
The Influence of Edaphic Factors on DNA Damage and Repair in Wild Wheat Triticum dicoccoides Körn. (Poaceae, Triticeae)


Abstract
:1. Introduction
2. Results
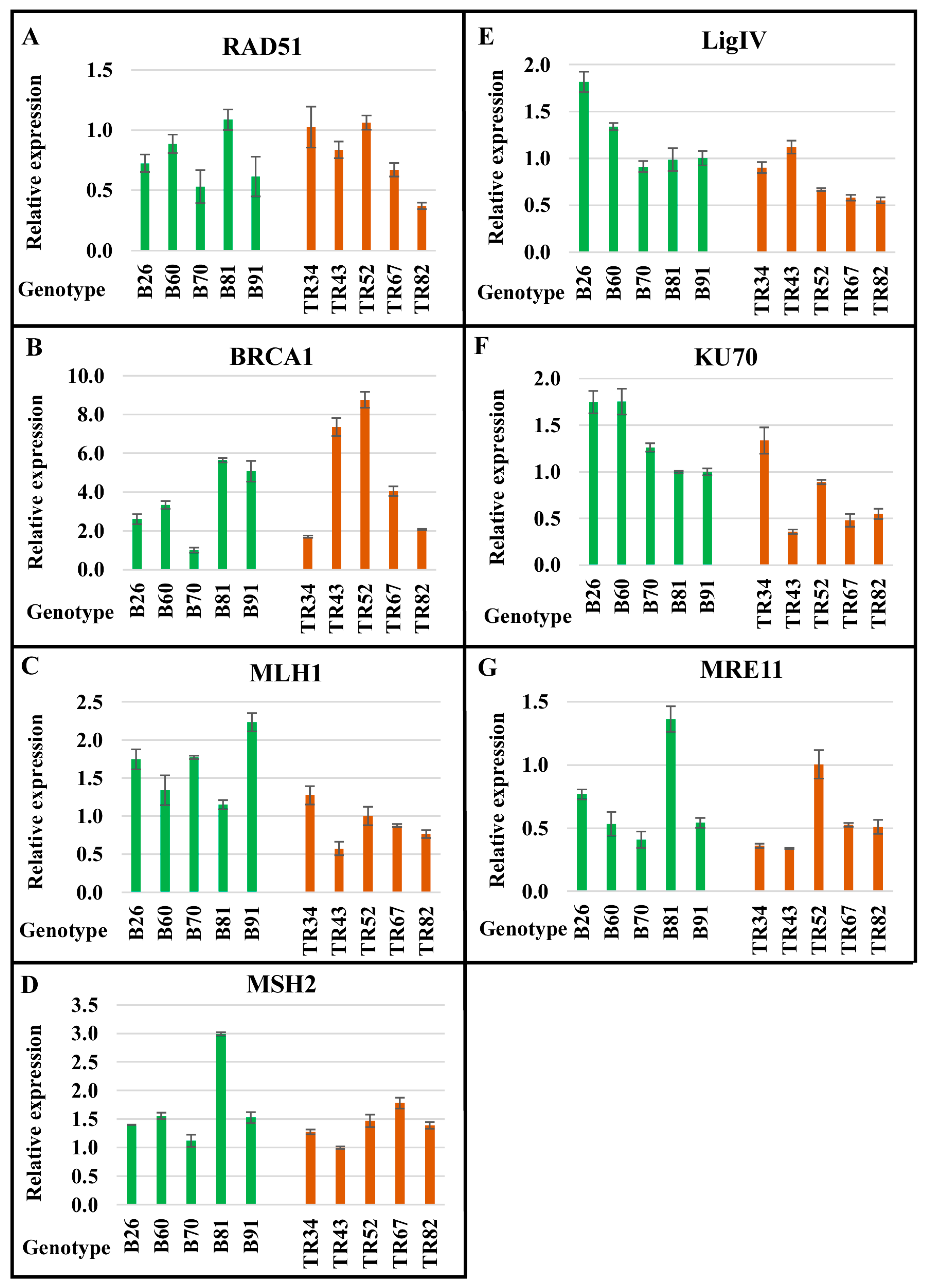
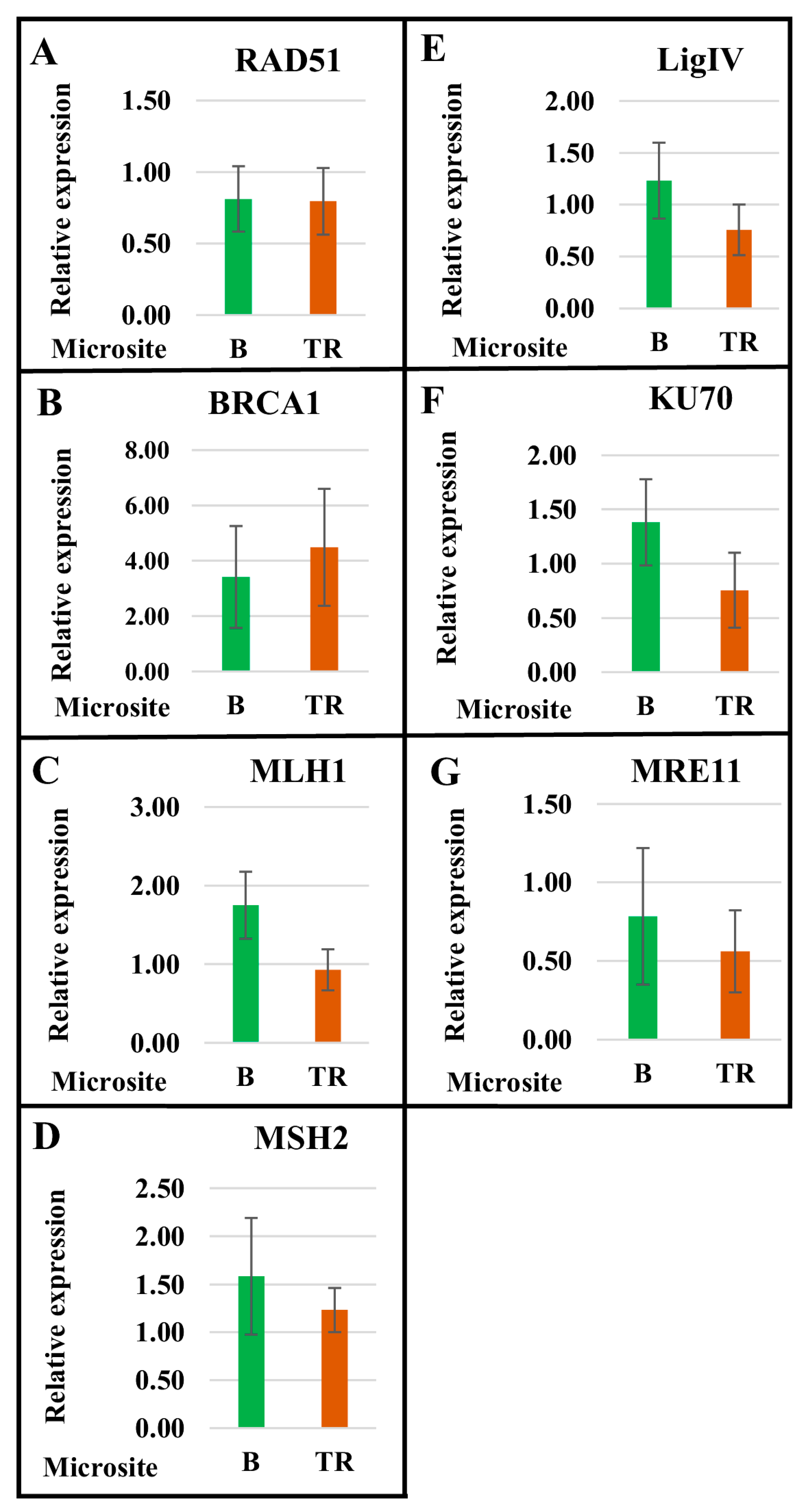
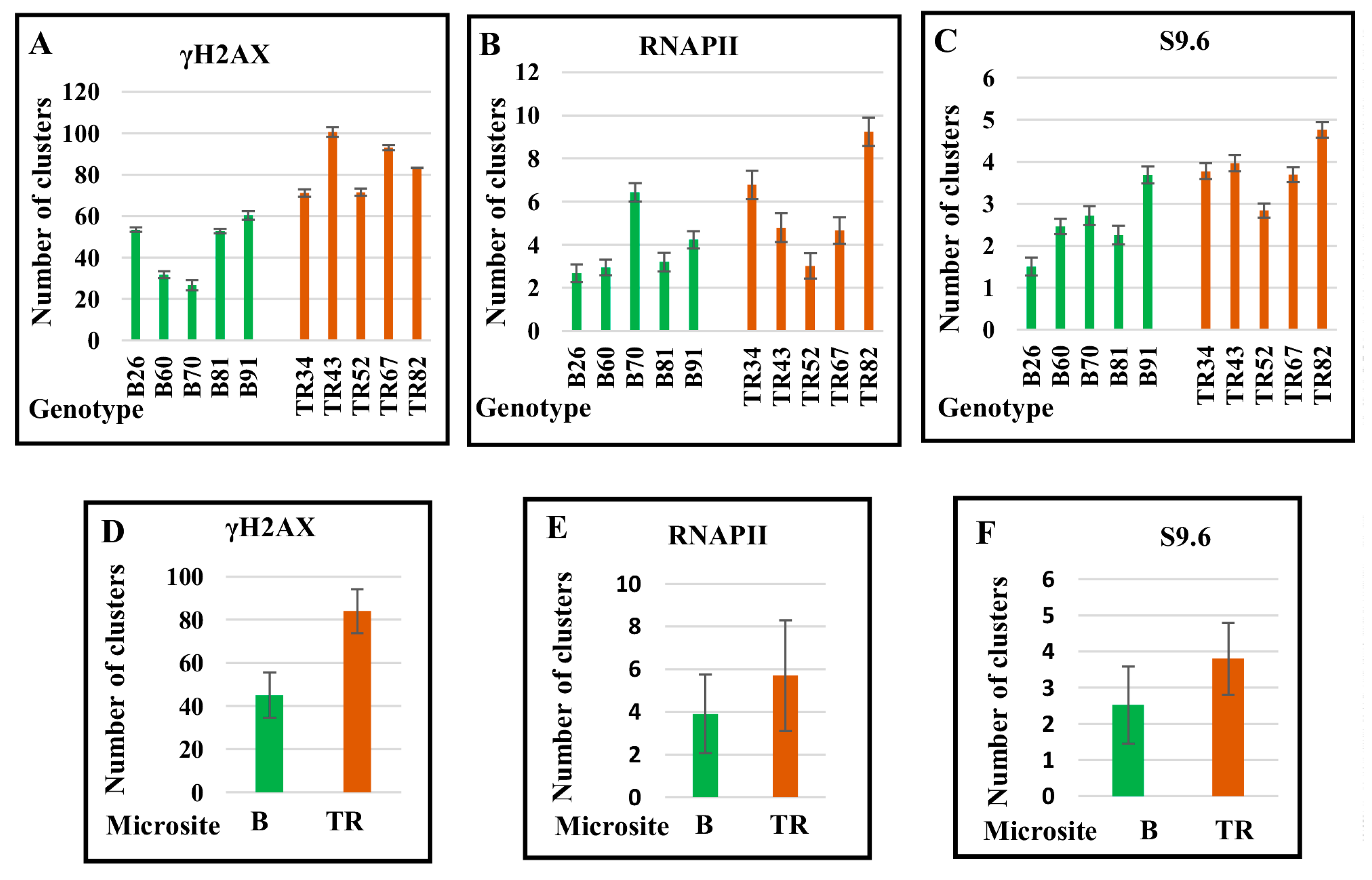
2.1. The Level of Expression of Genes Encoding Proteins Involved in DNA Repair Pathways Depends on the Genotype and Differs between the Basalt and the Terra Rossa Microsites
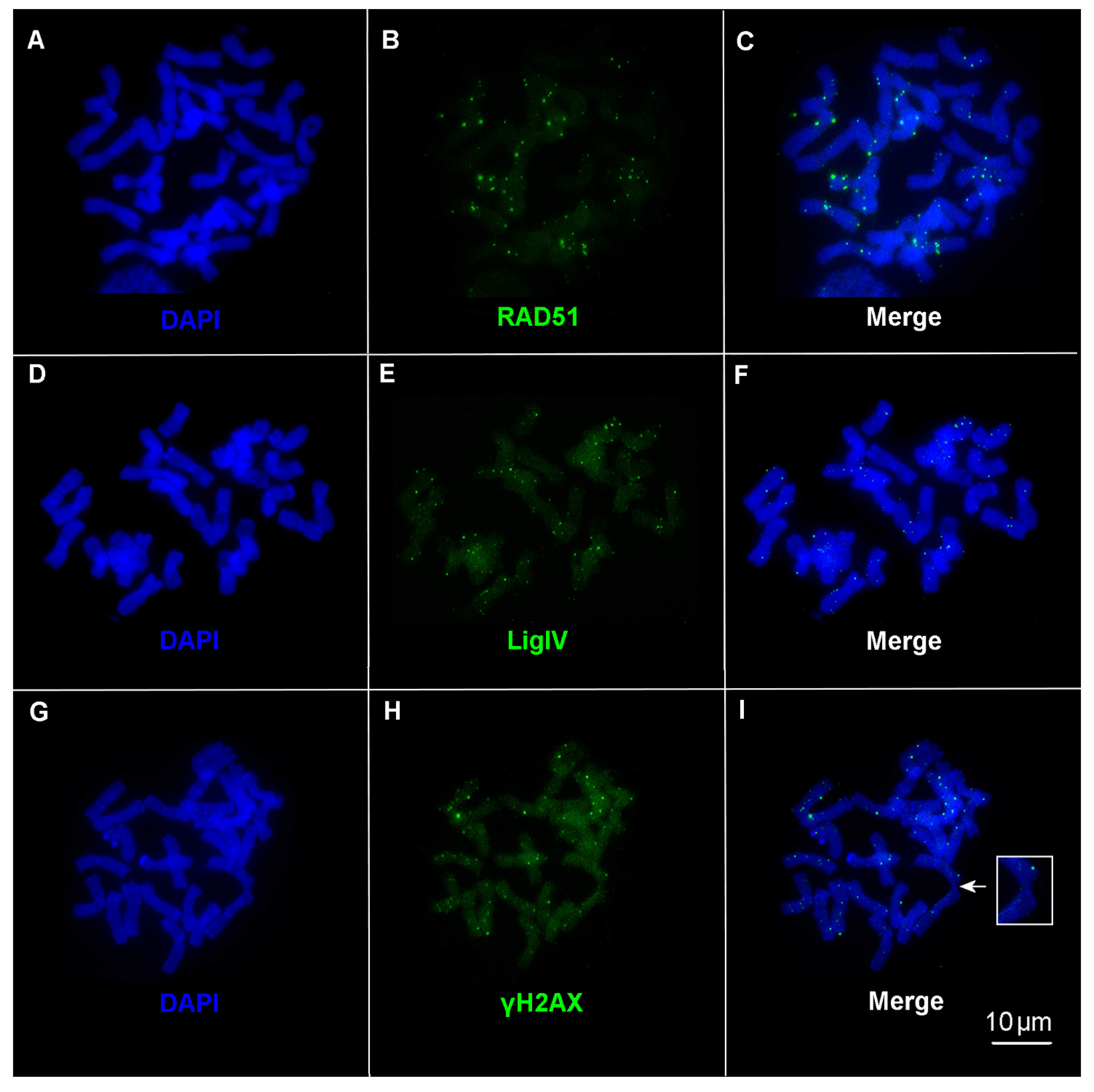
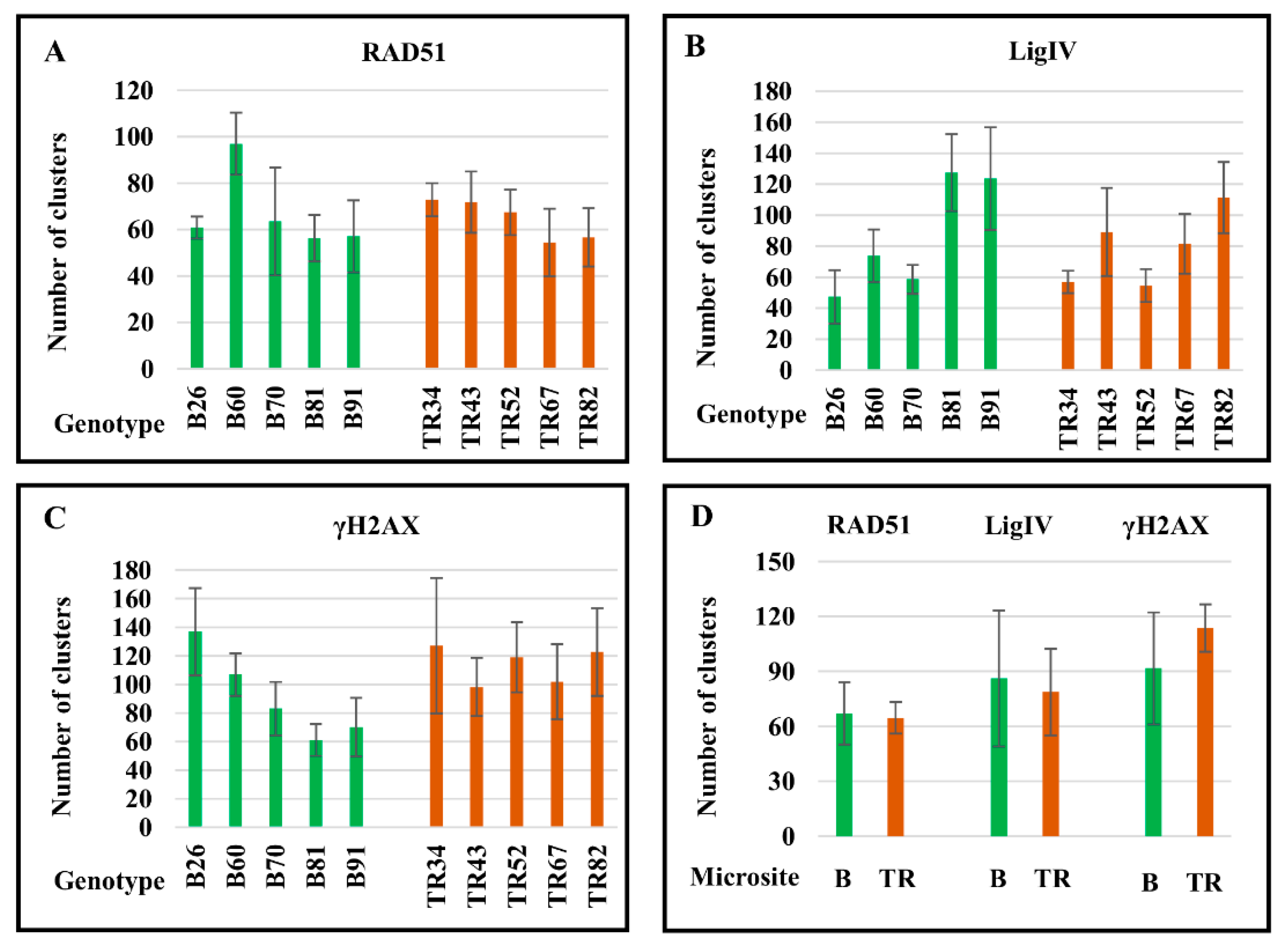
2.2. DNA Repair Continues in the M-Phase with Genotype-Dependent Efficiency
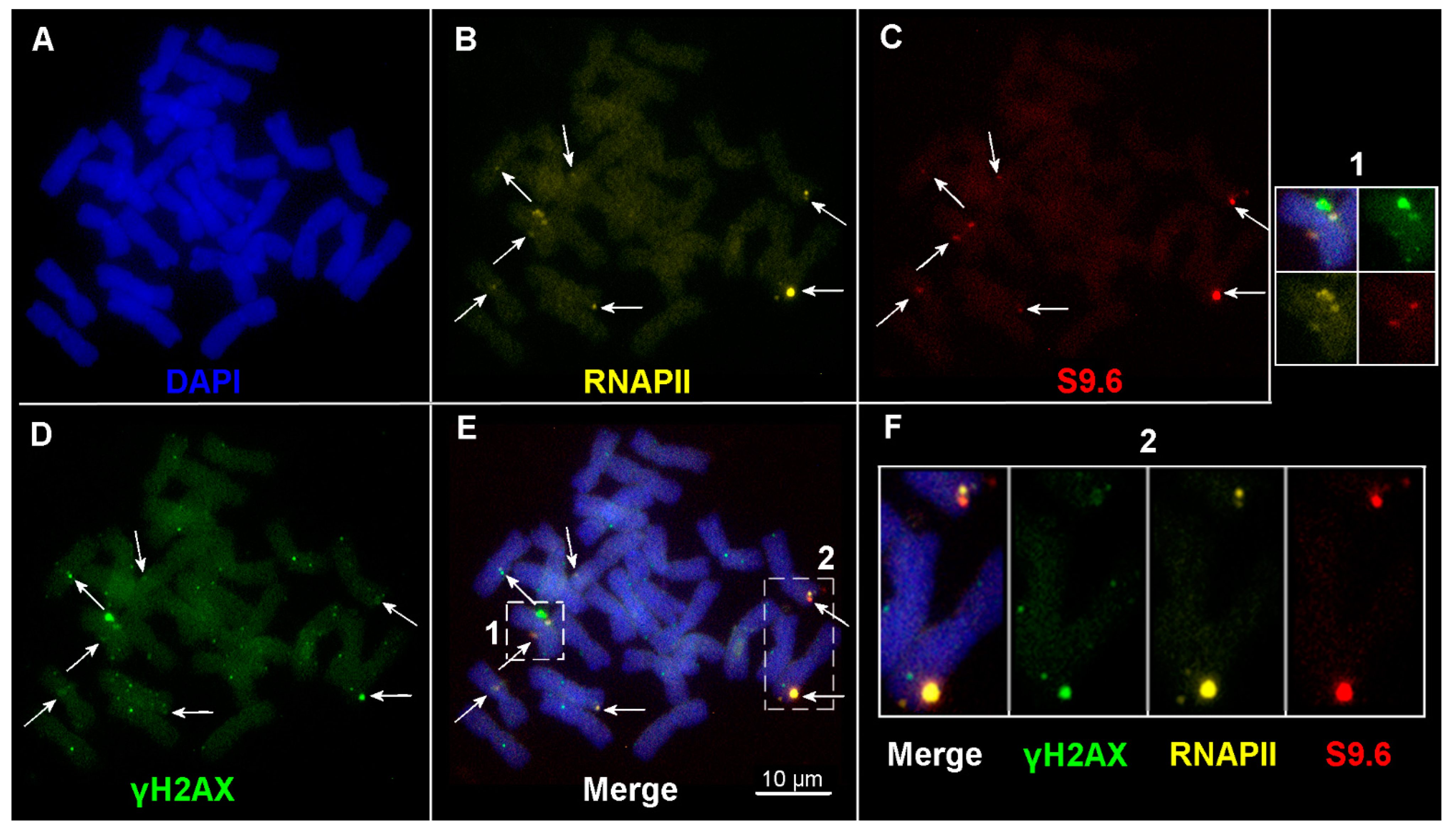
2.3. R-Loop Formation on Metaphase Chromosomes
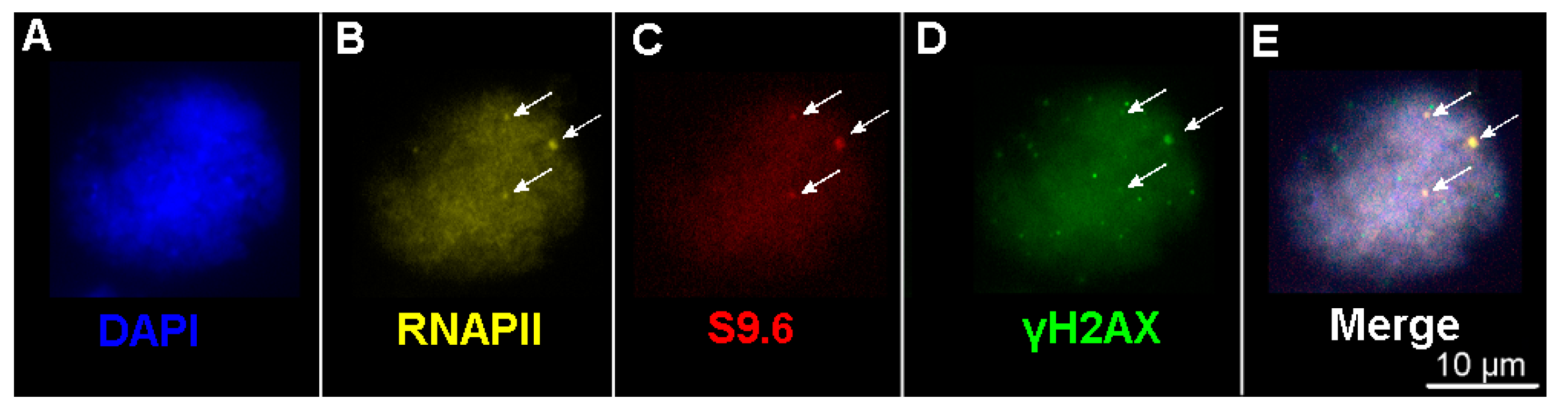
2.4. The Number of DSBs and Transcription Sites in Interphase Nuclei in Plants of the Terra Rossa Microsite Is Higher Than in Plants of the Basalt Microsite
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Total RNA Isolation and Reverse Transcription
4.3. Real-Time Quantitative PCR and Primer Design
4.4. Preparation of Cytological Slides
4.5. Fluorescence Immunolocalization
4.6. Microscopy and Image Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| BRCA1 | Breast cancer susceptibility gene 1 |
| DAPI | 4′,6-diamidino-2-phenylindole |
| DDR | DNA Damage Response |
| HR | Homologous Recombination |
| KU70 | X-ray cross complementing 6 |
| LigIV | DNA ligase 4 |
| MLH1 | MutL homolog 1 |
| MMR | Mismatch Repair |
| MRE11 | Meiotic Recombination 11 |
| MSH2 | MutS homolog 2 |
| NHEJ | Non-Homologous End Joining |
| RAD51 | Radiation sensitive 51 |
| RNAPII | RNA polymerase II |
| γH2AX | gamma-H2AX, phosphorylated H2AX |
References
- Soltis, D.E.; Soltis, P.S. Polyploidy: Recurrent formation and genome evolution. Trends Ecol. Evol. 1999, 14, 348–352. [Google Scholar] [CrossRef] [PubMed]
- Comai, L. The advantages and disadvantages of being polyploid. Nat. Rev. Genet. 2005, 6, 836–846. [Google Scholar] [CrossRef] [PubMed]
- Li, L.F.; Zhang, Z.B.; Wang, Z.H.; Li, N.; Sha, Y.; Wang, X.F.; Ding, N.; Li, Y.; Zhao, J.; Wu, Y.; et al. Genome sequences of five Sitopsis species of Aegilops and the origin of polyploid wheat B subgenome. Mol. Plant 2022, 15, 488–503. [Google Scholar] [CrossRef] [PubMed]
- Avni, R.; Lux, T.; Minz-Dub, A.; Millet, E.; Sela, H.; Distelfeld, A.; Deek, J.; Yu, G.; Steuernagel, B.; Pozniak, C.; et al. Genome sequences of three Aegilops species of the section Sitopsis reveal phylogenetic relationships and provide resources for wheat improvement. Plant J. 2022, 110, 179–192. [Google Scholar] [CrossRef]
- Levy, A.A.; Feldman, M. Evolution and origin of bread wheat. Plant Cell 2022, 34, 2549–2567. [Google Scholar] [CrossRef]
- Dvorak, J.; Akhunov, E.D. Tempos of gene locus deletions and duplications and their relationship to recombination rate during diploid and polyploid evolution in the Aegilops-Triticum alliance. Genetics 2005, 171, 323–332. [Google Scholar] [CrossRef] [Green Version]
- Özkan, H.; Willcox, G.; Graner, A.; Salamini, F.; Kilian, B. Geographic distribution and domestication of wild emmer wheat (Triticum dicoccoides). Genet. Resour. Crop Evol. 2011, 58, 11–53. [Google Scholar] [CrossRef]
- Feldman, M.; Levy, A.A. Allopolyploidy—A shaping force in the evolution of wheat genomes. Cytogenet. Genome Res. 2005, 109, 250–258. [Google Scholar] [CrossRef]
- Kimber, G.; Feldman, M. Wild Wheat, an Introduction; College of Agriculture University of Missouri Special Report: Columbia, MO, USA, 1987; Volume 353. [Google Scholar]
- Nevo, E. Evolution of wild emmer wheat and crop improvement. J. Syst. Evol. 2014, 52, 673–696. [Google Scholar] [CrossRef]
- Andersen, S.L.; Sekelsky, J. Meiotic versus mitotic recombination: Two different routes for double-strand break repair: The different functions of meiotic versus mitotic DSB repair are reflected in different pathway usage and different outcomes. BioEssays 2010, 32, 1058–1066. [Google Scholar] [CrossRef] [Green Version]
- Knoll, A.; Fauser, F.; Puchta, H. DNA recombination in somatic plant cells: Mechanisms and evolutionary consequences. Chromosom. Res. 2014, 22, 191–201. [Google Scholar] [CrossRef] [PubMed]
- Zeman, M.K.; Cimprich, K.A. Causes and consequences of replication stress. Nat. Cell Biol. 2014, 16, 2–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petermann, E.; Lan, L.; Zou, L. Sources, resolution and physiological relevance of R-loops and RNA–DNA hybrids. Nat. Rev. Mol. Cell Biol. 2022, 23, 521–540. [Google Scholar] [CrossRef]
- Zheng, D.; Li, M.; Yang, Y.; Huang, R.; Zhang, W. R-loops: Emerging key regulators in plants. J. Exp. Bot. 2022, erac433. [Google Scholar] [CrossRef] [PubMed]
- Yuan, W.; Zhou, J.; Tong, J.; Zhuo, W.; Wang, L.; Li, Y.; Sun, Q.; Qian, W. ALBA protein complex reads genic R-loops to maintain genome stability in Arabidopsis. Sci. Adv. 2019, 5, eaav9040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, W.; Li, K.; Li, S.; Hou, Q.; Zhang, Y.; Liu, K.; Sun, Q. The R-loop atlas of Arabidopsis development and responses to environmental stimuli. Plant Cell 2020, 32, 888–903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niehrs, C.; Luke, B. Regulatory R-loops as facilitators of gene expression and genome stability. Nat. Rev. Mol. Cell Biol. 2020, 21, 167–178. [Google Scholar] [CrossRef] [PubMed]
- Crossley, M.P.; Bocek, M.; Cimprich, K.A. R-Loops as Cellular Regulators and Genomic Threats. Mol. Cell 2019, 73, 398–411. [Google Scholar] [CrossRef] [Green Version]
- Spampinato, C.P. Protecting DNA from errors and damage: An overview of DNA repair mechanisms in plants compared to mammals. Cell. Mol. Life Sci. 2017, 74, 1693–1709. [Google Scholar] [CrossRef] [Green Version]
- Manova, V.; Gruszka, D. DNA damage and repair in plants–From models to crops. Front. Plant Sci. 2015, 6, 885. [Google Scholar] [CrossRef] [Green Version]
- Jackson, S.P.; Bartek, J. The DNA-damage response in human biology and disease. Nature 2009, 461, 1071–1078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nisa, M.; Huang, Y.; Benhamed, M.; Raynaud, C. The Plant DNA Damage Response: Signaling Pathways Leading to Growth Inhibition and Putative Role in Response to Stress Conditions. Front. Plant Sci. 2019, 10, 653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia, V.; Bruchet, H.; Camescasse, D.; Granier, F.; Bouchez, D.; Tissier, A. AtATM is essential for meiosis and the somatic response to DNA damage in plants. Plant Cell 2003, 15, 119–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Culligan, K.; Tissier, A.; Britt, A. ATR regulates a G2-phase cell-cycle checkpoint in Arabidopsis thaliana. Plant Cell 2004, 16, 1091–1104. [Google Scholar] [CrossRef] [Green Version]
- Amiard, S.; Charbonnel, C.; Allain, E.; Depeiges, A.; White, C.I.; Gallego, M.E. Distinct roles of the ATR kinase and the Mre11-Rad50-Nbs1 complex in the maintenance of chromosomal stability in Arabidopsis. Plant Cell 2010, 22, 3020–3033. [Google Scholar] [CrossRef] [Green Version]
- Löbrich, M.; Shibata, A.; Beucher, A.; Fisher, A.; Ensminger, M.; Goodarzi, A.A.; Barton, O.; Jeggo, P.A. γH2AX foci analysis for monitoring DNA double-strand break repair: Strengths, limitations and optimization. Cell Cycle 2010, 9, 662–669. [Google Scholar] [CrossRef] [Green Version]
- Waterworth, W.M.; Wilson, M.; Wang, D.; Nuhse, T.; Warward, S.; Selley, J.; West, C.E. Phosphoproteomic analysis reveals plant DNA damage signalling pathways with a functional role for histone H2AX phosphorylation in plant growth under genotoxic stress. Plant J. 2019, 100, 1007–1021. [Google Scholar] [CrossRef] [Green Version]
- Yoshiyama, K.O.; Kobayashi, J.; Ogita, N.; Ueda, M.; Kimura, S.; Maki, H.; Umeda, M. ATM-mediated phosphorylation of SOG1 is essential for the DNA damage response in Arabidopsis. EMBO Rep. 2013, 14, 817–822. [Google Scholar] [CrossRef] [Green Version]
- Yoshiyama, K.O.; Conklin, P.A.; Huefner, N.D.; Britt, A.B. Suppressor of gamma response 1 (SOG1) encodes a putative transcription factor governing multiple responses to DNA damage. Proc. Natl. Acad. Sci. USA 2009, 106, 12843–12848. [Google Scholar] [CrossRef] [Green Version]
- Puchta, H. The repair of double-strand breaks in plants: Mechanisms and consequences for genome evolution. J. Exp. Bot. 2005, 56, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Neal, J.A.; Meek, K. Choosing the right path: Does DNA-PK help make the decision? Mutat. Res.-Fundam. Mol. Mech. Mutagen. 2011, 711, 73–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spies, M.; Fishel, R. Mismatch repair and homeologous recombination. DNA Repair 2016, 38, 75–83. [Google Scholar] [CrossRef] [Green Version]
- Rabinovich-Vin, A.; Orshan, G. Ecological studies on the vegetation of the upper galilee, israel. II. factors determining the absence of batha and garigue components on middle eocenian strata. Isr. J. Bot. 1974, 23, 111–119. [Google Scholar]
- Nevo, E.; Beiles, A.; Krugman, T. Natural selection of allozyme polymorphisms: A microgeographic climatic differentiation in wild emmer wheat (Triticum dicoccoides). Theor. Appl. Genet. 1988, 75, 529–538. [Google Scholar] [CrossRef]
- Hatchi, E.; Skourti-Stathaki, K.; Ventz, S.; Pinello, L.; Yen, A.; Kamieniarz-Gdula, K.; Dimitrov, S.; Pathania, S.; McKinney, K.M.; Eaton, M.L.; et al. BRCA1 recruitment to transcriptional pause sites is required for R-loop-driven DNA damage repair. Mol. Cell 2015, 57, 636–647. [Google Scholar] [CrossRef] [Green Version]
- Lafarge, S.; Montané, M.H. Characterization of Arabidopsis thaliana ortholog of the human breast cancer susceptibility gene 1: AtBRCA1, strongly induced by gamma rays. Nucleic Acids Res. 2003, 31, 1148–1155. [Google Scholar] [CrossRef] [Green Version]
- Shinohara, M.; Hayashihara, K.; Grubb, J.T.; Bishop, D.K.; Shinohara, A. DNA damage response clamp 9-1-1 promotes assembly of ZMM proteins for formation of crossovers and synaptonemal complex. J. Cell Sci. 2015, 128, 1494–1506. [Google Scholar] [CrossRef] [Green Version]
- Melamed-Bessudo, C.; Shilo, S.; Levy, A.A. Meiotic recombination and genome evolution in plants. Curr. Opin. Plant Biol. 2016, 30, 82–87. [Google Scholar] [CrossRef]
- Pérez, R.; Cuadrado, Á.; Jouve, N.; de Bustos, A. Characterization of the Nbs1 gene and analysis of the expression of homologous and homoeologous MRN complex genes in meiocytes and somatic cells of different wheat species. Int. J. Plant Sci. 2011, 172, 959–969. [Google Scholar] [CrossRef]
- Wang, X.; Chen, Z.H.; Yang, C.; Zhang, X.; Jin, G.; Chen, G.; Wang, Y.; Holford, P.; Nevo, E.; Zhang, G.; et al. Genomic adaptation to drought in wild barley is driven by edaphic natural selection at the Tabigha Evolution Slope. Proc. Natl. Acad. Sci. USA 2018, 115, 5223–5228. [Google Scholar] [CrossRef] [Green Version]
- Cai, S.; Shen, Q.; Huang, Y.; Han, Z.; Wu, D.; Chen, Z.H.; Nevo, E.; Zhang, G. Multi-Omics Analysis Reveals the Mechanism Underlying the Edaphic Adaptation in Wild Barley at Evolution Slope (Tabigha). Adv. Sci. 2021, 8, 2101374. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Yin, H.; Jiao, C.; Fang, X.; Wang, G.; Li, G.; Ni, F.; Li, P.; Su, P.; Ge, W.; et al. Sympatric speciation of wild emmer wheat driven by ecology and chromosomal rearrangements. Proc. Natl. Acad. Sci. USA 2020, 117, 5955–5963. [Google Scholar] [CrossRef] [PubMed]
- Anand, R.P.; Tsaponina, O.; Greenwell, P.W.; Lee, C.S.; Du, W.; Petes, T.D.; Haber, J.E. Chromosome rearrangements via template switching between diverged repeated sequences. Genes Dev. 2014, 28, 2394–2406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gorbunova, V.; Levy, A.A. Non-homologous DNA end joining in plant cells is associated with deletions and filler DNA insertions. Nucleic Acids Res. 1997, 25, 4650–4657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wicker, T.; Buchmann, J.P.; Keller, B. Patching gaps in plant genomes results in gene movement and erosion of colinearity. Genome Res. 2010, 20, 1229–1237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raskina, O. Transposable Elements in the Organization and Diversification of the Genome of Aegilops speltoides Tausch (Poaceae, Triticeae). Int. J. Genom. 2018, 2018, 4373089. [Google Scholar] [CrossRef] [Green Version]
- Pollak, Y.; Zelinger, E.; Raskina, O. Repetitive DNA in the Architecture, Repatterning, and Diversification of the Genome of Aegilops speltoides Tausch (Poaceae, Triticeae). Front. Plant Sci. 2018, 9, 1779. [Google Scholar] [CrossRef]
- Raskina, O. Genotype- and Cell-Specific Dynamics of Tandem Repeat Patterns in Aegilops speltoides Tausch (Poaceae, Triticeae). Cytogenet. Genome Res. 2017, 153, 105–116. [Google Scholar] [CrossRef]
- Blackford, A.N.; Stucki, M. How Cells Respond to DNA Breaks in Mitosis. Trends Biochem. Sci. 2020, 45, 321–331. [Google Scholar] [CrossRef]
- Minocherhomji, S.; Ying, S.; Bjerregaard, V.A.; Bursomanno, S.; Aleliunaite, A.; Wu, W.; Mankouri, H.W.; Shen, H.; Liu, Y.; Hickson, I.D. Replication stress activates DNA repair synthesis in mitosis. Nature 2015, 528, 286–290. [Google Scholar] [CrossRef]
- Wassing, I.E.; Graham, E.; Saayman, X.; Rampazzo, L.; Ralf, C.; Bassett, A.; Esashi, F. The RAD51 recombinase protects mitotic chromatin in human cells. Nat. Commun. 2021, 12, 5380. [Google Scholar] [CrossRef] [PubMed]
- Audrey, A.; de Haan, L.; van Vugt, M.A.T.M.; de Boer, H.R. Processing DNA lesions during mitosis to prevent genomic instability. Biochem. Soc. Trans. 2022, 50, 1105–1118. [Google Scholar] [CrossRef]
- Godinez, V.G.; Kabbara, S.; Sherman, A.; Wu, T.; Cohen, S.; Kong, X.; Maravillas-Montero, J.L.; Shi, Z.; Preece, D.; Yokomori, K.; et al. DNA damage induced during mitosis undergoes DNA repair synthesis. PLoS ONE 2020, 15, e0227849. [Google Scholar] [CrossRef]
- Gomez-Godinez, V.; Wu, T.; Sherman, A.J.; Lee, C.S.; Liaw, L.H.; Zhongsheng, Y.; Yokomori, K.; Berns, M.W. Analysis of DNA double-strand break response and chromatin structure in mitosis using laser microirradiation. Nucleic Acids Res. 2010, 38, e202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakamoto, Y.; Kokuta, T.; Teshigahara, A.; Iijima, K.; Kitao, H.; Takata, M.; Tauchi, H. Mitotic cells can repair DNA double-strand breaks via a homology-directed pathway. J. Radiat. Res. 2021, 62, 25–33. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, Q.; Su, H.; Liu, K.; Xiao, X.; Li, W.; Sun, Q.; Birchler, J.A.; Han, F. Genome-wide mapping reveals R-loops associated with centromeric repeats in maize. Genome Res. 2021, 31, 1409–1418. [Google Scholar] [CrossRef]
- García-Muse, T.; Aguilera, A. R Loops: From Physiological to Pathological Roles. Cell 2019, 179, 604–618. [Google Scholar] [CrossRef]
- Cristini, A.; Ricci, G.; Britton, S.; Salimbeni, S.; Huang, S.; Yin, N.; Marinello, J.; Calsou, P.; Pommier, Y.; Favre, G.; et al. Dual Processing of R-Loops and Topoisomerase I Induces Transcription-Dependent DNA Double-Strand Breaks. Cell Rep. 2019, 28, 3167–3181. [Google Scholar] [CrossRef] [Green Version]
- Kabeche, L.; Nguyen, H.D.; Buisson, R. Zo A mitosis-specific and R loop–driven ATR pathway promotes faithful chromosome segregation. Science 2018, 359, 108–114. [Google Scholar] [CrossRef] [Green Version]
- Xu, W.; Xu, H.; Li, K.; Fan, Y.; Liu, Y.; Yang, X.; Sun, Q. The R-loop is a common chromatin feature of the Arabidopsis genome. Nat. Plants 2017, 3, 704–714. [Google Scholar] [CrossRef]
- Zeng, C.; Onoguchi, M.; Hamada, M. Association analysis of repetitive elements and R-loop formation across species. Mob. DNA 2021, 12, 3. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Su, H.; Zhang, J.; Liu, Y.; Feng, C.; Han, F. Back-spliced RNA from retrotransposon binds to centromere and regulates centromeric chromatin loops in maize. PLoS Biol. 2020, 18, e3000582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wicker, T.; Gundlach, H.; Spannagl, M.; Uauy, C.; Borrill, P.; Ramírez-González, R.H.; De Oliveira, R.; Mayer, K.F.X.; Paux, E.; Choulet, F. Impact of transposable elements on genome structure and evolution in bread wheat. Genome Biol. 2018, 19, 103. [Google Scholar] [CrossRef] [PubMed]
- Middleton, C.P.; Stein, N.; Keller, B.; Kilian, B.; Wicker, T. Comparative analysis of genome composition in Triticeae reveals strong variation in transposable element dynamics and nucleotide diversity. Plant J. 2013, 73, 347–356. [Google Scholar] [CrossRef] [PubMed]
- Raskina, O.; Belyayev, A.; Nevo, E. Repetitive DNAs of wild emmer wheat (Triticum dicoccoides) and their relation to S-genome species: Molecular cytogenetic analysis. Genome 2002, 45, 391–401. [Google Scholar] [CrossRef] [PubMed]
- Ruban, A.S.; Badaeva, E.D. Evolution of the S-Genomes in Triticum-Aegilops Alliance: Evidences From Chromosome Analysis. Front. Plant Sci. 2018, 9, 1756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raskina, O.; Barber, J.C.; Nevo, E.; Belyayev, A. Repetitive DNA and chromosomal rearrangements: Speciation-related events in plant genomes. Cytogenet. Genome Res. 2008, 120, 351–357. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Shams, I.; Raskina, O. Intraspecific and intraorganismal copy number dynamics of retrotransposons and tandem repeat in Aegilops speltoides Tausch (Poaceae, Triticeae). Protoplasma 2018, 255, 1023–1038. [Google Scholar] [CrossRef]
- Chelysheva, L.; Grandont, L.; Vrielynck, N.; le Guin, S.; Mercier, R.; Grelon, M. An Easy Protocol for Studying Chromatin and Recombination Protein Dynamics during Arabidopsis thaliana Meiosis: Immunodetection of Cohesins, Histones and MLH1. Cytogenet. Genome Res. 2010, 129, 143–153. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mean Number of Clusters per Chromosome Plate (SE) | |||||
|---|---|---|---|---|---|
| RAD51 | LigIV | γH2AX | RNAPII | S9.6 | |
| Basalt microsite | |||||
| Genotype | |||||
| B26 | 60.9 (±4.8) | 47.3 (±17.3) | 136.9 (±30.5) | NC | NC * |
| B60 | 97.0 (±13.3) | 73.8(±16.9) | 106.0 (±14.9) | NC | NC |
| B70 | 63.5 (±18.8) | 58.6(±9.3) | 83.1 (±0.7) | 4.5 (±0.7) | 0.8 (±0.9) |
| B81 | 56.3 (±10.1) | 127.4(±24.8) | 61.1 (±11.1) | 3.7 (±2.4) | 2.7 (±2.4) |
| B91 | 57.1 (±15.5) | 123.6 (±33.2) | 70.1 (±20.6) | NC | NC |
| average | 67.0 (±17.0) | 86.1 (±37.2) | 91.6 (±30.6) | 4.1 (±0.6) | 1.8 (±1.3) |
| Terra Rossa microsite | |||||
| Genotype | |||||
| TR34 | 72.9 (±7.1) | 56.8 (±7.2) | 127.1 (±47.2) | 4.5 (±2.8) | 1.5 (±1.8) |
| TR43 | 71.8 (±13.2) | 89.1 (±28.4) | 98.2 (±20.3) | NC | NC |
| TR52 | 67.4 (±9.8) | 54.5 (±10.6 | 119.0 (±24.6) | 5.6 (±2.3) | 3.2 (±2.2) |
| TR67 | 54.4 (±14.6) | 81.5 (±19.4) | 101.8 (±26.2) | NC | NC |
| TR82 | 56.6 (±12.6) | 111.3 (±22.9) | 122.7 (±30.7) | 9.1 (±4.4) | 6.4 (±2.4) |
| average | 64.6 (±8.6) | 78.7 (±23.7) | 113.8 (±12.9) | 6.4 (±2.4) | 3.7 (±2.5) |
| Number of Nuclei Analyzed | Mean Number of Clusters per Nucleus (SE) | |||
|---|---|---|---|---|
| γH2AX | RNAPII | S9.6 | ||
| Basalt microsite | ||||
| Genotype | ||||
| B26 | 56.0 | 53.4 | 2.7 | 1.5 |
| B60 | 72.0 | 31.7 | 2.9 | 2.5 |
| B70 | 53.0 | 26.6 | 6.4 | 2.7 |
| B81 | 52.0 | 52.8 | 3.2 | 2.3 |
| B91 | 60.0 | 60.3 | 4.2 | 3.7 |
| average | 45.0 (±10.4) | 3.9 (±1.8) | 2.5 (±1.1) | |
| Terra Rossa microsite | ||||
| Genotype | ||||
| TR34 | 53.0 | 71.2 | 6.8 | 3.8 |
| TR43 | 52.0 | 100.6 | 4.8 | 4.0 |
| TR52 | 67.0 | 71.6 | 3.0 | 2.8 |
| TR67 | 61.0 | 93.1 | 4.7 | 3.7 |
| TR82 | 54.0 | 83.4 | 9.2 | 4.8 |
| average | 84.0 (±10.2) | 5.7 (±2.6) | 3.8 (±1.0) | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Raskina, O.; Shklyar, B.; Nevo, E. The Influence of Edaphic Factors on DNA Damage and Repair in Wild Wheat Triticum dicoccoides Körn. (Poaceae, Triticeae). Int. J. Mol. Sci. 2023, 24, 6847. https://doi.org/10.3390/ijms24076847
Raskina O, Shklyar B, Nevo E. The Influence of Edaphic Factors on DNA Damage and Repair in Wild Wheat Triticum dicoccoides Körn. (Poaceae, Triticeae). International Journal of Molecular Sciences. 2023; 24(7):6847. https://doi.org/10.3390/ijms24076847
Chicago/Turabian StyleRaskina, Olga, Boris Shklyar, and Eviatar Nevo. 2023. "The Influence of Edaphic Factors on DNA Damage and Repair in Wild Wheat Triticum dicoccoides Körn. (Poaceae, Triticeae)" International Journal of Molecular Sciences 24, no. 7: 6847. https://doi.org/10.3390/ijms24076847