
Protective Effects of a Jellyfish-Derived Thioredoxin Fused with Cell-Penetrating Peptide TAT-PTD on H2O2-Induced Oxidative Damage


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
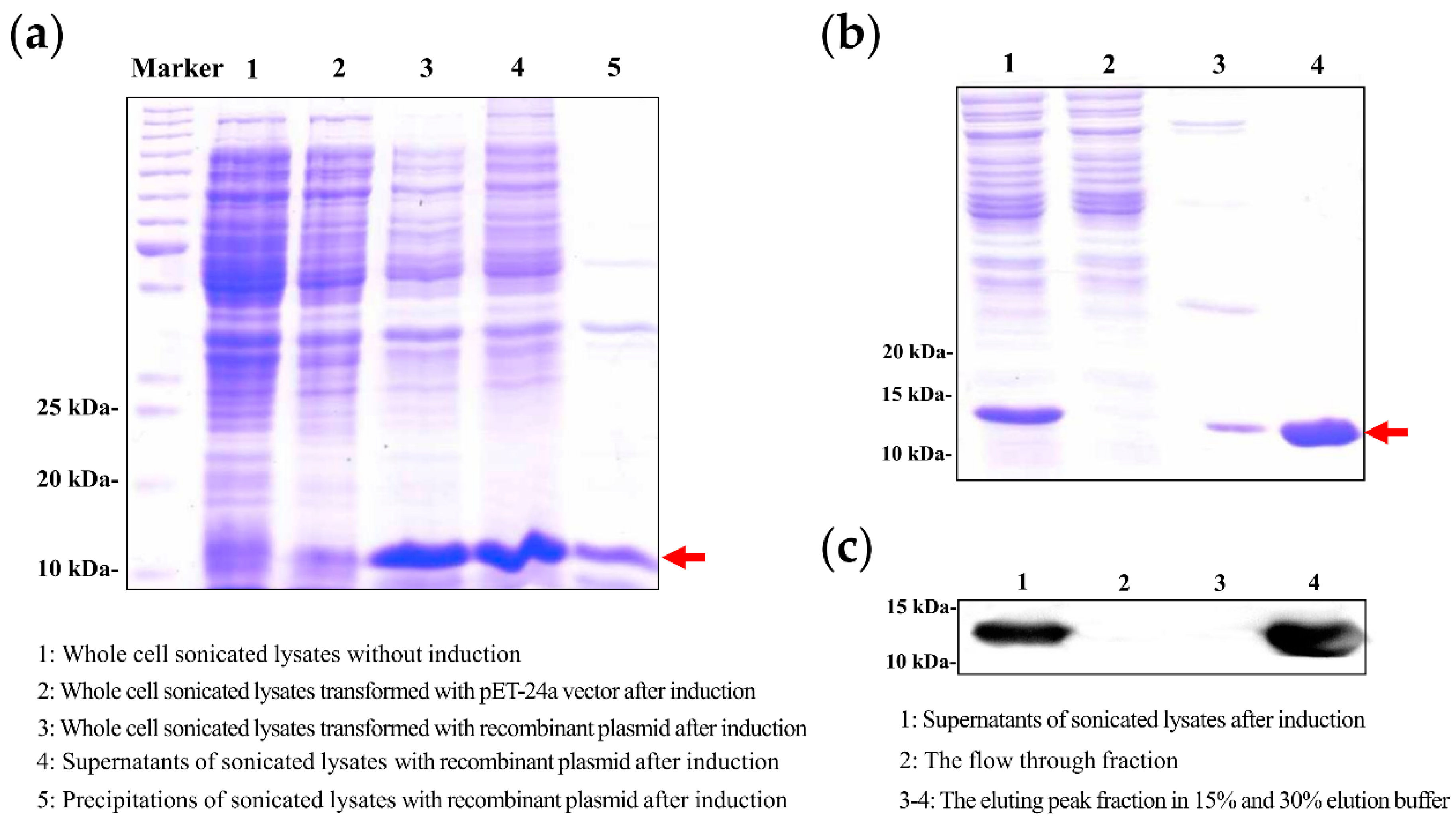
2.1. Characterization and Identification of PTD-CcTrx1 Coding Sequence
2.2. Expression and Purification of PTD-CcTrx1
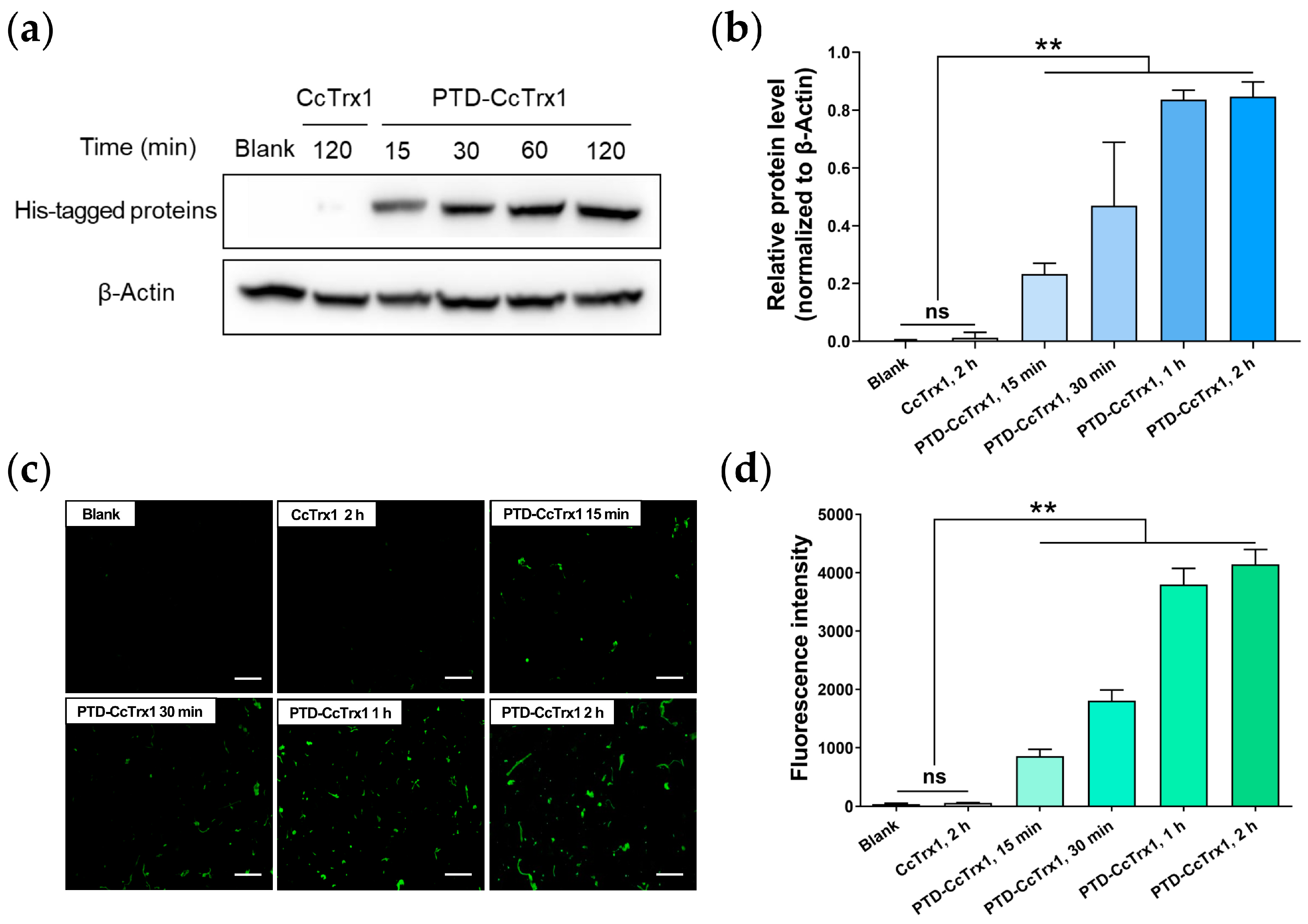
2.3. Transmembrane Delivery of PTD-CcTrx1 into HaCaT Cells
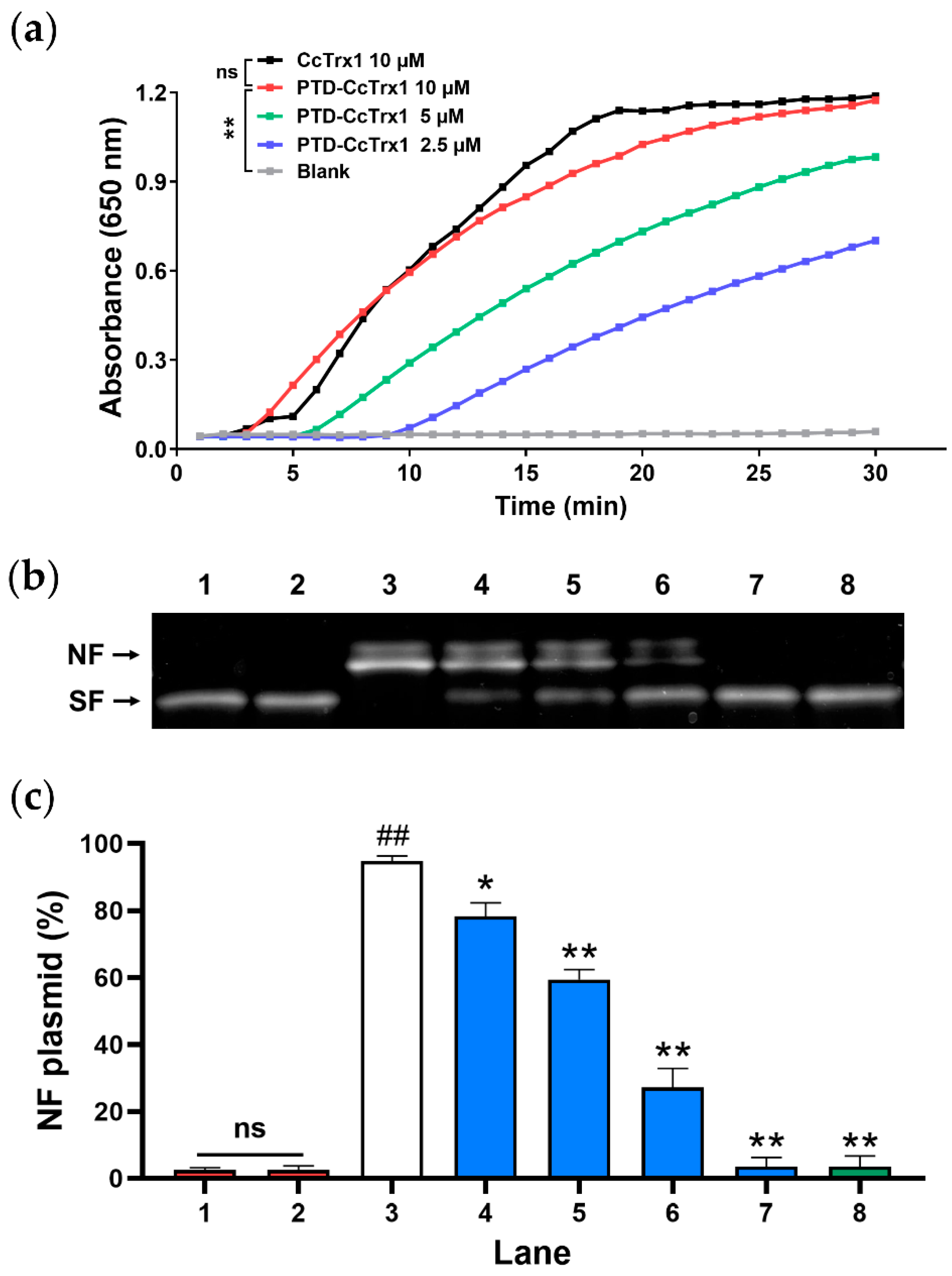
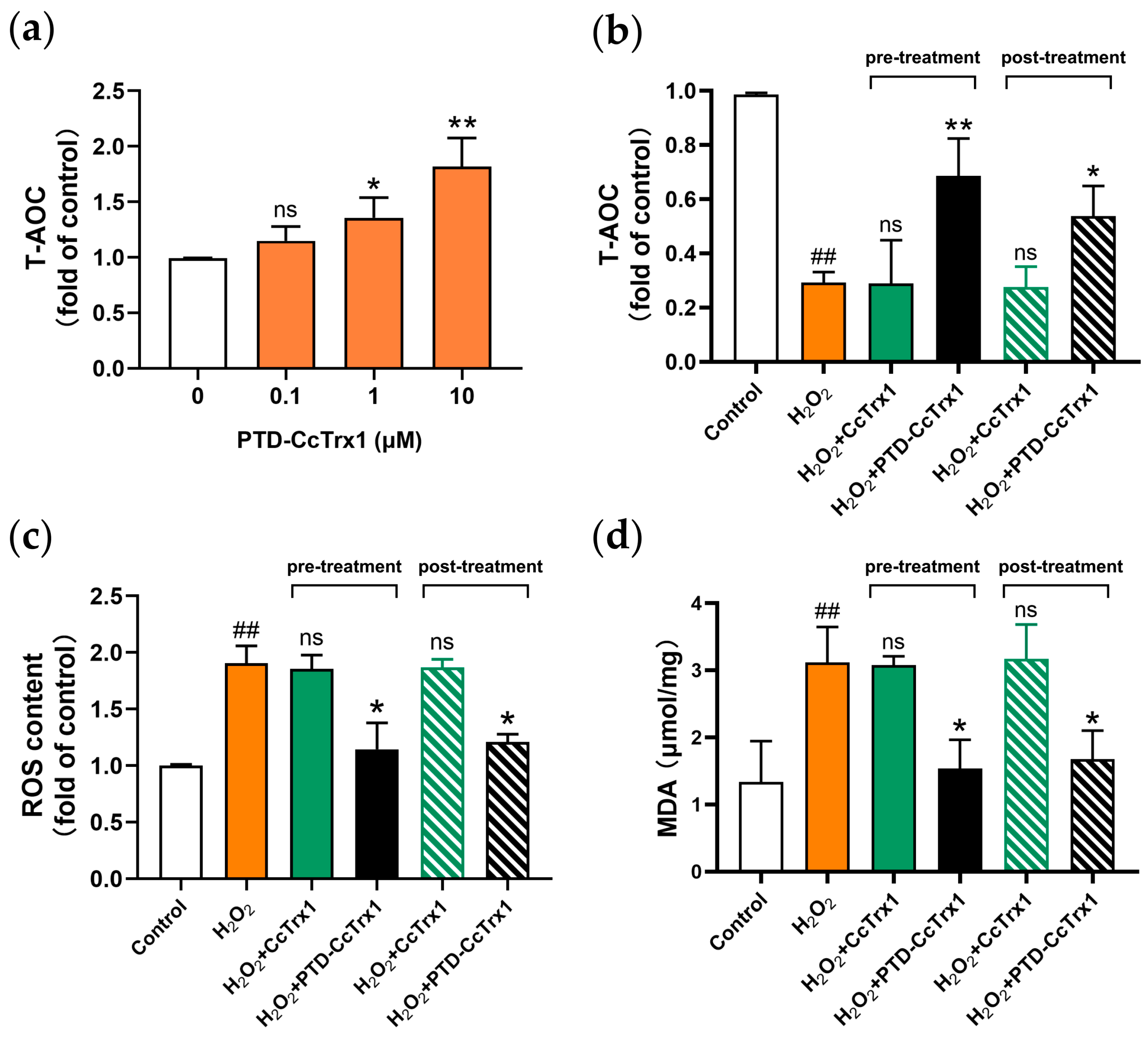
2.4. PTD-CcTrx1 Exhibited Specific Antioxidant Activities in Cell-Free System
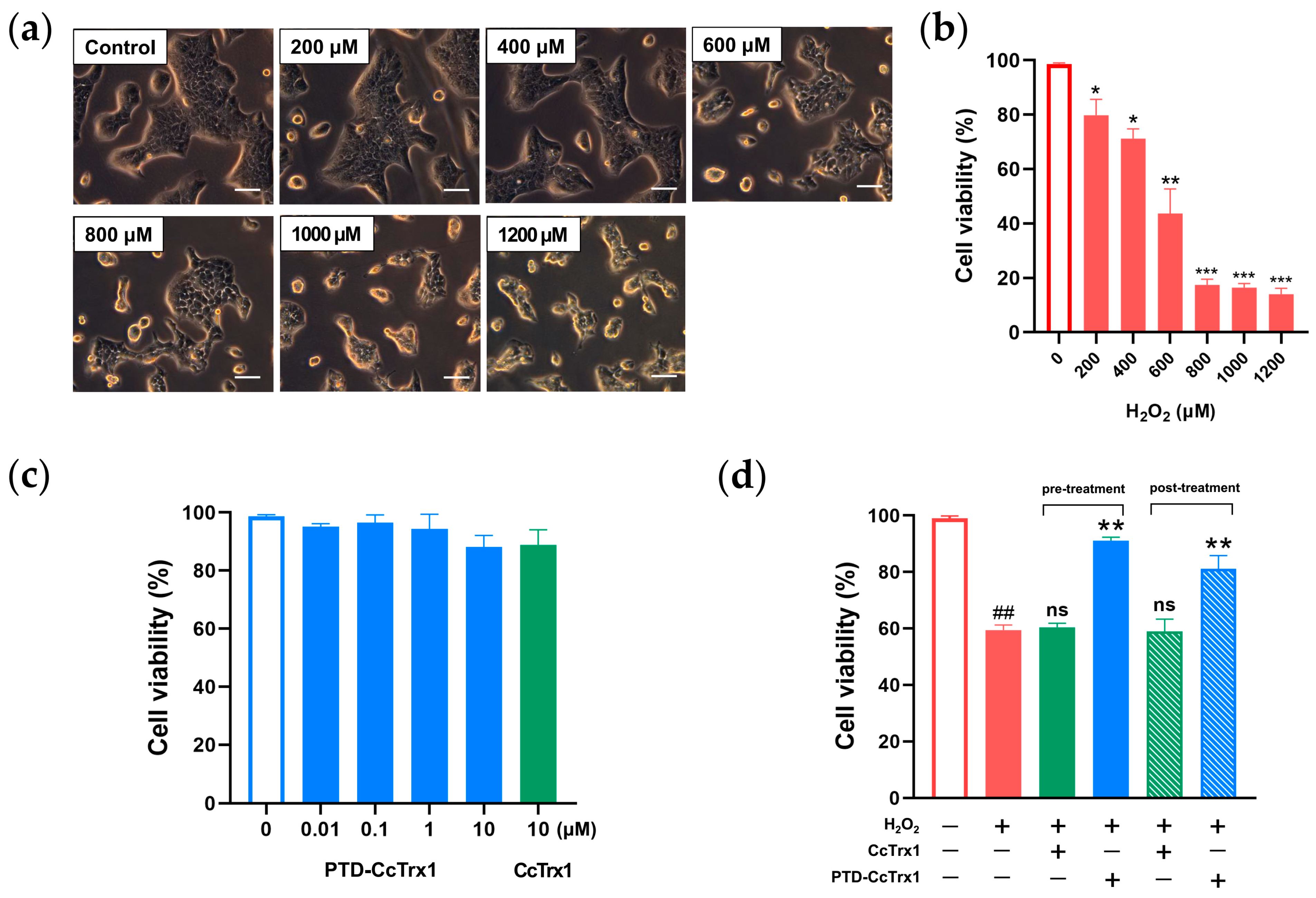
2.5. PTD-CcTrx1 Antagonized H2O2-Induced Cytotoxicity
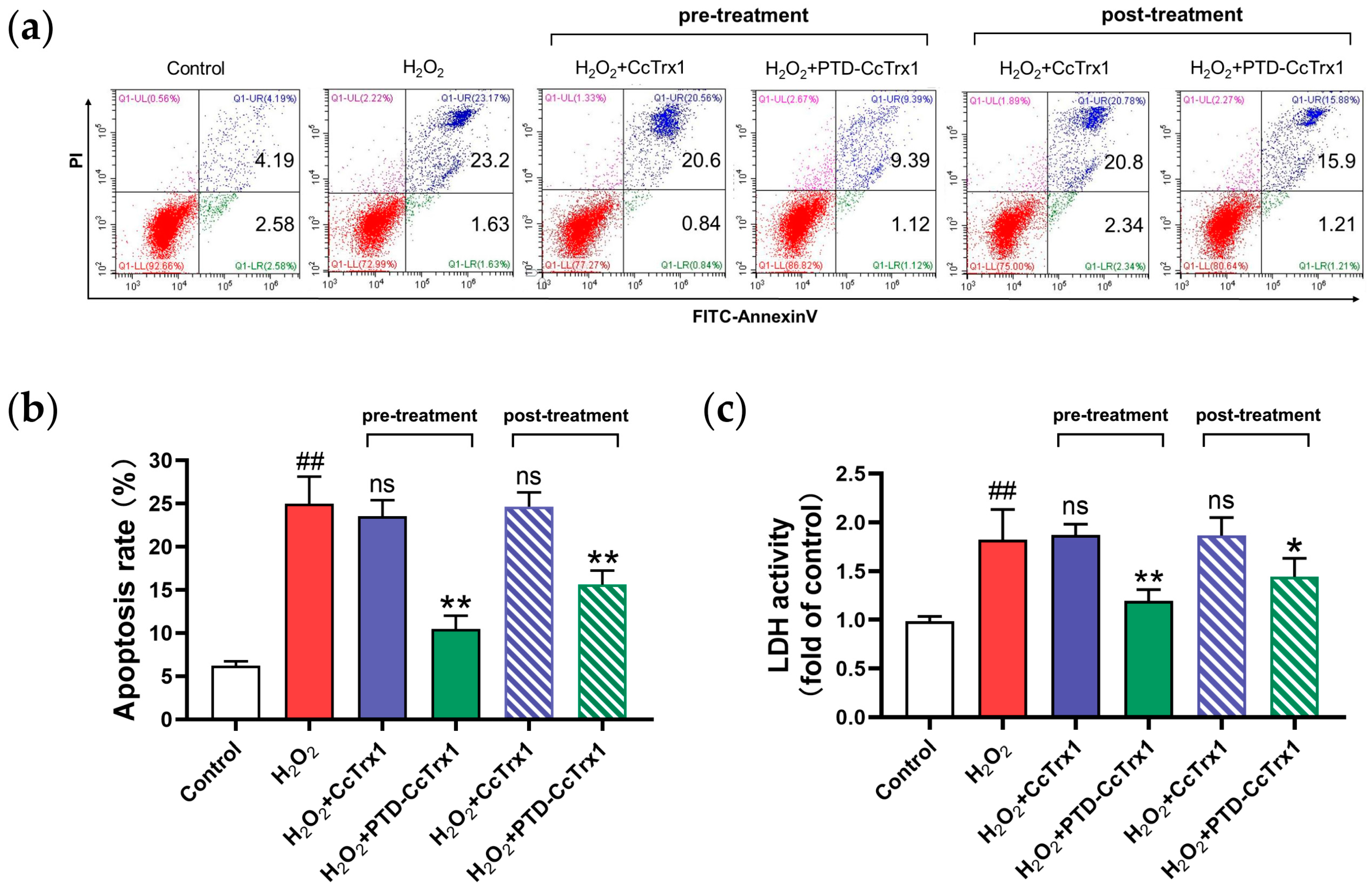
2.6. PTD-CcTrx1 Protected HaCaT Cells from H2O2-Induced Apoptosis
2.7. PTD-CcTrx1 Inhibited H2O2-Mediated Oxidative Stress in HaCaT Cells
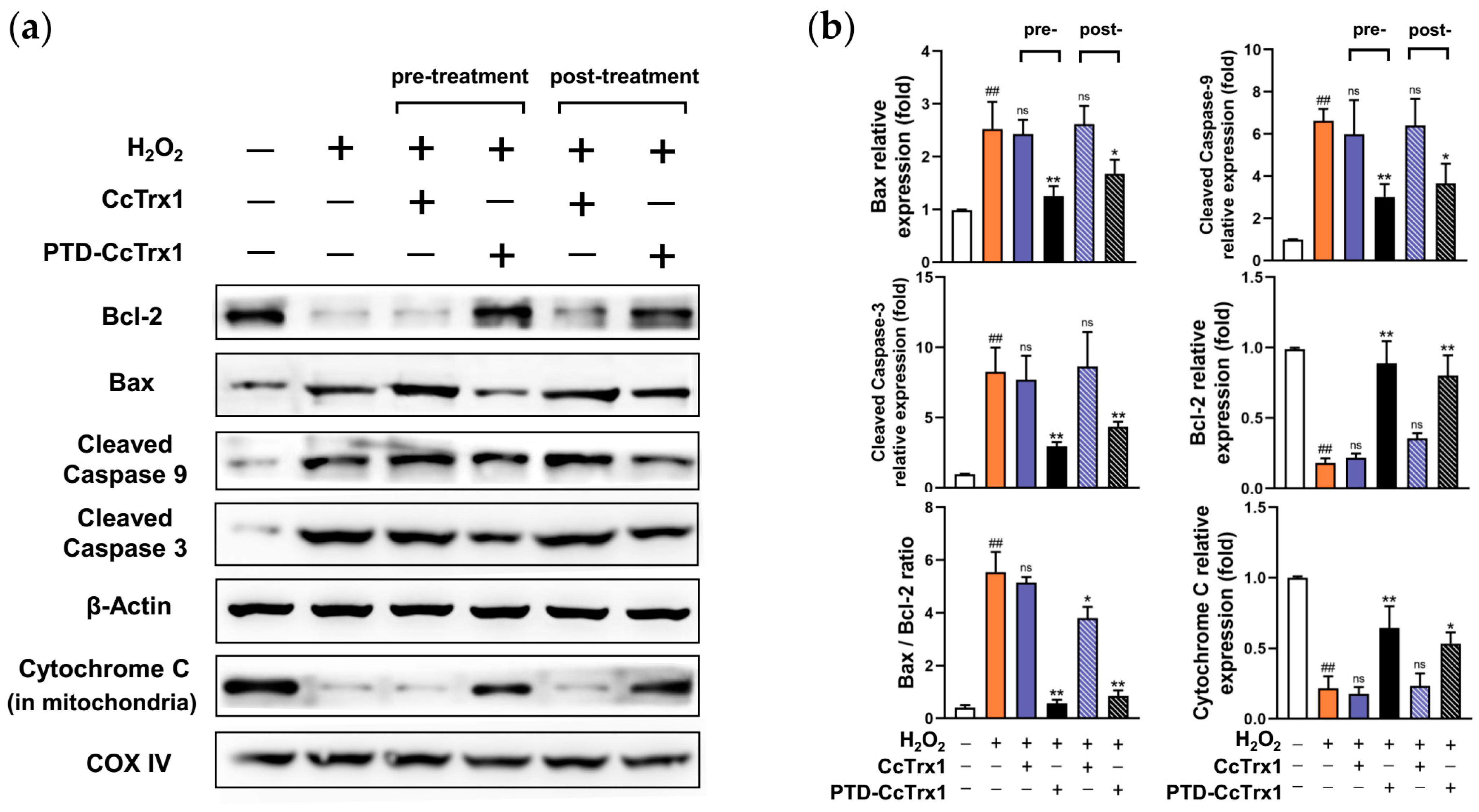
2.8. PTD-CcTrx1 Inhibited the Activation of Apoptosis Signaling Pathways Induced by H2O2
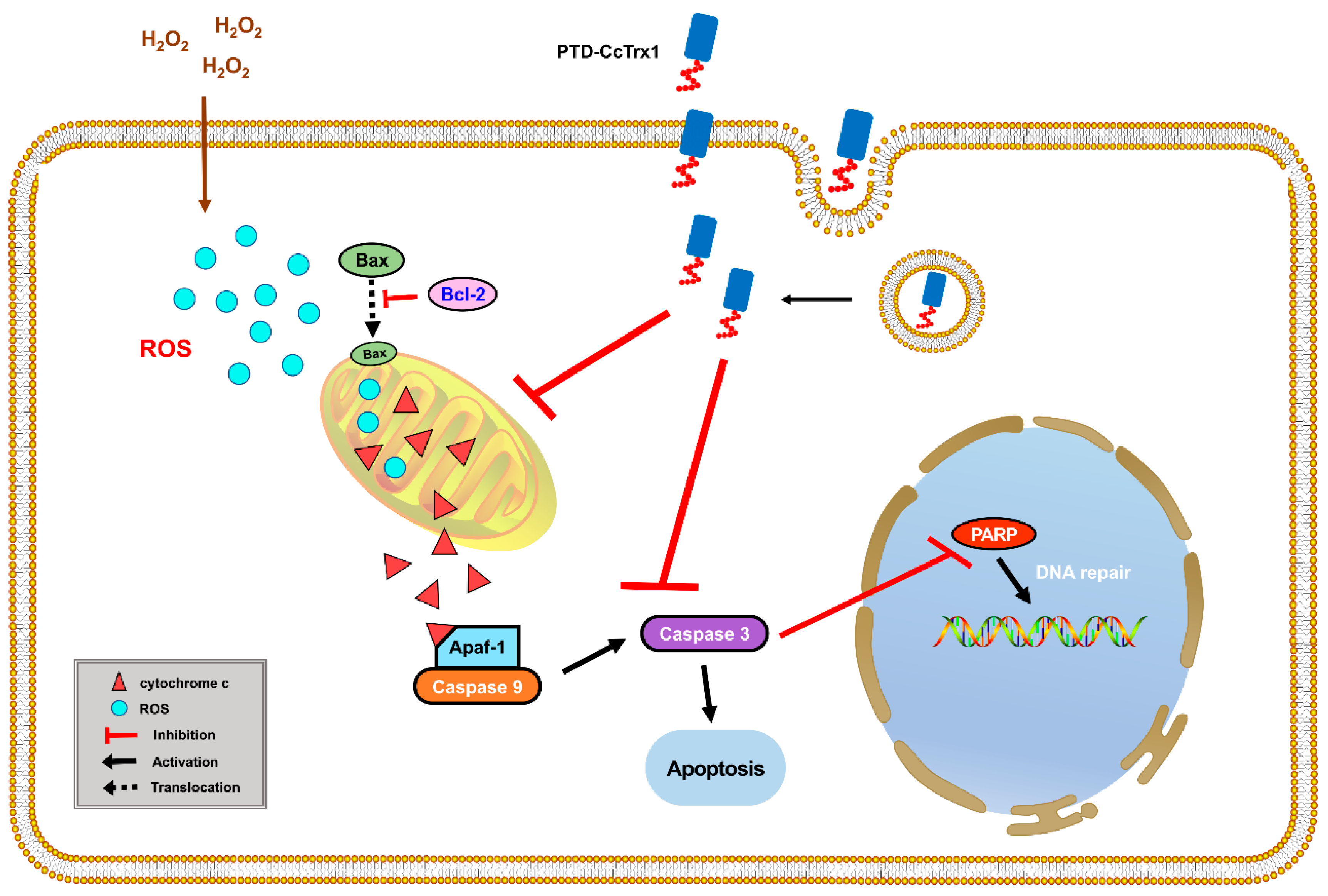
3. Discussion
4. Materials and Methods
4.1. Molecular Cloning
4.2. Expression and Purification of PTD-CcTrx1
4.3. Immunoblotting
4.4. Transmembrane Ability of PTD-CcTrx1
4.4.1. Western Blot Assay
4.4.2. Immunofluorescence Assay
4.5. Antioxidant Capacity Assays in Cell-Free System
4.5.1. Insulin Disulfide Reduction Assay by PTD-CcTrx1
4.5.2. DNA Cleavage Assay in a Metal-Catalyzed Oxidation System
4.6. Protective Effect of PTD-CcTrx1 against H2O2-Induced Cytotoxicity
4.7. Protective Effect of PTD-CcTrx1 on H2O2-Induced Apoptosis
4.7.1. Cell Apoptosis Assay
4.7.2. LDH Activity Assay
4.8. Total Antioxidant Capacity, ROS Level and Lipid Peroxidation Assays
4.9. Effects of PTD-CcTrx1 on Apoptosis Signaling Pathways Activated by H2O2 in HaCaT Cells
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mohania, D.; Chandel, S.; Kumar, P.; Verma, V.; Digvijay, K.; Tripathi, D.; Choudhury, K.; Mitten, S.K.; Shah, D. Ultraviolet radiations: Skin defense-damage mechanism. Adv. Exp. Med. Biol. 2017, 996, 71–87. [Google Scholar] [PubMed]
- D’Orazio, J.; Jarrett, S.; Amaro-Ortiz, A.; Scott, T. UV radiation and the skin. Int. J. Mol. Sci. 2013, 14, 12222–12248. [Google Scholar] [CrossRef] [PubMed]
- Ahn, K. The role of air pollutants in atopic dermatitis. J. Allergy Clin. Immunol. 2014, 134, 993–999. [Google Scholar] [CrossRef] [PubMed]
- Celebi Sözener, Z.; Cevhertas, L.; Nadeau, K.; Akdis, M.; Akdis, C.A. Environmental factors in epithelial barrier dysfunction. J. Allergy Clin. Immunol. 2020, 145, 1517–1528. [Google Scholar] [CrossRef] [PubMed]
- Gu, Y.; Han, J.; Jiang, C.; Zhang, Y. Biomarkers, oxidative stress and autophagy in skin aging. Ageing Res. Rev. 2020, 59, 101036. [Google Scholar] [CrossRef] [PubMed]
- Ansary, T.M.; Hossain, M.R.; Kamiya, K.; Komine, M.; Ohtsuki, M. Inflammatory Molecules Associated with Ultraviolet Radiation-Mediated Skin Aging. Int. J. Mol. Sci. 2021, 22, 3974. [Google Scholar] [CrossRef]
- Kammeyer, A.; Luiten, R.M. Oxidation events and skin aging. Ageing Res. Rev. 2015, 21, 16–29. [Google Scholar] [CrossRef]
- Romiszewska, A.; Nowak-Stępniowska, A. Photodynamic reaction and oxidative stress-influence of the photodynamic effect on the activity antioxidant enzymes. Postepy Biochem. 2014, 60, 355–364. [Google Scholar]
- Weydert, C.J.; Cullen, J.J. Measurement of superoxide dismutase, catalase and glutathione peroxidase in cultured cells and tissue. Nat. Protoc. 2010, 5, 51–66. [Google Scholar] [CrossRef]
- Ghareeb, H.; Metanis, N. The Thioredoxin System: A Promising Target for Cancer Drug Development. Chemistry 2020, 26, 10175–10184. [Google Scholar] [CrossRef]
- Mata-Pérez, C.; Spoel, S.H. Thioredoxin-mediated redox signalling in plant immunity. Plant. Sci. 2019, 279, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Holmgren, A. The thioredoxin antioxidant system. Free. Radic. Biol. Med. 2014, 66, 75–87. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Holmgren, A. Thioredoxin system in cell death progression. Antioxid. Redox Signal. 2012, 17, 1738–1747. [Google Scholar] [CrossRef] [PubMed]
- Myers, C.R.; Myers, J.M. The effects of acrolein on peroxiredoxins, thioredoxins, and thioredoxin reductase in human bronchial epithelial cells. Toxicology 2009, 257, 95–104. [Google Scholar] [CrossRef] [PubMed]
- Stancill, J.S.; Corbett, J.A. The Role of Thioredoxin/Peroxiredoxin in the β-Cell Defense Against Oxidative Damage. Front. Endocrinol. (Lausanne) 2021, 12, 718235. [Google Scholar] [CrossRef]
- Ruan, Z.; Liu, G.; Wang, B.; Zhou, Y.; Lu, J.; Wang, Q.; Zhao, J.; Zhang, L. First report of a peroxiredoxin homologue in jellyfish: Molecular cloning, expression and functional characterization of CcPrx4 from Cyanea capillata. Mar. Drugs 2014, 12, 214–231. [Google Scholar] [CrossRef]
- Stabili, L.; Rizzo, L.; Caprioli, R.; Leone, A.; Piraino, S. Jellyfish Bioprospecting in the Mediterranean Sea: Antioxidant and Lysozyme-Like Activities from Aurelia coerulea (Cnidaria, Scyphozoa) Extracts. Mar. Drugs 2021, 19, 619. [Google Scholar] [CrossRef]
- Amreen Nisa, S.; Vinu, D.; Krupakar, P.; Govindaraju, K.; Sharma, D.; Vivek, R. Jellyfish venom proteins and their pharmacological potentials: A review. Int. J. Biol. Macromol. 2021, 176, 424–436. [Google Scholar] [CrossRef]
- De Domenico, S.; De Rinaldis, G.; Paulmery, M.; Piraino, S.; Leone, A. Barrel Jellyfish (Rhizostoma pulmo) as Source of Antioxidant Peptides. Mar. Drugs 2019, 17, 134. [Google Scholar] [CrossRef]
- Ruan, Z.; Liu, G.; Guo, Y.; Zhou, Y.; Wang, Q.; Chang, Y.; Wang, B.; Zheng, J.; Zhang, L. First report of a thioredoxin homologue in jellyfish: Molecular cloning, expression and antioxidant activity of CcTrx1 from Cyanea capillata. PLoS ONE 2014, 9, e97509. [Google Scholar] [CrossRef]
- Schneider, A.F.L.; Kithil, M.; Cardoso, M.C.; Lehmann, M.; Hackenberger, C.P.R. Cellular uptake of large biomolecules enabled by cell-surface-reactive cell-penetrating peptide additives. Nat. Chem. 2021, 13, 530–539. [Google Scholar] [CrossRef] [PubMed]
- Raucher, D.; Ryu, J.S. Cell-penetrating peptides: Strategies for anticancer treatment. Trends Mol. Med. 2015, 21, 560–570. [Google Scholar] [CrossRef] [PubMed]
- Ramsey, J.D.; Flynn, N.H. Cell-penetrating peptides transport therapeutics into cells. Pharmacol. Ther. 2015, 154, 78–86. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhang, X.; Wang, F. Intracellular transduction and potential of Tat PTD and its analogs: From basic drug delivery mechanism to application. Expert. Opin. Drug. Deliv. 2012, 9, 457–472. [Google Scholar] [CrossRef] [PubMed]
- Yoon, J.S.; Jung, Y.T.; Hong, S.K.; Kim, S.H.; Shin, M.C.; Lee, D.G.; Shin, W.S.; Min, W.S.; Paik, S.Y. Characteristics of HIV-Tat protein transduction domain. J. Microbiol. 2004, 42, 328–335. [Google Scholar]
- Rizzuti, M.; Nizzardo, M.; Zanetta, C.; Ramirez, A.; Corti, S. Therapeutic applications of the cell-penetrating HIV-1 Tat peptide. Drug. Discov. Today 2015, 20, 76–85. [Google Scholar] [CrossRef]
- Frankel, A.D.; Pabo, C.O. Cellular uptake of the tat protein from human immunodeficiency virus. Cell. 1988, 55, 1189–1193. [Google Scholar] [CrossRef]
- Apel, K.; Hirt, H. Reactive oxygen species: Metabolism, oxidative stress, and signal transduction. Annu. Rev. Plant. Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef]
- Zorov, D.B.; Juhaszova, M.; Sollott, S.J. Mitochondrial reactive oxygen species (ROS) and ROS-induced ROS release. Physiol. Rev. 2014, 94, 909–950. [Google Scholar] [CrossRef]
- Cross, C.E.; van der Vliet, A.; Louie, S.; Thiele, J.J.; Halliwell, B. Oxidative stress and antioxidants at biosurfaces: Plants, skin, and respiratory tract surfaces. Environ. Health Perspect. 1998, 106, 1241–1251. [Google Scholar]
- Zhang, J.; Li, X.; Han, X.; Liu, R.; Fang, J. Targeting the Thioredoxin System for Cancer Therapy. Trends Pharmacol. Sci. 2017, 38, 794–808. [Google Scholar] [CrossRef] [PubMed]
- Sevilla, F.; Camejo, D.; Ortiz-Espín, A.; Calderón, A.; Lázaro, J.J.; Jiménez, A. The thioredoxin/peroxiredoxin/sulfiredoxin system: Current overview on its redox function in plants and regulation by reactive oxygen and nitrogen species. J. Exp. Bot. 2015, 66, 2945–2955. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Zhu, M.; Zhu, N.; Strul, J.M.; Dufresne, C.P.; Schneider, J.D.; Harmon, A.C.; Chen, S. Identification of thioredoxin targets in guard cell enriched epidermal peels using cysTMT proteomics. J. Proteomics 2016, 133, 48–53. [Google Scholar] [CrossRef]
- Miranda-Vizuete, A.; Ljung, J.; Damdimopoulos, A.E.; Gustafsson, J.A.; Oko, R.; Pelto-Huikko, M.; Spyrou, G. Characterization of Sptrx, a novel member of the thioredoxin family specifically expressed in human spermatozoa. J. Biol. Chem. 2001, 276, 31567–31574. [Google Scholar] [CrossRef] [PubMed]
- Tedetti, M.; Sempéré, R. Penetration of ultraviolet radiation in the marine environment. A review. Photochem. Photobiol. 2006, 82, 389–397. [Google Scholar] [CrossRef] [PubMed]
- Helbling, E.W.; Barbieri, E.S.; Marcoval, M.A.; Gonçalves, R.J.; Villafañe, V.E. Impact of solar ultraviolet radiation on marine phytoplankton of Patagonia, Argentina. Photochem. Photobiol. 2005, 81, 807–818. [Google Scholar] [CrossRef] [PubMed]
- Scaletti, F.; Hardie, J.; Lee, Y.W.; Luther, D.C.; Ray, M.; Rotello, V.M. Protein delivery into cells using inorganic nanoparticle-protein supramolecular assemblies. Chem. Soc. Rev. 2018, 47, 3421–3432. [Google Scholar] [CrossRef]
- Kim, J.; Narayana, A.; Patel, S.; Sahay, G. Advances in intracellular delivery through supramolecular self-assembly of oligonucleotides and peptides. Theranostics 2019, 9, 3191–3212. [Google Scholar] [CrossRef]
- McClements, D.J. Encapsulation, protection, and delivery of bioactive proteins and peptides using nanoparticle and microparticle systems: A review. Adv. Colloid. Interface Sci. 2018, 253, 1–22. [Google Scholar] [CrossRef]
- Lee, Y.W.; Luther, D.C.; Kretzmann, J.A.; Burden, A.; Jeon, T.; Zhai, S.; Rotello, V.M. Protein Delivery into the Cell Cytosol using Non-Viral Nanocarriers. Theranostics 2019, 9, 3280–3292. [Google Scholar] [CrossRef]
- Tian, S.; Liu, Y.; Appleton, E.; Wang, H.; Church, G.M.; Dong, M. Targeted intracellular delivery of Cas13 and Cas9 nucleases using bacterial toxin-based platforms. Cell. Rep. 2022, 38, 110476. [Google Scholar] [CrossRef]
- He, X.; Yang, J.; Ji, M.; Chen, Y.; Chen, Y.; Li, H.; Wang, H. A potential delivery system based on cholera toxin: A macromolecule carrier with multiple activities. J. Control. Release 2022, 343, 551–563. [Google Scholar] [CrossRef] [PubMed]
- Ruschig, M.; Marschall, A.L.J. Targeting the Inside of Cells with Biologicals: Toxin Routes in a Therapeutic Context. BioDrugs 2023, 37, 181–203. [Google Scholar] [CrossRef]
- Bolhassani, A.; Jafarzade, B.S.; Mardani, G. In vitro and in vivo delivery of therapeutic proteins using cell penetrating peptides. Peptides 2017, 87, 50–63. [Google Scholar] [CrossRef]
- Feni, L.; Neundorf, I. The Current Role of Cell-Penetrating Peptides in Cancer Therapy. Adv. Exp. Med. Biol. 2017, 1030, 279–295. [Google Scholar]
- Ma, J.; Xu, J.; Guan, L.; Hu, T.; Liu, Q.; Xiao, J.; Zhang, Y. Cell-penetrating peptides mediated protein cross-membrane delivery and its use in bacterial vector vaccine. Fish. Shellfish. Immunol. 2014, 39, 8–16. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Hu, Y.H. Cell-penetrating peptide-mediated subunit vaccine generates a potent immune response and protection against Streptococcus iniae in Japanese flounder (Paralichthys olivaceus). Vet. Immunol. Immunopathol. 2015, 167, 96–103. [Google Scholar] [CrossRef]
- Zhu, C.; Dong, Y.; Liu, H.; Ren, H.; Cui, Z. Hesperetin protects against H(2)O(2)-triggered oxidative damage via upregulation of the Keap1-Nrf2/HO-1 signal pathway in ARPE-19 cells. Biomed. Pharmacother. 2017, 88, 124–133. [Google Scholar] [CrossRef]
- Wang, T.; Jian, Z.; Baskys, A.; Yang, J.; Li, J.; Guo, H.; Hei, Y.; Xian, P.; He, Z.; Li, Z.; et al. MSC-derived exosomes protect against oxidative stress-induced skin injury via adaptive regulation of the NRF2 defense system. Biomaterials 2020, 257, 120264. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Pei, R.; Zhang, Z.; Liao, J.; Yu, W.; Qiao, N.; Han, Q.; Li, Y.; Hu, L.; Guo, J.; et al. Copper induces oxidative stress and apoptosis through mitochondria-mediated pathway in chicken hepatocytes. Toxicol. In Vitro 2019, 54, 310–316. [Google Scholar] [CrossRef]
- Tusi, S.K.; Khalaj, L.; Ashabi, G.; Kiaei, M.; Khodagholi, F. Alginate oligosaccharide protects against endoplasmic reticulum- and mitochondrial-mediated apoptotic cell death and oxidative stress. Biomaterials 2011, 32, 5438–5458. [Google Scholar] [CrossRef]
- Zhou, J.; Li, X.Y.; Liu, Y.J.; Feng, J.; Wu, Y.; Shen, H.M.; Lu, G.D. Full-coverage regulations of autophagy by ROS: From induction to maturation. Autophagy 2022, 18, 1240–1255. [Google Scholar] [CrossRef] [PubMed]
- Holmgren, A. Thioredoxin catalyzes the reduction of insulin disulfides by dithiothreitol and dihydrolipoamide. J. Biol. Chem. 1979, 254, 9627–9632. [Google Scholar] [CrossRef] [PubMed]
- Yao, P.; Hao, L.; Wang, F.; Chen, X.; Yan, Y.; Guo, X.; Xu, B. Molecular cloning, expression and antioxidant characterisation of a typical thioredoxin gene (AccTrx2) in Apis cerana cerana. Gene 2013, 527, 33–41. [Google Scholar] [CrossRef]
- Li, J.; Zhang, W.B.; Loukas, A.; Lin, R.Y.; Ito, A.; Zhang, L.H.; Jones, M.; McManus, D.P. Functional expression and characterization of Echinococcus granulosus thioredoxin peroxidase suggests a role in protection against oxidative damage. Gene 2004, 326, 157–165. [Google Scholar] [CrossRef]
- Qian, J.; Jiang, F.; Wang, B.; Yu, Y.; Zhang, X.; Yin, Z.; Liu, C. Ophiopogonin D prevents H2O2-induced injury in primary human umbilical vein endothelial cells. J. Ethnopharmacol. 2010, 128, 438–445. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.N.; Xu, H.H.; Zhou, W.; Yang, H.X.; Wang, J.; Ma, Z.C.; Gao, Y. Aconitine attenuates mitochondrial dysfunction of cardiomyocytes via promoting deacetylation of cyclophilin-D mediated by sirtuin-3. J. Ethnopharmacol. 2021, 270, 113765. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, B.; Zhang, P.; Wang, Q.; Zou, S.; Song, J.; Zhang, F.; Liu, G.; Zhang, L. Protective Effects of a Jellyfish-Derived Thioredoxin Fused with Cell-Penetrating Peptide TAT-PTD on H2O2-Induced Oxidative Damage. Int. J. Mol. Sci. 2023, 24, 7340. https://doi.org/10.3390/ijms24087340
Wang B, Zhang P, Wang Q, Zou S, Song J, Zhang F, Liu G, Zhang L. Protective Effects of a Jellyfish-Derived Thioredoxin Fused with Cell-Penetrating Peptide TAT-PTD on H2O2-Induced Oxidative Damage. International Journal of Molecular Sciences. 2023; 24(8):7340. https://doi.org/10.3390/ijms24087340
Chicago/Turabian StyleWang, Bo, Peipei Zhang, Qianqian Wang, Shuaijun Zou, Juxingsi Song, Fuhai Zhang, Guoyan Liu, and Liming Zhang. 2023. "Protective Effects of a Jellyfish-Derived Thioredoxin Fused with Cell-Penetrating Peptide TAT-PTD on H2O2-Induced Oxidative Damage" International Journal of Molecular Sciences 24, no. 8: 7340. https://doi.org/10.3390/ijms24087340