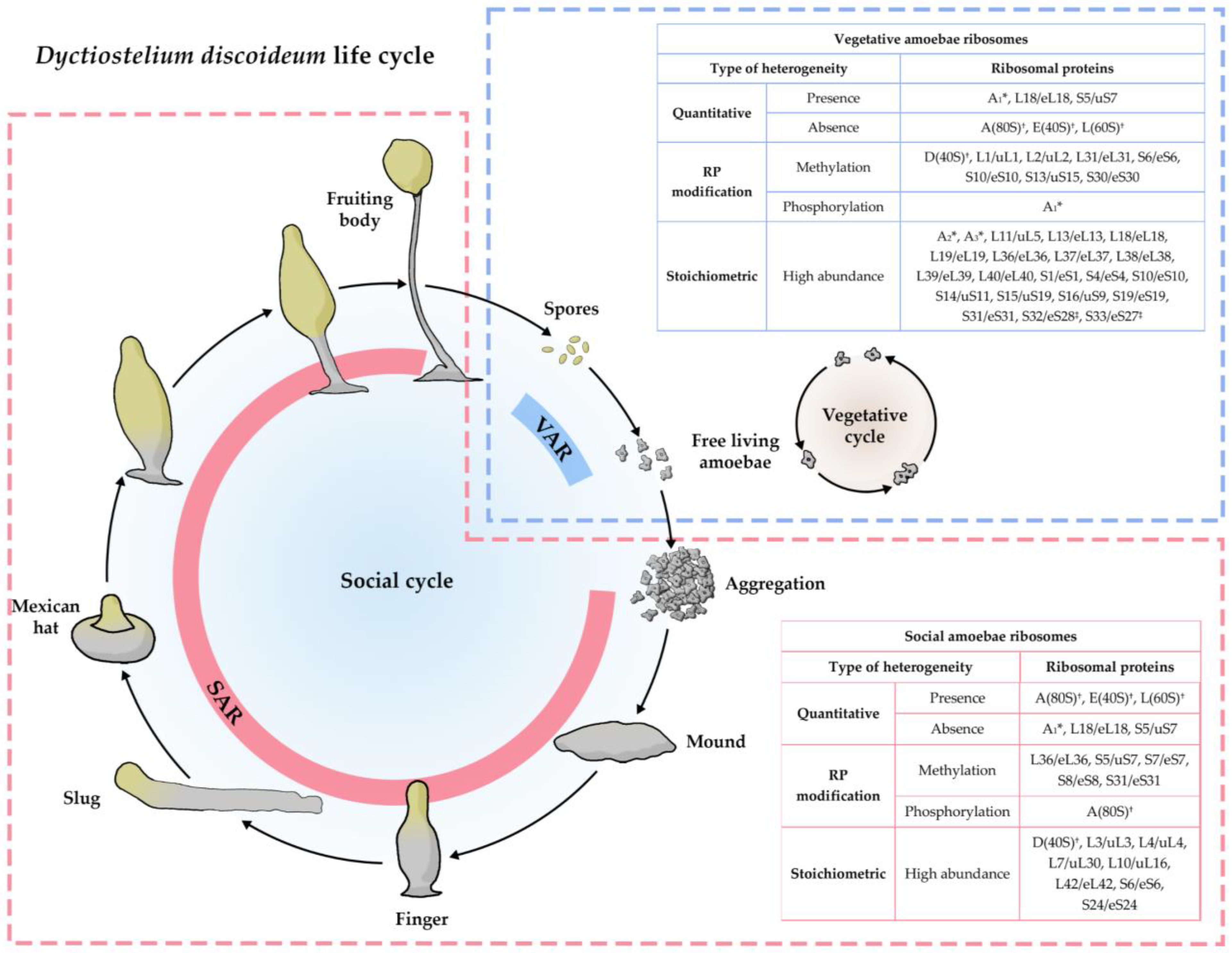
- Ramagopal, S.; Ennis, H.L. Regulation of synthesis of cell-specific ribosomal proteins during differentiation of Dictyostelium discoideum. Proc. Natl. Acad. Sci. USA 1981, 78, 3083–3087. [Google Scholar] [CrossRef]
- Ramagopal, S. Induction of cell-specific ribosomal proteins in aggregation-competent nonmorphogenetic Dictyostelium discoideum. Biochem. Cell Biol. 1990, 68, 1281–1287. [Google Scholar] [CrossRef]
- Ramagopal, S. Covalent modifications of ribosomal proteins in growing and aggregation-competent Dictyostelium discoideum: Phosphorylation and methylation. Biochem. Cell Biol. 1991, 69, 263–268. [Google Scholar] [CrossRef]
- Ramagopal, S. The Dictyostelium ribosome: Biochemistry, molecular biology, and developmental regulation. Biochem. Cell Biol. 1992, 70, 738–750. [Google Scholar] [CrossRef]
- Ban, N.; Beckmann, R.; Cate, J.H.; Dinman, J.D.; Dragon, F.; Ellis, S.R.; Lafontaine, D.L.; Lindahl, L.; Liljas, A.; Lipton, J.M.; et al. A new system for naming ribosomal proteins. Curr. Opin. Struct. Biol. 2014, 24, 165–169. [Google Scholar] [CrossRef]
- Moore, J. Studies of the Relationship between mRNA Stability and Gene Function in Saccharomyces cerevisiae; University of Glasgow: Glasgow, UK, 1991. [Google Scholar]
- Vembar, S.S.; Droll, D.; Scherf, A. Translational regulation in blood stages of the malaria parasite Plasmodium spp.: Systems-wide studies pave the way. Wiley Interdiscip. Rev. RNA 2016, 7, 772–792. [Google Scholar] [CrossRef]
- McCutchan, T.F.; Li, J.; McConkey, G.A.; Rogers, M.J.; Waters, A.P. The cytoplasmic ribosomal RNAs of Plasmodium spp. Parasitol. Today 1995, 11, 134–138. [Google Scholar] [CrossRef]
- Gunderson, J.H.; Sogin, M.L.; Wollett, G.; Hollingdale, M.; de la Cruz, V.F.; Waters, A.P.; McCutchan, T.F. Structurally distinct, stage-specific ribosomes occur in Plasmodium. Science 1987, 238, 933–937. [Google Scholar] [CrossRef]
- Waters, A.P.; Syin, C.; McCutchan, T.F. Developmental regulation of stage-specific ribosome populations in Plasmodium. Nature 1989, 342, 438–440. [Google Scholar] [CrossRef]
- Li, J.; McConkey, G.A.; Rogers, M.J.; Waters, A.P.; McCutchan, T.R. Plasmodium: The developmentally regulated ribosome. Exp. Parasitol. 1994, 78, 437–441. [Google Scholar] [CrossRef]
- Waters, A.P.; van Spaendonk, R.M.; Ramesar, J.; Vervenne, R.A.; Dirks, R.W.; Thompson, J.; Janse, C.J. Species-specific regulation and switching of transcription between stage-specific ribosomal RNA genes in Plasmodium berghei. J. Biol. Chem. 1997, 272, 3583–3589. [Google Scholar] [CrossRef]
- Thompson, J.; van Spaendonk, R.M.; Choudhuri, R.; Sinden, R.E.; Janse, C.J.; Waters, A.P. Heterogeneous ribosome populations are present in Plasmodium berghei during development in its vector. Mol. Microbiol. 1999, 31, 253–260. [Google Scholar] [CrossRef]
- Velichutina, I.V.; Rogers, M.J.; McCutchan, T.F.; Liebman, S.W. Chimeric rRNAs containing the GTPase centers of the developmentally regulated ribosomal rRNAs of Plasmodium falciparum are functionally distinct. RNA 1998, 4, 594–602. [Google Scholar] [CrossRef]
- Qi, Y.; Zhu, F.; Eastman, R.T.; Fu, Y.; Zilversmit, M.; Pattaradilokrat, S.; Hong, L.; Liu, S.; McCutchan, T.F.; Pan, W.; et al. Regulation of Plasmodium yoelii oocyst development by strain- and stage-specific small-subunit rRNA. mBio 2015, 6, e00117. [Google Scholar] [CrossRef] [PubMed]
- van Spaendonk, R.M.; Ramesar, J.; van Wigcheren, A.; Eling, W.; Beetsma, A.L.; van Gemert, G.J.; Hooghof, J.; Janse, C.J.; Waters, A.P. Functional equivalence of structurally distinct ribosomes in the malaria parasite, Plasmodium berghei. J. Biol. Chem. 2001, 276, 22638–22647. [Google Scholar] [CrossRef] [PubMed]
- Carraway, M.; Tzipori, S.; Widmer, G. Identification of genetic heterogeneity in the Cryptosporidium parvum ribosomal repeat. Appl. Environ. Microbiol. 1996, 62, 712–716. [Google Scholar] [CrossRef] [PubMed]
- Le Blancq, S.M.; Khramtsov, N.V.; Zamani, F.; Upton, S.J.; Wu, T.W. Ribosomal RNA gene organization in Cryptosporidium parvum. Mol. Biochem. Parasitol. 1997, 90, 463–478. [Google Scholar] [CrossRef]
- Erath, J.; Djuranovic, S. Association of the receptor for activated C-kinase 1 with ribosomes in Plasmodium falciparum. J. Biol. Chem. 2022, 298, 101954. [Google Scholar] [CrossRef]
- Vaidya, A.B.; Mather, M.W. Mitochondrial evolution and functions in malaria parasites. Annu. Rev. Microbiol. 2009, 63, 249–267. [Google Scholar] [CrossRef] [PubMed]
- Evers, F.; Cabrera-Orefice, A.; Elurbe, D.M.; Kea-Te Lindert, M.; Boltryk, S.D.; Voss, T.S.; Huynen, M.A.; Brandt, U.; Kooij, T.W.A. Composition and stage dynamics of mitochondrial complexes in Plasmodium falciparum. Nat. Commun. 2021, 12, 3820. [Google Scholar] [CrossRef]
- Feagin, J.E.; Harrell, M.I.; Lee, J.C.; Coe, K.J.; Sands, B.H.; Cannone, J.J.; Tami, G.; Schnare, M.N.; Gutell, R.R. The fragmented mitochondrial ribosomal RNAs of Plasmodium falciparum. PLoS ONE 2012, 7, e38320. [Google Scholar] [CrossRef]
- Dass, S.; Mather, M.W.; Morrisey, J.M.; Ling, L.; Vaidya, A.B.; Ke, H. Transcriptional changes in Plasmodium falciparum upon conditional knock down of mitochondrial ribosomal proteins RSM22 and L23. PLoS ONE 2022, 17, e0274993. [Google Scholar] [CrossRef]
- Gupta, A.; Shah, P.; Haider, A.; Gupta, K.; Siddiqi, M.I.; Ralph, S.A.; Habib, S. Reduced ribosomes of the apicoplast and mitochondrion of Plasmodium spp. and predicted interactions with antibiotics. Open Biol. 2014, 4, 140045. [Google Scholar] [CrossRef]
- Wilson, R.J.; Denny, P.W.; Preiser, P.R.; Rangachari, K.; Roberts, K.; Roy, A.; Whyte, A.; Strath, M.; Moore, D.J.; Moore, P.W.; et al. Complete gene map of the plastid-like DNA of the malaria parasite Plasmodium falciparum. J. Mol. Biol. 1996, 261, 155–172. [Google Scholar] [CrossRef]
- Arisue, N.; Hashimoto, T.; Mitsui, H.; Palacpac, N.M.; Kaneko, A.; Kawai, S.; Hasegawa, M.; Tanabe, K.; Horii, T. The Plasmodium apicoplast genome: Conserved structure and close relationship of P. ovale to rodent malaria parasites. Mol. Biol. Evol. 2012, 29, 2095–2099. [Google Scholar] [CrossRef]
- Ke, H.; Dass, S.; Morrisey, J.M.; Mather, M.W.; Vaidya, A.B. The mitochondrial ribosomal protein L13 is critical for the structural and functional integrity of the mitochondrion in Plasmodium falciparum. J. Biol. Chem. 2018, 293, 8128–8137. [Google Scholar] [CrossRef]
- Ling, L.; Mulaka, M.; Munro, J.; Dass, S.; Mather, M.W.; Riscoe, M.K.; Llinas, M.; Zhou, J.; Ke, H. Genetic ablation of the mitoribosome in the malaria parasite Plasmodium falciparum sensitizes it to antimalarials that target mitochondrial functions. J. Biol. Chem. 2020, 295, 7235–7248. [Google Scholar] [CrossRef]
- Kampoun, T.; Srichairatanakool, S.; Prommana, P.; Shaw, P.J.; Green, J.L.; Knuepfer, E.; Holder, A.A.; Uthaipibull, C. Apicoplast ribosomal protein S10-V127M enhances artemisinin resistance of a Kelch13 transgenic Plasmodium falciparum. Malar. J. 2022, 21, 302. [Google Scholar] [CrossRef]
- Zhang, M.; Wang, C.; Oberstaller, J.; Thomas, P.; Otto, T.D.; Casandra, D.; Boyapalle, S.; Adapa, S.R.; Xu, S.; Button-Simons, K.; et al. The apicoplast link to fever-survival and artemisinin-resistance in the malaria parasite. Nat. Commun. 2021, 12, 4563. [Google Scholar] [CrossRef]
- Simpson, L. Kinetoplast DNA in trypanosomid flagellates. Int. Rev. Cytol. 1986, 99, 119–179. [Google Scholar] [CrossRef]
- Walder, J.A.; Eder, P.S.; Engman, D.M.; Brentano, S.T.; Walder, R.Y.; Knutzon, D.S.; Dorfman, D.M.; Donelson, J.E. The 35-nucleotide spliced leader sequence is common to all trypanosome messenger RNA’s. Science 1986, 233, 569–571. [Google Scholar] [CrossRef]
- Lewdorowicz, M.; Yoffe, Y.; Zuberek, J.; Jemielity, J.; Stepinski, J.; Kierzek, R.; Stolarski, R.; Shapira, M.; Darzynkiewicz, E. Chemical synthesis and binding activity of the trypanosomatid cap-4 structure. RNA 2004, 10, 1469–1478. [Google Scholar] [CrossRef]
- White, T.C.; Rudenko, G.; Borst, P. Three small RNAs within the 10 kb trypanosome rRNA transcription unit are analogous to domain VII of other eukaryotic 28S rRNAs. Nucleic Acids Res. 1986, 14, 9471–9489. [Google Scholar] [CrossRef]
- Campbell, D.A.; Kubo, K.; Clark, C.G.; Boothroyd, J.C. Precise identification of cleavage sites involved in the unusual processing of trypanosome ribosomal RNA. J. Mol. Biol. 1987, 196, 113–124. [Google Scholar] [CrossRef] [PubMed]
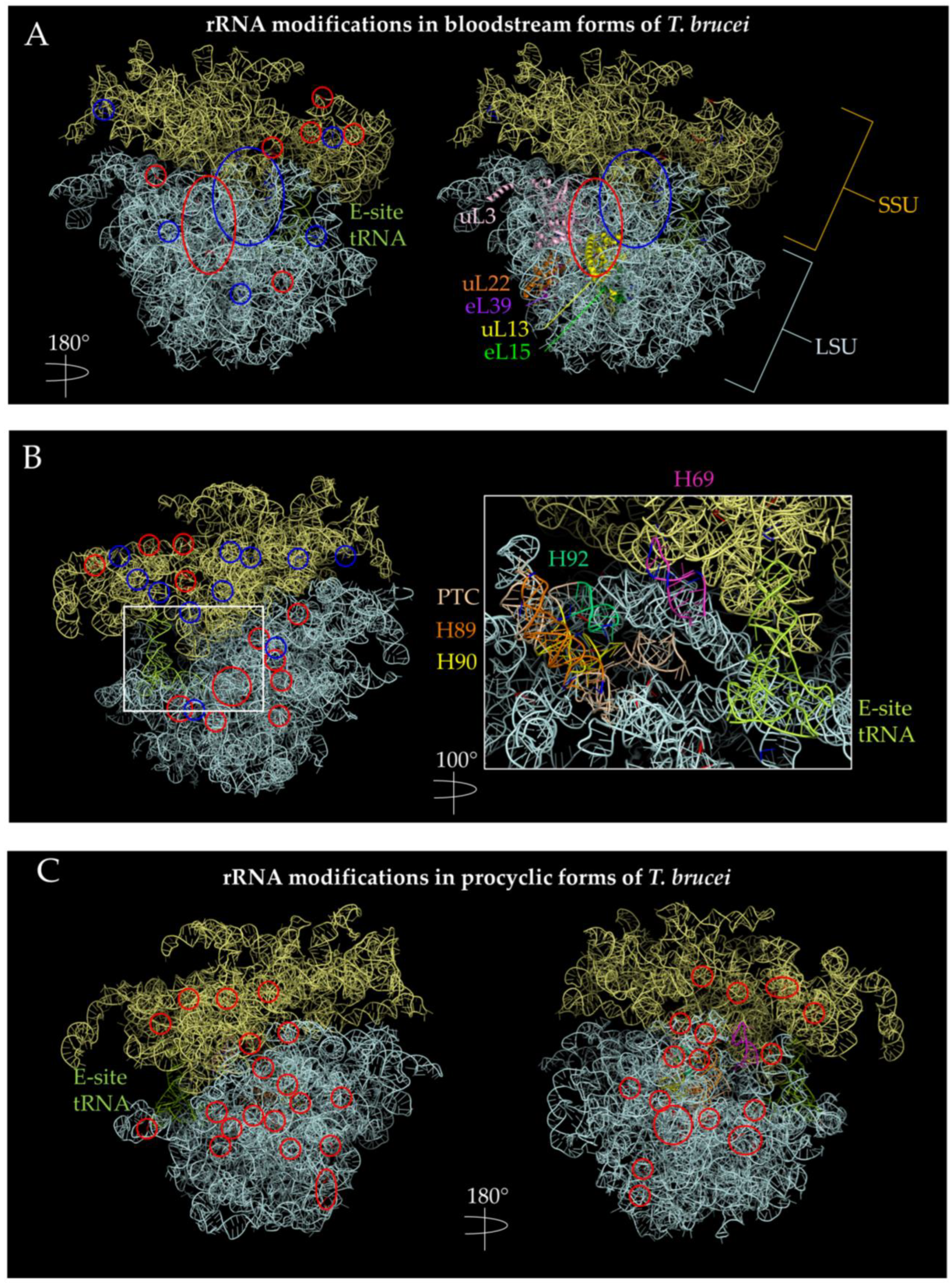
- Hashem, Y.; des Georges, A.; Fu, J.; Buss, S.N.; Jossinet, F.; Jobe, A.; Zhang, Q.; Liao, H.Y.; Grassucci, R.A.; Bajaj, C.; et al. High-resolution cryo-electron microscopy structure of the Trypanosoma brucei ribosome. Nature 2013, 494, 385–389. [Google Scholar] [CrossRef]
- Liu, Z.; Gutierrez-Vargas, C.; Wei, J.; Grassucci, R.A.; Ramesh, M.; Espina, N.; Sun, M.; Tutuncuoglu, B.; Madison-Antenucci, S.; Woolford, J.L., Jr.; et al. Structure and assembly model for the Trypanosoma cruzi 60S ribosomal subunit. Proc. Natl. Acad. Sci. USA 2016, 113, 12174–12179. [Google Scholar] [CrossRef]
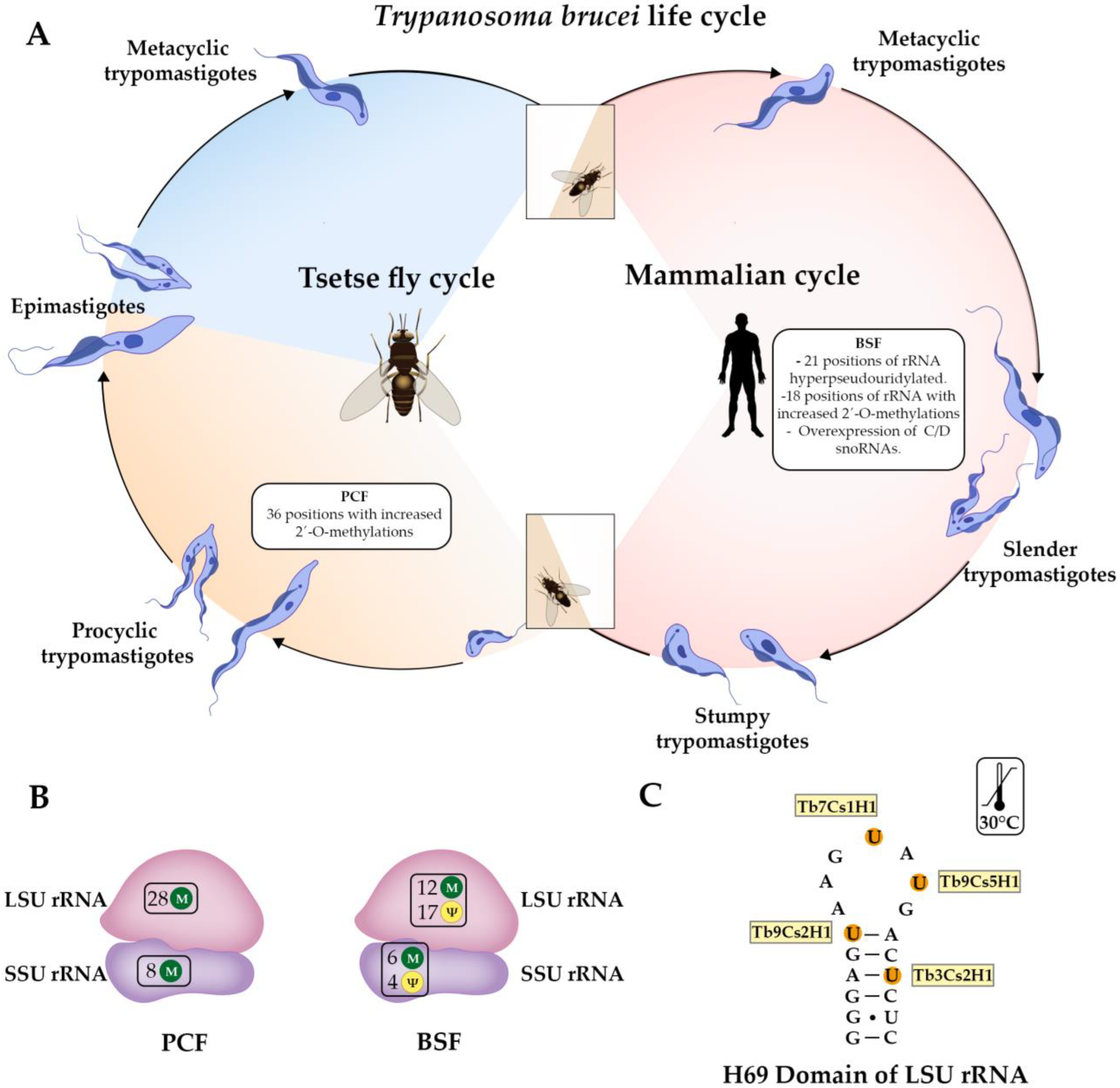
- Chikne, V.; Doniger, T.; Rajan, K.S.; Bartok, O.; Eliaz, D.; Cohen-Chalamish, S.; Tschudi, C.; Unger, R.; Hashem, Y.; Kadener, S.; et al. A pseudouridylation switch in rRNA is implicated in ribosome function during the life cycle of Trypanosoma brucei. Sci. Rep. 2016, 6, 25296. [Google Scholar] [CrossRef] [PubMed]
- Rajan, K.S.; Zhu, Y.; Adler, K.; Doniger, T.; Cohen-Chalamish, S.; Srivastava, A.; Shalev-Benami, M.; Matzov, D.; Unger, R.; Tschudi, C.; et al. The large repertoire of 2′-O-methylation guided by C/D snoRNAs on Trypanosoma brucei rRNA. RNA Biol. 2020, 17, 1018–1039. [Google Scholar] [CrossRef]
- Avila, C.C.; Mule, S.N.; Rosa-Fernandes, L.; Viner, R.; Barison, M.J.; Costa-Martins, A.G.; Oliveira, G.S.; Teixeira, M.M.G.; Marinho, C.R.F.; Silber, A.M.; et al. Proteome-Wide Analysis of Trypanosoma cruzi Exponential and Stationary Growth Phases Reveals a Subcellular Compartment-Specific Regulation. Genes 2018, 9, 413. [Google Scholar] [CrossRef]
- Regmi, S.; Rothberg, K.G.; Hubbard, J.G.; Ruben, L. The RACK1 signal anchor protein from Trypanosoma brucei associates with eukaryotic elongation factor 1A: A role for translational control in cytokinesis. Mol. Microbiol. 2008, 70, 724–745. [Google Scholar] [CrossRef]
- The PyMOL Molecular Graphics System, version 2.5; Schrödinger, LLC: New York, NY, USA, 2022.
- Piel, L.; Rajan, K.S.; Bussotti, G.; Varet, H.; Legendre, R.; Proux, C.; Douche, T.; Giai-Gianetto, Q.; Chaze, T.; Cokelaer, T.; et al. Experimental evolution links post-transcriptional regulation to Leishmania fitness gain. PLoS Pathog. 2022, 18, e1010375. [Google Scholar] [CrossRef] [PubMed]
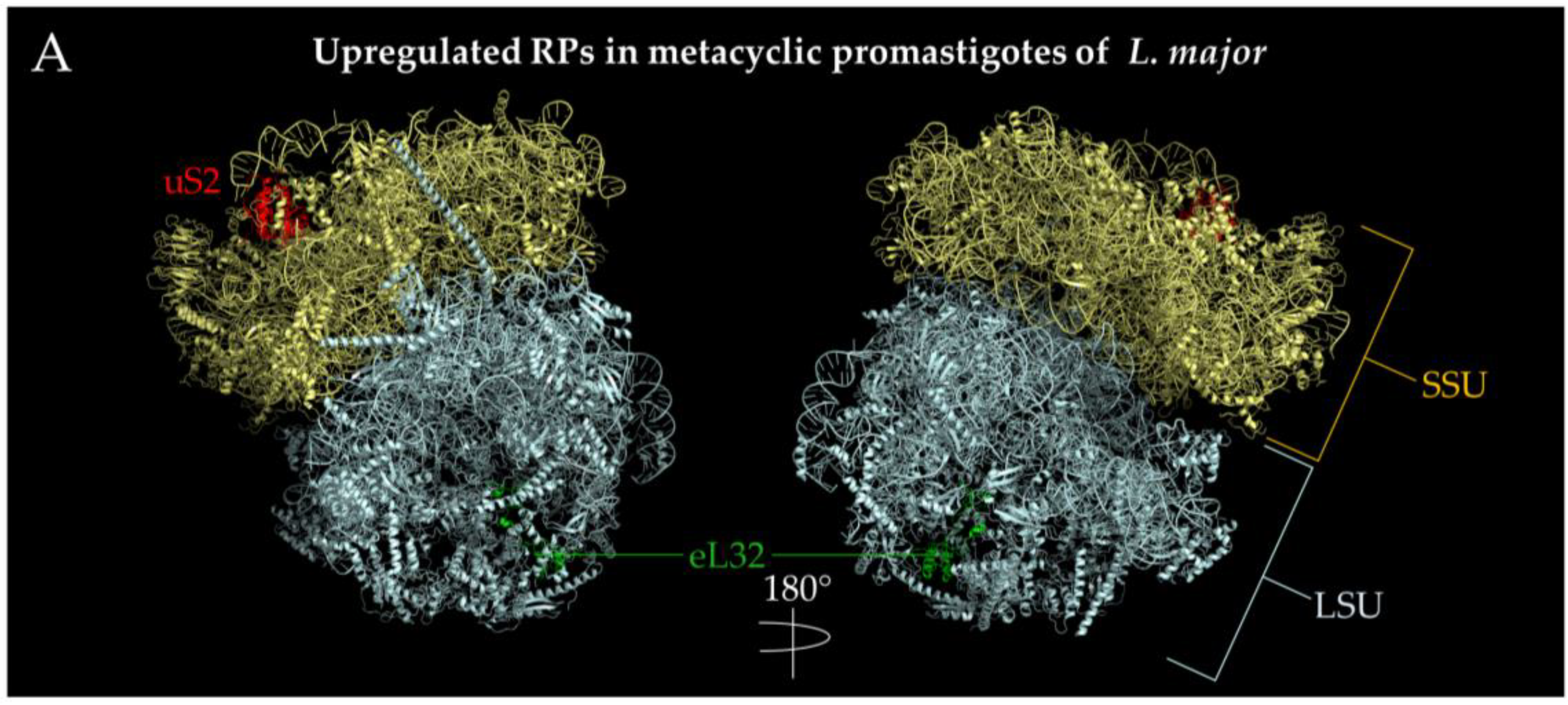
- Amiri-Dashatan, N.; Rezaei-Tavirani, M.; Zali, H.; Koushki, M.; Ahmadi, N. Quantitative proteomic analysis reveals differentially expressed proteins in Leishmania major metacyclogenesis. Microb. Pathog. 2020, 149, 104557. [Google Scholar] [CrossRef]
- Ashrafmansouri, M.; Amiri-Dashatan, N.; Ahmadi, N. Identification of protein profile in metacyclic and amastigote-like stages of Leishmania tropica: A proteomic approach. AMB Express 2022, 12, 142. [Google Scholar] [CrossRef]
- Biyani, N.; Madhubala, R. Quantitative proteomic profiling of the promastigotes and the intracellular amastigotes of Leishmania donovani isolates identifies novel proteins having a role in Leishmania differentiation and intracellular survival. Biochim. Biophys. Acta 2012, 1824, 1342–1350. [Google Scholar] [CrossRef]
- Bratkovic, T.; Rogelj, B. The many faces of small nucleolar RNAs. Biochim. Biophys. Acta 2014, 1839, 438–443. [Google Scholar] [CrossRef]
- van der Werf, J.; Chin, C.V.; Fleming, N.I. SnoRNA in Cancer Progression, Metastasis and Immunotherapy Response. Biology 2021, 10, 809. [Google Scholar] [CrossRef]
- Huang, Z.H.; Du, Y.P.; Wen, J.T.; Lu, B.F.; Zhao, Y. snoRNAs: Functions and mechanisms in biological processes, and roles in tumor pathophysiology. Cell Death Discov. 2022, 8, 259. [Google Scholar] [CrossRef]
- Chikne, V.; Shanmugha Rajan, K.; Shalev-Benami, M.; Decker, K.; Cohen-Chalamish, S.; Madmoni, H.; Biswas, V.K.; Kumar Gupta, S.; Doniger, T.; Unger, R.; et al. Small nucleolar RNAs controlling rRNA processing in Trypanosoma brucei. Nucleic Acids Res. 2019, 47, 2609–2629. [Google Scholar] [CrossRef]
- Martinez-Calvillo, S.; Florencio-Martinez, L.E.; Nepomuceno-Mejia, T. Nucleolar Structure and Function in Trypanosomatid Protozoa. Cells 2019, 8, 421. [Google Scholar] [CrossRef]
- Dupe, A.; Dumas, C.; Papadopoulou, B. Differential Subcellular Localization of Leishmania Alba-Domain Proteins throughout the Parasite Development. PLoS ONE 2015, 10, e0137243. [Google Scholar] [CrossRef]
- Zhang, X.; Lai, M.; Chang, W.; Yu, I.; Ding, K.; Mrazek, J.; Ng, H.L.; Yang, O.O.; Maslov, D.A.; Zhou, Z.H. Structures and stabilization of kinetoplastid-specific split rRNAs revealed by comparing leishmanial and human ribosomes. Nat. Commun. 2016, 7, 13223. [Google Scholar] [CrossRef]
- Dass, S.; Mather, M.W.; Ke, H. Divergent Mitochondrial Ribosomes in Unicellular Parasitic Protozoans. Trends Parasitol. 2020, 36, 318–321. [Google Scholar] [CrossRef]
- Gahura, O.; Chauhan, P.; Zikova, A. Mechanisms and players of mitoribosomal biogenesis revealed in trypanosomatids. Trends Parasitol. 2022, 38, 1053–1067. [Google Scholar] [CrossRef]
- Bieri, P.; Greber, B.J.; Ban, N. High-resolution structures of mitochondrial ribosomes and their functional implications. Curr. Opin. Struct. Biol. 2018, 49, 44–53. [Google Scholar] [CrossRef] [PubMed]
- Maslov, D.A.; Spremulli, L.L.; Sharma, M.R.; Bhargava, K.; Grasso, D.; Falick, A.M.; Agrawal, R.K.; Parker, C.E.; Simpson, L. Proteomics and electron microscopic characterization of the unusual mitochondrial ribosome-related 45S complex in Leishmania tarentolae. Mol. Biochem. Parasitol. 2007, 152, 203–212. [Google Scholar] [CrossRef]
- Ridlon, L.; Skodova, I.; Pan, S.; Lukes, J.; Maslov, D.A. The importance of the 45 S ribosomal small subunit-related complex for mitochondrial translation in Trypanosoma brucei. J. Biol. Chem. 2013, 288, 32963–32978. [Google Scholar] [CrossRef] [PubMed]
- Aphasizheva, I.; Maslov, D.A.; Aphasizhev, R. Kinetoplast DNA-encoded ribosomal protein S12: A possible functional link between mitochondrial RNA editing and translation in Trypanosoma brucei. RNA Biol. 2013, 10, 1679–1688. [Google Scholar] [CrossRef] [PubMed]
- Brown, S.V.; Hosking, P.; Li, J.; Williams, N. ATP synthase is responsible for maintaining mitochondrial membrane potential in bloodstream form Trypanosoma brucei. Eukaryot. Cell 2006, 5, 45–53. [Google Scholar] [CrossRef]
- Schnaufer, A.; Clark-Walker, G.D.; Steinberg, A.G.; Stuart, K. The F1-ATP synthase complex in bloodstream stage trypanosomes has an unusual and essential function. EMBO J. 2005, 24, 4029–4040. [Google Scholar] [CrossRef]
- Ginger, M.L. Niche metabolism in parasitic protozoa. Philos. Trans. R. Soc. B Biol. Sci. 2006, 361, 101–118. [Google Scholar] [CrossRef]
- Zhang, X.; Deitsch, K.W.; Kirkman, L.A. The contribution of extrachromosomal DNA to genome plasticity in malaria parasites. Mol. Microbiol. 2021, 115, 503–507. [Google Scholar] [CrossRef]
- Laffitte, M.N.; Leprohon, P.; Papadopoulou, B.; Ouellette, M. Plasticity of the Leishmania genome leading to gene copy number variations and drug resistance. F1000Res 2016, 5, 2350. [Google Scholar] [CrossRef] [PubMed]
- Santi, A.M.M.; Murta, S.M.F. Impact of Genetic Diversity and Genome Plasticity of Leishmania spp. in Treatment and the Search for Novel Chemotherapeutic Targets. Front. Cell. Infect. Microbiol. 2022, 12, 826287. [Google Scholar] [CrossRef]
- Cruz-Saavedra, L.; Vallejo, G.A.; Guhl, F.; Messenger, L.A.; Ramirez, J.D. Transcriptional remodeling during metacyclogenesis in Trypanosoma cruzi I. Virulence 2020, 11, 969–980. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Saavedra, L.; Vallejo, G.A.; Guhl, F.; Ramirez, J.D. Transcriptomic changes across the life cycle of Trypanosoma cruzi II. PeerJ 2020, 8, e8947. [Google Scholar] [CrossRef] [PubMed]
- Fiebig, M.; Kelly, S.; Gluenz, E. Comparative Life Cycle Transcriptomics Revises Leishmania mexicana Genome Annotation and Links a Chromosome Duplication with Parasitism of Vertebrates. PLoS Pathog. 2015, 11, e1005186. [Google Scholar] [CrossRef]
- Vasquez, J.J.; Hon, C.C.; Vanselow, J.T.; Schlosser, A.; Siegel, T.N. Comparative ribosome profiling reveals extensive translational complexity in different Trypanosoma brucei life cycle stages. Nucleic Acids Res. 2014, 42, 3623–3637. [Google Scholar] [CrossRef] [PubMed]
- Atwood, J.A., 3rd; Weatherly, D.B.; Minning, T.A.; Bundy, B.; Cavola, C.; Opperdoes, F.R.; Orlando, R.; Tarleton, R.L. The Trypanosoma cruzi proteome. Science 2005, 309, 473–476. [Google Scholar] [CrossRef]
- Amorim, J.C.; Batista, M.; da Cunha, E.S.; Lucena, A.C.R.; Lima, C.V.P.; Sousa, K.; Krieger, M.A.; Marchini, F.K. Quantitative proteome and phosphoproteome analyses highlight the adherent population during Trypanosoma cruzi metacyclogenesis. Sci. Rep. 2017, 7, 9899. [Google Scholar] [CrossRef]
- Amiri-Dashatan, N.; Ahmadi, N.; Rezaei-Tavirani, M.; Koushki, M. Identification of differential protein expression and putative drug target in metacyclic stage of Leishmania major and Leishmania tropica: A quantitative proteomics and computational view. Comp. Immunol. Microbiol. Infect. Dis. 2021, 75, 101617. [Google Scholar] [CrossRef]
- Wang, Z.X.; Zhou, C.X.; Elsheikha, H.M.; He, S.; Zhou, D.H.; Zhu, X.Q. Proteomic Differences between Developmental Stages of Toxoplasma gondii Revealed by iTRAQ-Based Quantitative Proteomics. Front. Microbiol. 2017, 8, 985. [Google Scholar] [CrossRef]
- Wilson, D.M.; Deutsch, W.A.; Kelley, M.R. Drosophila ribosomal protein S3 contains an activity that cleaves DNA at apurinic/apyrimidinic sites. J. Biol. Chem. 1994, 269, 25359–25364. [Google Scholar] [CrossRef]
- Graifer, D.; Malygin, A.; Zharkov, D.O.; Karpova, G. Eukaryotic ribosomal protein S3: A constituent of translational machinery and an extraribosomal player in various cellular processes. Biochimie 2014, 99, 8–18. [Google Scholar] [CrossRef]
- Carvalho, C.M.; Santos, A.A.; Pires, S.R.; Rocha, C.S.; Saraiva, D.I.; Machado, J.P.; Mattos, E.C.; Fietto, L.G.; Fontes, E.P. Regulated nuclear trafficking of rpL10A mediated by NIK1 represents a defense strategy of plant cells against virus. PLoS Pathog. 2008, 4, e1000247. [Google Scholar] [CrossRef]
- Mazumder, B.; Poddar, D.; Basu, A.; Kour, R.; Verbovetskaya, V.; Barik, S. Extraribosomal l13a is a specific innate immune factor for antiviral defense. J. Virol. 2014, 88, 9100–9110. [Google Scholar] [CrossRef]
- Kuhn, J.F.; Tran, E.J.; Maxwell, E.S. Archaeal ribosomal protein L7 is a functional homolog of the eukaryotic 15.5kD/Snu13p snoRNP core protein. Nucleic Acids Res. 2002, 30, 931–941. [Google Scholar] [CrossRef]
- Warner, J.R.; McIntosh, K.B. How common are extraribosomal functions of ribosomal proteins? Mol. Cell 2009, 34, 3–11. [Google Scholar] [CrossRef]
- Lindstrom, M.S. Emerging functions of ribosomal proteins in gene-specific transcription and translation. Biochem. Biophys. Res. Commun. 2009, 379, 167–170. [Google Scholar] [CrossRef]
- Lu, H.; Zhu, Y.F.; Xiong, J.; Wang, R.; Jia, Z. Potential extra-ribosomal functions of ribosomal proteins in Saccharomyces cerevisiae. Microbiol. Res. 2015, 177, 28–33. [Google Scholar] [CrossRef]
- Wool, I.G. Extraribosomal functions of ribosomal proteins. Trends Biochem. Sci. 1996, 21, 164–165. [Google Scholar] [CrossRef]
- Mishra, P.C.; Kumar, A.; Sharma, A. Analysis of small nucleolar RNAs reveals unique genetic features in malaria parasites. BMC Genom. 2009, 10, 68. [Google Scholar] [CrossRef]
- Wang, L.; Ciganda, M.; Williams, N. Defining the RNA-protein interactions in the trypanosome preribosomal complex. Eukaryot. Cell 2013, 12, 559–566. [Google Scholar] [CrossRef]
- Liang, X.; Hart, K.J.; Dong, G.; Siddiqui, F.A.; Sebastian, A.; Li, X.; Albert, I.; Miao, J.; Lindner, S.E.; Cui, L. Puf3 participates in ribosomal biogenesis in malaria parasites. J. Cell Sci. 2018, 131, jcs212597. [Google Scholar] [CrossRef]
- Fricker, R.; Brogli, R.; Luidalepp, H.; Wyss, L.; Fasnacht, M.; Joss, O.; Zywicki, M.; Helm, M.; Schneider, A.; Cristodero, M.; et al. A tRNA half modulates translation as stress response in Trypanosoma brucei. Nat. Commun. 2019, 10, 118. [Google Scholar] [CrossRef] [PubMed]
- Rajan, K.S.; Chikne, V.; Decker, K.; Waldman Ben-Asher, H.; Michaeli, S. Unique Aspects of rRNA Biogenesis in Trypanosomatids. Trends Parasitol. 2019, 35, 778–794. [Google Scholar] [CrossRef] [PubMed]
- Nepomuceno-Mejia, T.; Florencio-Martinez, L.E.; Pineda-Garcia, I.; Martinez-Calvillo, S. Identification of factors involved in ribosome assembly in the protozoan parasite Leishmania major. Acta Trop. 2022, 228, 106315. [Google Scholar] [CrossRef] [PubMed]
- Pircher, A.; Gebetsberger, J.; Polacek, N. Ribosome-associated ncRNAs: An emerging class of translation regulators. RNA Biol. 2014, 11, 1335–1339. [Google Scholar] [CrossRef] [PubMed]
- Mleczko, A.M.; Machtel, P.; Walkowiak, M.; Wasilewska, A.; Pietras, P.J.; Bakowska-Zywicka, K. Levels of sdRNAs in cytoplasm and their association with ribosomes are dependent upon stress conditions but independent from snoRNA expression. Sci. Rep. 2019, 9, 18397. [Google Scholar] [CrossRef]
- Dumas, C.; Chow, C.; Muller, M.; Papadopoulou, B. A novel class of developmentally regulated noncoding RNAs in Leishmania. Eukaryot. Cell 2006, 5, 2033–2046. [Google Scholar] [CrossRef]
- Rajan, K.S.; Doniger, T.; Cohen-Chalamish, S.; Rengaraj, P.; Galili, B.; Aryal, S.; Unger, R.; Tschudi, C.; Michaeli, S. Developmentally Regulated Novel Non-coding Anti-sense Regulators of mRNA Translation in Trypanosoma brucei. iScience 2020, 23, 101780. [Google Scholar] [CrossRef]
- Guegan, F.; Rajan, K.S.; Bento, F.; Pinto-Neves, D.; Sequeira, M.; Guminska, N.; Mroczek, S.; Dziembowski, A.; Cohen-Chalamish, S.; Doniger, T.; et al. A long noncoding RNA promotes parasite differentiation in African trypanosomes. Sci. Adv. 2022, 8, eabn2706. [Google Scholar] [CrossRef]
- Leroux, L.P.; Lorent, J.; Graber, T.E.; Chaparro, V.; Masvidal, L.; Aguirre, M.; Fonseca, B.D.; van Kempen, L.C.; Alain, T.; Larsson, O.; et al. The Protozoan Parasite Toxoplasma gondii Selectively Reprograms the Host Cell Translatome. Infect. Immun. 2018, 86. [Google Scholar] [CrossRef]
- Vesper, O.; Amitai, S.; Belitsky, M.; Byrgazov, K.; Kaberdina, A.C.; Engelberg-Kulka, H.; Moll, I. Selective translation of leaderless mRNAs by specialized ribosomes generated by MazF in Escherichia coli. Cell 2011, 147, 147–157. [Google Scholar] [CrossRef]
- Kurylo, C.M.; Parks, M.M.; Juette, M.F.; Zinshteyn, B.; Altman, R.B.; Thibado, J.K.; Vincent, C.T.; Blanchard, S.C. Endogenous rRNA Sequence Variation Can Regulate Stress Response Gene Expression and Phenotype. Cell Rep. 2018, 25, 236–248.e236. [Google Scholar] [CrossRef]
- Lopez-Lopez, A.; Benlloch, S.; Bonfa, M.; Rodriguez-Valera, F.; Mira, A. Intragenomic 16S rDNA divergence in Haloarcula marismortui is an adaptation to different temperatures. J. Mol. Evol. 2007, 65, 687–696. [Google Scholar] [CrossRef]
- Chavez-Fumagalli, M.A.; Costa, M.A.; Oliveira, D.M.; Ramirez, L.; Costa, L.E.; Duarte, M.C.; Martins, V.T.; Oliveira, J.S.; Olortegi, C.C.; Bonay, P.; et al. Vaccination with the Leishmania infantum ribosomal proteins induces protection in BALB/c mice against Leishmania chagasi and Leishmania amazonensis challenge. Microbes Infect. 2010, 12, 967–977. [Google Scholar] [CrossRef]
- Ramirez, L.; Santos, D.M.; Souza, A.P.; Coelho, E.A.; Barral, A.; Alonso, C.; Escutia, M.R.; Bonay, P.; de Oliveira, C.I.; Soto, M. Evaluation of immune responses and analysis of the effect of vaccination of the Leishmania major recombinant ribosomal proteins L3 or L5 in two different murine models of cutaneous leishmaniasis. Vaccine 2013, 31, 1312–1319. [Google Scholar] [CrossRef]
- Prava, J.; Pan, A. In silico analysis of Leishmania proteomes and protein-protein interaction network: Prioritizing therapeutic targets and drugs for repurposing to treat leishmaniasis. Acta Trop. 2022, 229, 106337. [Google Scholar] [CrossRef]
- Clayton, C.E. Life without transcriptional control? From fly to man and back again. EMBO J. 2002, 21, 1881–1888. [Google Scholar] [CrossRef]
- Smircich, P.; Eastman, G.; Bispo, S.; Duhagon, M.A.; Guerra-Slompo, E.P.; Garat, B.; Goldenberg, S.; Munroe, D.J.; Dallagiovanna, B.; Holetz, F.; et al. Ribosome profiling reveals translation control as a key mechanism generating differential gene expression in Trypanosoma cruzi. BMC Genom. 2015, 16, 443. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
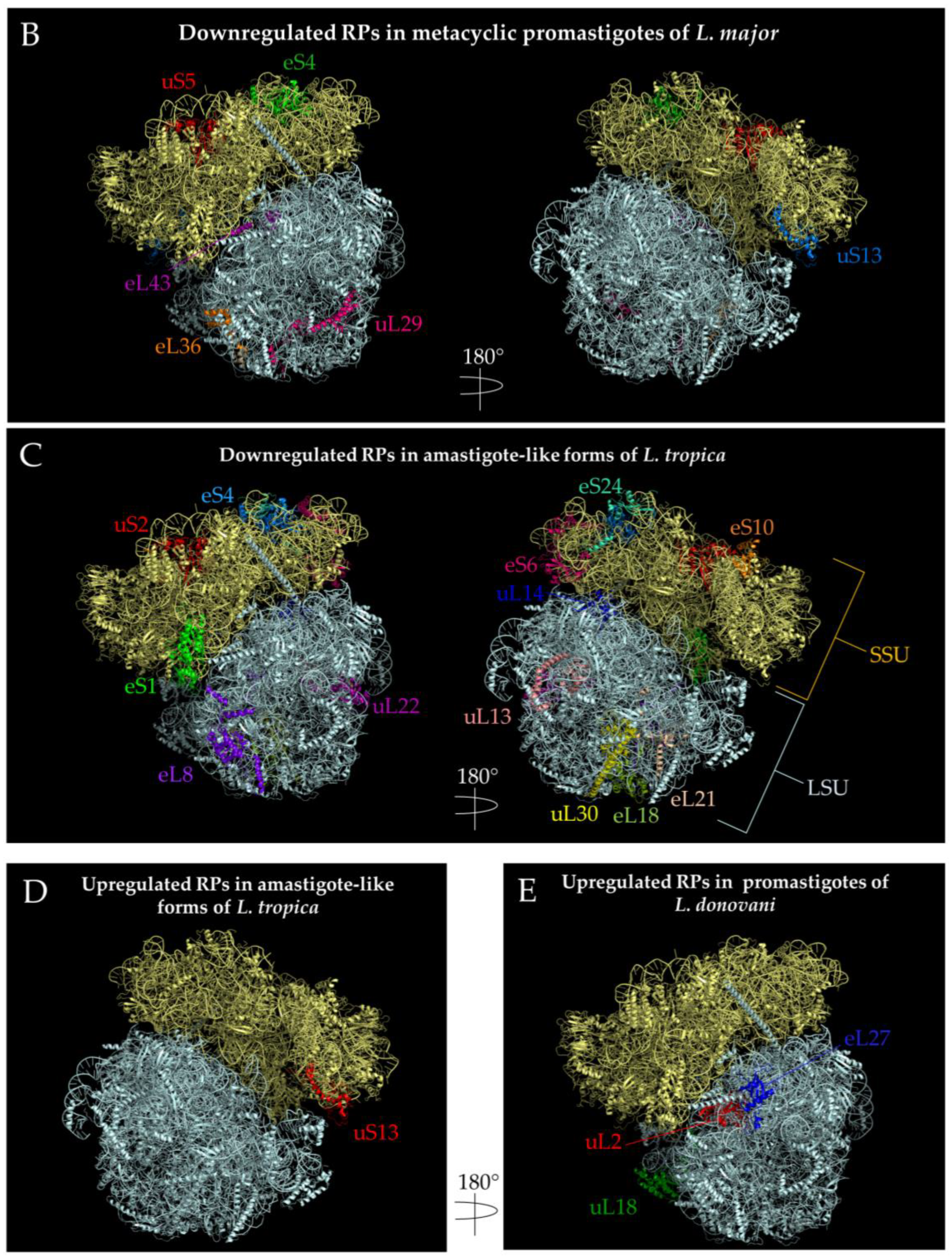{kind=link}
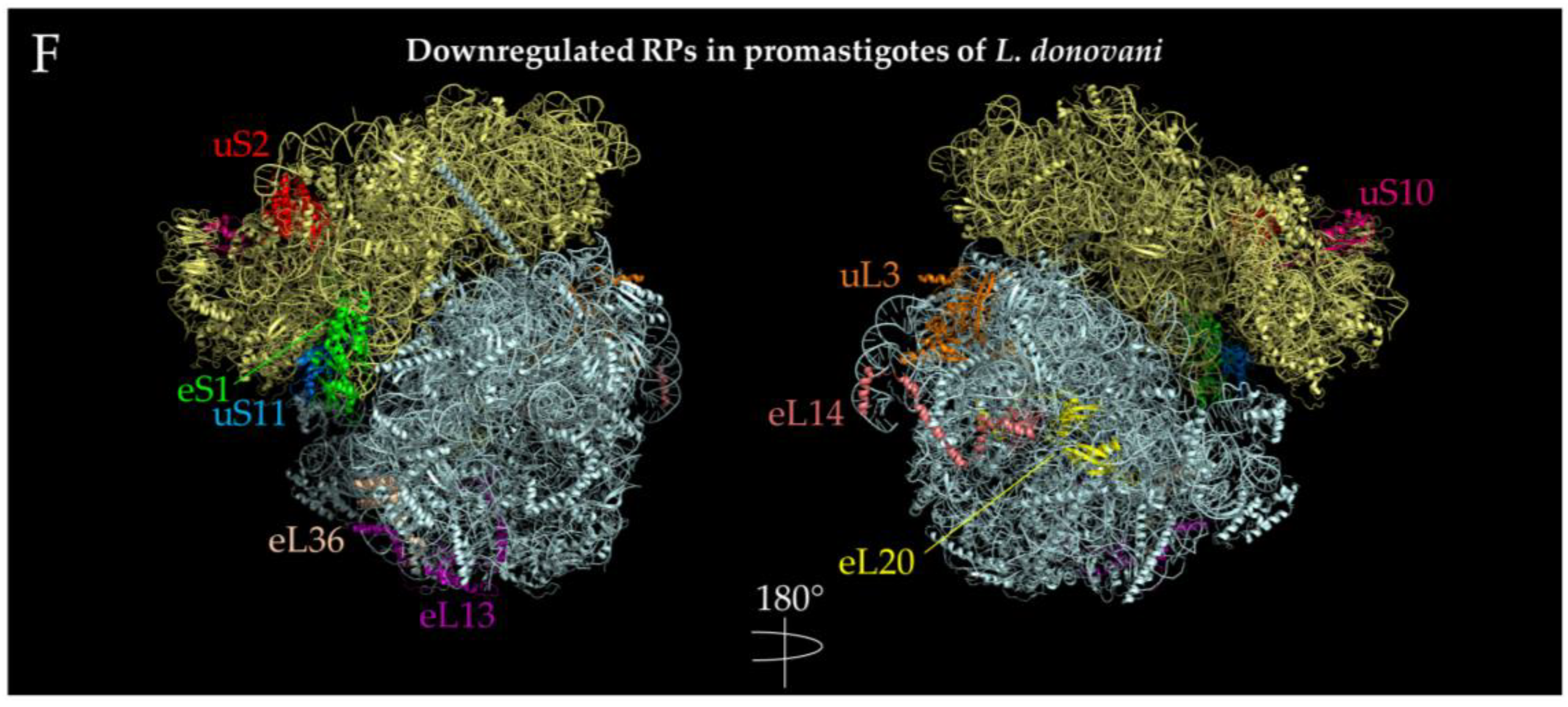{kind=link}
| E. coli Ribosome [61] | S. cerevisiae Mitoribosome [62] | T. brucei Mitoribosome [63] | H. sapiens Mitoribosome [64] | Eukaryotic Cytosol Ribosomes [61] | |
|---|---|---|---|---|---|
| Ribosome | |||||
| Sedimentation coefficient | 70S | 74S | ~60S [65] | 55S | 80S |
| Molecular weight | 2.3 MDa | 3–3.3 MDa | 4.5 MDa | 2.7 MDa | 3.3–4.3 MDa |
| Number of rRNAs | 3 | 2 | 2 | 3 | 4 |
| Number of proteins | 55 | 80 | 127 | 83 | 79–80 |
| Large subunit | |||||
| Sedimentation coefficient | 50S | 54S | 40S * | 39S | 60S |
| Number of rRNAs | 23S (2904 nt) | 21S (3296 nt) | 12S (1176 nt) | 16S (1569 nt) | 26S–28S (3396–5034 nt) |
| 5.8S (156–158 nt) | |||||
| 5S (120 nt) | tRNA (73–75 nt) | 5S (120–121 nt) | |||
| Number of proteins | 33 | 46 | 72 | 50 | 46–47 |
| Small subunit | |||||
| Sedimentation coefficient | 30S | 37S | 30S * | 28S | 40S |
| Number of rRNAs | 16S (1542 nt) | 15S (1649 nt) | 9S (620 nt) | 12S (962 nt) | 18S (1800–1870 nt) |
| Number of proteins | 22 | 34 | 55 | 30 | 33 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodríguez-Almonacid, C.C.; Kellogg, M.K.; Karamyshev, A.L.; Karamysheva, Z.N. Ribosome Specialization in Protozoa Parasites. Int. J. Mol. Sci. 2023, 24, 7484. https://doi.org/10.3390/ijms24087484
Rodríguez-Almonacid CC, Kellogg MK, Karamyshev AL, Karamysheva ZN. Ribosome Specialization in Protozoa Parasites. International Journal of Molecular Sciences. 2023; 24(8):7484. https://doi.org/10.3390/ijms24087484
Chicago/Turabian StyleRodríguez-Almonacid, Cristian Camilo, Morgana K. Kellogg, Andrey L. Karamyshev, and Zemfira N. Karamysheva. 2023. "Ribosome Specialization in Protozoa Parasites" International Journal of Molecular Sciences 24, no. 8: 7484. https://doi.org/10.3390/ijms24087484