
Bioactive Constituents of Verbena officinalis Alleviate Inflammation and Enhance Killing Efficiency of Natural Killer Cells

 , , and
, , and 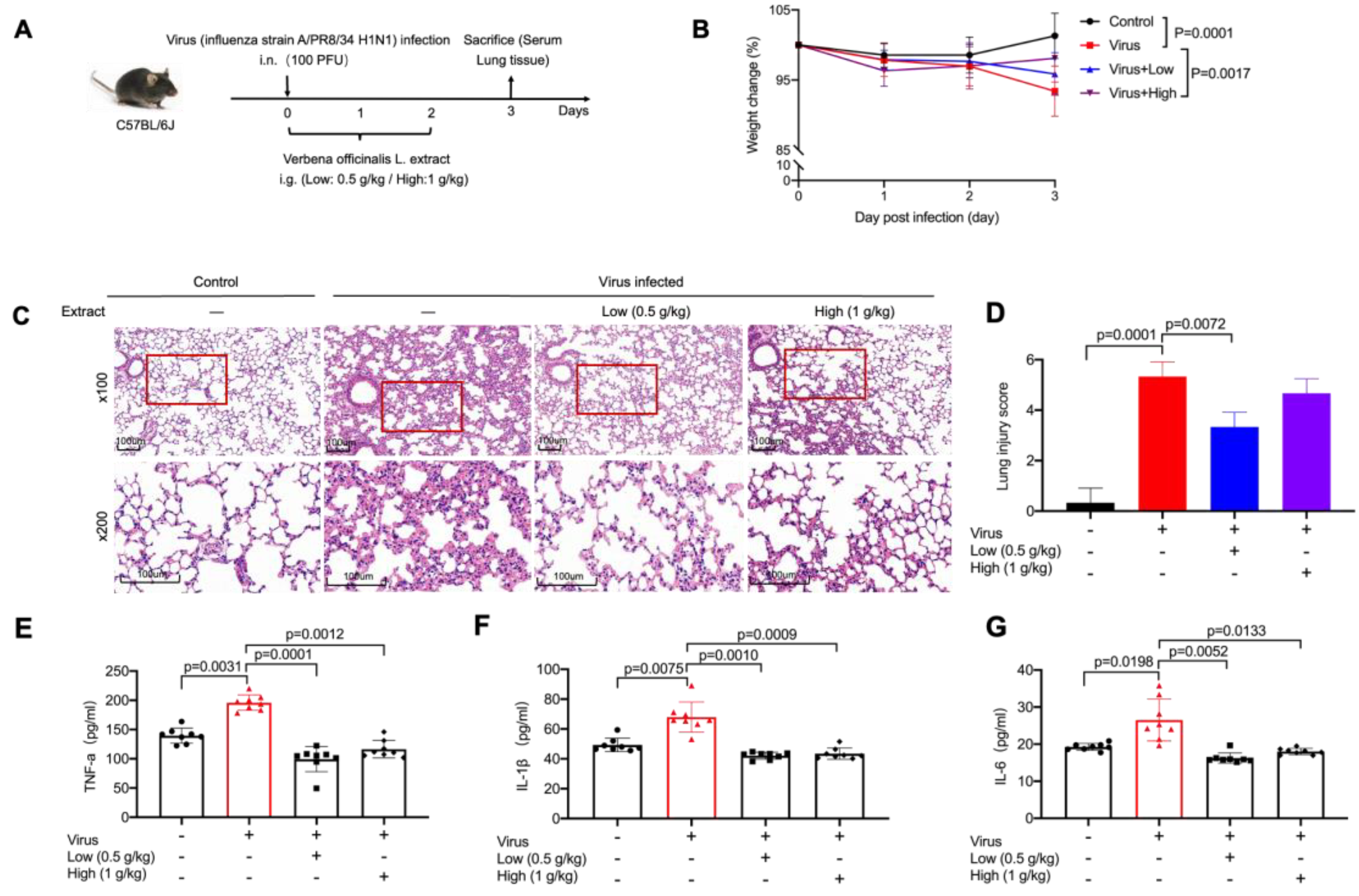{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. VO Extract Attenuated the Acute Lung Damage Induced by Virus Infection
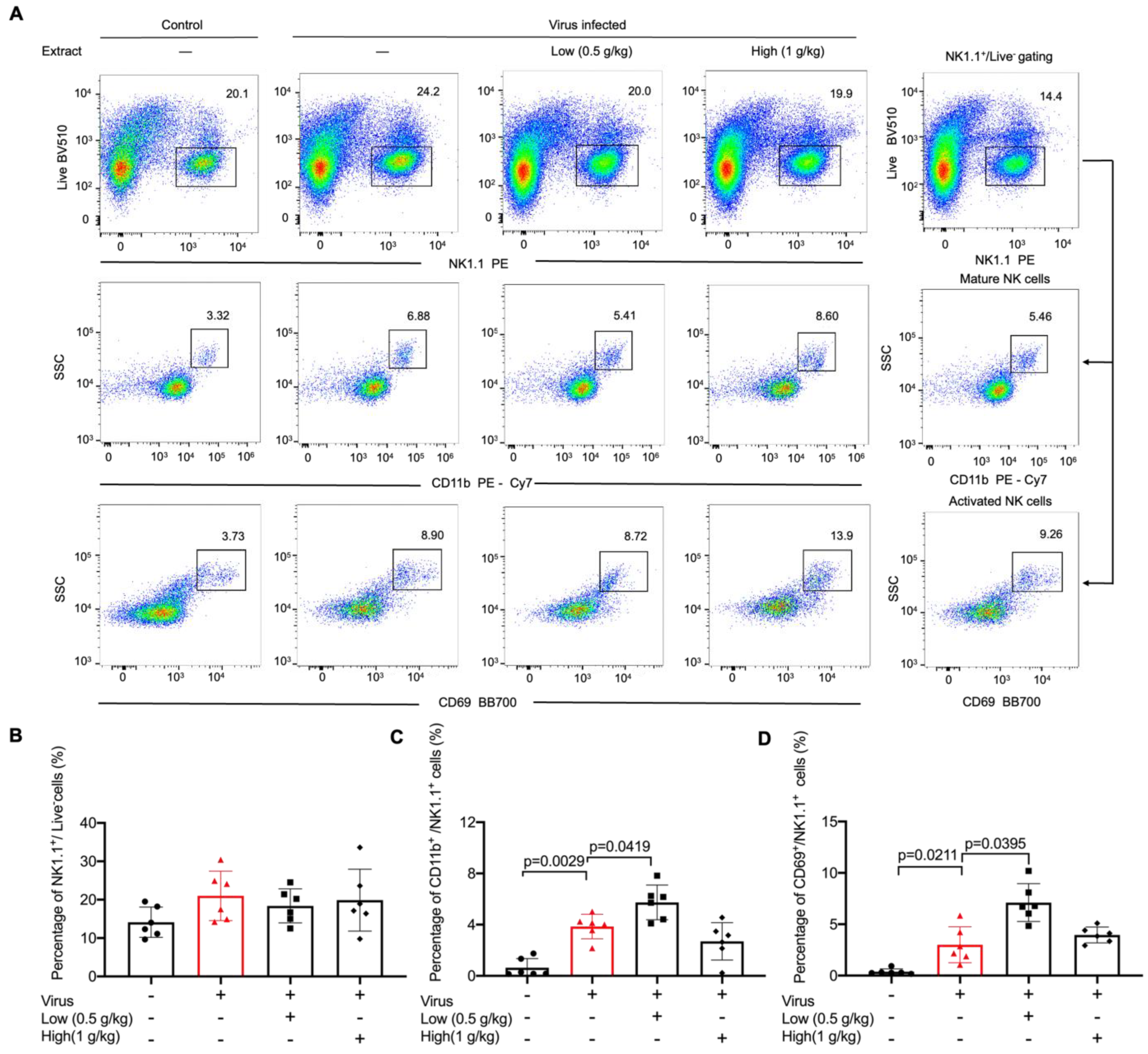
2.2. VO Extract Promoted Maturation and Activation of NK Cells in the Lungs in Response to Viral Infection
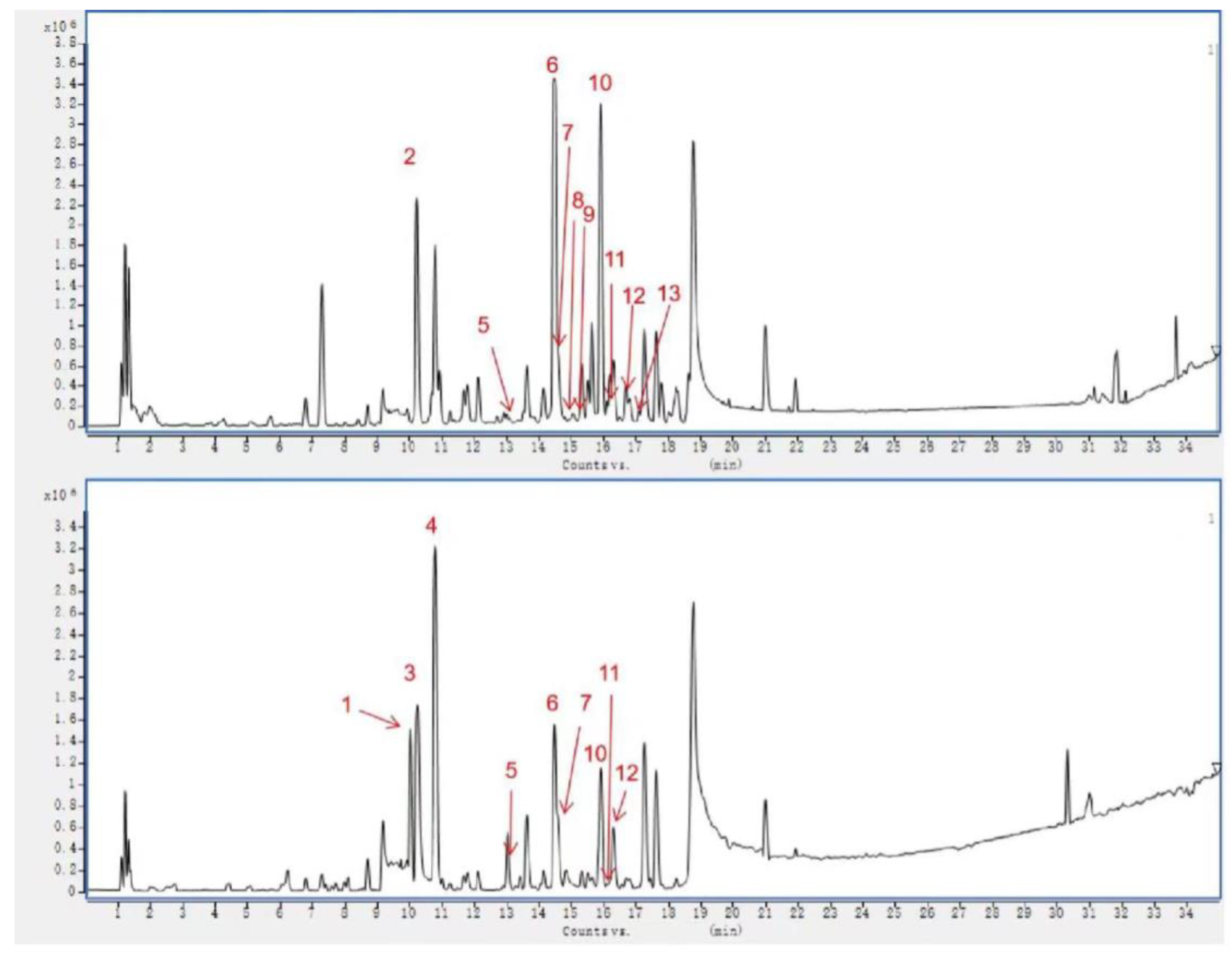
2.3. Identification of Chemical Composition from VO Extract by UPLC-Q-TOF-MS
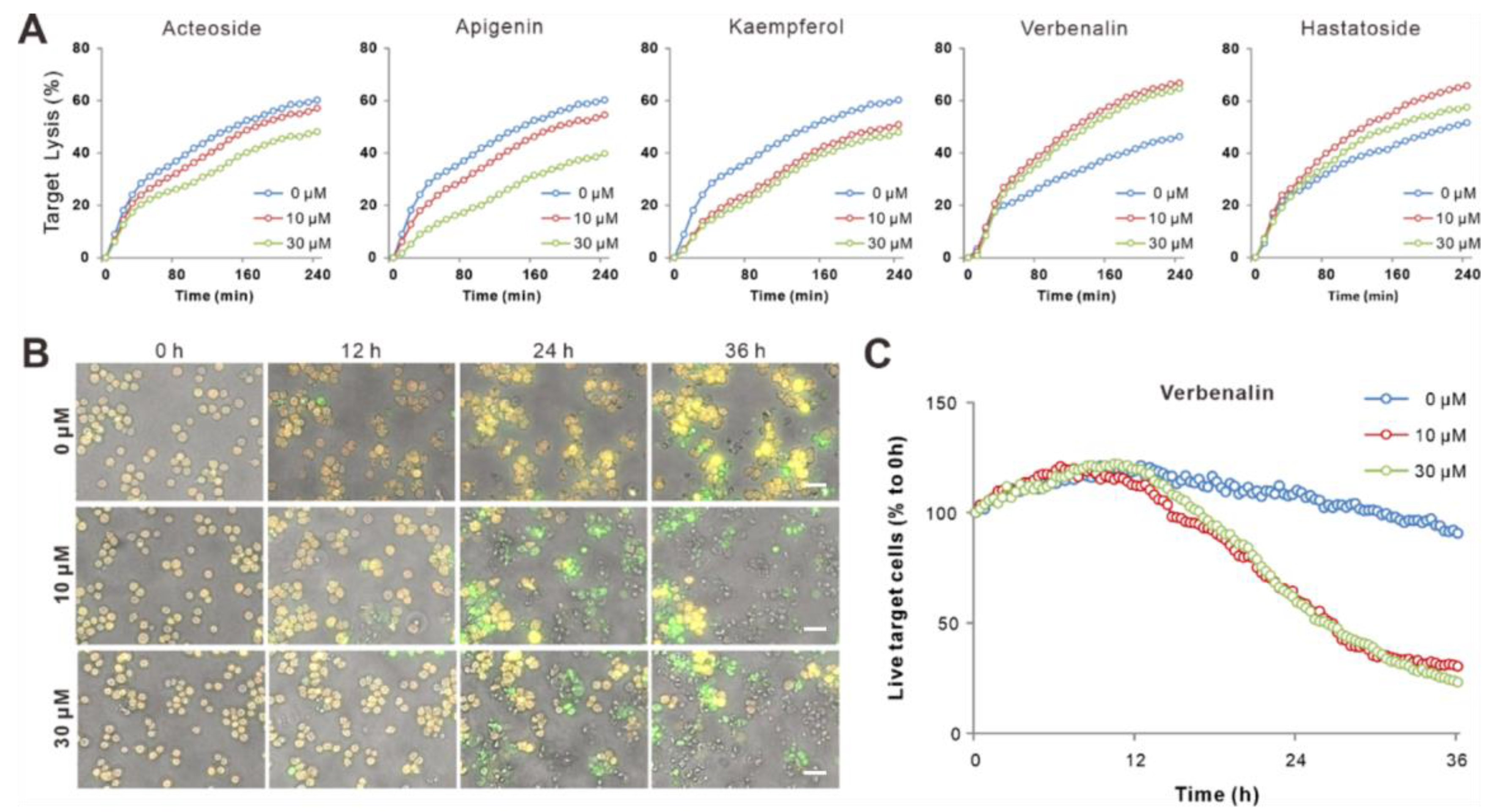
2.4. Bioactive Components of V. officinalis Enhanced NK Killing Efficiency
2.5. Verbenalin Accelerated NK Killing Processes
3. Discussion
4. Materials and Methods
4.1. Preparation of VO Extract
4.2. UPLC-Q-TOF-MS Analysis
4.3. Quantification of the Active Compounds from VO Extract
4.4. Mice and Virus
4.5. Virus Infection
4.6. Administration of VO Extract
4.7. Flow Cytometry Assay
4.8. Hematoxylin–Eosin (HE) Staining
4.9. Detection of IL-6, TNF-α, and IL-1β Levels in Serum
4.10. NK Celsl Preparation and Cell Culture
4.11. Real-Time Killing Assay
4.12. 3D Killing Assay and Live Cell Imaging
4.13. Proliferation Assay
4.14. Determination of Cytotoxic Protein Expression
4.15. CD107a Degranulation Assay
4.16. Molecular Docking
4.17. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| IS | immunological synapse |
| LG | lytic granules |
| MHC | major histocompatibility complex |
| NK | natural killer |
| PBMCs | peripheral blood mononuclear cells |
| VO extract | Verbena officinalis extract. |
References
- Kubica, P.; Szopa, A.; Kokotkiewicz, A.; Miceli, N.; Taviano, M.F.; Maugeri, A.; Cirmi, S.; Synowiec, A.; Gniewosz, M.; Elansary, H.O.; et al. Production of Verbascoside, Isoverbascoside and Phenolic Acids in Callus, Suspension, and Bioreactor Cultures of Verbena officinalis and Biological Properties of Biomass Extracts. Molecules 2020, 25, 5609. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.W.; Khan, A.U.; Ahmed, T. Anticonvulsant, Anxiolytic, and Sedative Activities of Verbena officinalis. Front. Pharmacol. 2016, 7, 499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kubica, P.; Szopa, A.; Dominiak, J.; Luczkiewicz, M.; Ekiert, H. Verbena officinalis (Common Vervain)—A Review on the Investigations of This Medicinally Important Plant Species. Planta Medica 2011, 86, 1241–1257. [Google Scholar] [CrossRef] [PubMed]
- Kubica, P.; Szopa, A.; Ekiert, H. Production of verbascoside and phenolic acids in biomass of Verbena officinalis L. (vervain) cultured under different in vitro conditions. Nat. Prod. Res. 2017, 31, 1663–1668. [Google Scholar] [CrossRef]
- Shu, J.; Chou, G.; Wang, Z. Two New Iridoids from Verbena officinalis L. Molecules 2014, 19, 10473–10479. [Google Scholar] [CrossRef] [Green Version]
- Casanova, E.; Garcia-Mina, J.M.; Calvo, M.I. Antioxidant and Antifungal Activity of Verbena officinalis L. Leaves. Plant Foods Hum. Nutr. 2008, 63, 93–97. [Google Scholar] [CrossRef]
- Lopez-Jornet, P.; Camacho-Alonso, F.; Gómez-Garcia, F.; Miñano, F.M.; Cañas, X.; Serafín, A.; Castillo, J.; Vicente-Ortega, V. Effects of potassium apigenin and verbena extract on the wound healing process of SKH-1 mouse skin. Int. Wound J. 2014, 11, 489–495. [Google Scholar] [CrossRef]
- Speroni, E.; Cervellati, R.; Costa, S.; Guerra, M.C.; Utan, A.; Govoni, P.; Berger, A.; Müller, A.; Stuppner, H. Effects of differential extraction of Verbena officinalis on rat models of inflammation, cicatrization and gastric damage. Planta Med. 2007, 73, 227–235. [Google Scholar] [CrossRef] [Green Version]
- Gazit, R.; Gruda, R.; Elboim, M.; Arnon, T.; Katz, G.; Achdout, H.; Hanna, J.H.; Qimron, U.; Landau, G.; Greenbaum, E.; et al. Lethal influenza infection in the absence of the natural killer cell receptor gene Ncr1. Nat. Immunol. 2006, 7, 517–523. [Google Scholar] [CrossRef]
- Nogusa, S.; Ritz, B.W.; Kassim, S.H.; Jennings, S.R.; Gardner, E.M. Characterization of age-related changes in natural killer cells during primary influenza infection in mice. Mech. Ageing Dev. 2008, 129, 223–230. [Google Scholar] [CrossRef]
- Stein-Streilein, J.; Guffee, J. In vivo treatment of mice and hamsters with antibodies to asialo GM1 increases morbidity and mortality to pulmonary influenza infection. J. Immunol. 1986, 136, 1435–1441. [Google Scholar] [CrossRef] [PubMed]
- Zhou, G.; Juang, S.W.W.; Kane, K.P. NK cells exacerbate the pathology of influenza virus infection in mice. Eur. J. Immunol. 2013, 43, 929–938. [Google Scholar] [CrossRef] [PubMed]
- Abdul-Careem, M.F.; Mian, M.F.; Yue, G.; Gillgrass, A.; Chenoweth, M.; Barra, N.G.; Chew, M.V.; Chan, T.; Al-Garawi, A.A.; Jordana, M.; et al. Critical Role of Natural Killer Cells in Lung Immunopathology During Influenza Infection in Mice. J. Infect. Dis. 2012, 206, 167–177. [Google Scholar] [CrossRef] [PubMed]
- Domizio, J.D.; Gulen, M.F.; Saidoune, F.; Thacker, V.V.; Yatim, A.; Sharma, K.; Nass, T.; Guenova, E.; Schaller, M.; Conrad, C.; et al. The cGAS-STING pathway drives type I IFN immunopathology in COVID-19. Nature 2022, 603, 145–151. [Google Scholar] [CrossRef]
- Kiener, M.; Roldan, N.; Machahua, C.; Sengupta, A.; Geiser, T.; Guenat, O.T.; Funke-Chambour, M.; Hobi, N.; Julio, M.K.-D. Human-Based Advanced in vitro Approaches to Investigate Lung Fibrosis and Pulmonary Effects of COVID-19. Front. Med. 2021, 8, 644678. [Google Scholar] [CrossRef] [PubMed]
- Sisay, M.; Bussa, N.; Gashaw, T. Evaluation of the Antispasmodic and Antisecretory Activities of the 80% Methanol Extracts of Verbena officinalis L: Evidence From In Vivo Antidiarrheal Study. J. Evid.-Based Integr. Med. 2019, 24, 2515690X19853264. [Google Scholar] [CrossRef] [Green Version]
- Laura, C.; Julie, C.; Nicolas, F.; Claude, R.; Eric, V.; Thierry, W. Maturation of mouse NK cells is a 4-stage developmental program. Blood 2009, 113, 5488–5496. [Google Scholar]
- Borrego, F.; Robertson, M.J.; Ritz, J.; Peña, J.; Solana, R. CD69 is a stimulatory receptor for natural killer cell and its cytotoxic effect is blocked by CD94 inhibitory receptor. Immunology 1999, 97, 159–165. [Google Scholar] [CrossRef]
- Kummerow, C.; Schwarz, E.C.; Bufe, B.; Zufall, F.; Hoth, M.; Qu, B. A simple, economic, time-resolved killing assay. Eur. J. Immunol. 2014, 44, 1870–1872. [Google Scholar] [CrossRef]
- Backes, C.S.; Friedmann, K.S.; Mang, S.; Knörck, A.; Hoth, M.; Kummerow, C. Natural killer cells induce distinct modes of cancer cell death: Discrimination, quantification, and modulation of apoptosis, necrosis, and mixed forms. J. Biol. Chem. 2018, 293, 16348–16363. [Google Scholar] [CrossRef] [Green Version]
- Bhat, R.; Watzl, C. Serial Killing of Tumor Cells by Human Natural Killer Cells—Enhancement by Therapeutic Antibodies. PLoS ONE 2007, 2, e326. [Google Scholar] [CrossRef] [PubMed]
- Prager, I.; Liesche, C.; van Ooijen, H.; Urlaub, D.; Verron, Q.; Sandström, N.; Fasbender, F.; Claus, M.; Eils, R.; Beaudouin, J.; et al. NK cells switch from granzyme B to death receptor-mediated cytotoxicity during serial killing. Exp Med. 2019, 216, 2113–2127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ross, S.M. An integrative approach to rhinosinusitis in children, part II. Holist. Nurs. Pract. 2010, 24, 49–51. [Google Scholar] [CrossRef] [PubMed]
- Xiong, W.-Z.; Wang, G.; Du, J.; Ai, W. Efficacy of herbal medicine (Xuanfei Baidu decoction) combined with conventional drug in treating COVID-19:A pilot randomized clinical trial. Integr. Med. Res. 2020, 9, 100489. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Fisher, E.M.; Murasko, D.M. CD8 T cell responses to influenza virus infection in aged mice. Ageing Res. Rev. 2011, 10, 422–427. [Google Scholar] [CrossRef] [Green Version]
- Tian, X.; Eikmans, M.; van der Hoorn, M.-L. The Role of Macrophages in Oocyte Donation Pregnancy: A Systematic Review. Int. J. Mol. Sci. 2020, 21, 939. [Google Scholar] [CrossRef] [Green Version]
- Maeda, S.; Omata, M. Inflammation and cancer: Role of nuclear factor-kappaB activation. Cancer Sci. 2008, 99, 836–842. [Google Scholar] [CrossRef]
- Fontenelle, T.P.C.; Lima, G.C.; Mesquita, J.X.; Lopes, J.L.D.S.; de Brito, T.V.; Júnior, F.D.C.V.; Sales, A.B.; Aragão, K.S.; Souza, M.H.L.P.; Barbosa, A.L.D.R.; et al. Lectin obtained from the red seaweed Bryothamnion triquetrum: Secondary structure and anti-inflammatory activity in mice. Int. J. Biol. Macromol. 2018, 112, 1122–1130. [Google Scholar] [CrossRef]
- Weber, A.; Wasiliew, P.; Kracht, M. Interleukin-1 (IL-1) Pathway. Sci. Signal. 2010, 3, cm1. [Google Scholar] [CrossRef]
- Koppinger, M.P.; Lopez-Pier, M.A.; Skaria, R.; Harris, P.R.; Konhilas, J.P. Lactobacillus reuteri attenuates cardiac injury without lowering cholesterol in low-density lipoprotein receptor-deficient mice fed standard chow. Am. J. Physiol. Heart Circ. Physiol. 2020, 319, H32–H41. [Google Scholar] [CrossRef]
- Viji, V.; Shobha, B.; Kavitha, S.K.; Ratheesh, M.; Kripa, K.; Helen, A. Betulinic acid isolated from Bacopa monniera (L.) Wettst suppresses lipopolysaccharide stimulated interleukin-6 production through modulation of nuclear factor-kappaB in peripheral blood mononuclear cells. Int. Immunopharmacol. 2010, 10, 843–849. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.L.; Li, H.B.; Miao, M.S. Effect of total glucosides of Verbena officinalis L. on rat model for chronic nonbacterial prostatitis. China Med. Herald. 2016, 13, 4–7. [Google Scholar]
- Dong, S.M.; Gao, P.H.; Liang, W.H. Effects of Verbenalin on Airway Inflammation, Airway Smooth Muscle Cell Proliferation and RhoA Expression in Rats with Bronchial Asthma Through NF-κB/MAPK Signaling Pathway. Guid. J. Tradit. Chin. Med. Pharm. 2021, 27, 23–27. [Google Scholar]
- Prager, I.; Watzl, C. Mechanisms of natural killer cell-mediated cellular cytotoxicity. J. Leukoc. Biol. 2019, 105, 1319–1329. [Google Scholar] [CrossRef]
- Orange, J.S. Formation and function of the lytic NK-cell immunological synapse. Nat. Rev. Immunol. 2008, 8, 713–725. [Google Scholar] [CrossRef] [Green Version]
- Pegram, H.J.; Andrews, D.M.; Smyth, M.J.; Darcy, P.K.; Kershaw, M.H. Activating and inhibitory receptors of natural killer cells. Immunol. Cell Biol. 2011, 89, 216–224. [Google Scholar] [CrossRef]
- Lanier, L.L. On guard--activating NK cell receptors. Nat. Immunol. 2001, 2, 23–27. [Google Scholar] [CrossRef]
- Sivori, S.; Vacca, P.; Del Zotto, G.; Munari, E.; Mingari, M.C.; Moretta, L. Human NK cells: Surface receptors, inhibitory checkpoints, and translational applications. Cell. Mol. Immunol. 2019, 16, 430–441. [Google Scholar] [CrossRef]
- Gross, C.C.; Brzostowski, J.A.; Liu, D.; Long, E.O. Tethering of Intercellular Adhesion Molecule on Target Cells Is Required for LFA-1–Dependent NK Cell Adhesion and Granule Polarization. J. Immunol. 2010, 185, 2918–2926. [Google Scholar] [CrossRef] [Green Version]
- Mentlik, A.N.; Sanborn, K.B.; Holzbaur, E.L.; Orange, J.S. Rapid Lytic Granule Convergence to the MTOC in Natural Killer Cells Is Dependent on Dynein But Not Cytolytic Commitment. Mol. Biol. Cell 2010, 21, 2241–2256. [Google Scholar] [CrossRef] [Green Version]
- Qu, B.; Pattu, V.; Junker, C.; Schwarz, E.C.; Bhat, S.S.; Kummerow, C.; Marshall, M.; Matti, U.; Neumann, F.; Pfreundschuh, M.; et al. Docking of Lytic Granules at the Immunological Synapse in Human CTL Requires Vti1b-Dependent Pairing with CD3 Endosomes. J. Immunol. 2011, 186, 6894–6904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertrand, F.; Müller, S.; Roh, K.-H.; Laurent, C.; Dupré, L.; Valitutti, S. An initial and rapid step of lytic granule secretion precedes microtubule organizing center polarization at the cytotoxic T lymphocyte/target cell synapse. Proc. Natl. Acad. Sci. USA 2013, 110, 6073–6078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tuomela, K.; Ambrose, A.R.; Davis, D.M. Escaping Death: How Cancer Cells and Infected Cells Resist Cell-Mediated Cytotoxicity. Front. Immunol. 2022, 13, 867098. [Google Scholar] [CrossRef]
- Kong, L.; Deng, J.; Zhou, X.; Cai, B.; Zhang, B.; Chen, X.; Chen, Z.; Wang, W. Sitagliptin activates the p62–Keap1–Nrf2 signalling pathway to alleviate oxidative stress and excessive autophagy in severe acute pancreatitis-related acute lung injury. Cell Death Dis. 2021, 12, 928. [Google Scholar] [CrossRef]
- Schmidt, J.; Rattner, D.W.; Lewandrowski, K.; Compton, C.C.; Mandavilli, U.; Knoefel, W.T.; Warshaw, A.L. A Better Model of Acute Pancreatitis for Evaluating Therapy. Ann. Surg. 1992, 215, 44–56. [Google Scholar] [CrossRef] [PubMed]
- Matute-Bello, G.; Winn, R.K.; Jonas, M.; Chi, E.Y.; Martin, T.R.; Liles, W.C. Fas (CD95) Induces Alveolar Epithelial Cell Apoptosis in Vivo: Implications for Acute Pulmonary Inflammation. Am. J. Pathol. 2001, 158, 153–161. [Google Scholar] [CrossRef]
- Zhao, R.; Zhou, X.; Khan, E.S.; Alansary, D.; Friedmann, K.S.; Yang, W.; Schwarz, E.C.; del Campo, A.; Hoth, M.; Qu, B. Targeting the Microtubule-Network Rescues CTL Killing Efficiency in Dense 3D Matrices. Front. Immunol. 2021, 12, 729820. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dai, X.; Zhou, X.; Shao, R.; Zhao, R.; Yanamandra, A.K.; Xing, Z.; Ding, M.; Wang, J.; Liu, T.; Zheng, Q.; et al. Bioactive Constituents of Verbena officinalis Alleviate Inflammation and Enhance Killing Efficiency of Natural Killer Cells. Int. J. Mol. Sci. 2023, 24, 7144. https://doi.org/10.3390/ijms24087144
Dai X, Zhou X, Shao R, Zhao R, Yanamandra AK, Xing Z, Ding M, Wang J, Liu T, Zheng Q, et al. Bioactive Constituents of Verbena officinalis Alleviate Inflammation and Enhance Killing Efficiency of Natural Killer Cells. International Journal of Molecular Sciences. 2023; 24(8):7144. https://doi.org/10.3390/ijms24087144
Chicago/Turabian StyleDai, Xiangdong, Xiangda Zhou, Rui Shao, Renping Zhao, Archana K. Yanamandra, Zhimei Xing, Mingyu Ding, Junhong Wang, Tao Liu, Qi Zheng, and et al. 2023. "Bioactive Constituents of Verbena officinalis Alleviate Inflammation and Enhance Killing Efficiency of Natural Killer Cells" International Journal of Molecular Sciences 24, no. 8: 7144. https://doi.org/10.3390/ijms24087144