
Local Microtubule and F-Actin Distributions Fully Constrain the Spatial Geometry of Drosophila Sensory Dendritic Arbors

Abstract
:1. Introduction
2. Results
2.1. Distinct Dendritic Architecture of Sensory Neurons
2.2. Influence of Local Cytoskeleton Composition on Branch Orientation and Straightness
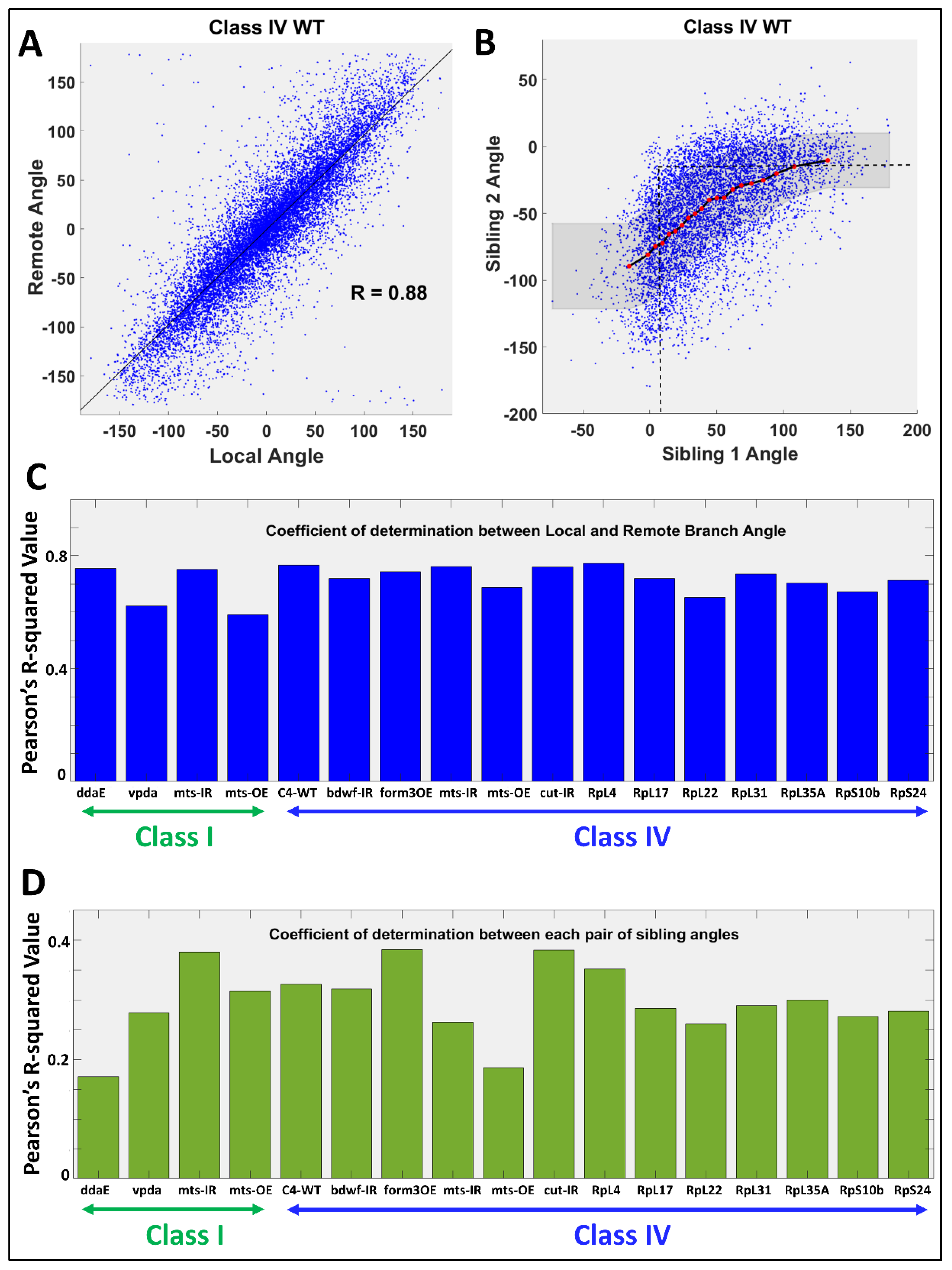
2.3. Relation between Angular Deviations of Sibling Branches
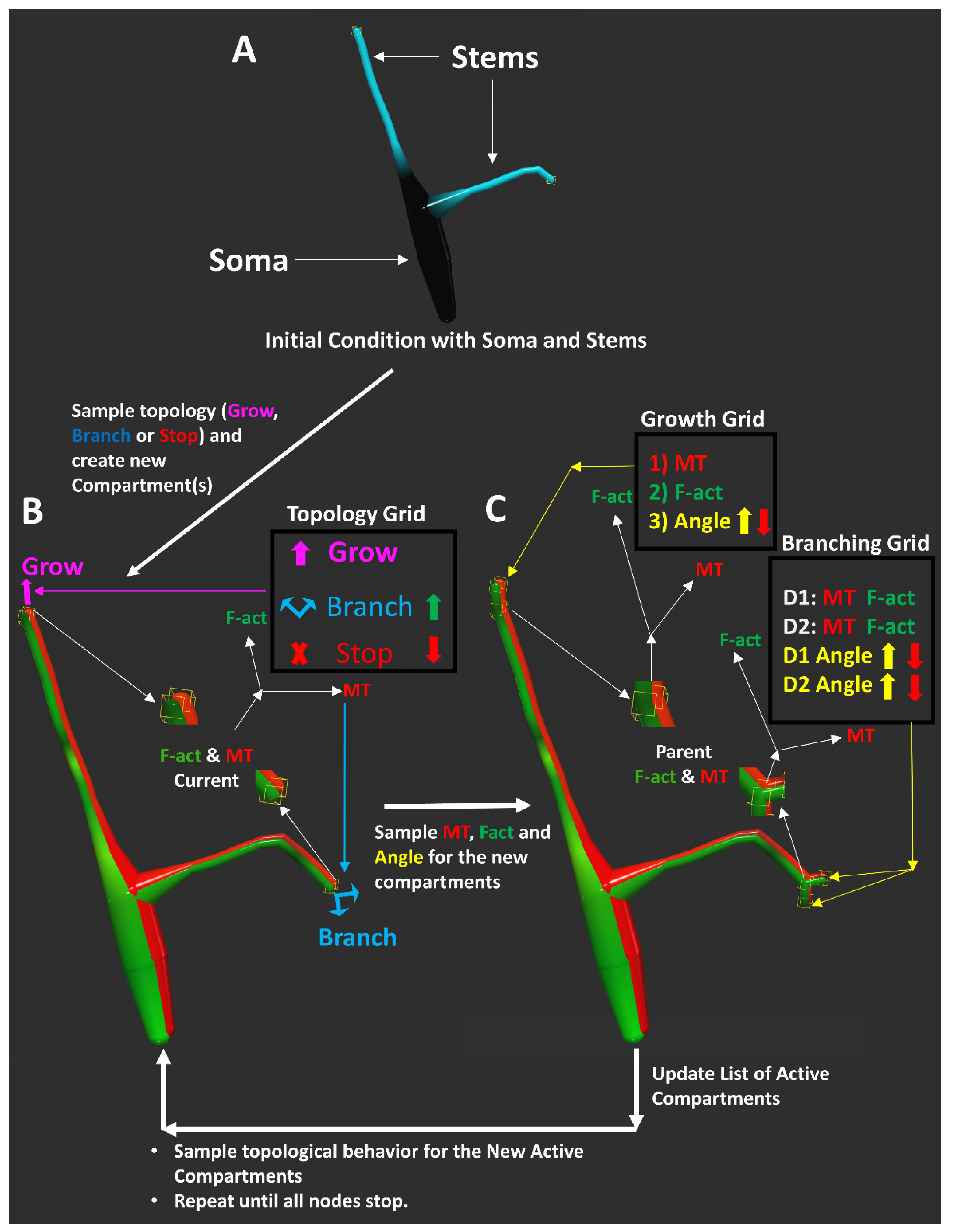
2.4. A Cytoskeleton-Driven Generative Model of Dendritic Morphology
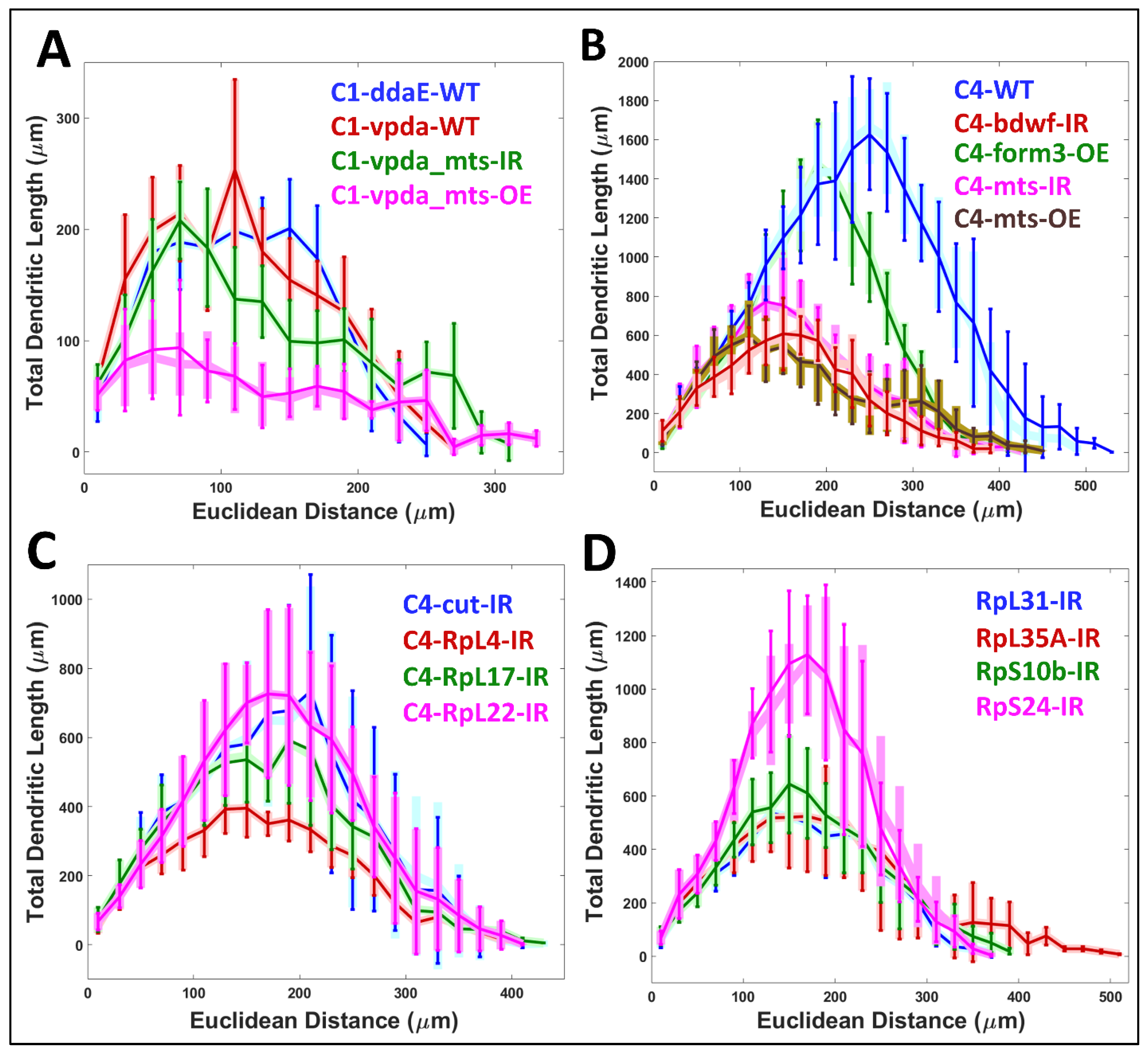
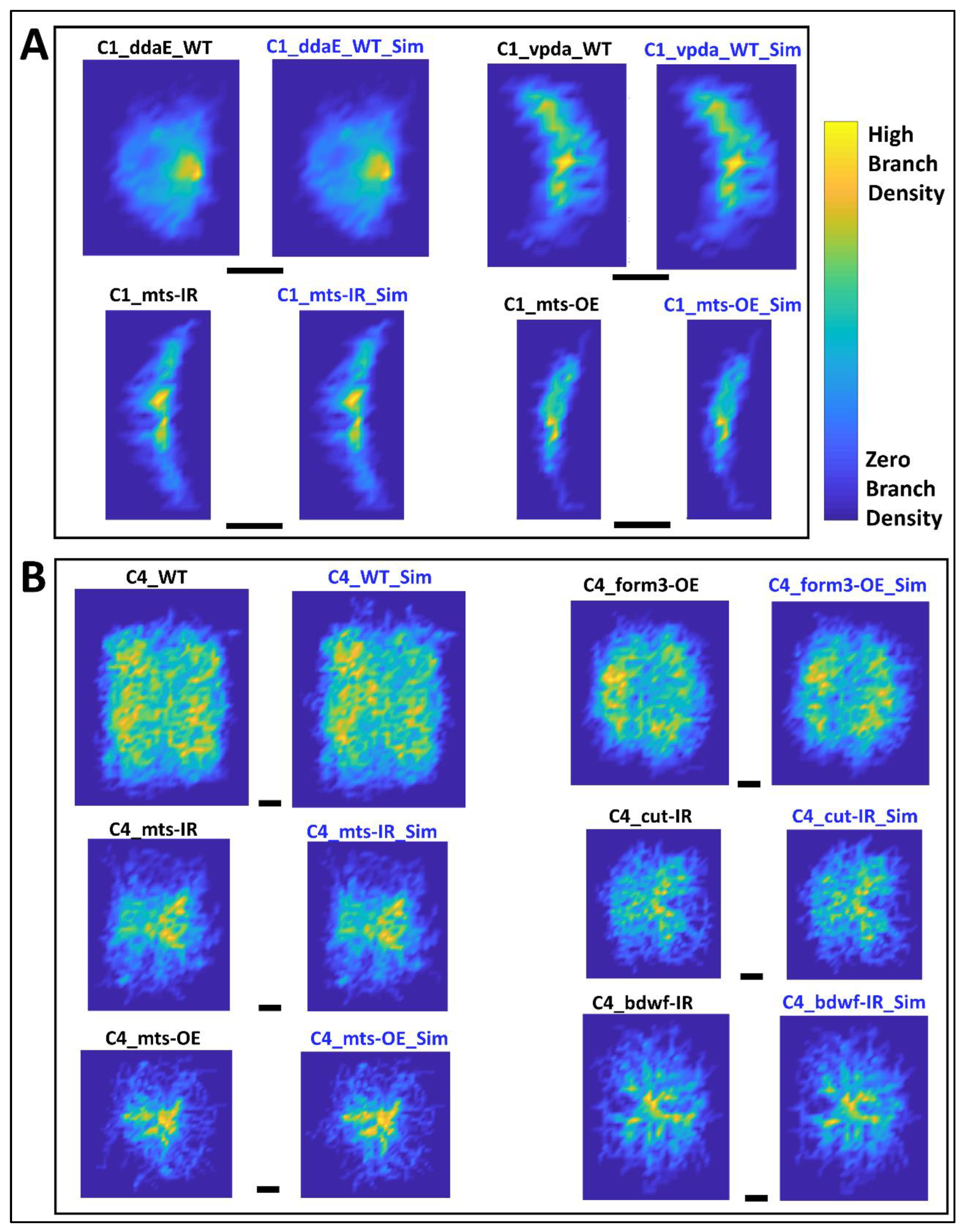
2.5. Simulated Neurons Reproduce all Morphological Attributes across Neuron Groups
3. Discussion
4. Materials and Methods
4.1. Drosophila Strains and Live Confocal Imaging
4.2. Morphological Reconstruction and Editing
4.3. Measurement and Correlation of Morphological and Cytoskeletal Properties
4.4. Model of Dendritic Arbor Generation
4.5. Comparison of Real and Simulated Arbor Properties
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jan, Y.-N.; Jan, L.Y. Branching out: Mechanisms of Dendritic Arborization. Nat. Rev. Neurosci. 2010, 11, 316–328. [Google Scholar] [CrossRef] [PubMed]
- Parekh, R.; Ascoli, G.A. Quantitative Investigations of Axonal and Dendritic Arbors. Neuroscientist 2015, 21, 241–254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lefebvre, J.L.; Sanes, J.R.; Kay, J.N. Development of Dendritic Form and Function. Annu. Rev. Cell Dev. Biol. 2015, 31, 741–777. [Google Scholar] [CrossRef] [PubMed]
- Lefebvre, J.L. Molecular Mechanisms That Mediate Dendrite Morphogenesis. Curr. Top. Dev. Biol. 2021, 142, 233–282. [Google Scholar] [CrossRef] [PubMed]
- Falke, E.; Nissanov, J.; Mitchell, T.W.; Bennett, D.A.; Trojanowski, J.Q.; Arnold, S.E. Subicular Dendritic Arborization in Alzheimer’s Disease Correlates with Neurofibrillary Tangle Density. Am. J. Pathol. 2003, 163, 1615–1621. [Google Scholar] [CrossRef]
- Emoto, K. Dendrite Remodeling in Development and Disease. Dev. Growth Differ. 2011, 53, 277–286. [Google Scholar] [CrossRef] [PubMed]
- Forrest, M.P.; Parnell, E.; Penzes, P. Dendritic Structural Plasticity and Neuropsychiatric Disease. Nat. Rev. Neurosci. 2018, 19, 215–234. [Google Scholar] [CrossRef]
- Brown, K.; Gillette, T.; Ascoli, G. Quantifying Neuronal Size: Summing up Trees and Splitting the Branch Difference. Semin. Cell Dev. Biol. 2008, 19, 485–493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Komendantov, A.O.; Ascoli, G.A. Dendritic Excitability and Neuronal Morphology as Determinants of Synaptic Efficacy. J. Neurophysiol. 2009, 101, 1847–1866. [Google Scholar] [CrossRef] [Green Version]
- Stiso, J.; Bassett, D.S. Spatial Embedding Imposes Constraints on Neuronal Network Architectures. Trends Cogn. Sci. 2018, 22, 1127–1142. [Google Scholar] [CrossRef]
- Ropireddy, D.; Ascoli, G.A. Potential Synaptic Connectivity of Different Neurons onto Pyramidal Cells in a 3D Reconstruction of the Rat Hippocampus. Front. Neuroinform. 2011, 5, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferreira Castro, A.; Baltruschat, L.; Stürner, T.; Bahrami, A.; Jedlicka, P.; Tavosanis, G.; Cuntz, H. Achieving Functional Neuronal Dendrite Structure through Sequential Stochastic Growth and Retraction. Elife 2020, 9, e60920. [Google Scholar] [CrossRef]
- Nanda, S.; Das, R.; Bhattacharjee, S.; Cox, D.N.; Ascoli, G.A. Morphological Determinants of Dendritic Arborization Neurons in Drosophila Larva. Brain Struct. Funct. 2018, 223, 1107–1120. [Google Scholar] [CrossRef]
- Das, R.; Bhattacharjee, S.; Patel, A.A.; Harris, J.M.; Bhattacharya, S.; Letcher, J.M.; Clark, S.G.; Nanda, S.; Iyer, E.P.R.; Ascoli, G.A.; et al. Dendritic Cytoskeletal Architecture Is Modulated by Combinatorial Transcriptional Regulation in Drosophila Melanogaster. Genetics 2017, 207, 1401–1421. [Google Scholar] [CrossRef] [Green Version]
- Bhattacharjee, S.; Lottes, E.N.; Nanda, S.; Golshir, A.; Patel, A.A.; Ascoli, G.A.; Cox, D.N. PP2A Phosphatase Regulates Cell-Type Specific Cytoskeletal Organization to Drive Dendrite Diversity. Front. Mol. Neurosci. 2022, 15, 926567. [Google Scholar] [CrossRef] [PubMed]
- Franker, M.; Hoogenraad, C. Microtubule-Based Transport—Basic Mechanisms, Traffic Rules and Role in Neurological Pathogenesis. J. Cell Sci. 2013, 126, 2319–2329. [Google Scholar] [CrossRef] [Green Version]
- Ledda, F.; Paratcha, G. Mechanisms Regulating Dendritic Arbor Patterning. Cell. Mol. Life Sci. 2017, 74, 4511–4537. [Google Scholar] [CrossRef]
- Nagel, J.C.; Delandre, C.; Zhang, Y.; Förstner, F.; Moore, A.W.; Tavosanis, G. Fascin Controls Neuronal Class-Specific Dendrite Arbor Morphology. Development 2012, 139, 2999–3009. [Google Scholar] [CrossRef] [Green Version]
- Nanda, S.; Bhattacharjee, S.; Cox, D.N.; Ascoli, G.A. Distinct Relations of Microtubules and Actin Filaments with Dendritic Architecture. iScience 2020, 23, 101865. [Google Scholar] [CrossRef] [PubMed]
- Coles, C.H.; Bradke, F. Coordinating Neuronal Actin-Microtubule Dynamics. Curr. Biol. 2015, 25, R677–R691. [Google Scholar] [CrossRef] [Green Version]
- Kapitein, L.C.; Hoogenraad, C.C. Building the Neuronal Microtubule Cytoskeleton. Neuron 2015, 87, 492–506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corty, M.M.; Tam, J.; Grueber, W.B. Dendritic Diversification through Transcription-Factor Mediated Suppression of Alternative Morphologies. Development 2016, 143, 1351–1362. [Google Scholar] [CrossRef] [Green Version]
- Das, R.; Bhattacharjee, S.; Letcher, J.M.; Harris, J.M.; Nanda, S.; Foldi, I.; Lottes, E.N.; Bobo, H.M.; Grantier, B.D.; Mihály, J.; et al. Formin 3 Directs Dendritic Architecture via Microtubule Regulation and Is Required for Somatosensory Nociceptive Behavior. Development 2021, 148, dev187609. [Google Scholar] [CrossRef] [PubMed]
- de la Torre-Ubieta, L.; Bonni, A. Transcriptional Regulation of Neuronal Polarity and Morphogenesis in the Mammalian Brain. Neuron 2011, 72, 22–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iyer, S.C.; Wang, D.; Iyer, E.P.R.; Trunnell, S.A.; Meduri, R.; Shinwari, R.; Sulkowski, M.J.; Cox, D.N. The Rhogef Trio Functions in Sculpting Class Specific Dendrite Morphogenesis in Drosophila Sensory Neurons. PLoS ONE 2012, 7, e33634. [Google Scholar] [CrossRef] [Green Version]
- Nanda, S.; Das, R.; Cox, D.N.; Ascoli, G.A. Structural Plasticity in Dendrites: Developmental Neurogenetics, Morphological Reconstructions, and Computational Modeling. In Neurobiological and Psychological Aspects of Brain Recovery; Petrosini, L., Ed.; Contemporary Clinical Neuroscience Series; Springer: Cham, Switzerland, 2017; Volume 30, pp. 1–34. [Google Scholar]
- Hely, T.A.; Graham, B.; Van Ooyen, A. A Computational Model of Dendrite Elongation and Branching Based on MAP2 Phosphorylation. J. Theor. Biol. 2001, 210, 375–384. [Google Scholar] [CrossRef] [Green Version]
- Fernandez-Gonzalez, P.; Benavides-Piccione, R.; Leguey, I.; Bielza, C.; Larrañaga, P.; DeFelipe, J. Dendritic-Branching Angles of Pyramidal Neurons of the Human Cerebral Cortex. Brain Struct. Funct. 2017, 222, 1847–1859. [Google Scholar] [CrossRef] [Green Version]
- Nanda, S.; Chen, H.; Das, R.; Bhattacharjee, S.; Cuntz, H.; Torben-Nielsen, B.; Peng, H.; Cox, D.N.; Schutter, E.D.; Ascoli, G.A. Design and Implementation of Multi-Signal and Time-Varying Neural Reconstructions. Sci. Data 2018, 5, 170207. [Google Scholar] [CrossRef] [Green Version]
- Bhattacharjee, S.; Iyer, E.P.R.; Iyer, S.C.; Nanda, S.; Rubaharan, M.; Ascoli, G.A.; Cox, D.N. The Zinc-BED Transcription Factor Bedwarfed Promotes Proportional Dendritic Growth and Branching through Transcriptional and Translational Regulation in Drosophila. bioRxiv 2023, 6344. [Google Scholar] [CrossRef]
- Nanda, S.; Bhattacharjee, S.; Cox, D.N.; Ascoli, G.A. Local microtubule and F-actin distributions fully determine the spatial geometry of Drosophila sensory dendritic arbors. bioRxiv 2023. [Google Scholar] [CrossRef]
- Samsonovich, A.V.; Ascoli, G.A. Morphological Homeostasis in Cortical Dendrites. Proc. Natl. Acad. Sci. USA 2006, 103, 1569–1574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Wang, D.; Ascoli, G.A.; Mitra, P.; Wang, Y. Metrics for Comparing Neuronal Tree Shapes Based on Persistent Homology. PLoS ONE 2017, 12, e0182184. [Google Scholar] [CrossRef] [Green Version]
- Cuntz, H.; Forstner, F.; Borst, A.; Häusser, M. One Rule to Grow Them All: A General Theory of Neuronal Branching and Its Practical Application. PLoS Comput. Biol. 2010, 6, e1000877. [Google Scholar] [CrossRef]
- Conde, C.; Cáceres, A. Microtubule Assembly, Organization and Dynamics in Axons and Dendrites. Nat. Rev. Neurosci. 2009, 10, 319–332. [Google Scholar] [CrossRef] [PubMed]
- Georges, P.C.; Hadzimichalis, N.M.; Sweet, E.S.; Firestein, B.L. The Yin–Yang of Dendrite Morphology: Unity of Actin and Microtubules. Mol. Neurobiol. 2008, 38, 270–284. [Google Scholar] [CrossRef]
- Sasaki, Y.; Cheng, C.; Uchida, Y.; Nakajima, O.; Ohshima, T.; Yagi, T.; Taniguchi, M.; Nakayama, T.; Kishida, R.; Kudo, Y.; et al. Fyn and Cdk5 Mediate Semaphorin-3A Signaling, Which Is Involved in Regulation of Dendrite Orientation in Cerebral Cortex. Neuron 2002, 35, 907–920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsui, A.; Tran, M.; Yoshida, A.C.; Kikuchi, S.S.; U, M.; Ogawa, M.; Shimogori, T. BTBD3 Controls Dendrite Orientation toward Active Axons in Mammalian Neocortex. Science 2013, 342, 1114–1118. [Google Scholar] [CrossRef] [Green Version]
- Samsonovich, A.V.; Ascoli, G.A. Statistical Morphological Analysis of Hippocampal Principal Neurons Indicates Cell-Specific Repulsion of Dendrites from Their Own Cell. J. Neurosci. Res. 2003, 71, 173–187. [Google Scholar] [CrossRef]
- Marks, W.B.; Burke, R.E. Simulation of Motoneuron Morphology in Three Dimensions. I. Building Individual Dendritic Trees. J. Comp. Neurol. 2007, 503, 685–700. [Google Scholar] [CrossRef]
- Donohue, D.E.; Ascoli, G.A. A Comparative Computer Simulation of Dendritic Morphology. PLoS Comput. Biol. 2008, 4, e1000089. [Google Scholar] [CrossRef]
- Bielza, C.; Benavides-Piccione, R.; López-Cruz, P.; Larrañaga, P.; Defelipe, J. Branching Angles of Pyramidal Cell Dendrites Follow Common Geometrical Design Principles in Different Cortical Areas. Sci. Rep. 2014, 4, 5909. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Green, E.J.; Juraska, J.M. The Dendritic Morphology of Hippocampal Dentate Granule Cells Varies with Their Position in the Granule Cell Layer: A Quantitative Golgi Study. Exp. Brain Res. 1985, 59, 582–586. [Google Scholar] [CrossRef] [PubMed]
- Schneider-Mizell, C.M.; Gerhard, S.; Longair, M.; Kazimiers, T.; Li, F.; Zwart, M.F.; Champion, A.; Midgley, F.M.; Fetter, R.D.; Saalfeld, S.; et al. Quantitative Neuroanatomy for Connectomics in Drosophila. Elife 2016, 5, e12059. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nithianandam, V.; Chien, C.-T. Actin Blobs Prefigure Dendrite Branching Sites. J. Cell Biol. 2018, 217, 3731–3746. [Google Scholar] [CrossRef] [Green Version]
- Stürner, T.; Tatarnikova, A.; Mueller, J.; Schaffran, B.; Cuntz, H.; Zhang, Y.; Nemethova, M.; Bogdan, S.; Small, V.; Tavosanis, G. Transient Localization of the Arp2/3 Complex Initiates Neuronal Dendrite Branching in Vivo. Development 2019, 146, dev171397. [Google Scholar] [CrossRef] [Green Version]
- Ritzenthaler, S.; Suzuki, E.; Chiba, A. Postsynaptic Filopodia in Muscle Cells Interact with Innervating Motoneuron Axons. Nat. Neurosci. 2000, 3, 1012–1017. [Google Scholar] [CrossRef]
- Stürner, T.; Ferreira Castro, A.; Philipps, M.; Cuntz, H.; Tavosanis, G. The Branching Code: A Model of Actin-Driven Dendrite Arborization. Cell Rep. 2022, 39, 110746. [Google Scholar] [CrossRef]
- Rada, L.; Kilic, B.; Erdil, E.; Ramiro-Cortés, Y.; Israely, I.; Unay, D.; Cetin, M.; Argunsah, A.Ö. Tracking-Assisted Detection of Dendritic Spines in Time-Lapse Microscopic Images. Neuroscience 2018, 394, 189–205. [Google Scholar] [CrossRef]
- He, H.Y.; Cline, H.T. Diadem X: Automated 4 Dimensional Analysis of Morphological Data. Neuroinformatics 2011, 9, 107–112. [Google Scholar] [CrossRef] [Green Version]
- Sheintuch, L.; Rubin, A.; Brande-Eilat, N.; Geva, N.; Sadeh, N.; Pinchasof, O.; Ziv, Y. Tracking the Same Neurons across Multiple Days in Ca2+ Imaging Data. Cell Rep. 2017, 21, 1102–1115. [Google Scholar] [CrossRef] [Green Version]
- Nanda, S.; Bhattacharjee, S.; Cox, D.N.; Ascoli, G.A. An Imaging Analysis Protocol to Trace, Quantify, and Model Multi-Signal Neuron Morphology. STAR Protoc. 2021, 2, 100567. [Google Scholar] [CrossRef] [PubMed]
- Gleeson, P.; Davison, A.P.; Silver, R.A.; Ascoli, G.A. A Commitment to Open Source in Neuroscience. Neuron 2017, 96, 964–965. [Google Scholar] [CrossRef] [Green Version]
- Akram, M.A.; Nanda, S.; Maraver, P.; Armañanzas, R.; Ascoli, G.A. An Open Repository for Single-Cell Reconstructions of the Brain Forest. Sci. Data 2018, 5, 180006. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, H.; Takasu, E.; Aigaki, T.; Kato, K.; Hayashi, S.; Nose, A. Formin3 Is Required for Assembly of the F-Actin Structure That Mediates Tracheal Fusion in Drosophila. Dev. Biol. 2004, 274, 413–425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Y.; Roesel, C.; Flockhart, I.; Perkins, L.; Perrimon, N.; Mohr, S.E. UP-TORR: Online Tool for Accurate and up-to-Date Annotation of RNAi Reagents. Genetics 2013, 195, 37–45. [Google Scholar] [CrossRef] [Green Version]
- Feng, L.; Zhao, T.; Kim, J. NeuTube 1.0: A New Design for Efficient Neuron Reconstruction Software Based on the SWC Format. eneuro 2015, 2, ENEURO.0049-14.2014. [Google Scholar] [CrossRef]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An Open-Source Platform for Biological-Image Analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scorcioni, R.; Polavaram, S.; Ascoli, G.A. L-Measure: A Web-Accessible Tool for the Analysis, Comparison and Search of Digital Reconstructions of Neuronal Morphologies. Nat. Protoc. 2008, 3, 866–876. [Google Scholar] [CrossRef] [PubMed]
- Bijari, K.; Valera, G.; López-Schier, H.; Ascoli, G.A. Quantitative Neuronal Morphometry by Supervised and Unsupervised Learning. STAR Protoc. 2021, 2, 100867. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Abbreviation | Neuron Class | Neuron Subtype | Genotype | Morphology Description | Number of Neurons | Source Publication |
|---|---|---|---|---|---|---|
| C1-ddaE-WT | Class I | ddaE | Wild type | Simple dendritic arbors, secondary arbors tend to extend towards one direction | 19 | [15] |
| C1-vpda-WT | Class I | vpda | Wild type | Simple dendritic arbors, secondary arbors tend to extend towards both direction | 8 | [15] |
| C1-vpda-mts-IR | Class I | vpda | mts-Knockdown | Increased terminal complexity | 9 | [15] |
| C1-vpda-mts-OE | Class I | vpda | mts-Overexpression | Reduced size, and complexity | 12 | [15] |
| C4_WT | Class IV | ddaC | Wild type | Large complex dendritic arbors | 10 | [15] |
| C4_bdwf-IR | Class IV | ddaC | CG3995/bdwf Knockdown | Reduced size and complexity | 8 | [30] |
| C4_form3-OE | Class IV | ddaC | Formin 3 Overexpression | Reduced size | 9 | [23] |
| C4_mts-IR | Class IV | ddaC | mts-Knockdown | Reduced size | 11 | [15] |
| C4_mts-OE | Class IV | ddaC | mts Overexpression | Reduced size | 10 | [15] |
| C4_cut_IR | Class IV | ddaC | cut Knockdown | Reduced size and terminal complexity | 8 | [30] |
| C4_RpL4-IR | Class IV | ddaC | RpL4-Knockdown | Reduced size and complexity | 10 | [30] |
| C4_RpL17-IR | Class IV | ddaC | RpL17-Knockdown | Reduced size and complexity | 6 | [31] |
| C4_RpL22-IR | Class IV | ddaC | RpL22-Knockdown | Reduced size and complexity | 10 | [30] |
| C4_RpL31-IR | Class IV | ddaC | RpL31-Knockdown | Reduced size and complexity | 9 | [30] |
| C4_RpL35A-IR | Class IV | ddaC | RpL35A-Knockdown | Reduced size and complexity | 13 | [31] |
| C4_RpS10b-IR | Class IV | ddaC | RpS10b-Knockdown | Reduced size and complexity | 8 | [31] |
| C4_RpS24-IR | Class IV | ddaC | RpS24-Knockdown | Reduced size and complexity | 7 | [30] |
| Neuron Groups | MT vs. Remote Angle | F-Act vs. Remote Angle | Integral MT vs. Remote Angle | MT vs. Branch Straightness | F-Act vs. Branch Straightness | Integral MT vs. Branch Straightness |
|---|---|---|---|---|---|---|
| C1-ddaE-WT | −0.80 | −0.68 | 0 | 0.64 | 0 | 0.64 |
| C1-vpda-WT | −0.40 | 0 | 0 | 0 | 0 | 0 |
| C1-vpda-mts-IR | −0.58 | 0 | 0 | 0 | 0 | 0 |
| C1-vpda-mts-OE | −0.23 | 0 | 0 | 0 | 0 | 0 |
| C4_WT | −0.82 | −0.88 | −0.51 | 0.72 | 0.84 | 0.11 |
| C4_bdwf-IR | −0.73 | 0 | 0 | 0.69 | 0.68 | 0 |
| C4_form3-OE | −0.71 | −0.88 | −0.67 | 0.66 | 0.86 | 0.83 |
| C4_mts-IR | −0.72 | −0.63 | 0 | 0 | 0.89 | 0 |
| C4_mts-OE | −0.88 | 0 | 0 | 0.82 | 0.61 | 0.46 |
| C4_cut_IR | −0.75 | 0.80 | 0 | 0.85 | 0 | 0.35 |
| C4_RpL4-IR | −0.72 | 0 | 0 | 0.63 | 0.74 | 0.53 |
| C4_RpL17-IR | −0.62 | 0 | −0.33 | 0.72 | 0.72 | 0.33 |
| C4_RpL22-IR | −0.14 | 0 | −0.38 | 0.00 | 0.53 | 0 |
| C4_RpL31-IR | −0.62 | 0 | 0 | 0.62 | 0 | 0.50 |
| C4_RpL35A-IR | −0.62 | 0 | −0.54 | 0.67 | 0.48 | 0.66 |
| C4_RpS10b-IR | −0.59 | −0.15 | 0 | 0.68 | 0.75 | 0.49 |
| C4_RpS24-IR | −0.80 | −0.14 | −0.46 | 0.68 | 0.55 | 0.30 |
| Neuron Groups | Number of Branches | Height | Width | Length | Max Path Distance | Max Euclidean Distance | Max Branch Order | Contraction | Partition Asymmetry | Local Bifurcation Angle |
|---|---|---|---|---|---|---|---|---|---|---|
| C1-ddaE-WT | 1 | 1 | 0.99 | 1 | 1 | 1 | 1 | 0.99 | 1 | 1 |
| C1-vpda-WT | 1 | 0.98 | 1 | 0.96 | 0.90 | 0.93 | 1 | 0.90 | 1 | 0.96 |
| C1-vpda-mts-IR | 1 | 1 | 1 | 1 | 0.98 | 0.97 | 1 | 1 | 1 | 0.93 |
| C1-vpda-mts-OE | 0.94 | 0.98 | 0.77 | 0.88 | 0.92 | 0.79 | 0.79 | 0.95 | 0.68 | 0.95 |
| C4_WT | 1 | 0.97 | 0.85 | 1 | 0.94 | 0.76 | 0.79 | 0.73 | 0.82 | 0.91 |
| C4_bdwf-IR | 1 | 0.90 | 0.94 | 0.94 | 0.90 | 0.86 | 0.90 | 0.99 | 0.66 | 1 |
| C4_form3-OE | 1 | 0.65 | 0.53 | 0.92 | 0.95 | 0.62 | 0.96 | 1 | 0.85 | 0.93 |
| C4_mts-IR | 1 | 0.98 | 0.96 | 1 | 0.99 | 1 | 0.74 | 0.98 | 0.91 | 0.97 |
| C4_mts-OE | 1 | 0.68 | 0.94 | 1 | 0.97 | 0.85 | 0.73 | 1 | 1 | 0.85 |
| C4_cut_IR | 1 | 0.82 | 0.90 | 0.78 | 0.98 | 0.93 | 0.94 | 0.96 | 0.94 | 0.99 |
| C4_RpL4-IR | 1 | 0.97 | 1 | 1 | 0.97 | 0.97 | 1 | 1 | 1 | 1 |
| C4_RpL17-IR | 1 | 0.66 | 0.85 | 0.92 | 0.97 | 1 | 1 | 0.94 | 0.91 | 1 |
| C4_RpL22-IR | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 0.91 | 0.68 | 0.94 |
| C4_RpL31-IR | 1 | 0.97 | 0.98 | 0.95 | 0.88 | 0.98 | 0.96 | 0.95 | 1 | 0.93 |
| C4_RpL35A-IR | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 0.95 |
| C4_RpS10b-IR | 1 | 1 | 0.98 | 0.96 | 1 | 0.93 | 1 | 0.94 | 0.96 | 0.96 |
| C4_RpS24-IR | 0.96 | 0.97 | 0.83 | 0.92 | 0.64 | 0.97 | 0.58 | 0.93 | 0.78 | 0.80 |
| Neuron Group | Persistence Vectors | Arbor Density |
|---|---|---|
| C1-ddaE-WT | 0.92 | 1 |
| C1-vpda-WT | 0.93 | 0.99 |
| C1-vpda-mts-IR | 0.93 | 0.99 |
| C1-vpda-mts-OE | 0.91 | 1 |
| C4_WT | 0.99 | 0.99 |
| C4_bdwf-IR | 0.94 | 1 |
| C4_form3-OE | 0.82 | 0.99 |
| C4_mts-IR | 0.88 | 1 |
| C4_mts-OE | 0.84 | 1 |
| C4_cut_IR | 0.91 | 1 |
| C4_RpL4-IR | 0.99 | 1 |
| C4_RpL17-IR | 0.72 | 0.99 |
| C4_RpL22-IR | 0.85 | 1 |
| C4_RpL31-IR | 0.82 | 1 |
| C4_RpL35A-IR | 0.87 | 1 |
| C4_RpS10b-IR | 0.98 | 1 |
| C4_RpS24-IR | 0.98 | 0.99 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nanda, S.; Bhattacharjee, S.; Cox, D.N.; Ascoli, G.A. Local Microtubule and F-Actin Distributions Fully Constrain the Spatial Geometry of Drosophila Sensory Dendritic Arbors. Int. J. Mol. Sci. 2023, 24, 6741. https://doi.org/10.3390/ijms24076741
Nanda S, Bhattacharjee S, Cox DN, Ascoli GA. Local Microtubule and F-Actin Distributions Fully Constrain the Spatial Geometry of Drosophila Sensory Dendritic Arbors. International Journal of Molecular Sciences. 2023; 24(7):6741. https://doi.org/10.3390/ijms24076741
Chicago/Turabian StyleNanda, Sumit, Shatabdi Bhattacharjee, Daniel N. Cox, and Giorgio A. Ascoli. 2023. "Local Microtubule and F-Actin Distributions Fully Constrain the Spatial Geometry of Drosophila Sensory Dendritic Arbors" International Journal of Molecular Sciences 24, no. 7: 6741. https://doi.org/10.3390/ijms24076741