
Doxorubicin–Mediated miR–433 Expression on Exosomes Promotes Bystander Senescence in Multiple Myeloma Cells in a DDR–Independent Manner
 ,
,  , , , and
, , , and
Abstract
:1. Introduction
2. Results
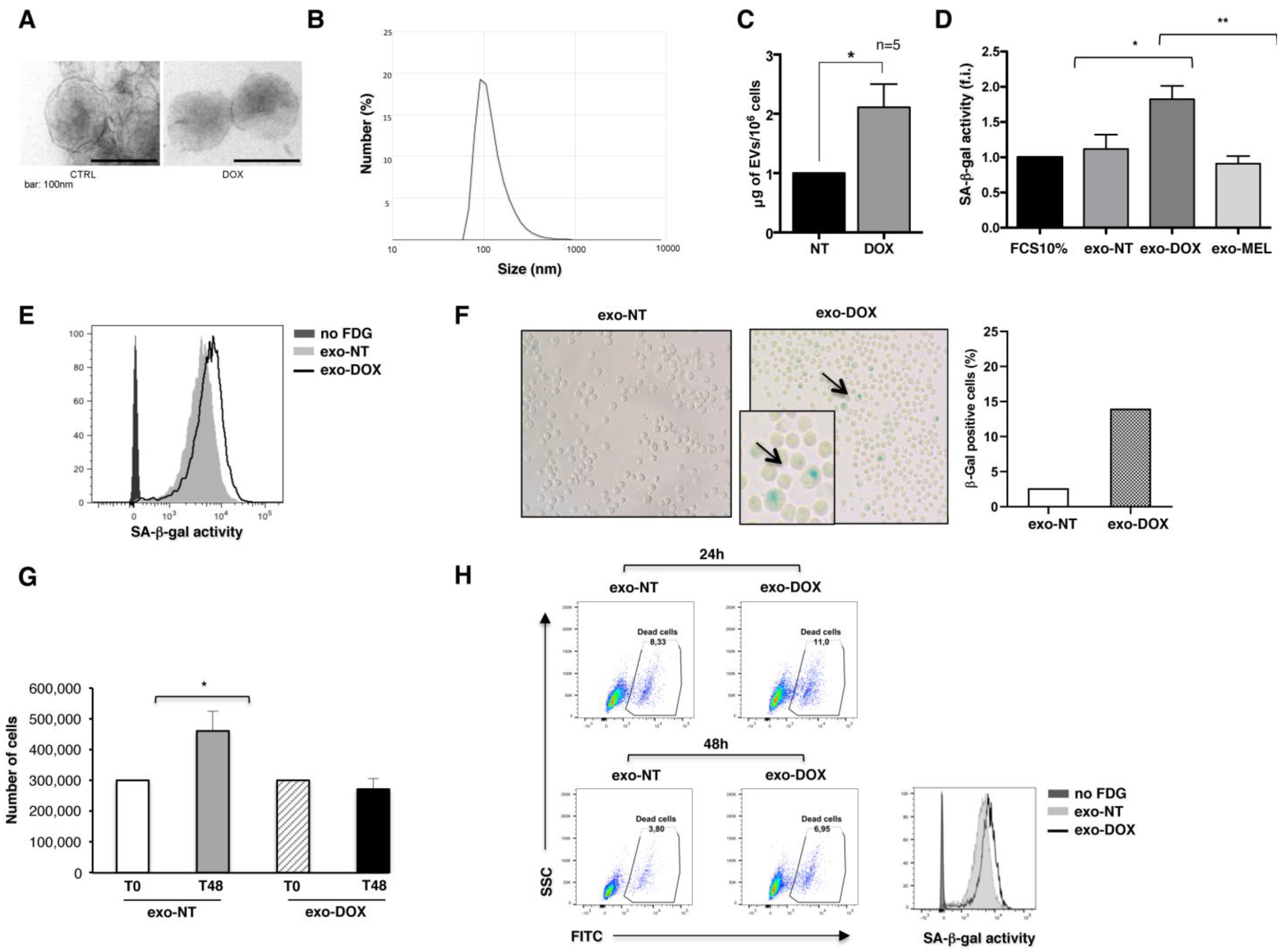
2.1. Exosomes Released by DOX-Treated Multiple Myeloma Cells Induce a Senescence-Like Phenotype on Bystander Cells
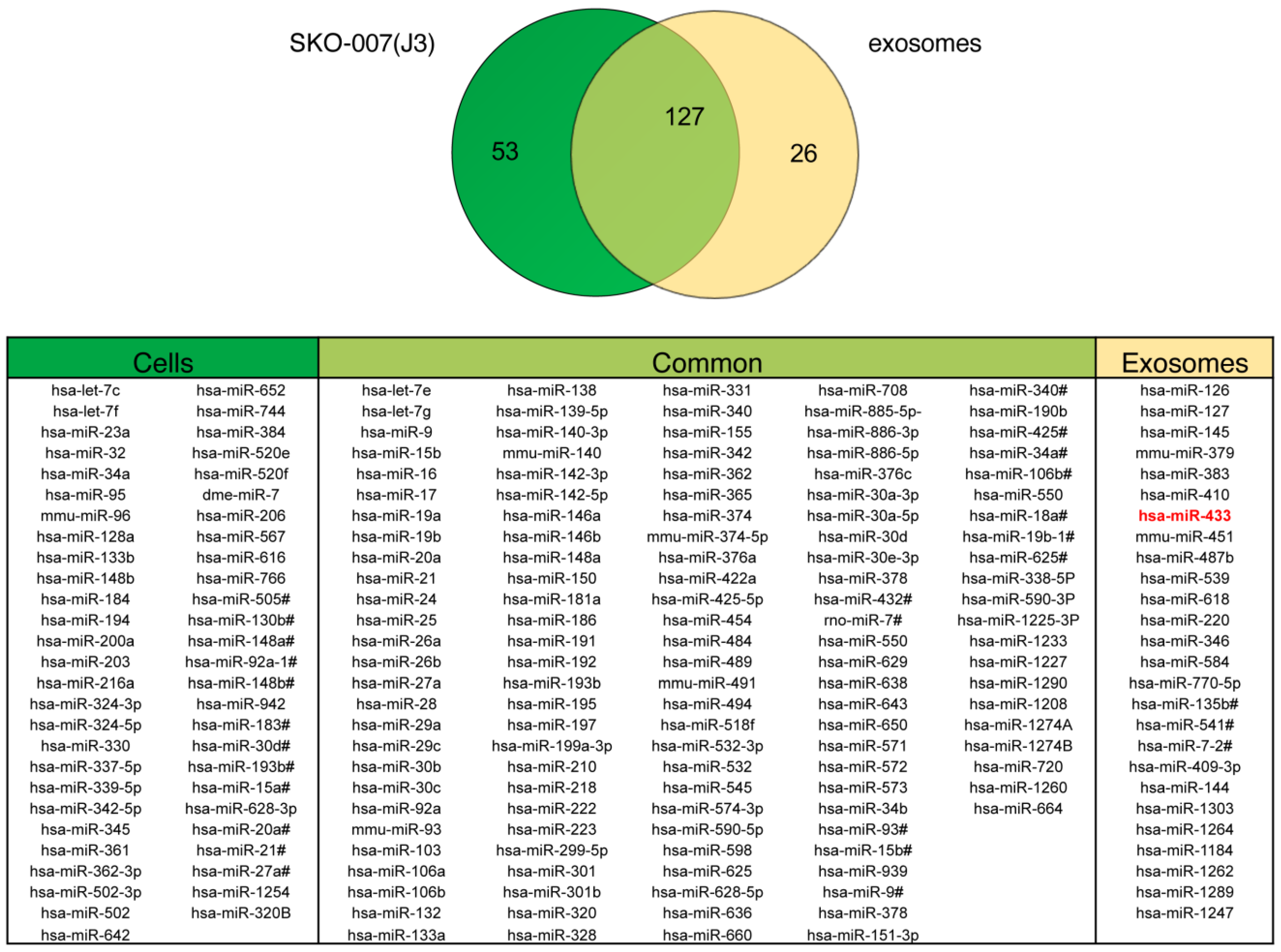
2.2. Exosomes Released by DOX-Treated MM Cells Convey miR-433
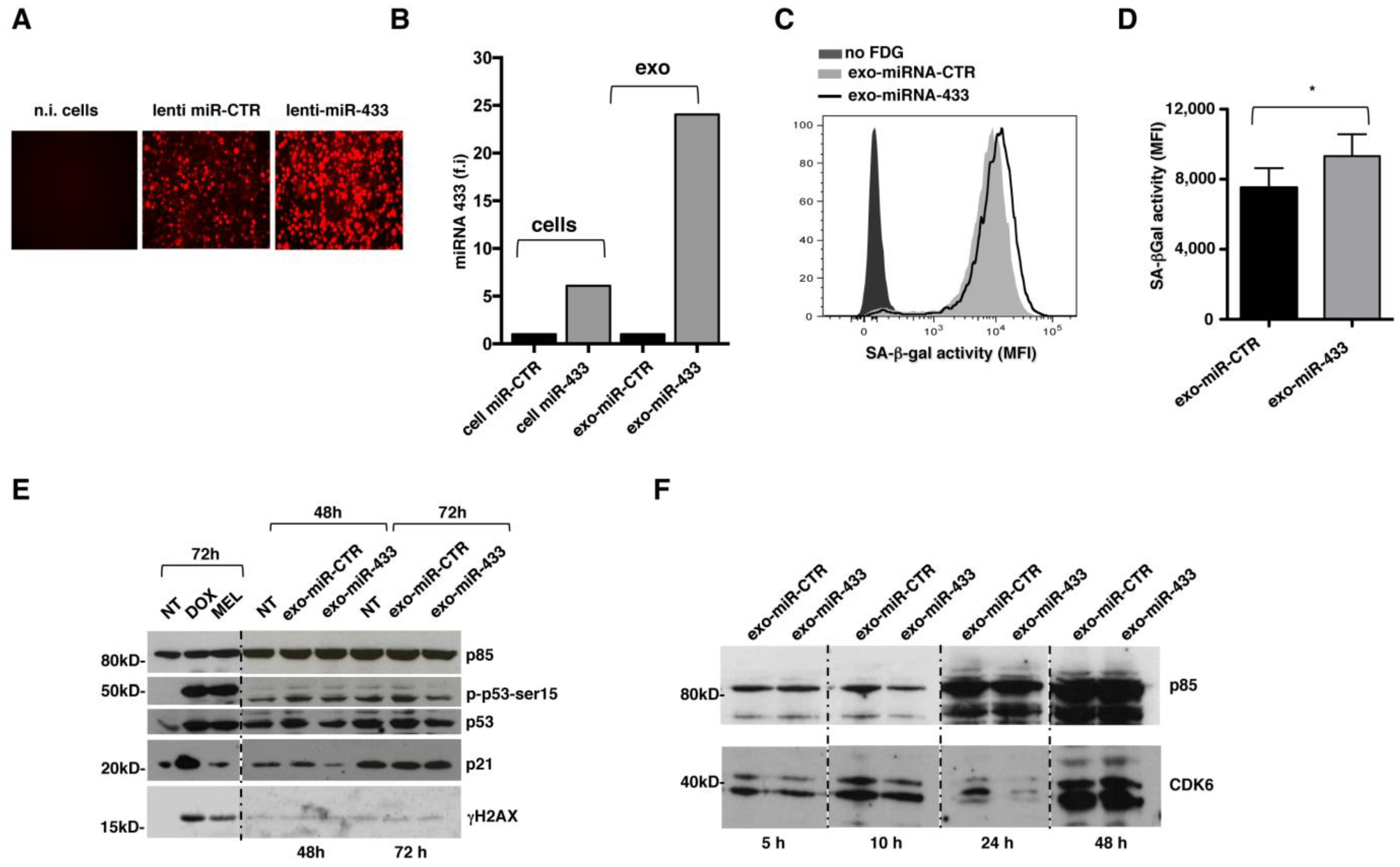
2.3. Overexpression of miR-433 in MM Cells Results in the Induction of a Senescent Phenotype through CDK6 Down-Modulation
3. Discussion
4. Materials and Methods
4.1. Cell Lines and Clinical Samples
4.2. Treatments
4.3. Senescence Associated-β-Galactosidase Staining
4.4. Immunofluorescence and Flow Cytometry
4.5. Exosome Isolation
4.6. Transmission Electron Microscopy
4.7. Size Experiments through Dynamic Light Scattering (DLS)
4.8. Analysis of Exosomes by Flow Cytometry
4.9. Exosomal RNA Isolation
4.10. Exosome miRNA Profiling and Real-Time PCR Analysis
4.11. SDS-PAGE and Western Blot
4.12. Virus Production and In Vitro Transduction
4.13. Degranulation Assay
4.14. Statistics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Palumbo, A.; Anderson, K. Multiple Myeloma. N. Engl. J. Med. 2011, 364, 1046–1060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giannakoulas, N.; Ntanasis-Stathopoulos, I.; Terpos, E. The Role of Marrow Microenvironment in the Growth and Development of Malignant Plasma Cells in Multiple Myeloma. Int. J. Mol. Sci. 2021, 22, 4462. [Google Scholar] [CrossRef]
- Soriani, A.; Fionda, C.; Ricci, B.; Iannitto, M.L.; Cippitelli, M.; Santoni, A. Chemotherapy—Elicited Upregulation of NKG2D and DNAM-1 Ligands as a Therapeutic Target in Multiple Myeloma. Oncoimmunology 2013, 2, e26663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soriani, A.; Zingoni, A.; Cerboni, C.; Iannitto, M.L.; Ricciardi, M.R.; Di Gialleonardo, V.; Cippitelli, M.; Fionda, C.; Petrucci, M.T.; Guarini, A.; et al. ATM-ATR-Dependent up-Regulation of DNAM-1 and NKG2D Ligands on Multiple Myeloma Cells by Therapeutic Agents Results in Enhanced NK-Cell Susceptibility and Is Associated with a Senescent Phenotype. Blood 2009, 113, 3503–3511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borrelli, C.; Ricci, B.; Vulpis, E.; Fionda, C.; Ricciardi, M.R.; Petrucci, M.T.; Masuelli, L.; Peri, A.; Cippitelli, M.; Zingoni, A.; et al. Drug-Induced Senescent Multiple Myeloma Cells Elicit NK Cell Proliferation by Direct or Exosome-Mediated IL15 Trans-Presentation. Cancer Immunol. Res. 2018, 6, 860–869. [Google Scholar] [CrossRef] [Green Version]
- Antonangeli, F.; Soriani, A.; Ricci, B.; Ponzetta, A.; Benigni, G.; Morrone, S.; Bernardini, G.; Santoni, A. Natural Killer Cell Recognition of In Vivo Drug-Induced Senescent Multiple Myeloma Cells. Oncoimmunology 2016, 5, e1218105. [Google Scholar] [CrossRef] [Green Version]
- Faget, D.V.; Ren, Q.; Stewart, S.A. Unmasking Senescence: Context-Dependent Effects of SASP in Cancer. Nat. Rev. Cancer 2019, 19, 439–453. [Google Scholar] [CrossRef]
- Nelson, G.; Kucheryavenko, O.; Wordsworth, J.; von Zglinicki, T. The Senescent Bystander Effect Is Caused by ROS-Activated NF-ΚB Signalling. Mech. Ageing Dev. 2018, 170, 30–36. [Google Scholar] [CrossRef]
- Coppé, J.-P.; Desprez, P.-Y.; Krtolica, A.; Campisi, J. The Senescence-Associated Secretory Phenotype: The Dark Side of Tumor Suppression. Annu. Rev. Pathol. 2010, 5, 99–118. [Google Scholar] [CrossRef] [Green Version]
- Takasugi, M.; Okada, R.; Takahashi, A.; Virya Chen, D.; Watanabe, S.; Hara, E. Small Extracellular Vesicles Secreted from Senescent Cells Promote Cancer Cell Proliferation through EphA2. Nat. Commun. 2017, 8, 15729. [Google Scholar] [CrossRef] [Green Version]
- Weilner, S.; Schraml, E.; Wieser, M.; Messner, P.; Schneider, K.; Wassermann, K.; Micutkova, L.; Fortschegger, K.; Maier, A.B.; Westendorp, R.; et al. Secreted Microvesicular MiR-31 Inhibits Osteogenic Differentiation of Mesenchymal Stem Cells. Aging Cell 2016, 15, 744–754. [Google Scholar] [CrossRef]
- Jakhar, R. Exosomes as Emerging Pro-Tumorigenic Mediators of the Senescence-Associated Secretory Phenotype. Int. J. Mol. Sci. 2019, 20, 2547. [Google Scholar] [CrossRef] [Green Version]
- Roccaro, A.M.; Sacco, A.; Maiso, P.; Azab, A.K.; Tai, Y.-T.; Reagan, M.; Azab, F.; Flores, L.M.; Campigotto, F.; Weller, E.; et al. BM Mesenchymal Stromal Cell-Derived Exosomes Facilitate Multiple Myeloma Progression. J. Clin. Investig. 2013, 123, 1542–1555. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, T.; Kristensen, S.R.; Gregersen, H.; Teodorescu, E.M.; Christiansen, G.; Pedersen, S. Extracellular Vesicle-Associated Procoagulant Phospholipid and Tissue Factor Activity in Multiple Myeloma. PLoS ONE 2019, 14, e0210835. [Google Scholar] [CrossRef]
- Saltarella, I.; Lamanuzzi, A.; Apollonio, B.; Desantis, V.; Bartoli, G.; Vacca, A.; Frassanito, M.A. Role of Extracellular Vesicle-Based Cell-to-Cell Communication in Multiple Myeloma Progression. Cells 2021, 10, 3185. [Google Scholar] [CrossRef]
- Zhang, L.; Lei, Q.; Wang, H.; Xu, C.; Liu, T.; Kong, F.; Yang, C.; Yan, G.; Sun, L.; Zhao, A.; et al. Tumor-Derived Extracellular Vesicles Inhibit Osteogenesis and Exacerbate Myeloma Bone Disease. Theranostics 2019, 9, 196–209. [Google Scholar] [CrossRef] [PubMed]
- Vulpis, E.; Cecere, F.; Molfetta, R.; Soriani, A.; Fionda, C.; Peruzzi, G.; Caracciolo, G.; Palchetti, S.; Masuelli, L.; Simonelli, L.; et al. Genotoxic Stress Modulates the Release of Exosomes from Multiple Myeloma Cells Capable of Activating NK Cell Cytokine Production: Role of HSP70/TLR2/NF-KB Axis. Oncoimmunology 2017, 6, e1279372. [Google Scholar] [CrossRef] [Green Version]
- Yang, T.; Martin, P.; Fogarty, B.; Brown, A.; Schurman, K.; Phipps, R.; Yin, V.P.; Lockman, P.; Bai, S. Exosome Delivered Anticancer Drugs Across the Blood-Brain Barrier for Brain Cancer Therapy in Danio Rerio. Pharm. Res 2015, 32, 2003–2014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weiner-Gorzel, K.; Dempsey, E.; Milewska, M.; McGoldrick, A.; Toh, V.; Walsh, A.; Lindsay, S.; Gubbins, L.; Cannon, A.; Sharpe, D.; et al. Overexpression of the MicroRNA MiR-433 Promotes Resistance to Paclitaxel through the Induction of Cellular Senescence in Ovarian Cancer Cells. Cancer Med. 2015, 4, 745–758. [Google Scholar] [CrossRef]
- Garcia-Martin, R.; Wang, G.; Brandão, B.B.; Zanotto, T.M.; Shah, S.; Kumar Patel, S.; Schilling, B.; Kahn, C.R. MicroRNA Sequence Codes for Small Extracellular Vesicle Release and Cellular Retention. Nature 2022, 601, 446–451. [Google Scholar] [CrossRef] [PubMed]
- Soriani, A.; Iannitto, M.L.; Ricci, B.; Fionda, C.; Malgarini, G.; Morrone, S.; Peruzzi, G.; Ricciardi, M.R.; Petrucci, M.T.; Cippitelli, M.; et al. Reactive Oxygen Species- and DNA Damage Response-Dependent NK Cell Activating Ligand Upregulation Occurs at Transcriptional Levels and Requires the Transcriptional Factor E2F1. J. Immunol. 2014, 193, 950–960. [Google Scholar] [CrossRef] [Green Version]
- Coppé, J.P.; Patil, C.K.; Rodier, F.; Sun, Y.; Muñoz, D.P.; Goldstein, J.; Nelson, P.S.; Desprez, P.Y.; Campisi, J. Senescence-Associated Secretory Phenotypes Reveal Cell-Nonautonomous Functions of Oncogenic RAS and the P53 Tumor Suppressor. PLoS Biol. 2008, 6, e301. [Google Scholar] [CrossRef] [PubMed]
- Iannello, A.; Thompson, T.W.; Ardolino, M.; Lowe, S.W.; Raulet, D.H. P53-Dependent Chemokine Production by Senescent Tumor Cells Supports NKG2D-Dependent Tumor Elimination by Natural Killer Cells. J. Exp. Med. 2013, 210, 2057–2069. [Google Scholar] [CrossRef]
- Davalos, A.R.; Coppe, J.P.; Campisi, J.; Desprez, P.Y. Senescent Cells as a Source of Inflammatory Factors for Tumor Progression. Cancer Metastasis Rev. 2010, 29, 273–283. [Google Scholar] [CrossRef] [Green Version]
- Krizhanovsky, V.; Yon, M.; Dickins, R.A.; Hearn, S.; Simon, J.; Miething, C.; Yee, H.; Zender, L.; Lowe, S.W. Senescence of Activated Stellate Cells Limits Liver Fibrosis. Cell 2008, 134, 657–667. [Google Scholar] [CrossRef] [Green Version]
- Raposo, G.; Stoorvogel, W. Extracellular Vesicles: Exosomes, Microvesicles, and Friends. J. Cell Biol. 2013, 200, 373–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernández-Messina, L.; Gutiérrez-Vázquez, C.; Rivas-García, E.; Sánchez-Madrid, F.; de la Fuente, H. Immunomodulatory Role of MicroRNAs Transferred by Extracellular Vesicles. Biol. Cell 2015, 107, 61–77. [Google Scholar] [CrossRef] [Green Version]
- Lehmann, B.D.; Paine, M.S.; Brooks, A.M.; McCubrey, J.A.; Renegar, R.H.; Wang, R.; Terrian, D.M. Senescence-Associated Exosome Release from Human Prostate Cancer Cells. Cancer Res. 2008, 68, 7864–7871. [Google Scholar] [CrossRef] [Green Version]
- Takasugi, M. Emerging Roles of Extracellular Vesicles in Cellular Senescence and Aging. Aging Cell 2018, 17, e12734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suh, N. MicroRNA Controls of Cellular Senescence. BMB Rep. 2018, 51, 493–499. [Google Scholar] [CrossRef] [Green Version]
- Xu, D.; Tahara, H. The Role of Exosomes and MicroRNAs in Senescence and Aging. Adv. Drug Deliv. Rev. 2013, 65, 368–375. [Google Scholar] [CrossRef]
- Schmitt, C.A. Senescence, Apoptosis and Therapy—Cutting the Lifelines of Cancer. Nat. Rev. Cancer 2003, 3, 286–295. [Google Scholar] [CrossRef]
- Villarroya-Beltri, C.; Gutiérrez-Vázquez, C.; Sánchez-Cabo, F.; Pérez-Hernández, D.; Vázquez, J.; Martin-Cofreces, N.; Martinez-Herrera, D.J.; Pascual-Montano, A.; Mittelbrunn, M.; Sánchez-Madrid, F. Sumoylated HnRNPA2B1 Controls the Sorting of MiRNAs into Exosomes through Binding to Specific Motifs. Nat. Commun. 2013, 4, 2980. [Google Scholar] [CrossRef] [Green Version]
- Guo, L.-H.; Li, H.; Wang, F.; Yu, J.; He, J.-S. The Tumor Suppressor Roles of MiR-433 and MiR-127 in Gastric Cancer. Int. J. Mol. Sci. 2013, 14, 14171–14184. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.; Tsuchiya, H.; Zhang, Y.; Hartnett, M.E.; Wang, L. MicroRNA-433 Inhibits Liver Cancer Cell Migration by Repressing the Protein Expression and Function of CAMP Response Element-Binding Protein. J. Biol. Chem. 2013, 288, 28893–28899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, S.; Wang, X.; Xu, X.; Di, H.; Du, J.; Xu, B.; Wang, Q.; Wang, J. MiR-433-3p Suppresses Cell Growth and Enhances Chemosensitivity by Targeting CREB in Human Glioma. Oncotarget 2017, 8, 5057–5068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, T.; Wang, X.; Zhou, D.; Qiu, H.; Zhang, J.; Yang, W. Circ_0003489 Facilitates Multiple Myeloma Progression by Targeting MiR-433-3p/PBX3 Axis. Hematology 2022, 27, 951–959. [Google Scholar] [CrossRef]
- Debacq-Chainiaux, F.; Erusalimsky, J.D.; Campisi, J.; Toussaint, O. Protocols to Detect Senescence-Associated Beta-Galactosidase (SA-Βgal) Activity, a Biomarker of Senescent Cells in Culture and in vivo. Nat. Protoc. 2009, 4, 1798–1806. [Google Scholar] [CrossRef] [PubMed]
- Vulpis, E.; Loconte, L.; Peri, A.; Molfetta, R.; Caracciolo, G.; Masuelli, L.; Tomaipitinca, L.; Peruzzi, G.; Petillo, S.; Petrucci, M.T.; et al. Impact on NK Cell Functions of Acute versus Chronic Exposure to Extracellular Vesicle-Associated MICA: Dual Role in Cancer Immunosurveillance. J. Extracell. Vesicles 2022, 11, e12176. [Google Scholar] [CrossRef]
- Soriani, A.; Borrelli, C.; Ricci, B.; Molfetta, R.; Zingoni, A.; Fionda, C.; Carnevale, S.; Abruzzese, M.P.; Petrucci, M.T.; Ricciardi, M.R.; et al. P38 MAPK Differentially Controls NK Activating Ligands at Transcriptional and Post-Transcriptional Level on Multiple Myeloma Cells. Oncoimmunology 2017, 6, e1264564. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| N° | Sex | Age | Disease Stage | % PCs in BM |
|---|---|---|---|---|
| 1 | M | 75 | relapse | 50 |
| 2 | F | 70 | relapse | 40 |
| 3 | M | 66 | onset | 42 |
| 4 | F | 63 | onset | 51 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vulpis, E.; Cuollo, L.; Borrelli, C.; Antonangeli, F.; Masuelli, L.; Cippitelli, M.; Fionda, C.; Caracciolo, G.; Petrucci, M.T.; Santoni, A.; et al. Doxorubicin–Mediated miR–433 Expression on Exosomes Promotes Bystander Senescence in Multiple Myeloma Cells in a DDR–Independent Manner. Int. J. Mol. Sci. 2023, 24, 6862. https://doi.org/10.3390/ijms24076862
Vulpis E, Cuollo L, Borrelli C, Antonangeli F, Masuelli L, Cippitelli M, Fionda C, Caracciolo G, Petrucci MT, Santoni A, et al. Doxorubicin–Mediated miR–433 Expression on Exosomes Promotes Bystander Senescence in Multiple Myeloma Cells in a DDR–Independent Manner. International Journal of Molecular Sciences. 2023; 24(7):6862. https://doi.org/10.3390/ijms24076862
Chicago/Turabian StyleVulpis, Elisabetta, Lorenzo Cuollo, Cristiana Borrelli, Fabrizio Antonangeli, Laura Masuelli, Marco Cippitelli, Cinzia Fionda, Giulio Caracciolo, Maria Teresa Petrucci, Angela Santoni, and et al. 2023. "Doxorubicin–Mediated miR–433 Expression on Exosomes Promotes Bystander Senescence in Multiple Myeloma Cells in a DDR–Independent Manner" International Journal of Molecular Sciences 24, no. 7: 6862. https://doi.org/10.3390/ijms24076862