
HuR Promotes the Differentiation of Goat Skeletal Muscle Satellite Cells by Regulating Myomaker mRNA Stability

 , , , , , and
, , , , , and 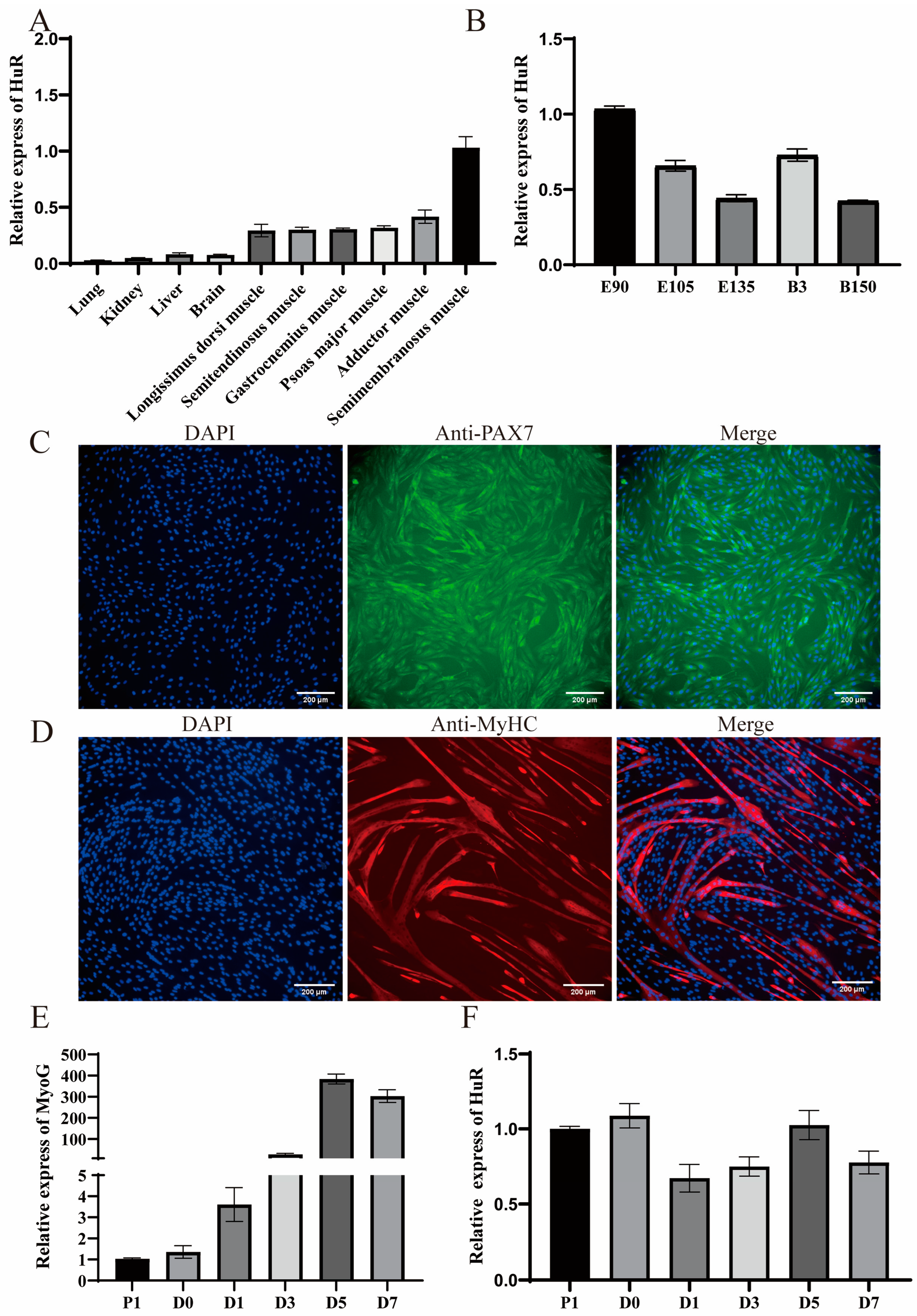{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Expression Patterns of HuR
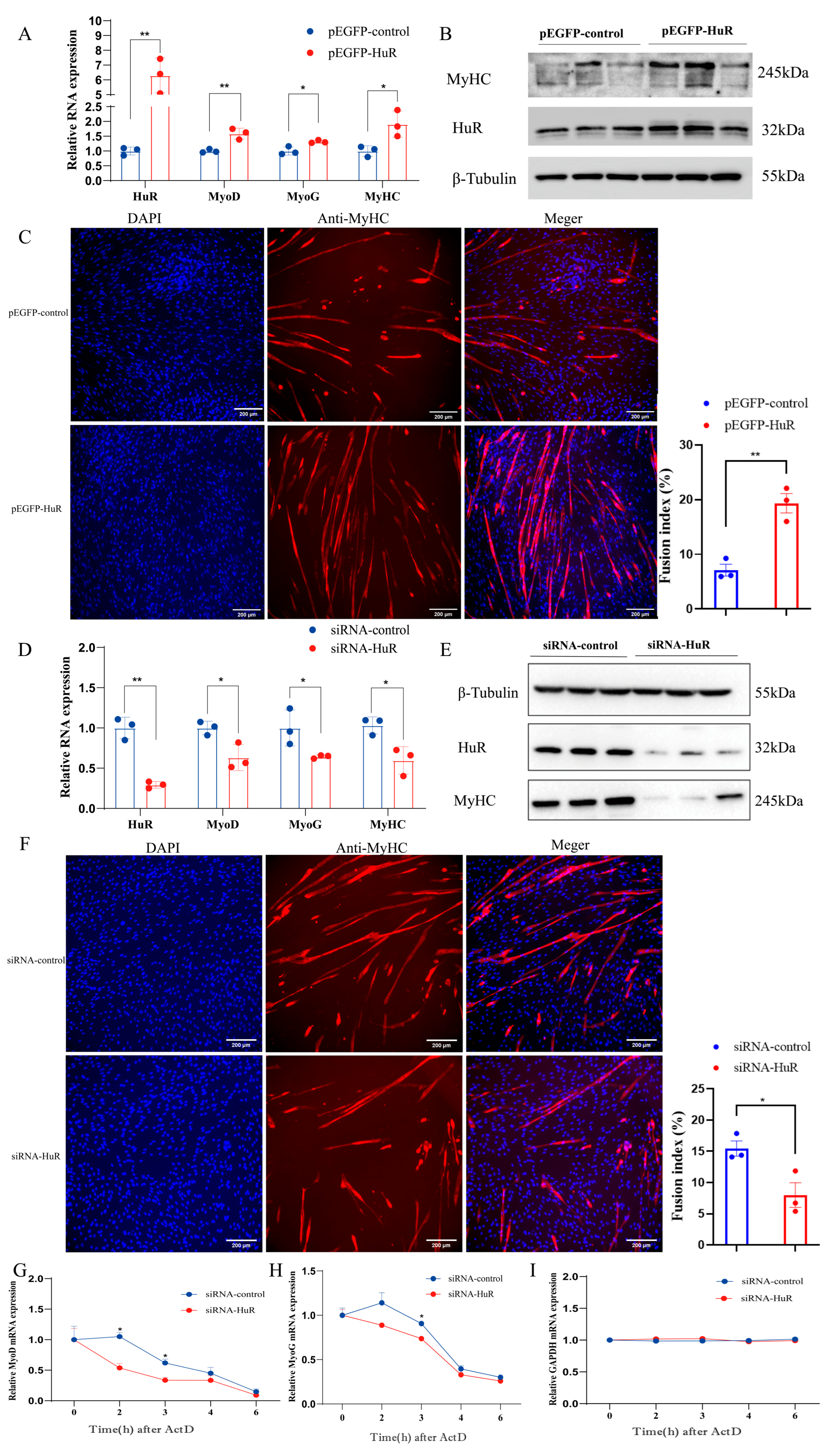
2.2. HuR Promotes MuSC Differentiation in Goats
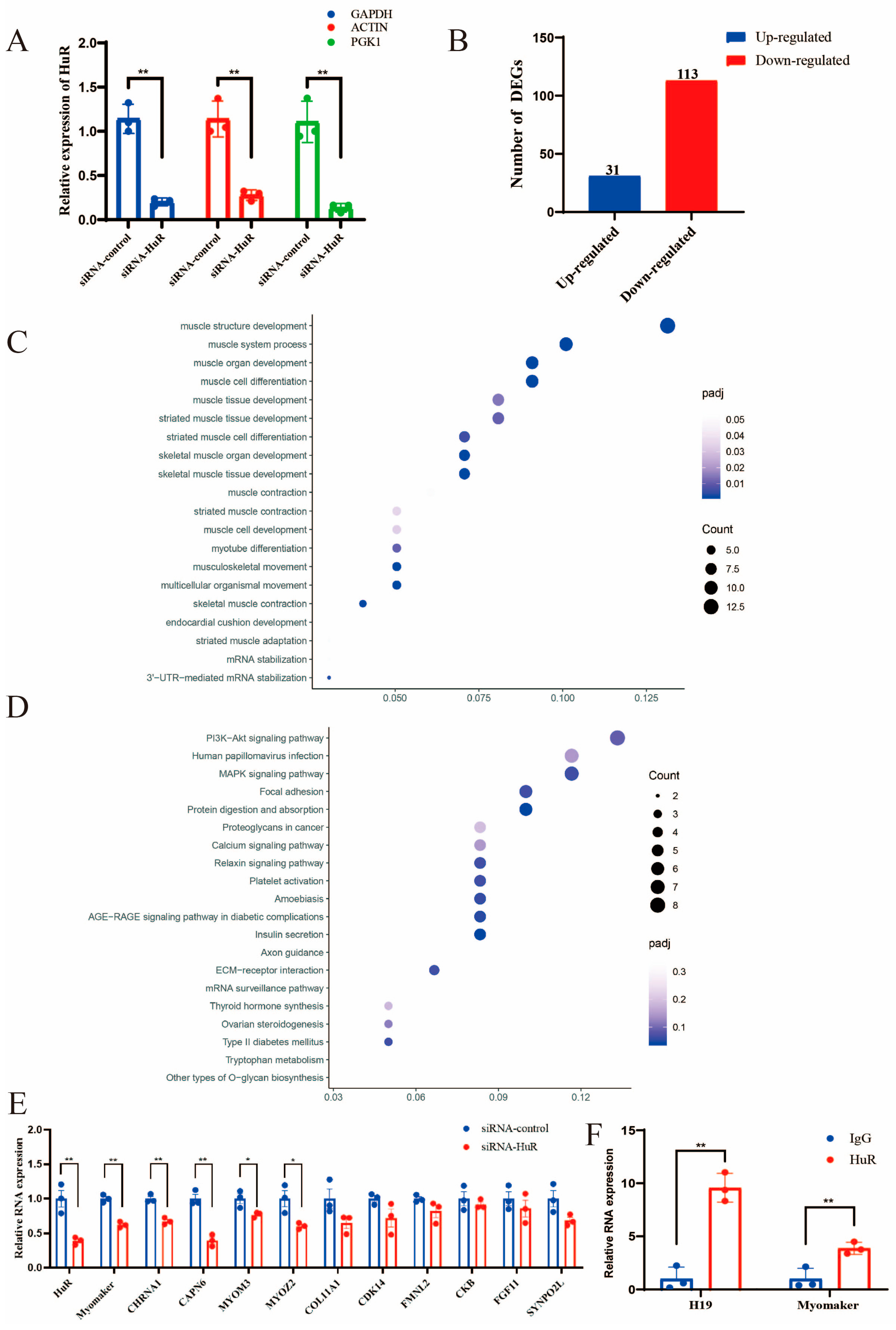
2.3. Identification of HuR Downstream Targets
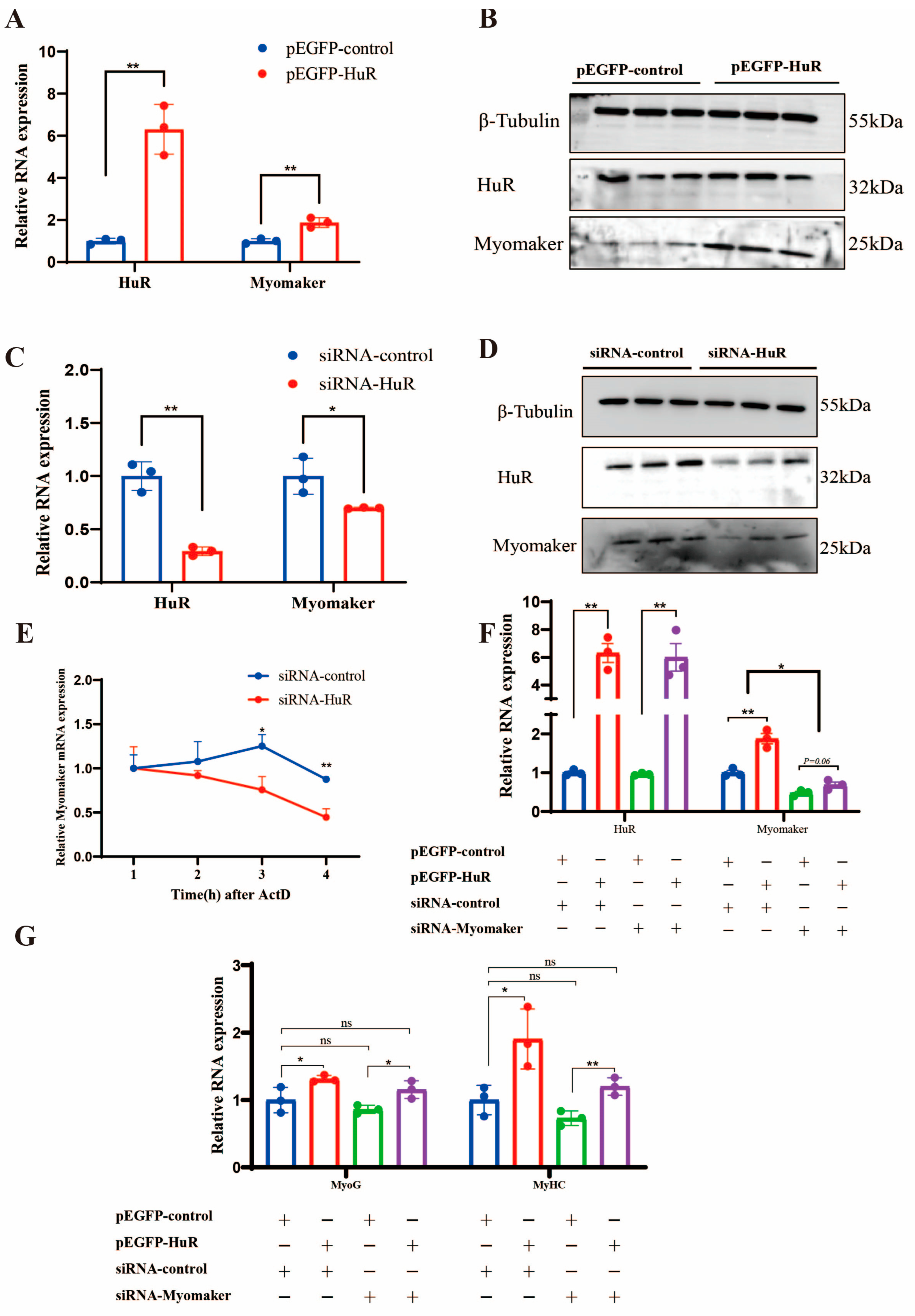
2.4. HuR Promotes the Differentiation of Goat MuSCs by Regulating Myomaker mRNA Stability
3. Discussion
4. Materials and Methods
4.1. Sample Preparation
4.2. Cell Culture and Transfection
4.3. Extraction of Total RNA and qRT-PCR
4.4. Plasmid Construction and siRNAs
4.5. Western Blot Analysis
4.6. Immunofluorescence Analyses
4.7. RNA Stability Analyses
4.8. RNA Immunoprecipitation Assay
4.9. RNA-Seq Analyses
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Khanna, S.; Merriam, A.P.; Gong, B.; Leahy, P.; Porter, J.D. Comprehensive expression profiling by muscle tissue class and identification of the molecular niche of extraocular muscle. FASEB J. 2003, 17, 1370–1372. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; Price, F.; Rudnicki, M.A. Satellite cells and the muscle stem cell niche. Physiol. Rev. 2013, 93, 23–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bentzinger, C.F.; Wang, Y.X.; Rudnicki, M.A. Building muscle: Molecular regulation of myogenesis. Cold Spring Harb. Perspect. Biol. 2012, 4, a008342. [Google Scholar] [CrossRef] [Green Version]
- Yamanouchi, K.; Hosoyama, T.; Murakami, Y.; Nakano, S.; Nishihara, M. Satellite cell differentiation in goat skeletal muscle single fiber culture. J. Reprod. Dev. 2009, 55, 252–255. [Google Scholar] [CrossRef] [PubMed]
- Bhagavati, S.; Song, X.; Siddiqui, M.A. RNAi inhibition of Pax3/7 expression leads to markedly decreased expression of muscle determination genes. Mol. Cell. Biochem. 2007, 302, 257–262. [Google Scholar] [CrossRef]
- Taylor, M.V.; Hughes, S.M. Mef2 and the skeletal muscle differentiation program. Semin. Cell Dev. Biol. 2017, 72, 33–44. [Google Scholar] [CrossRef]
- Ott, M.O.; Bober, E.; Lyons, G.; Arnold, H.; Buckingham, M. Early expression of the myogenic regulatory gene, myf-5, in precursor cells of skeletal muscle in the mouse embryo. Development 1991, 111, 1097–1107. [Google Scholar] [CrossRef]
- Zhan, S.; Qin, C.; Li, D.; Zhao, W.; Nie, L.; Cao, J.; Guo, J.; Zhong, T.; Wang, L.; Li, L.; et al. A Novel Long Noncoding RNA, lncR-125b, Promotes the Differentiation of Goat Skeletal Muscle Satellite Cells by Sponging miR-125b. Front. Genet. 2019, 10, 1171. [Google Scholar] [CrossRef] [Green Version]
- Zhao, W.; Yang, H.; Li, J.; Chen, Y.; Cao, J.; Zhong, T.; Wang, L.; Guo, J.; Li, L.; Zhang, H. MiR-183 promotes preadipocyte differentiation by suppressing Smad4 in goats. Gene 2018, 666, 158–164. [Google Scholar] [CrossRef]
- Li, L.; Chen, Y.; Nie, L.; Ding, X.; Zhang, X.; Zhao, W.; Xu, X.; Kyei, B.; Dai, D.; Zhan, S.; et al. MyoD-induced circular RNA CDR1as promotes myogenic differentiation of skeletal muscle satellite cells. Biochim. Biophys. Acta Gene Regul. Mech. 2019, 1862, 807–821. [Google Scholar] [CrossRef]
- Zhou, G.; Yang, Y.; Zhang, X.; Wang, J. Msx1 cooperates with Runx1 for inhibiting myoblast differentiation. Protein Expr. Purif. 2021, 179, 105797. [Google Scholar] [CrossRef] [PubMed]
- Shi, D.L.; Grifone, R. RNA-Binding Proteins in the Post-transcriptional Control of Skeletal Muscle Development, Regeneration and Disease. Front. Cell Dev. Biol. 2021, 9, 738978. [Google Scholar] [CrossRef] [PubMed]
- Figueroa, A.; Cuadrado, A.; Fan, J.; Atasoy, U.; Muscat, G.E.; Muñoz-Canoves, P.; Gorospe, M.; Muñoz, A. Role of HuR in Skeletal Myogenesis through Coordinate Regulation of Muscle Differentiation Genes. Mol. Cell. Biol. 2003, 23, 4991–5004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van der Giessen, K.; Di-Marco, S.; Clair, E.; Gallouzi, I.E. RNAi-mediated HuR depletion leads to the inhibition of muscle cell differentiation. J. Biol. Chem. 2003, 278, 47119–47128. [Google Scholar] [CrossRef] [Green Version]
- Dormoy-Raclet, V.; Cammas, A.; Celona, B.; Lian, X.J.; Van Der Giessen, K.; Zivojnovic, M.; Brunelli, S.; Riuzzi, F.; Sorci, G.; Wilhelm, B.T.; et al. HuR and miR-1192 regulate myogenesis by modulating the translation of HMGB1 mRNA. Nat. Commun. 2013, 4, 2388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cammas, A.; Sanchez, B.J.; Lian, X.J.; Dormoy-Raclet, V.; Van Der Giessen, K.; Lopez de Silanes, I.; Ma, J.; Wilusz, C.; Richardson, J.; Gorospe, M.; et al. Destabilization of nucleophosmin mRNA by the HuR/KSRP complex is required for muscle fibre formation. Nat. Commun. 2014, 5, 4190. [Google Scholar] [CrossRef] [Green Version]
- Cesana, M.; Cacchiarelli, D.; Legnini, I.; Santini, T.; Sthandier, O.; Chinappi, M.; Tramontano, A.; Bozzoni, I. A long noncoding RNA controls muscle differentiation by functioning as a competing endogenous RNA. Cell 2011, 147, 358–369. [Google Scholar] [CrossRef] [Green Version]
- Legnini, I.; Morlando, M.; Mangiavacchi, A.; Fatica, A.; Bozzoni, I. A feedforward regulatory loop between HuR and the long noncoding RNA linc-MD1 controls early phases of myogenesis. Mol. Cell 2014, 53, 506–514. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.H.; Chang, M.W.; Pandey, P.R.; Tsitsipatis, D.; Yang, X.; Martindale, J.L.; Munk, R.; De, S.; Abdelmohsen, K.; Gorospe, M. Interaction of OIP5-AS1 with MEF2C mRNA promotes myogenic gene expression. Nucleic Acids Res. 2020, 48, 12943–12956. [Google Scholar] [CrossRef]
- Mynatt, R.L.; Noland, R.C.; Elks, C.M.; Vandanmagsar, B.; Bayless, D.S.; Stone, A.C.; Ghosh, S.; Ravussin, E.; Warfel, J.D. The RNA binding protein HuR influences skeletal muscle metabolic flexibility in rodents and humans. Metabolism 2019, 97, 40–49. [Google Scholar] [CrossRef]
- Mubaid, S.; Ma, J.F.; Omer, A.; Ashour, K.; Lian, X.J.; Sanchez, B.J.; Robinson, S.; Cammas, A.; Dormoy-Raclet, V.; Di Marco, S.; et al. HuR counteracts miR-330 to promote STAT3 translation during inflammation-induced muscle wasting. Proc. Natl. Acad. Sci. USA 2019, 116, 17261–17270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanchez, B.J.; Mubaid, S.; Busque, S.; de Los Santos, Y.L.; Ashour, K.; Sadek, J.; Lian, X.J.; Khattak, S.; Di Marco, S.; Gallouzi, I.E. The formation of HuR/YB1 complex is required for the stabilization of target mRNA to promote myogenesis. Nucleic Acids Res. 2023, 51, 1375–1392. [Google Scholar] [CrossRef] [PubMed]
- Beauchamp, P.; Nassif, C.; Hillock, S.; Van Der Giessen, K.; Von Roretz, C.; Jasmin, B.J.; Gallouzi, I.E. The cleavage of HuR interferes with its transportin-2-mediated nuclear import and promotes muscle fiber formation. Cell Death Differ. 2010, 17, 1588–1599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lai, W.S.; Arvola, R.M.; Goldstrohm, A.C.; Blackshear, P.J. Inhibiting transcription in cultured metazoan cells with actinomycin D to monitor mRNA turnover. Methods 2019, 155, 77–87. [Google Scholar] [CrossRef]
- Millay, D.P.; O’Rourke, J.R.; Sutherland, L.B.; Bezprozvannaya, S.; Shelton, J.M.; Bassel-Duby, R.; Olson, E.N. Myomaker is a membrane activator of myoblast fusion and muscle formation. Nature 2013, 499, 301–305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Q.; Vashisht, A.A.; O’Rourke, J.; Corbel, S.Y.; Moran, R.; Romero, A.; Miraglia, L.; Zhang, J.; Durrant, E.; Schmedt, C.; et al. The microprotein Minion controls cell fusion and muscle formation. Nat. Commun. 2017, 8, 15664. [Google Scholar] [CrossRef] [Green Version]
- Quinn, M.E.; Goh, Q.; Kurosaka, M.; Gamage, D.G.; Petrany, M.J.; Prasad, V.; Millay, D.P. Myomerger induces fusion of non-fusogenic cells and is required for skeletal muscle development. Nat. Commun. 2017, 8, 15665. [Google Scholar] [CrossRef] [Green Version]
- Lv, W.; Jin, J.; Xu, Z.; Luo, H.; Guo, Y.; Wang, X.; Wang, S.; Zhang, J.; Zuo, H.; Bai, W.; et al. lncMGPF is a novel positive regulator of muscle growth and regeneration. J. Cachexia Sarcopenia Muscle 2020, 11, 1723–1746. [Google Scholar] [CrossRef]
- Zhou, A.; Shi, G.; Kang, G.J.; Xie, A.; Liu, H.; Jiang, N.; Liu, M.; Jeong, E.M.; Dudley Jr, S.C. RNA Binding Protein, HuR, Regulates SCN5A Expression Through Stabilizing MEF2C transcription factor mRNA. J. Am. Heart Assoc. 2018, 7, e007802. [Google Scholar] [CrossRef]
- Kim, H.H.; Kuwano, Y.; Srikantan, S.; Lee, E.K.; Martindale, J.L.; Gorospe, M. HuR recruits let-7/RISC to repress c-Myc expression. Genes Dev. 2009, 23, 1743–1748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poria, D.K.; Guha, A.; Nandi, I.; Ray, P.S. RNA-binding protein HuR sequesters microRNA-21 to prevent translation repression of proinflammatory tumor suppressor gene programmed cell death 4. Oncogene 2016, 35, 1703–1715. [Google Scholar] [CrossRef] [Green Version]
- Sun, Q.; Tripathi, V.; Yoon, J.H.; Singh, D.K.; Hao, Q.; Min, K.W.; Davila, S.; Zealy, R.W.; Li, X.L.; Polycarpou-Schwarz, M.; et al. MIR100 host gene-encoded lncRNAs regulate cell cycle by modulating the interaction between HuR and its target mRNAs. Nucleic Acids Res. 2018, 46, 10405–10416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.; Li, F.; Zhang, H.; Hu, X.; Chen, Y.; Huang, C. RAB5C, a new mRNA binding target of HuR, regulates breast cancer cell proliferation. Cell Biol. Int. 2023, 47, 374–382. [Google Scholar] [CrossRef]
- Duan, X.H.; Chen, R.; Li, D.S.; Luo, A.H.; Guo, L.L. HuR affects chemoresistance of small cell lung cancer by regulating FGFRL1 expression. Exp. Ther. Med. 2022, 24, 1–6. [Google Scholar] [CrossRef] [PubMed]
- van der Giessen, K.; Gallouzi, I.E. Involvement of transportin 2-mediated HuR import in muscle cell differentiation. Mol. Biol. Cell 2007, 18, 2619–2629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abmayr, S.M.; Balagopalan, L.; Galletta, B.J.; Hong, S.J. Cell and molecular biology of myoblast fusion. Int. Rev. Cytol. 2003, 225, 33–89. [Google Scholar] [CrossRef] [PubMed]
- Luo, W.; Li, E.; Nie, Q.; Zhang, X. Myomaker, Regulated by MYOD, MYOG and miR-140-3p, Promotes Chicken Myoblast Fusion. Int. J. Mol. Sci. 2015, 16, 26186–26201. [Google Scholar] [CrossRef] [Green Version]
- Zheng, S.; Li, L.; Zhou, H.; Zhang, X.; Xu, X.; Dai, D.; Zhan, S.; Cao, J.; Guo, J.; Zhong, T.; et al. CircTCF4 Suppresses Proliferation and Differentiation of Goat Skeletal Muscle Satellite Cells Independent from AGO2 Binding. Int. J. Mol. Sci. 2022, 23, 12868. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Qiu, L.Q.; Lai, W.S.; Stumpo, D.J.; Blackshear, P.J. Measurement of mRNA Decay in Mouse Embryonic Fibroblasts. Bio-protocol 2016, 6, e1858. [Google Scholar] [CrossRef] [Green Version]
- Song, T.; Yang, Y.; Wei, H.; Xie, X.; Lu, J.; Zeng, Q.; Peng, J.; Zhou, Y.; Jiang, S.; Peng, J. Zfp217 mediates m6A mRNA methylation to orchestrate transcriptional and post-transcriptional regulation to promote adipogenic differentiation. Nucleic Acids Res. 2019, 47, 6130–6144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghosh, M.; Aguila, H.L.; Michaud, J.; Ai, Y.; Wu, M.T.; Hemmes, A.; Ristimaki, A.; Guo, C.; Furneaux, H.; Hla, T. Essential role of the RNA-binding protein HuR in progenitor cell survival in mice. J. Clin. Investig. 2009, 119, 3530–3543. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, Y.; Zhan, S.; Zhao, S.; Zhong, T.; Wang, L.; Guo, J.; Dai, D.; Li, D.; Cao, J.; Li, L.; et al. HuR Promotes the Differentiation of Goat Skeletal Muscle Satellite Cells by Regulating Myomaker mRNA Stability. Int. J. Mol. Sci. 2023, 24, 6893. https://doi.org/10.3390/ijms24086893
Sun Y, Zhan S, Zhao S, Zhong T, Wang L, Guo J, Dai D, Li D, Cao J, Li L, et al. HuR Promotes the Differentiation of Goat Skeletal Muscle Satellite Cells by Regulating Myomaker mRNA Stability. International Journal of Molecular Sciences. 2023; 24(8):6893. https://doi.org/10.3390/ijms24086893
Chicago/Turabian StyleSun, Yanjin, Siyuan Zhan, Sen Zhao, Tao Zhong, Linjie Wang, Jiazhong Guo, Dinghui Dai, Dandan Li, Jiaxue Cao, Li Li, and et al. 2023. "HuR Promotes the Differentiation of Goat Skeletal Muscle Satellite Cells by Regulating Myomaker mRNA Stability" International Journal of Molecular Sciences 24, no. 8: 6893. https://doi.org/10.3390/ijms24086893