
Compositional Analysis of Glycosaminoglycans in Different Lung Cancer Types—A Pilot Study
 , ,
, ,  , , , , ,
, , , , ,  and
and
Abstract
:1. Introduction
2. Results
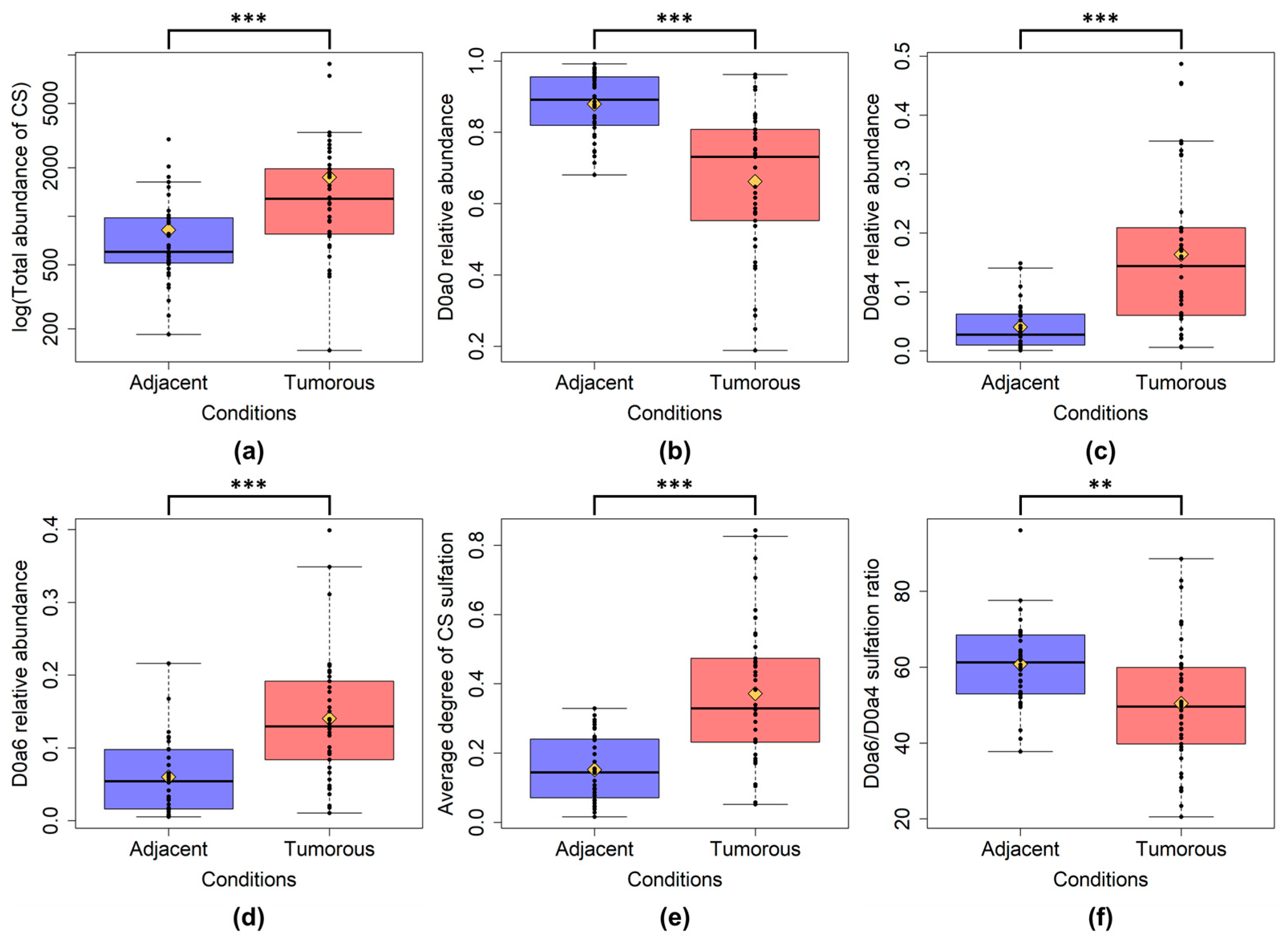
2.1. CS and HS Content and Sulfation Characteristics between All Tumor and All Tumor Adjacent Normal Samples
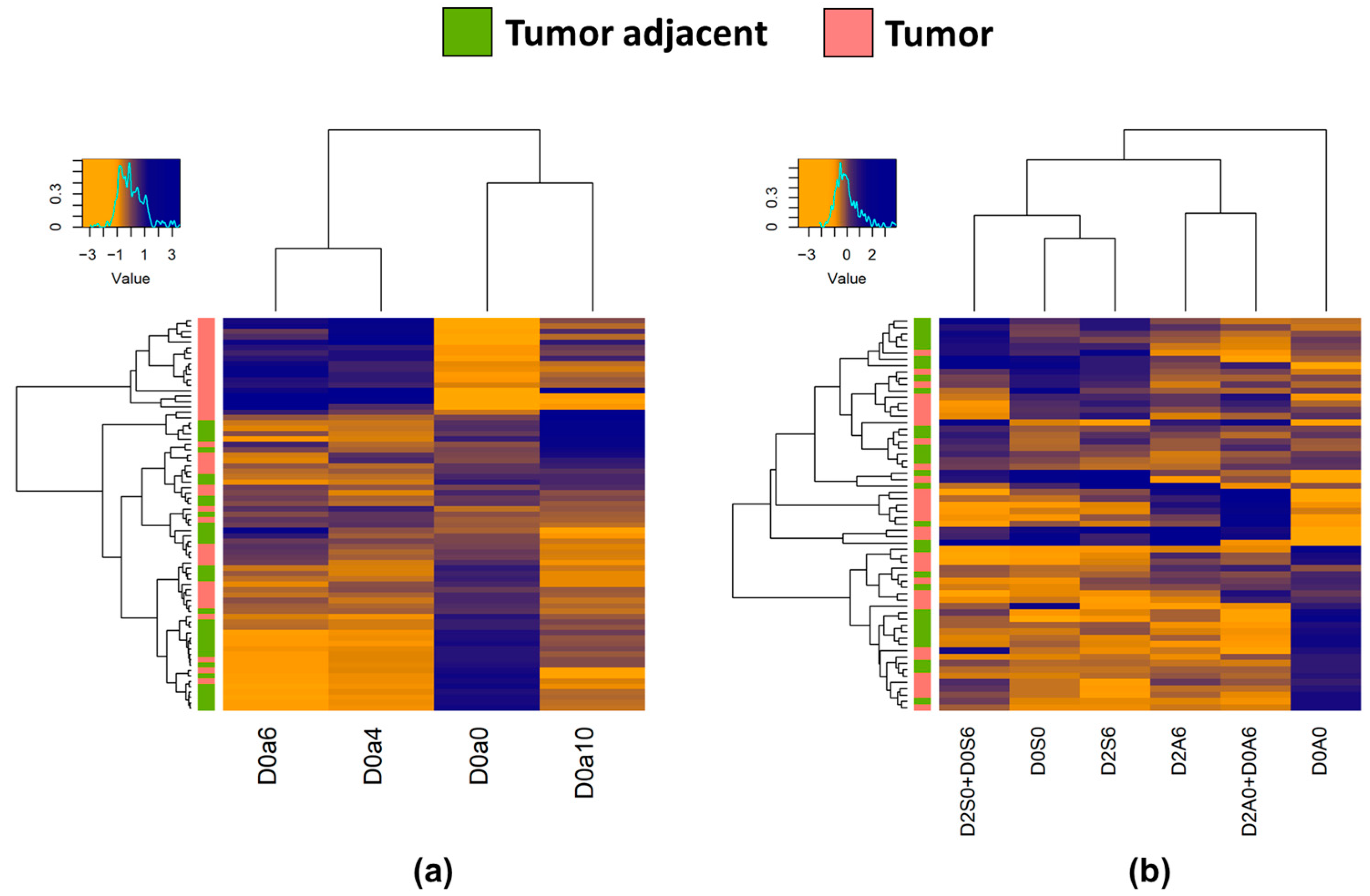
2.2. CS and HS Content and Sulfation Characteristics between Tumor and Corresponding Tumor Adjacent Normal Samples
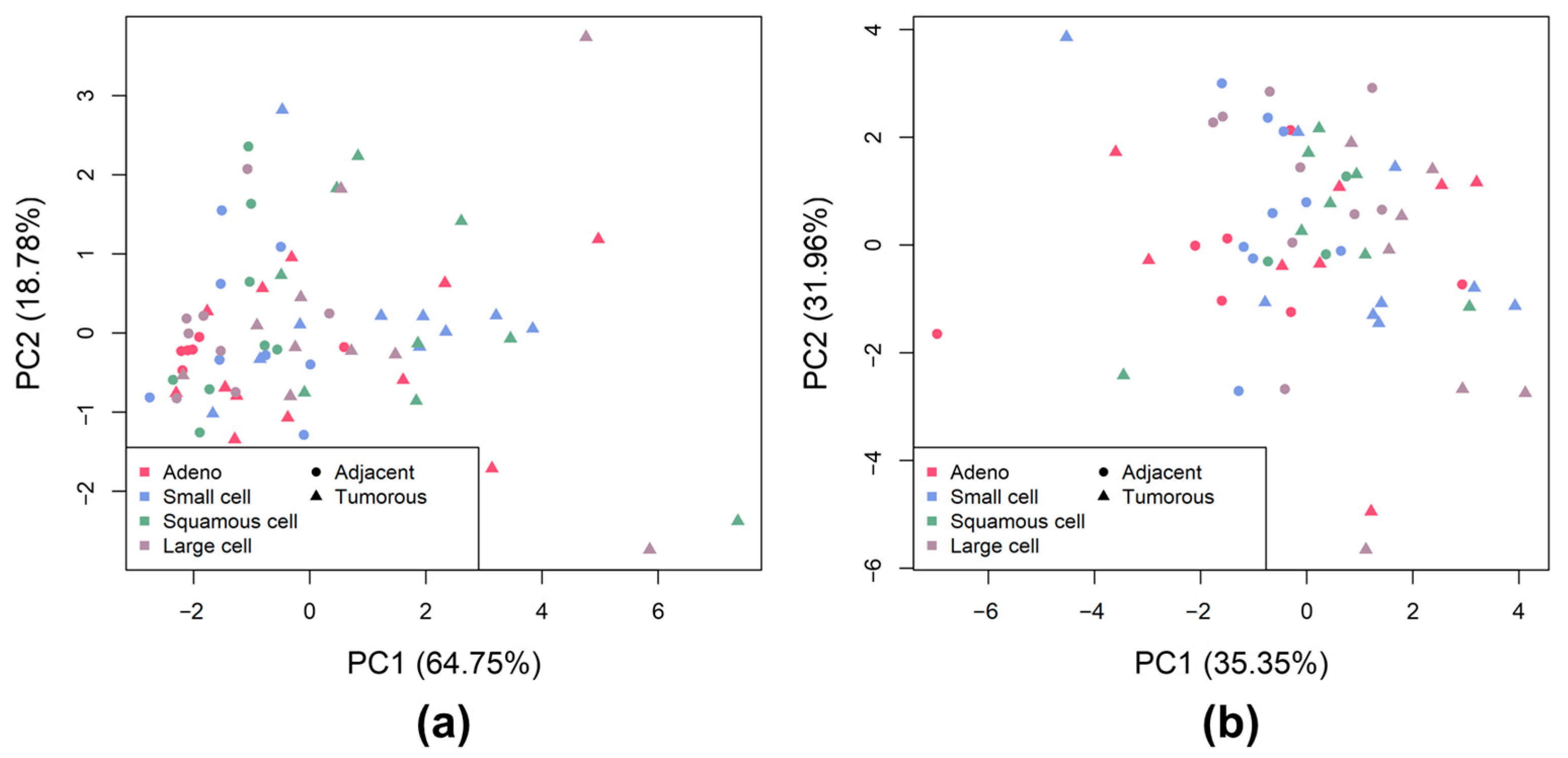
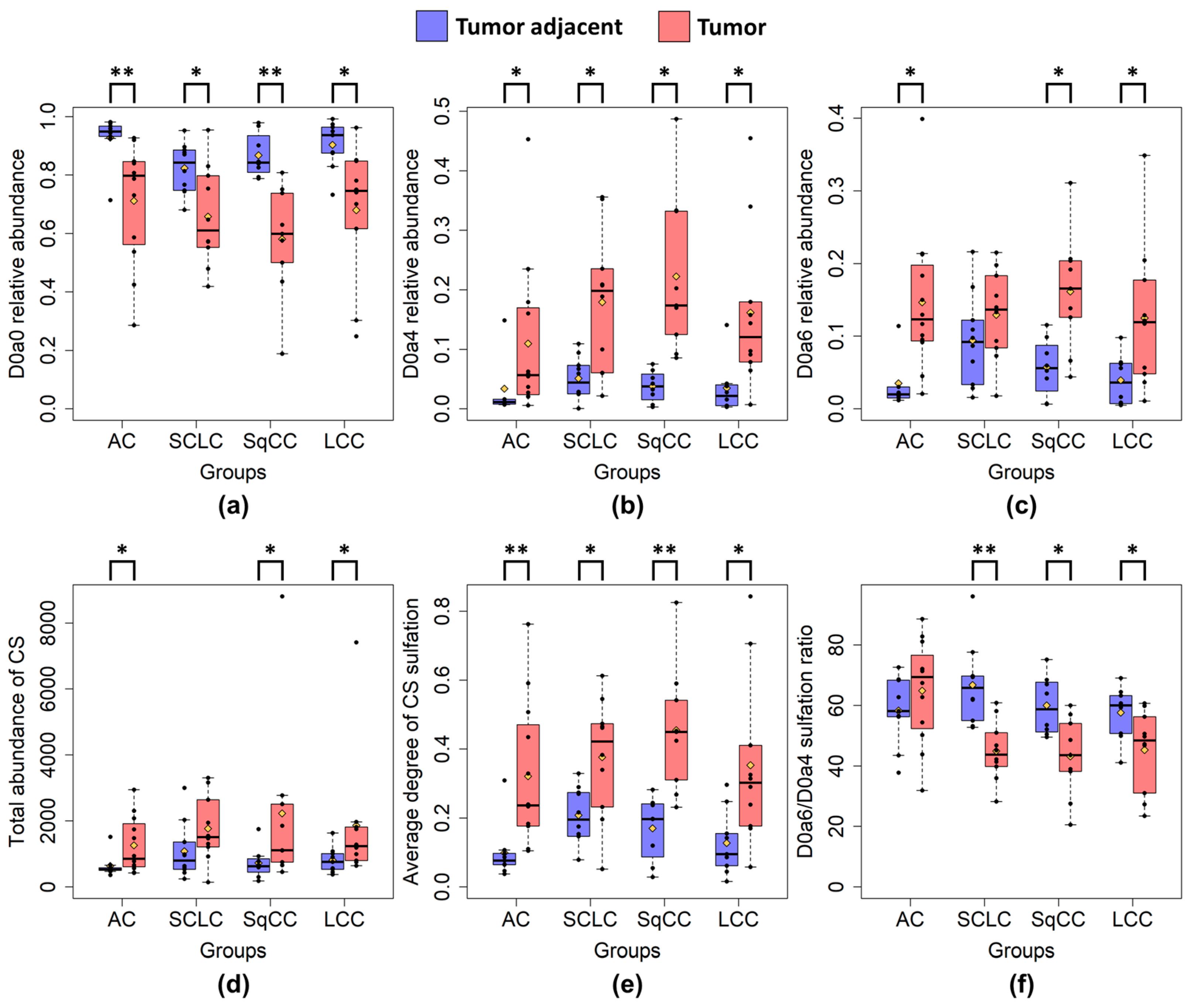
2.3. CS and HS Sulfation between Lung Tumor Phenotypes
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Tissue Sample Selection and Preparation
4.3. Chondroitin Sulfate Digestion on Tissue Surface
4.4. Heparan Sulfate Digestion on Tissue Surface
4.5. Sample Cleanup with Combined Extraction Method (Cotton Wool and Graphite Solid-Phase Extraction)
4.6. Liquid Chromatography-Mass Spectrometry
4.7. Data Evaluation and Interpretation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Nooreldeen, R.; Bach, H. Current and Future Development in Lung Cancer Diagnosis. Int. J. Mol. Sci. 2021, 22, 8661. [Google Scholar] [CrossRef]
- Inamura, K. Lung Cancer: Understanding Its Molecular Pathology and the 2015 WHO Classification. Front. Oncol. 2017, 7, 193. [Google Scholar] [CrossRef] [Green Version]
- Ettinger, D.S.; Akerley, W.; Bepler, G.; Blum, M.G.; Chang, A.; Cheney, R.T.; Yang, S.C. Non-small-cell lung cancer. Nat. Rev. Dis. Prim. 2015, 1, 15009. [Google Scholar]
- Travis, W.D. Pathology of lung cancer. Clin. Chest Med. 2002, 23, 65–81. [Google Scholar] [CrossRef]
- Travis, W.D.; Brambilla, E.; Riely, G.J. New Pathologic Classification of Lung Cancer: Relevance for Clinical Practice and Clinical Trials. J. Clin. Oncol. 2013, 31, 992–1001. [Google Scholar] [CrossRef] [Green Version]
- Stowell, S.R.; Ju, T.; Cummings, R.D. Protein Glycosylation in Cancer. Annu. Rev. Pathol. Mech. Dis. 2015, 10, 473–510. [Google Scholar] [CrossRef] [Green Version]
- Lemjabbar-Alaoui, H.; McKinney, A.; Yang, Y.-W.; Tran, V.M.; Phillips, J.J. Chapter Nine-Glycosylation Alterations in Lung and Brain Cancer. In Glycosylation and Cancer; Drake, R.R., Ball, L.E.B.T.-A., Eds.; Academic Press: Cambridge, MA, USA, 2015; Volume 126, pp. 305–344. [Google Scholar]
- Esko, J.D.; Kimata, K.; Lindahl, U. Proteoglycans and Sulfated Glycosaminoglycans. In Essentials of Glycobiology, 2nd ed.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2017. [Google Scholar]
- Alper, J. Turning Sweet on Cancer. Science 2003, 301, 159–160. [Google Scholar] [CrossRef]
- Hakomori, S. Aberrant Glycosylation in Tumors and Tumor-Associated Carbohydrate Antigens; Vande Woude, G.F., Klein, G.B.T.-A., Eds.; Academic Press: Cambridge, MA, USA, 1989; Volume 52, pp. 257–331. [Google Scholar]
- Asada, M.; Furukawa, K.; Segawa, K.; Endo, T.; Kobata, A. Increased Expression of Highly Branched N-Glycans at Cell Surface Is Correlated with the Malignant Phenotypes of Mouse Tumor Cells. Cancer Res. 1997, 57, 1073–1080. [Google Scholar]
- Mi, W.; Gu, Y.; Han, C.; Liu, H.; Fan, Q.; Zhang, X.; Yu, W. O-GlcNAcylation is a novel regulator of lung and colon cancer malignancy. Biochim. Biophys. Acta-Mol. Basis Dis. 2011, 1812, 514–519. [Google Scholar] [CrossRef] [Green Version]
- Couchman, J.R.; Pataki, C.A. An Introduction to Proteoglycans and Their Localization. J. Histochem. Cytochem. 2012, 60, 885–897. [Google Scholar] [CrossRef] [PubMed]
- Gibbs, R.V. Cytokines and Glycosaminoglycans (GAGS). In Glycobiology and Medicine; Axford, J.S., Ed.; Springer: New York, NY, USA, 2003; pp. 125–143. [Google Scholar]
- Kjellén, L.; Lindahl, U. Specificity of glycosaminoglycan–protein interactions. Curr. Opin. Struct. Biol. 2018, 50, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Kreuger, J.; Kjellén, L. Heparan Sulfate Biosynthesis: Regulation and Variability. J. Histochem. Cytochem. 2012, 60, 898–907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kovalszky, I.; Pogany, G.; Molnar, G.; Jeney, A.; Lapis, K.; Karacsonyi, S.; Szecseny, A.; Iozzo, R.V. Altered glycosaminoglycan composition in reactive and neoplastic human liver. Biochem. Biophys. Res. Commun. 1990, 167, 883–890. [Google Scholar] [CrossRef]
- Tóth, G.; Sugár, S.; Pál, D.; Fügedi, K.D.; Drahos, L.; Schlosser, G.; Turiák, L. Glycosaminoglycan Analysis of FFPE Tissues from Prostate Cancer and Benign Prostate Hyperplasia Patients Reveals Altered Regulatory Functions and Independent Markers for Survival. Cancers 2022, 14, 4867. [Google Scholar] [CrossRef]
- Tóth, G.; Pál, D.; Sugár, S.; Kovalszky, I.; Dezső, K.; Schlosser, G.; Turiák, L. Expression of glycosaminoglycans in cirrhotic liver and hepatocellular carcinoma—A pilot study including etiology. Anal. Bioanal. Chem. 2022, 414, 3837–3846. [Google Scholar] [CrossRef]
- Hatae, Y.; Atsuta, T.; Makita, A. Glycosaminoglycans in human lung carcinoma. GANN Jpn. J. Cancer Res. 1977, 68, 59–63. [Google Scholar]
- Horai, T.; Nakamura, N.; Tateishi, R.; Hattori, S. Glycosaminoglycans in human lung cancer. Cancer 1981, 48, 2016–2021. [Google Scholar] [CrossRef]
- Li, G.; Li, L.; Joo, E.J.; Son, J.W.; Kim, Y.J.; Kang, J.K.; Lee, K.B.; Zhang, F.; Linhardt, R.J. Glycosaminoglycans and glycolipids as potential biomarkers in lung cancer. Glycoconj. J. 2017, 34, 661–669. [Google Scholar] [CrossRef]
- Sugár, S.; Bugyi, F.; Tóth, G.; Pápay, J.; Kovalszky, I.; Tornóczky, T.; Drahos, L.; Turiák, L. Proteomic Analysis of Lung Cancer Types—A Pilot Study. Cancers 2022, 14, 2629. [Google Scholar] [CrossRef]
- Solakyildirim, K. Recent advances in glycosaminoglycan analysis by various mass spectrometry techniques. Anal. Bioanal. Chem. 2019, 311, 3731–3741. [Google Scholar] [CrossRef]
- Zaia, J. Principles of mass spectrometry of glycosaminoglycans. J. Biomacromol. Mass. Spectrom. 2005, 1, 3–36. [Google Scholar]
- Lawrence, R.; Lu, H.; Rosenberg, R.D.; Esko, J.D.; Zhang, L. Disaccharide structure code for the easy representation of constituent oligosaccharides from glycosaminoglycans. Nat. Methods 2008, 5, 291–292. [Google Scholar] [CrossRef]
- Turiák, L.; Shao, C.; Meng, L.; Khatri, K.; Leymarie, N.; Wang, Q.; Pantazopoulos, H.; Leon, D.R.; Zaia, J. Workflow for Combined Proteomics and Glycomics Profiling from Histological Tissues. Anal. Chem. 2014, 86, 9670–9678. [Google Scholar] [CrossRef] [Green Version]
- Tóth, G.; Vékey, K.; Sugár, S.; Kovalszky, I.; Drahos, L.; Turiák, L. Salt gradient chromatographic separation of chondroitin sulfate disaccharides. J. Chromatogr. A 2020, 1619, 460979. [Google Scholar] [CrossRef]
- Tóth, G.; Vékey, K.; Drahos, L.; Horváth, V.; Turiák, L. Salt and solvent effects in the microscale chromatographic separation of heparan sulfate disaccharides. J. Chromatogr. A 2020, 1610, 460548. [Google Scholar] [CrossRef] [Green Version]
- Turiák, L.; Ozohanics, O.; Tóth, G.; Ács, A.; Révész, Á.; Vékey, K.; Telekes, A.; Drahos, L. High sensitivity proteomics of prostate cancer tissue microarrays to discriminate between healthy and cancerous tissue. J. Proteom. 2019, 197, 82–91. [Google Scholar] [CrossRef]
- Tóth, G.; Turiák, L. HPLC-MS Characterization of Tissue-Derived Heparan Sulfate and Chondroitin Sulfate. In Proteoglycans: Methods and Protocols; Karamanos, N.K., Ed.; Springer: New York, NY, USA, 2023; pp. 71–90. [Google Scholar] [CrossRef]
- Kawai, T.; Suzuki, M.; Kageyama, K. Glycosaminoglycans in lung carcinoma. Hum. Pathol. 1988, 19, 1288–1292. [Google Scholar] [CrossRef]
- Iozzo, R.V.; Schaefer, L. Proteoglycan form and function: A comprehensive nomenclature of proteoglycans. Matrix Biol. 2015, 42, 11–55. [Google Scholar] [CrossRef]
- Guo, J.-Y.; Chiu, C.-H.; Wang, M.-J.; Li, F.-A.; Chen, J.-Y. Proteoglycan serglycin promotes non-small cell lung cancer cell migration through the interaction of its glycosaminoglycans with CD44. J. Biomed. Sci. 2020, 27, 2. [Google Scholar] [CrossRef] [Green Version]
- Pudełko, A.; Wisowski, G.; Olczyk, K.; Koźma, E.M. The dual role of the glycosaminoglycan chondroitin-6-sulfate in the development, progression and metastasis of cancer. FEBS J. 2019, 286, 1815–1837. [Google Scholar] [CrossRef] [Green Version]
- Spliid, C.B.; Toledo, A.G.; Sanderson, P.; Mao, Y.; Gatto, F.; Gustavsson, T.; Clausen, T.M. The specificity of the malarial VAR2CSA protein for chondroitin sulfate depends on 4-O-sulfation and ligand accessibility. J. Biol. Chem. 2021, 297, 6. [Google Scholar] [CrossRef] [PubMed]
- Oo, H.Z.; Lohinai, Z.; Khazamipour, N.; Lo, J.; Kumar, G.; Pihl, J.; Adomat, H.; Nabavi, N.; Behmanesh, H.; Zhai, B.; et al. Oncofetal Chondroitin Sulfate Is a Highly Expressed Therapeutic Target in Non-Small Cell Lung Cancer. Cancers 2021, 13, 4489. [Google Scholar] [CrossRef]
- Watanabe, Y.; Aoki-Kinoshita, K.F.; Ishihama, Y.; Okuda, S. GlycoPOST realizes FAIR principles for glycomics mass spectrometry data. Nucleic Acids Res. 2021, 49, D1523–D1528. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. of Patients | No. of Tumor Samples | No. of Tumor Adjacent Samples | |
|---|---|---|---|
| Total No. | 42 | 81 (CS:41, HS: 40) | 72 (CS: 36, HS: 36) |
| Age | 65.5 (54–79) | 65.5 (54–79) | 65 (54–75) |
| Male | 44 | 40 | |
| CS | 22 | 22 | 20 |
| HS | 22 | 22 | 20 |
| Female | 37 | 32 | |
| CS | 19 | 19 | 16 |
| HS | 18 | 18 | 16 |
| Histology | |||
| Adenocarcinoma | 22 | 18 | |
| CS | 12 | 12 | 9 |
| HS | 10 | 10 | 9 |
| Large cell carcinoma | 20 | 18 | |
| CS | 10 | 10 | 9 |
| HS | 10 | 10 | 9 |
| Small cell lung carcinoma | 20 | 20 | |
| CS | 10 | 10 | 10 |
| HS | 10 | 10 | 10 |
| Squamous cell carcinoma | 19 | 16 | |
| CS | 9 | 9 | 8 |
| HS | 10 | 10 | 8 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pál, D.; Tóth, G.; Sugár, S.; Fügedi, K.D.; Szabó, D.; Kovalszky, I.; Papp, D.; Schlosser, G.; Tóth, C.; Tornóczky, T.; et al. Compositional Analysis of Glycosaminoglycans in Different Lung Cancer Types—A Pilot Study. Int. J. Mol. Sci. 2023, 24, 7050. https://doi.org/10.3390/ijms24087050
Pál D, Tóth G, Sugár S, Fügedi KD, Szabó D, Kovalszky I, Papp D, Schlosser G, Tóth C, Tornóczky T, et al. Compositional Analysis of Glycosaminoglycans in Different Lung Cancer Types—A Pilot Study. International Journal of Molecular Sciences. 2023; 24(8):7050. https://doi.org/10.3390/ijms24087050
Chicago/Turabian StylePál, Domonkos, Gábor Tóth, Simon Sugár, Kata Dorina Fügedi, Dániel Szabó, Ilona Kovalszky, Dávid Papp, Gitta Schlosser, Csaba Tóth, Tamás Tornóczky, and et al. 2023. "Compositional Analysis of Glycosaminoglycans in Different Lung Cancer Types—A Pilot Study" International Journal of Molecular Sciences 24, no. 8: 7050. https://doi.org/10.3390/ijms24087050