
The Role of Tyr-His-Trp Triad and Water Molecule Near the N1-Atom of 2-Hydroperoxycoelenterazine in Bioluminescence of Hydromedusan Photoproteins: Structural and Mutagenesis Study
 , , , , and
, , , , and
Abstract
:1. Introduction

2. Results
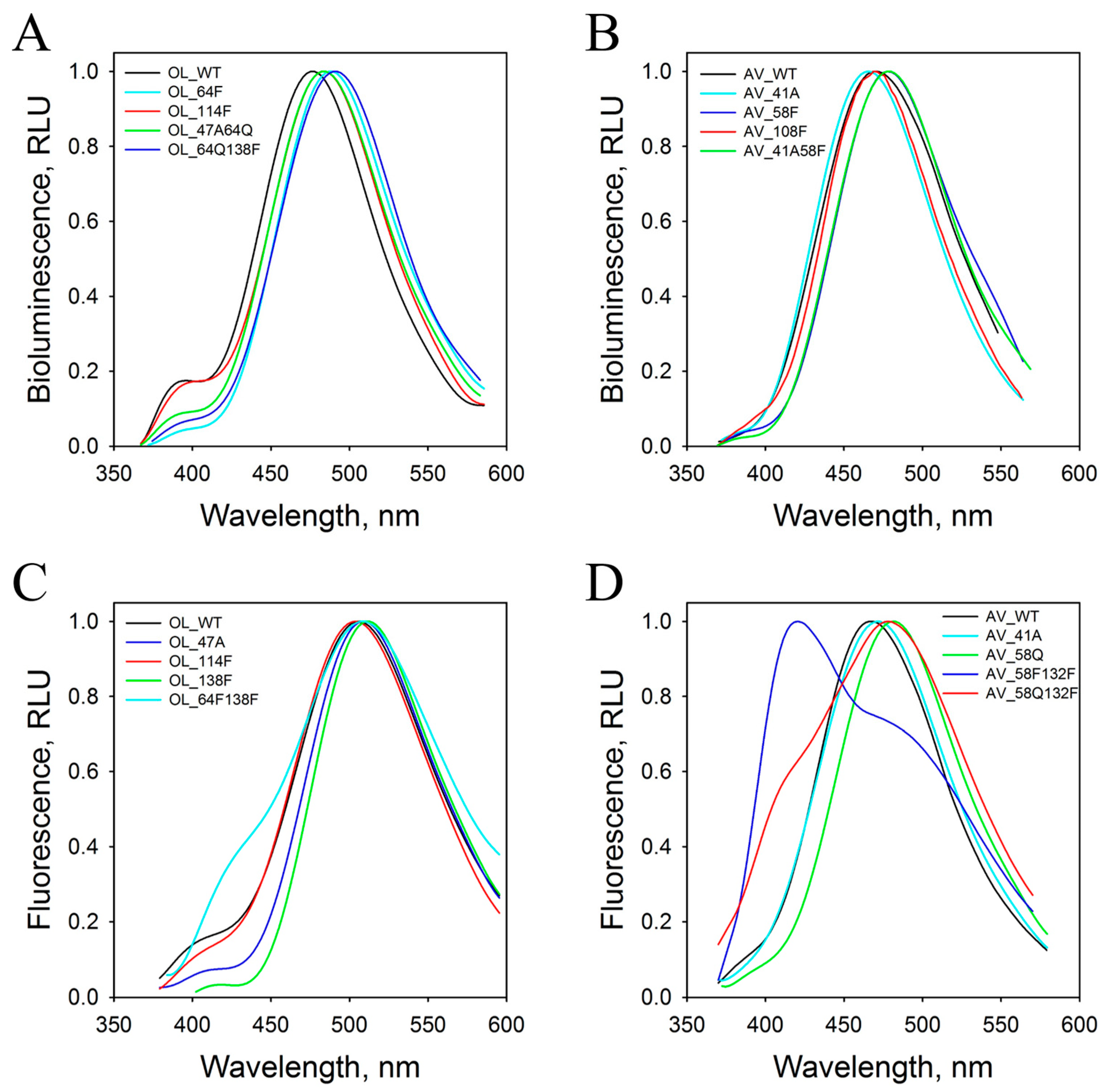
2.1. Bioluminescence Activity and Spectral Properties
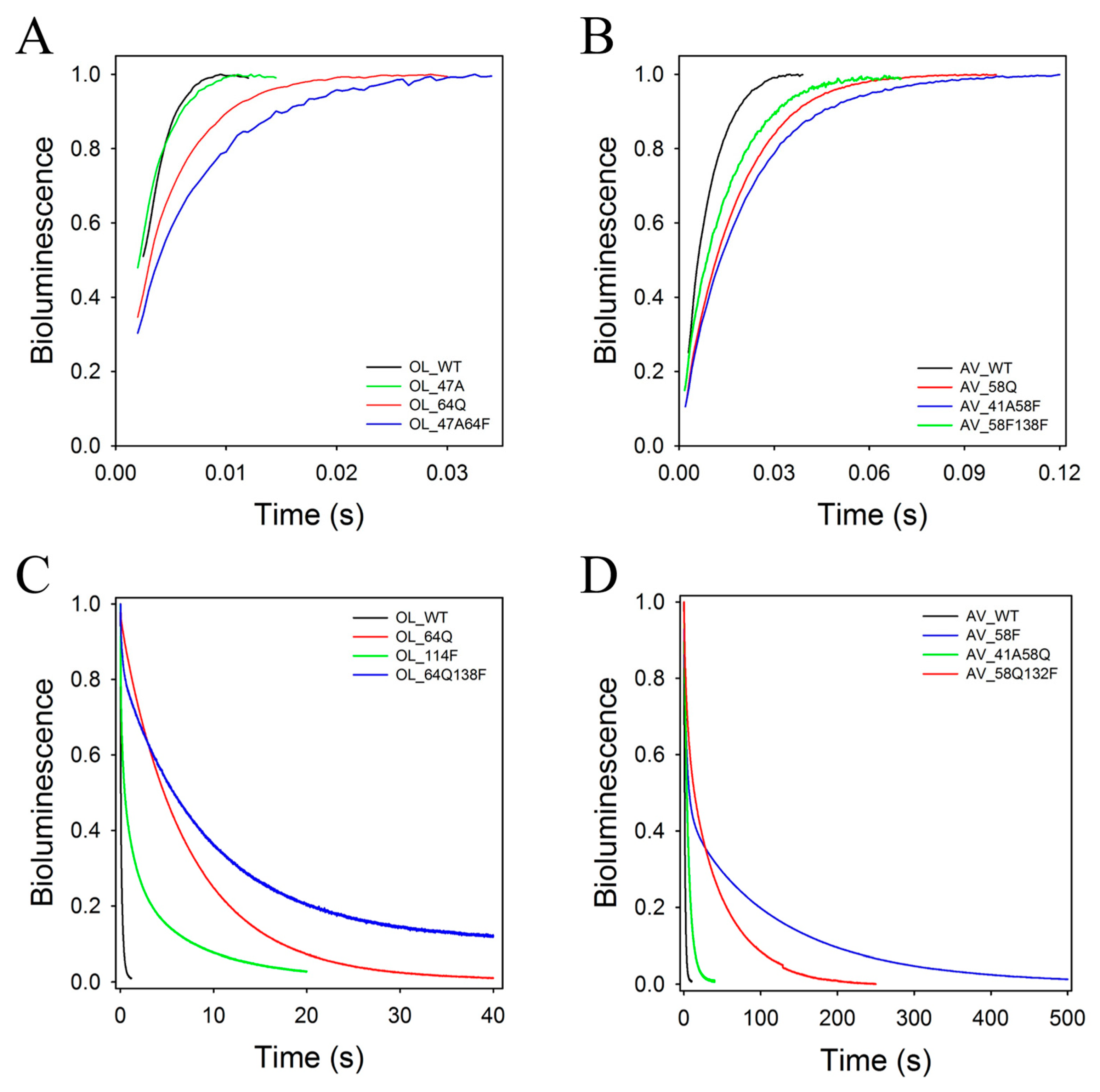
2.2. Rapid-Mixing Stopped-Flow Kinetics

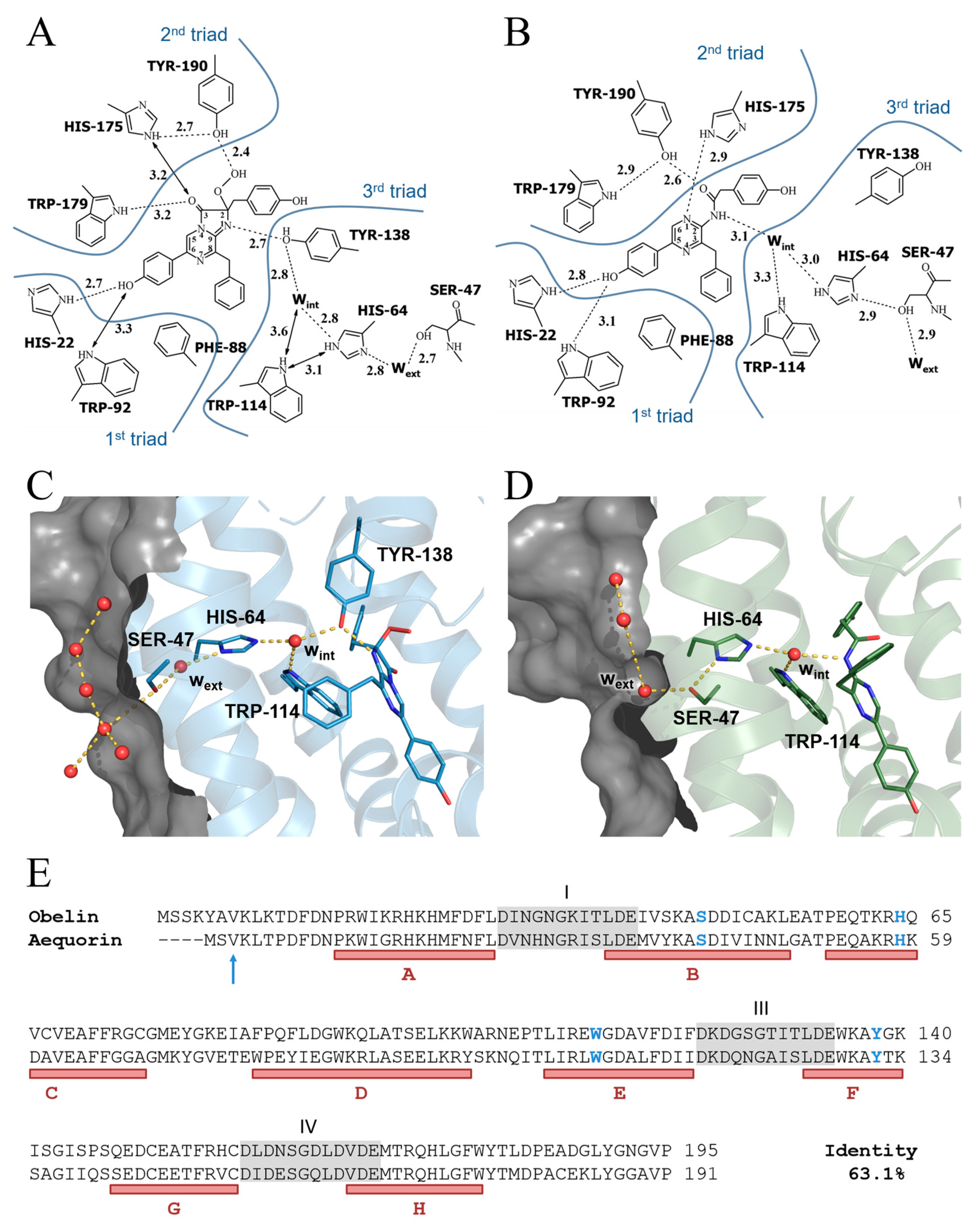
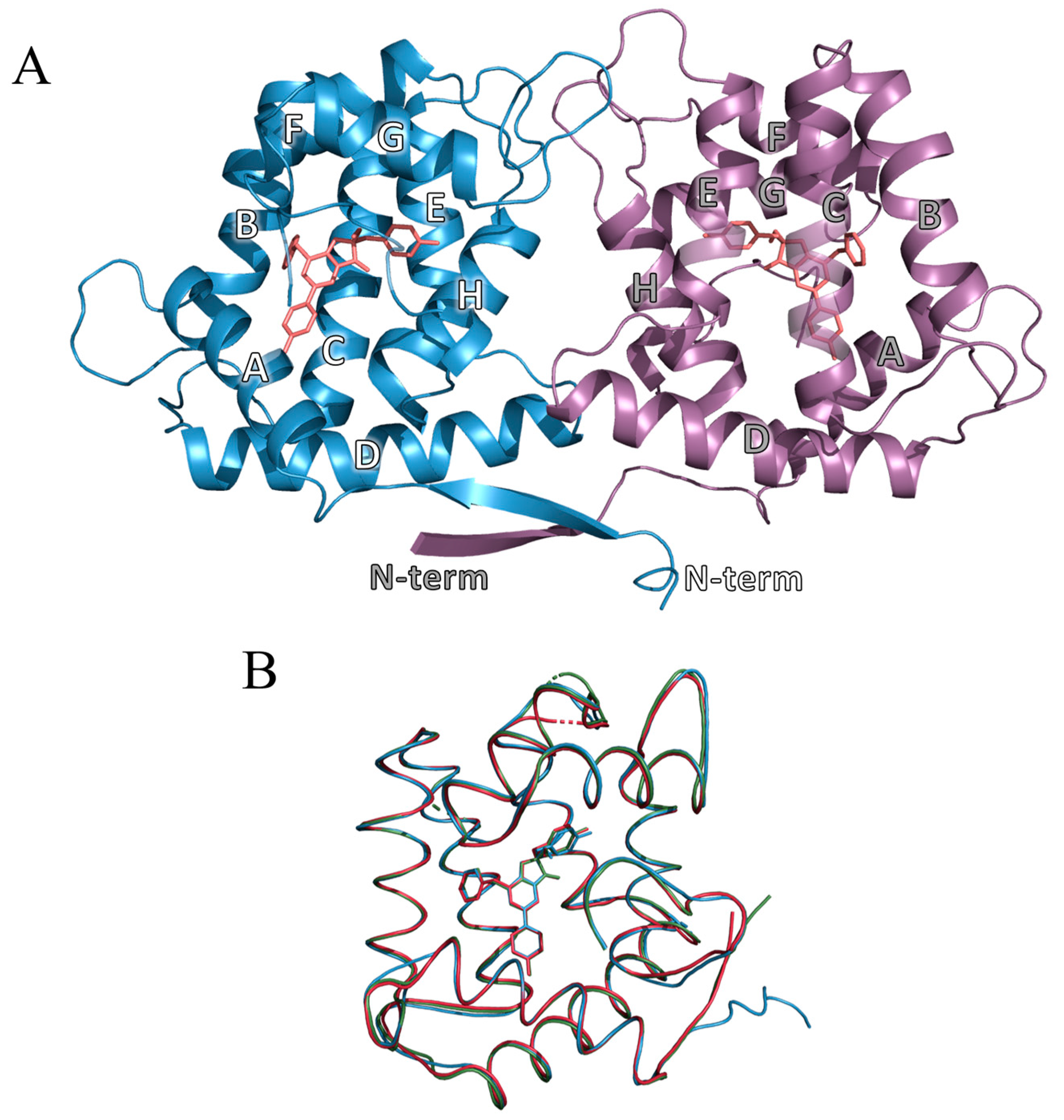
2.3. Overall Structure of OL_H64F Mutant
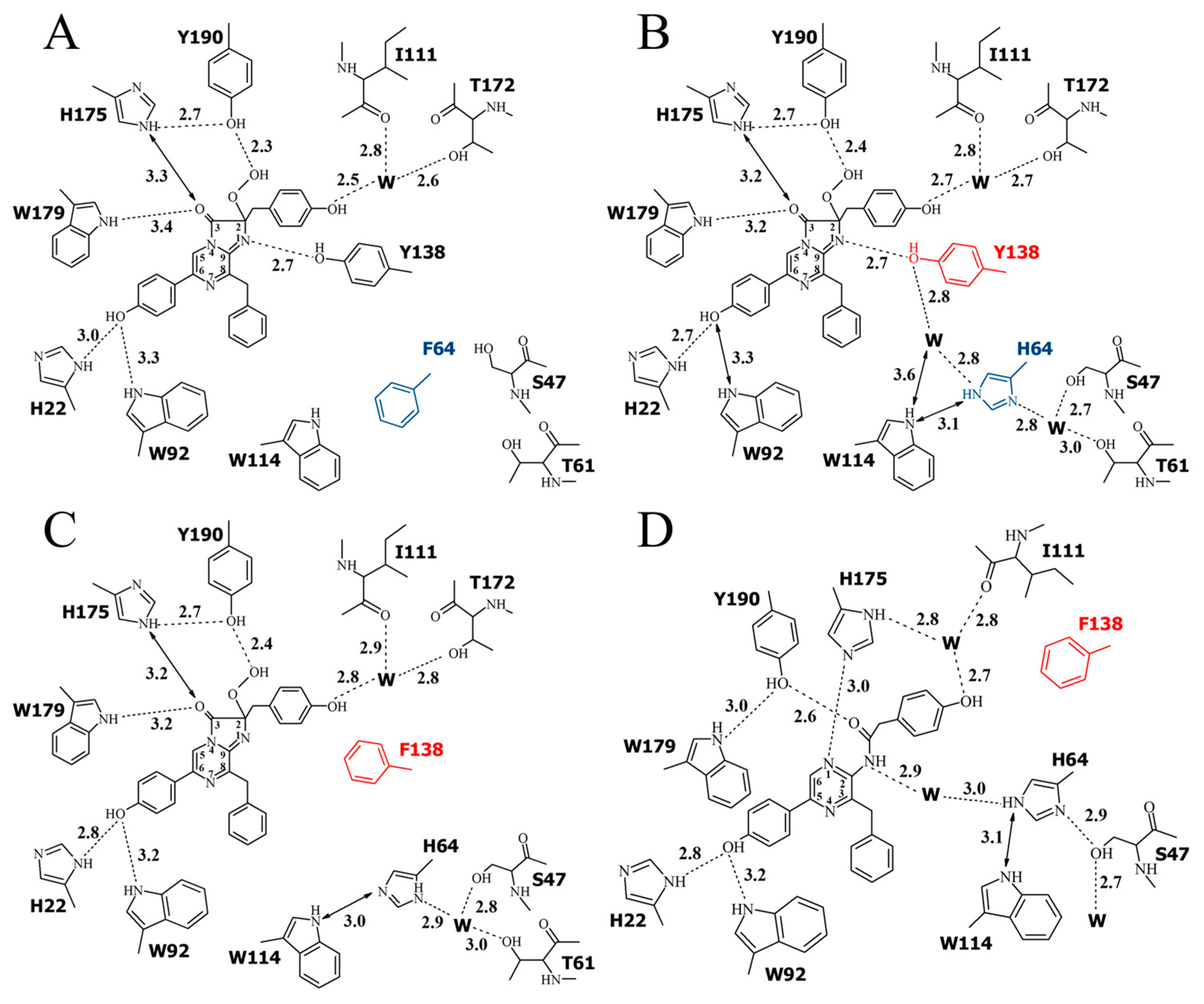
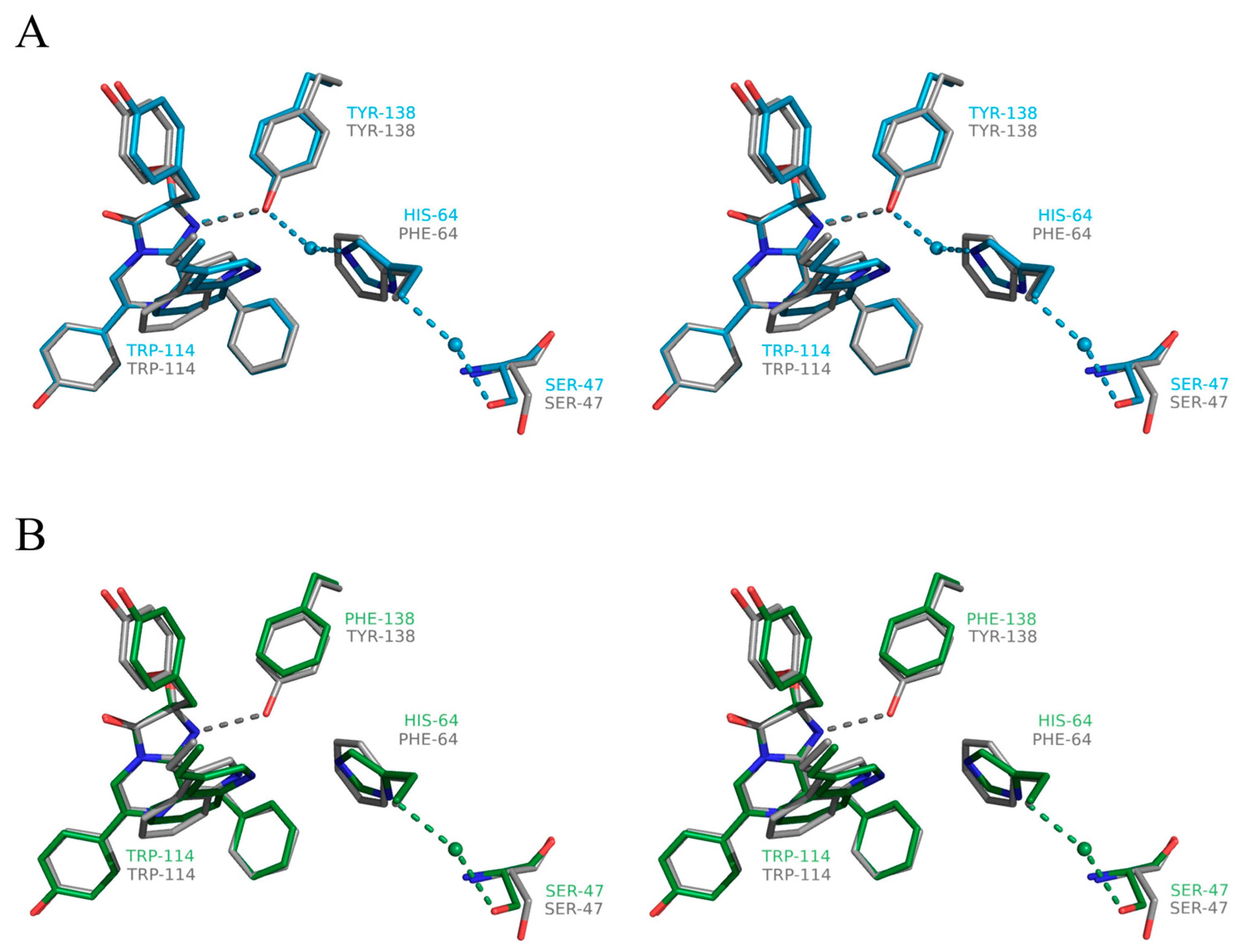
2.4. Structure of the Substrate-Binding Cavity of OL_H64F Mutant


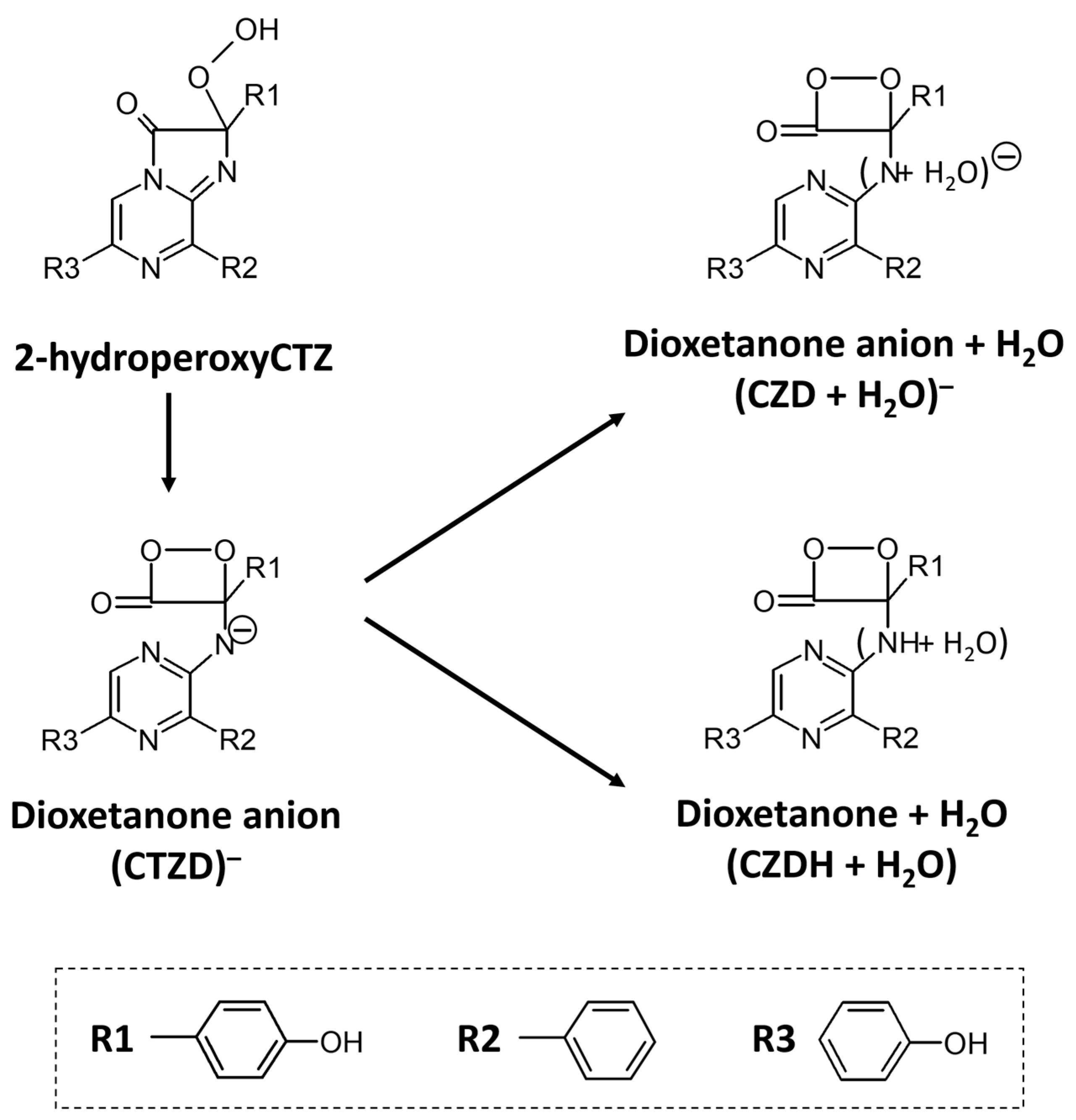
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Site-Directed Mutagenesis and Preparation of the Photoprotein Samples
4.3. Crystallization, Data Collection, Structure Solution, and Crystallographic Refinement
4.4. Bioluminescence Assay
4.5. Spectral Measurements
4.6. Rapid-Mixing Kinetics Measurements
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shimomura, O. Bioluminescence: Chemical Principles and Methods; World Scientific Publishing Co. Pte. Ltd.: Singapore, 2006. [Google Scholar]
- Vysotski, E.S.; Markova, S.V.; Frank, L.A. Calcium-regulated photoproteins of marine coelenterates. Mol. Biol. 2006, 40, 355–367. [Google Scholar] [CrossRef]
- Shimomura, O.; Johnson, F.H. Structure of the light-emitting moiety of aequorin. Biochemistry 1972, 11, 1602–1608. [Google Scholar] [CrossRef]
- Cormier, M.J.; Hori, K.; Karkhanis, Y.D.; Anderson, J.M.; Wampler, J.M.; Morin, J.G.; Hastings, J.W. Evidence for similar biochemical requirements for bioluminescence among the coelenterate. J. Cell. Physiol. 1973, 81, 291–297. [Google Scholar] [CrossRef]
- Vysotski, E.S.; Lee, J. Ca2+-regulated photoproteins: Structural insight into the bioluminescence mechanism. Acc. Chem. Res. 2004, 37, 405–415. [Google Scholar] [CrossRef]
- Prasher, D.; McCann, R.O.; Cormier, M.J. Cloning and expression of the cDNA coding for aequorin, a bioluminescent calcium-binding protein. Biochem. Biophys. Res. Commun. 1985, 126, 1259–1268. [Google Scholar] [CrossRef] [PubMed]
- Prasher, D.; McCann, R.O.; Longiaru, M.; Cormier, M.J. Sequence comparisons of complementary DNAs encoding aequorin isotypes. Biochemistry 1987, 26, 1326–1332. [Google Scholar] [CrossRef]
- Inouye, S.; Noguchi, M.; Sakaki, Y.; Takagi, Y.; Miyata, T.; Iwanaga, S.; Miyata, T.; Tsuji, F.I. Cloning and sequence analysis of cDNA for the luminescent protein aequorin. Proc. Natl. Acad. Sci. USA 1985, 82, 3154–3158. [Google Scholar] [CrossRef] [Green Version]
- Inouye, S.; Tsuji, F.I. Cloning and sequence analysis of cDNA for the Ca2+-activated photoprotein, clytin. FEBS Lett. 1993, 315, 343–346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inouye, S. Cloning, expression, purification and characterization of an isotype of clytin, a calcium-binding photoprotein from the luminous hydromedusa Clytia gregarium. J. Biochem. 2008, 143, 711–717. [Google Scholar] [CrossRef] [PubMed]
- Markova, S.V.; Burakova, L.P.; Frank, L.A.; Golz, S.; Korostileva, K.A.; Vysotski, E.S. Green-fluorescent protein from the bioluminescent jellyfish Clytia gregaria: cDNA cloning, expression, and characterization of novel recombinant protein. Photochem. Photobiol. Sci. 2010, 9, 757–765. [Google Scholar] [CrossRef] [PubMed]
- Fourrage, C.; Swann, K.; Gonzalez Garcia, J.R.; Campbell, A.K.; Houliston, E. An endogenous green fluorescent protein-photoprotein pair in Clytia hemisphaerica eggs shows co-targeting to mitochondria and efficient bioluminescence energy transfer. Open Biol. 2014, 4, 130206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fagan, T.F.; Ohmiya, Y.; Blinks, J.R.; Inouye, S.; Tsuji, F.I. Cloning, expression and sequence analysis of cDNA for the Ca2+-binding photoprotein, mitrocomin. FEBS Lett. 1993, 333, 301–305. [Google Scholar] [CrossRef] [Green Version]
- Burakova, L.P.; Natashin, P.V.; Markova, S.V.; Eremeeva, E.V.; Malikova, N.P.; Cheng, C.; Liu, Z.J.; Vysotski, E.S. Mitrocomin from the jellyfish Mitrocoma cellularia with deleted C-terminal tyrosine reveals a higher bioluminescence activity compared to wild type photoprotein. J. Photochem. Photobiol. 2016, 162, 286–297. [Google Scholar] [CrossRef]
- Illarionov, B.A.; Markova, S.V.; Bondar, V.S.; Vysotski, E.S.; Gitelson, J.I. Cloning and expression of cDNA for the Ca2+-activated photoprotein obelin from the hydroid polyp Obelia longissima. Dokl. Akad. Nauk 1992, 326, 911–913. [Google Scholar]
- Illarionov, B.A.; Bondar, V.S.; Illarionova, V.A.; Vysotski, E.S. Sequence of the cDNA encoding the Ca2+-activated photoprotein obelin from the hydroid polyp Obelia longissima. Gene 1995, 153, 273–274. [Google Scholar] [CrossRef] [PubMed]
- Markova, S.V.; Vysotski, E.S.; Blinks, J.R.; Burakova, L.P.; Wang, B.C.; Lee, J. Obelin from the bioluminescent marine hydroid Obelia geniculata: Cloning, expression, and comparison of some properties with those of other Ca2+-regulated photoproteins. Biochemistry 2002, 41, 2227–2236. [Google Scholar] [CrossRef] [PubMed]
- Ohmiya, Y.; Hirano, T. Shining the light: The mechanism of the bioluminescence reaction of calcium-binding photoproteins. Chem. Biol. 1996, 3, 337–347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawasaki, H.; Nakayama, S.; Kretsinger, R.H. Classification and evolution of EF-hand proteins. Biometals 1998, 11, 277–295. [Google Scholar] [CrossRef] [PubMed]
- Shimomura, O.; Johnson, F.H. Regeneration of the photoprotein aequorin. Nature 1975, 256, 236–238. [Google Scholar] [CrossRef] [PubMed]
- Eremeeva, E.V.; Natashin, P.V.; Song, L.; Zhou, Y.; van Berkel, W.J.; Liu, Z.J.; Vysotski, E.S. Oxygen activation of apo-obelin-coelenterazine complex. Chembiochem 2013, 14, 739–745. [Google Scholar] [CrossRef]
- Head, J.F.; Inouye, S.; Teranishi, K.; Shimomura, O. The crystal structure of the photoprotein aequorin at 2.3 Å resolution. Nature 2000, 405, 372–376. [Google Scholar] [CrossRef]
- Liu, Z.J.; Vysotski, E.S.; Chen, C.J.; Rose, J.P.; Lee, J.; Wang, B.C. Structure of the Ca2+-regulated photoprotein obelin at 1.7 Å resolution determined directly from ist sulfur substructure. Protein Sci. 2000, 9, 2085–2093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Z.J.; Vysotski, E.S.; Deng, L.; Lee, J.; Rose, J.; Wang, B.C. Atomic resolution structure of obelin: Soaking with calcium enhances electron density of the second oxygen atom substituted at the C2-position of coelenterazine. Biochem. Biophys. Res. Commun. 2003, 311, 433–439. [Google Scholar] [CrossRef]
- Titushin, M.S.; Feng, Y.; Stepanyuk, G.A.; Li, Y.; Markova, S.V.; Golz, S.; Wang, B.C.; Lee, J.; Wang, J.; Vysotski, E.S.; et al. NMR-derived topology of a GFP-photoprotein energy transfer complex. J. Biol. Chem. 2010, 285, 40891–40900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vysotski, E.S.; Lee, J. Bioluminescent mechanism of Ca2+-regulated photoproteins from three-dimensional structures. In Luciferases and Fluorescent proteins: Principles and Advances in Biotechnology and Bioimaging; Viviani, V.R., Ohmiya, Y., Eds.; Transworld Research Network: Kerala, India, 2007; pp. 19–41. [Google Scholar]
- Ohmiya, Y.; Ohashi, M.; Tsuji, F.I. Two excited states in aequorin bioluminescence induced by tryptophan modification. FEBS Lett. 1992, 301, 197–201. [Google Scholar] [CrossRef] [Green Version]
- Deng, L.; Vysotski, E.S.; Liu, Z.J.; Markova, S.V.; Malikova, N.P.; Lee, J.; Rose, J.; Wang, B.C. Structural basis for the emission of violet bioluminescence from a W92F obelin mutant. FEBS Lett. 2001, 506, 281–285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vysotski, E.S.; Liu, Z.J.; Markova, S.V.; Blinks, J.R.; Deng, L.; Frank, L.A.; Herko, M.; Malikova, N.P.; Rose, J.P.; Wang, B.C.; et al. Violet bioluminescence and fast kinetics from W92F obelin: Structure-based proposals for the bioluminescence triggering and the identification of the emitting species. Biochemistry 2003, 42, 6013–6024. [Google Scholar] [CrossRef] [PubMed]
- Malikova, N.P.; Stepanyuk, G.A.; Frank, L.A.; Markova, S.V.; Vysotski, E.S.; Lee, J. Spectral tuning of obelin bioluminescence by mutations of Trp92. FEBS Lett. 2003, 554, 184–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stepanyuk, G.A.; Golz, S.; Markova, S.V.; Frank, L.A.; Lee, J.; Vysotski, E.S. Interchange of aequorin and obelin bioluminescence color is determined by substitution of one active site residue of each photoprotein. FEBS Lett. 2005, 579, 1008–1014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Natashin, P.V.; Markova, S.V.; Lee, J.; Vysotski, E.S.; Liu, Z.J. Crystal structures of the F88Y obelin mutant before and after bioluminescence provide molecular insight into spectral tuning among hydromedusan photoproteins. FEBS J. 2014, 281, 1432–1445. [Google Scholar] [CrossRef] [PubMed]
- Gao, M.; Ding, B.W.; Liu, Y.J. Tuning the fluorescence of calcium-discharged photoprotein obelin via mutating at the His22-Phe88-Trp92 triad—A QM/MM study. Photochem. Photobiol. Sci. 2019, 18, 1823–1832. [Google Scholar] [CrossRef]
- Eremeeva, E.V.; Markova, S.V.; Frank, L.A.; Visser, A.J.; van Berkel, W.J.; Vysotski, E.S. Bioluminescent and spectroscopic properties of His-Trp-Tyr triad mutants of obelin and aequorin. Photochem. Photobiol. Sci. 2013, 12, 1016–1024. [Google Scholar] [CrossRef]
- Eremeeva, E.V.; Markova, S.V.; van Berkel, W.J.; Vysotski, E.S. Role of key residues of obelin in coelenterazine binding and conversion into 2-hydroperoxy adduct. J. Photochem. Photobiol. 2013, 127, 133–139. [Google Scholar] [CrossRef]
- Liu, Z.J.; Stepanyuk, G.A.; Vysotski, E.S.; Lee, J.; Markova, S.V.; Malikova, N.P.; Wang, B.C. Crystal structure of obelin after Ca2+-triggered bioluminescence suggests neutral coelenteramide as the primary excited state. Proc. Natl. Acad. Sci. USA 2006, 103, 2570–2575. [Google Scholar] [CrossRef] [Green Version]
- Markova, S.V.; Vysotski, E.S.; Lee, J. Obelin hyperexpression in E. coli, purification and characterization. In Bioluminescence and Chemiluminescence; Case, J.F., Herring, P.J., Robison, B.H., Haddock, S.H.D., Kricka, L.J., Stanley, P.E., Eds.; World Scientific Publishing Co.: Singapore, 2001; pp. 115–119. [Google Scholar]
- Deng, L.; Vysotski, E.S.; Markova, S.V.; Liu, Z.J.; Lee, J.; Rose, J.; Wang, B.C. All three Ca2+-binding loops of photoproteins bind calcium ions: The crystal structures of calcium-loaded apo-aequorin and apo-obelin. Protein Sci. 2005, 14, 663–675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Natashin, P.V.; Ding, W.; Eremeeva, E.V.; Markova, S.V.; Lee, J.; Vysotski, E.S.; Liu, Z.J. Structures of the Ca2+-regulated photoprotein obelin Y138F mutant before and after bioluminescence support the catalytic function of a water molecule in the reaction. Acta Cryst. Biol. Crystallogr. 2014, 70, 720–732. [Google Scholar] [CrossRef] [PubMed]
- Allen, D.G.; Blinks, J.R.; Prendergast, F.G. Aequorin luminescence: Relation of light emission to calcium concentration—A calcium-independent component. Science 1977, 195, 996–998. [Google Scholar] [CrossRef] [PubMed]
- Eremeeva, E.V.; Vysotski, E.S. Exploring bioluminescence function of the Ca2+-regulated photoproteins with site-directed mutagenesis. Photochem. Photobiol. 2019, 95, 8–23. [Google Scholar] [CrossRef] [Green Version]
- Burakova, L.P.; Vysotski, E.S. Recombinant Ca2+-regulated photoproteins of ctenophores: Current knowledge and application prospects. Appl. Microbiol. Biotechnol. 2019, 103, 5929–5946. [Google Scholar] [CrossRef] [PubMed]
- Tomilin, F.N.; Rogova, A.V.; Burakova, L.P.; Tchaikovskaya, O.N.; Avramov, P.V.; Fedorov, D.G.; Vysotski, E.S. Unusual shift in the visible absorption spectrum of an active ctenophore photoprotein elucidated by time-dependent density functional theory. Photochem. Photobiol. Sci. 2021, 20, 559–570. [Google Scholar] [CrossRef]
- Shimomura, O.; Teranishi, K. Light-emitters involved in the luminescence of coelenterazine. Luminescence 2000, 15, 51–58. [Google Scholar] [CrossRef]
- Malikova, N.P.; Eremeeva, E.V.; Gulnov, D.V.; Natashin, P.V.; Nemtseva, E.V.; Vysotski, E.S. Specific Activities of Hydromedusan Ca2+-Regulated Photoproteins. Photochem. Photobiol. 2022, 98, 275–283. [Google Scholar] [CrossRef]
- Malikova, N.P.; Burakova, L.P.; Markova, S.V.; Vysotski, E.S. Characterization of hydromedusan Ca2+-regulated photoproteins as a tool for measurement of Ca2+ concentration. Anal. Bioanal. Chem. 2014, 406, 5715–5726. [Google Scholar] [CrossRef]
- Hastings, J.W.; Mitchell, G.; Mattingly, P.H.; Blinks, J.R.; Van Leeuwen, M. Response of aequorin bioluminescence to rapid changes in calcium concentration. Nature 1969, 222, 1047–1050. [Google Scholar] [CrossRef]
- Stephenson, D.G.; Sutherland, P.J. Studies on the luminescent response of the Ca2+-activated photoprotein, obelin. Biochim. Biophys. Acta 1981, 678, 65–75. [Google Scholar] [CrossRef] [PubMed]
- Eremeeva, E.V.; Bartsev, S.I.; van Berkel, W.J.; Vysotski, E.S. Unanimous model for describing the fast bioluminescence kinetics of Ca2+-regulated photoproteins of different organisms. Photochem. Photobiol. 2017, 93, 495–502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, L.; Markova, S.V.; Vysotski, E.S.; Liu, Z.J.; Lee, J.; Rose, J.; Wang, B.C. Crystal structure of a Ca2+-discharged photoprotein: Implications for mechanisms of the calcium trigger and bioluminescence. J. Biol. Chem. 2004, 279, 33647–33652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Natashin, P.V.; Eremeeva, E.V.; Shevtsov, M.B.; Kovaleva, M.I.; Bukhdruker, S.S.; Dmitrieva, D.A.; Gulnov, D.V.; Nemtseva, E.V.; Gordeliy, V.I.; Mishin, A.V.; et al. Crystal structure of semi-synthetic obelin-v after calcium induced bioluminescence implies coelenteramine as the main reaction product. Sci. Rep. 2022, 12, 19613. [Google Scholar] [CrossRef] [PubMed]
- Eremeeva, E.V.; Burakova, L.P.; Krasitskaya, V.V.; Kudryavtsev, A.N.; Shimomura, O.; Frank, L.A. Hydrogen-bond networks between the C-terminus and Arg from the first α-helix stabilize photoprotein molecules. Photochem. Photobiol. Sci. 2014, 13, 541–547. [Google Scholar] [CrossRef]
- McCapra, F.; Chang, Y.C. Chemiluminescence of a Cypridina luciferin analogue. Chem. Commun. 1967, 19, 1011–1012. [Google Scholar] [CrossRef]
- Hori, K.; Wampler, J.E.; Matthews, J.C.; Cormier, M.J. Identification of the product excited states during the chemiluminescent and bioluminescent oxidation of Renilla (sea pansy) luciferin and certain of its analogues. Biochemistry 1973, 12, 4463–4468. [Google Scholar] [CrossRef]
- Imai, Y.; Shibata, T.; Maki, S.; Niwa, H.; Ohashi, M.; Hirano, T. Fluorescence properties of phenolate anions of coelenteramide analogues: The light-emitter structure in aequorin bioluminescence. J. Photochem. Photobiol. Chem. 2001, 146, 95–107. [Google Scholar] [CrossRef]
- Ireland, J.F.; Wyatt, P.A.H. Acid-Base Properties of Electronically Excited States of Organic Molecules. In Advances in Physical Organic Chemistry; Gold, V., Ed.; Academic Press: New York, NY, USA, 1976; Volume 12, pp. 131–221. [Google Scholar]
- van Oort, B.; Eremeeva, E.V.; Koehorst, R.B.; Laptenok, S.P.; van Amerongen, H.; van Berkel, W.J.; Malikova, N.P.; Markova, S.V.; Vysotski, E.S.; Visser, A.J.; et al. Picosecond fluorescence relaxation spectroscopy of the calcium-discharged photoproteins aequorin and obelin. Biochemistry 2009, 48, 10486–10491. [Google Scholar] [CrossRef]
- Usami, K.; Isobe, M. Low temperature oxidation of coelenterate luciferin analog. Synthesis and proof of a 1,2-dioxetanone as a luminescence intermediate. Tetrahedron 1996, 52, 12061–12090. [Google Scholar] [CrossRef]
- Yue, L. QM/MM investigations on the bioluminescent decomposition of coelenterazine dioxetanone in obelin. Chem. Res. Chin. Univ. 2018, 34, 758–766. [Google Scholar] [CrossRef]
- Ding, B.W.; Liu, Y.J. Bioluminescence of firefly squid via mechanism of single electron-transfer oxygenation and charge-transfer-induced luminescence. J. Am. Chem. Soc. 2017, 139, 1106–1119. [Google Scholar] [CrossRef] [PubMed]
- Yue, L.; Liu, Y.J.; Fang, W.H. Mechanistic insight into the chemiluminescent decomposition of firefly dioxetanone. J. Am. Chem. Soc. 2012, 134, 11632–11639. [Google Scholar] [CrossRef]
- Chen, S.F.; Vysotski, E.S.; Liu, Y.J. H2O-bridged proton-transfer channel in emitter species formation in obelin bioluminescence. J. Phys. Chem. 2021, 125, 10452–10458. [Google Scholar] [CrossRef]
- Pinto da Silva, L.; Magalhães, C.M.; Crista, D.M.A.; Esteves da Silva, J.C.G. Theoretical modulation of singlet/triplet chemiexcitation of chemiluminescent imidazopyrazinone dioxetanone via C8-substitution. Photochem. Photobiol. Sci. 2017, 16, 897–907. [Google Scholar] [CrossRef]
- Min, C.G.; Ferreira, P.J.O.; Pinto da Silva, L. Theoretically obtained insight into the mechanism and dioxetanone species responsible for the singlet chemiexcitation of Coelenterazine. J. Photochem. Photobiol. 2017, 174, 18–26. [Google Scholar] [CrossRef]
- Nery, A.L.P.; Ropke, S.; Catalani, L.H.; Baader, W.J. Fluoride-triggered decomposition of m-sililoxyphenyl-substituted dioxetanes by an intramolecular electron transfer (CIEEL) mechanism. Tetrahedron. Lett. 1999, 40, 2443–2446. [Google Scholar] [CrossRef]
- Matsumoto, M.; Ito, Y.; Murakami, M.; Watanabe, N. Synthesis of 5-tert-butyl-1-(3-tert-butyldimethylsiloxy)phenyl-4,4-dimethyl-2,6,7-trioxabicyclo[3.2.0]heptanes and their fluoride-induced chemiluminescent decomposition: Effect of a phenolic electron donor on the CIEEL decay rate in aprotic polar solvent. Luminescence 2002, 17, 305–312. [Google Scholar] [CrossRef] [PubMed]
- Tanimura, M.; Watanabe, N.; Ijuin, H.K.; Matsumoto, M. Intramolecular charge-transfer-induced decomposition promoted by an aprotic polar solvent for bicyclic dioxetanes bearing a 4-(benzothiazol-2-yl)-3-hydroxyphenyl moiety. J. Org. Chem. 2011, 76, 902–908. [Google Scholar] [CrossRef]
- Adam, W.; Baader, W.J. Effects of methylation on the thermal stability and chemiluminescence properties of 1,2-dioxetanes. J. Am. Chem. Soc. 1985, 107, 410. [Google Scholar] [CrossRef]
- Magalhães, C.M.; Esteves da Silva, J.C.G.; Pinto da Silva, L. Comparative study of the chemiluminescence of coelenterazine, coelenterazine-e and Cypridina luciferin with an experimental and theoretical approach. J. Photochem. Photobiol. 2019, 190, 21–31. [Google Scholar] [CrossRef] [PubMed]
- Rhys, N.H.; Soper, A.K.; Dougan, L. The hydrogen-bonding ability of the amino acid glutamine revealed by neutron diffraction experiments. J. Phys. Chem. 2012, 116, 13308–13319. [Google Scholar] [CrossRef] [PubMed]
- Illarionov, B.A.; Frank, L.A.; Illarionova, V.A.; Bondar, V.S.; Vysotski, E.S.; Blinks, J.R. Recombinant obelin: Cloning and expression of cDNA purification, and characterization as a calcium indicator. Methods Enzymol. 2000, 305, 223–249. [Google Scholar] [PubMed]
- Vysotski, E.S.; Liu, Z.J.; Rose, J.; Wang, B.C.; Lee, J. Preparation and X-ray crystallographic analysis of recombinant obelin crystals diffracting to beyond 1.1 Å. Acta Cryst. Biol. Crystallogr. 2001, 57, 1919–1921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inouye, S.; Hojo, H. Revalidation of recombinant aequorin as a light emission standard: Estimation of specific activity of Gaussia luciferase. Biochem. Biophys. Res. Commun. 2018, 507, 242–245. [Google Scholar] [CrossRef] [PubMed]
- Mishin, A.; Gabdulkhakov, A.; Boyko, K.; Shevtsov, M.; Bukhdruker, S.; Borshchevskiy, V. Russian BAG for Xtallography and BioSAXS. Eur. Synchrotron Radiat. Facil. 2022. [Google Scholar] [CrossRef]
- Kabsch, W. Integration, scaling, space-group assignment and post-refinement. Acta Cryst. Biol. Crystallogr. 2010, 66, 133–144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panjikar, S.; Parthasarathy, V.; Lamzin, V.S.; Weiss, M.S.; Tucker, P.A. On the combination of molecular replacement and single-wavelength anomalous diffraction phasing for automated structure determination. Acta Cryst. Biol. Crystallogr. 2009, 65, 1089–1097. [Google Scholar] [CrossRef] [PubMed]
- Adams, P.D.; Afonine, P.V.; Bunkóczi, G.; Chen, V.B.; Davis, I.W.; Echols, N.; Headd, J.J.; Hung, L.W.; Kapral, G.J.; Grosse-Kunstleve, R.W.; et al. PHENIX: A comprehensive Python-based system for macromolecular structure solution. Acta Cryst. Biol Crystallogr. 2010, 66, 213–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Emsley, P.; Cowtan, K. Coot: Model-building tools for molecular graphics. Acta Cryst. Biol. Crystallogr. 2004, 60, 2126–2132. [Google Scholar] [CrossRef] [Green Version]
- Kabsch, W.; Sander, C. Dictionary of protein secondary structure: Pattern recognition of hydrogen-bonded and geometrical features. Biopolymers 1983, 22, 2577–2637. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Photoprotein | Maximal Light Intensity (Quanta/mg) a, ×1013 | Specific Activity (Quanta/mg) a, ×1015 | Relative Specific Activity | Ca2+-Independent Luminescence (Quanta/mg) a, ×103 |
|---|---|---|---|---|
| Obelin | ||||
| OL_wt | 62.19 ± 2.00 (1) b | 6.72 ± 0.04 | 1 | 70.80 ± 0.88 (1) c |
| OL_S47A | 9.34 ± 0.74 (0.15) | 6.44 ± 0.05 | 0.96 | 21.91 ± 0.31 (0.31) |
| OL_H64F | 0.32 ± 0.004 (0.05) | 2.41 ± 0.06 | 0.36 | 1.13 ± 0.12 (0.016) |
| OL_H64Q | 1.44 ± 0.08 (0.02) | 5.40 ± 0.11 | 0.80 | 17.10 ± 0.30 (0.24) |
| OL_W114F | 4.91 ± 0.71 (0.08) | 0.94 ± 0.01 | 0.14 | – |
| OL_Y138F | 0.48 ± 0.01 (0.01) | 3.52 ± 0.02 | 0.52 | 50.72 ± 0.84 (0.72) |
| OL_S47A/H64F | 0.03 ± 0.005 (0.0005) | 2.01 ± 0.04 | 0.30 | 0.66 ± 0.02 (0.009) |
| OL_S47A/H64Q | 1.27 ± 0.11 (0.02) | 6.18 ± 0.03 | 0.92 | 27.30 ± 0.60 (0.39) |
| OL_H64F/Y138F | 0.20 ± 0.05 (0.003) | 0.25 ± 0.002 | 0.04 | – |
| OL_H64Q/Y138F | 0.04 ± 0.001 (0.001) | 1.59 ± 0.07 | 0.24 | 11.63 ± 0.31 (0.16) |
| Aequorin | ||||
| AV_wt | 5.43 ± 0.27 (1) | 5.12 ± 0.09 | 1 | 17.01 ± 0.27 (1) |
| AV_S41A | 1.87 ± 0.14 (0.34) | 4.34 ± 0.17 | 0.85 | 3.20 ± 0.18 (0.19) |
| AV_H58F | 0.02 ± 0.002 (0.004) | 1.53 ± 0.05 | 0.30 | 0.11 ± 0.01 (0.006) |
| AV_H58Q | 1.47 ± 0.04 (0.27) | 3.63 ± 0.02 | 0.71 | 8.75 ± 0.21 (0.51) |
| AV_W108F | 0.33 ± 0.03 (0.06) | 0.52 ± 0.001 | 0.10 | – |
| AV_Y132F | 0.28 ± 0.002 (0.05) | 2.62 ± 0.17 | 0.51 | 14.32 ± 0.44 (0.84) |
| AV_S41A/H58F | 0.05 ± 0.003 (0.009) | 1.75 ± 0.01 | 0.34 | 0.60 ± 0.03 (0.035) |
| AV_S41A/H58Q | 1.20 ± 0.07 (0.22) | 4.25 ± 0.05 | 0.83 | 8.38 ± 0.24 (0.49) |
| AV_H58F/Y132F | 0.03 ± 0.002 (0.006) | 0.14 ± 0.003 | 0.03 | – |
| AV_H58Q/Y132F | 0.08 ± 0.006 (0.02) | 1.79 ± 0.01 | 0.35 | 6.07 ± 0.23 (0.36) |
| Photoprotein | Absorption Spectral Maximum λmax (nm) | Bioluminescence Spectral Maximum λshoulder (*)/λmax (nm) | Fluorescence Spectral Maximum λshoulder (*)/λmax (nm) |
|---|---|---|---|
| Obelin | |||
| OL_wt | 465 | 400 (0.17)/480 | 510 |
| OL_S47A | 465 | 400 (0.18)/480 | 420 (0.07)/508 |
| OL_H64F | 465 | 400 (0.05)/485 | 508 |
| OL_H64Q | 465 | 400 (0.08)/483 | 504 |
| OL_W114F | 345/450 | 400 (0.17)/485 | 410 (0.13)/506 |
| OL_Y138F | 455 | 400 (0.07)/487 | 420 (0.03)/512 |
| OL_S47A/H64F | 465 | 400 (0.05)/485 | 490 |
| OL_S47A/H64Q | 465 | 400 (0.09)/485 | 420 (0.05)/508 |
| OL_H64F/Y138F | 345/450 | 400 (0.07)/487 | 425 (0.37)/508 |
| OL_H64Q/Y138F | 455 | 400 (0.07)/492 | 425 (0.10)/510 |
| Aequorin | |||
| AV_wt | 455 | 470 | 470 |
| AV_S41A | 435/470 | 465 | 470 |
| AV_H58F | 460 | 390 (0.04)/480 | 476 |
| AV_H58Q | 455 | 390 (0.04)/480 | 480 |
| AV_W108F | 345/445 | 470 | 470 |
| AV_Y132F | 445 | 395 (0.03)/475 | 395 (0.16)/480 |
| AV_S41A/H58F | 460 | 390 (0.03)/480 | 460 |
| AV_S41A/H58Q | 455 | 480 | 476 |
| AV_H58F/Y132F | 345/450 | 395 (0.06)/485 | 420/486 (0.71) |
| AV_H58Q/Y132F | 445 | 480 | 420 (0.64)/480 |
| Photoprotein | krise | kdecay | |
|---|---|---|---|
| kdecay1 | kdecay2 | ||
| Obelin | |||
| OL_WT | 510.0 ± 5.0 | 40.00 ± 1.75 (0.66) a | 4.80 ± 0.05 (0.34) |
| OL_S47A | 416.9 ± 3.5 | 44.88 ± 1.59 (0.23) | 0.74 ± 0.002 (0.77) |
| OL_H64F | 144.2 ± 4.2 | 0.0035 ± 0.0006 | |
| OL_H64Q | 236.8 ± 0.6 | 0.49 ± 0.07 (0.07) | 0.13 ± 0.001 (0.93) |
| OL_W114F | 406.7 ± 4.2 | 2.25 ± 0.02 (0.53) | 0.21 ± 0.001 (0.47) |
| OL_Y138F | 305.3 ± 1.4 | 13.7 ± 0.30 (0.80) | 0.06 ± 0003 (0.20) |
| OL_S47A/H64F | 158.1 ± 2.0 | 0.09 ± 0.01 (0.13) | 0.003 ± 0.0006 (0.87) |
| OL_S47A/H64Q | 214.5 ± 0.8 | 0.112 ± 0.0003 | |
| OL_H64F/Y138F | 218.8 ± 1.6 | 0.111 ± 0.0003 | |
| OL_H64Q/Y138F | 278.1 ± 2.2 | 4.10 ± 0.26 (0.18) | 0.10 ± 0.001 (0.82) |
| Aequorin | |||
| AV_WT | 123.0 ± 1.0 | 0.81 ± 0.01 | |
| AV_S41A | 134.6 ± 1.4 | 0.36 ± 0.02 (0.43) | 0.18 ± 0.01 (0.57) |
| AV_H58F | 119.8 ± 0.4 | 0.26 ± 0.001 (0.52) | 0.01 ± 0.0001 (0.48) |
| AV_H58Q | 60.0 ± 0.2 | 0.35 ± 0.02 (0.46) | 0.23 ± 0.01 (0.54) |
| AV_W108F | 238.7 ± 3.8 | 0.88 ± 0.02 (0.43) | 0.17 ± 0.01 (0.57) |
| AV_Y132F | 32.4 ± 0.2 | 1.6 ± 0.21 (0.13) | 0.05 ± 0.001 (0.87) |
| AV_S41A/H58F | 51.4 ± 0.4 | 0.17 ± 0.01 (0.12) | 0.013 ± 0.0001 (0.88) |
| AV_S41A/H58Q | 70.1 ± 0.6 | 0.21 ± 0.002 (0.85) | 0.10 ± 0.002 (0.15) |
| AV_H58F/Y132F | 72.5 ± 0.6 | 0.87 ± 0.06 (0.07) | 0.19 ± 0.001 (0.93) |
| AV_H58Q/Y132F | 51.1 ± 0.8 | 0.21 ± 0.01 (0.35) | 0.02 ± 0.001 (0.65) |
| Obelin H64F Mutant (8C6O) | |
|---|---|
| Wavelength, Å | 0.9677 |
| Resolution range, Å | 38.76–2.2 (2.279–2.2) |
| Space group | P 61 |
| Unit cell, Å ° | 77.51 77.51 180.22 90 90 120 |
| Total reflections | 161,998 (16,720) |
| Unique reflections | 29,727 (3029) |
| Multiplicity | 5.4 (5.5) |
| Completeness, % | 95.9 (97.3) |
| Mean I/sigma (I) | 8.73 (0.94) |
| Wilson B-factor, Å2 | 40.6 |
| R-merge, % | 16.4 (220.1) |
| R-meas, % | 18.2 (242.7) |
| R-pim, % | 7.6 (100) |
| CC1/2, % | 99.5 (27.7) |
| CC*, % | 99.9 (65.8) |
| Reflections used in refinement | 29,719 (3028) |
| Reflections used for R-free | 844 (60) |
| R-work, % | 17.2 (30.2) |
| R-free, % | 20.3 (30.2) |
| CC (work), % | 96.2 (62.9) |
| CC (free), % | 94.2 (62.0) |
| Number of non-hydrogen atoms | 3446 |
| macromolecules | 3112 |
| ligands | 70 |
| solvent | 264 |
| Protein residues | 391 |
| RMS (bonds), Å | 0.010 |
| RMS (angles), ° | 0.96 |
| Ramachandran favored, % | 98.2 |
| Ramachandran allowed, % | 1.8 |
| Ramachandran outliers, % | 0.00 |
| Rotamer outliers, % | 0.31 |
| Clashscore | 1.15 |
| Average B-factor, Å2 | 43.2 |
| macromolecules | 42.9 |
| ligands | 44.3 |
| solvent | 47.4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Natashin, P.V.; Burakova, L.P.; Kovaleva, M.I.; Shevtsov, M.B.; Dmitrieva, D.A.; Eremeeva, E.V.; Markova, S.V.; Mishin, A.V.; Borshchevskiy, V.I.; Vysotski, E.S. The Role of Tyr-His-Trp Triad and Water Molecule Near the N1-Atom of 2-Hydroperoxycoelenterazine in Bioluminescence of Hydromedusan Photoproteins: Structural and Mutagenesis Study. Int. J. Mol. Sci. 2023, 24, 6869. https://doi.org/10.3390/ijms24076869
Natashin PV, Burakova LP, Kovaleva MI, Shevtsov MB, Dmitrieva DA, Eremeeva EV, Markova SV, Mishin AV, Borshchevskiy VI, Vysotski ES. The Role of Tyr-His-Trp Triad and Water Molecule Near the N1-Atom of 2-Hydroperoxycoelenterazine in Bioluminescence of Hydromedusan Photoproteins: Structural and Mutagenesis Study. International Journal of Molecular Sciences. 2023; 24(7):6869. https://doi.org/10.3390/ijms24076869
Chicago/Turabian StyleNatashin, Pavel V., Ludmila P. Burakova, Margarita I. Kovaleva, Mikhail B. Shevtsov, Daria A. Dmitrieva, Elena V. Eremeeva, Svetlana V. Markova, Alexey V. Mishin, Valentin I. Borshchevskiy, and Eugene S. Vysotski. 2023. "The Role of Tyr-His-Trp Triad and Water Molecule Near the N1-Atom of 2-Hydroperoxycoelenterazine in Bioluminescence of Hydromedusan Photoproteins: Structural and Mutagenesis Study" International Journal of Molecular Sciences 24, no. 7: 6869. https://doi.org/10.3390/ijms24076869