
Photodegradable by Yellow-Orange Light degFusionRed Optogenetic Module with Autocatalytically Formed Chromophore
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
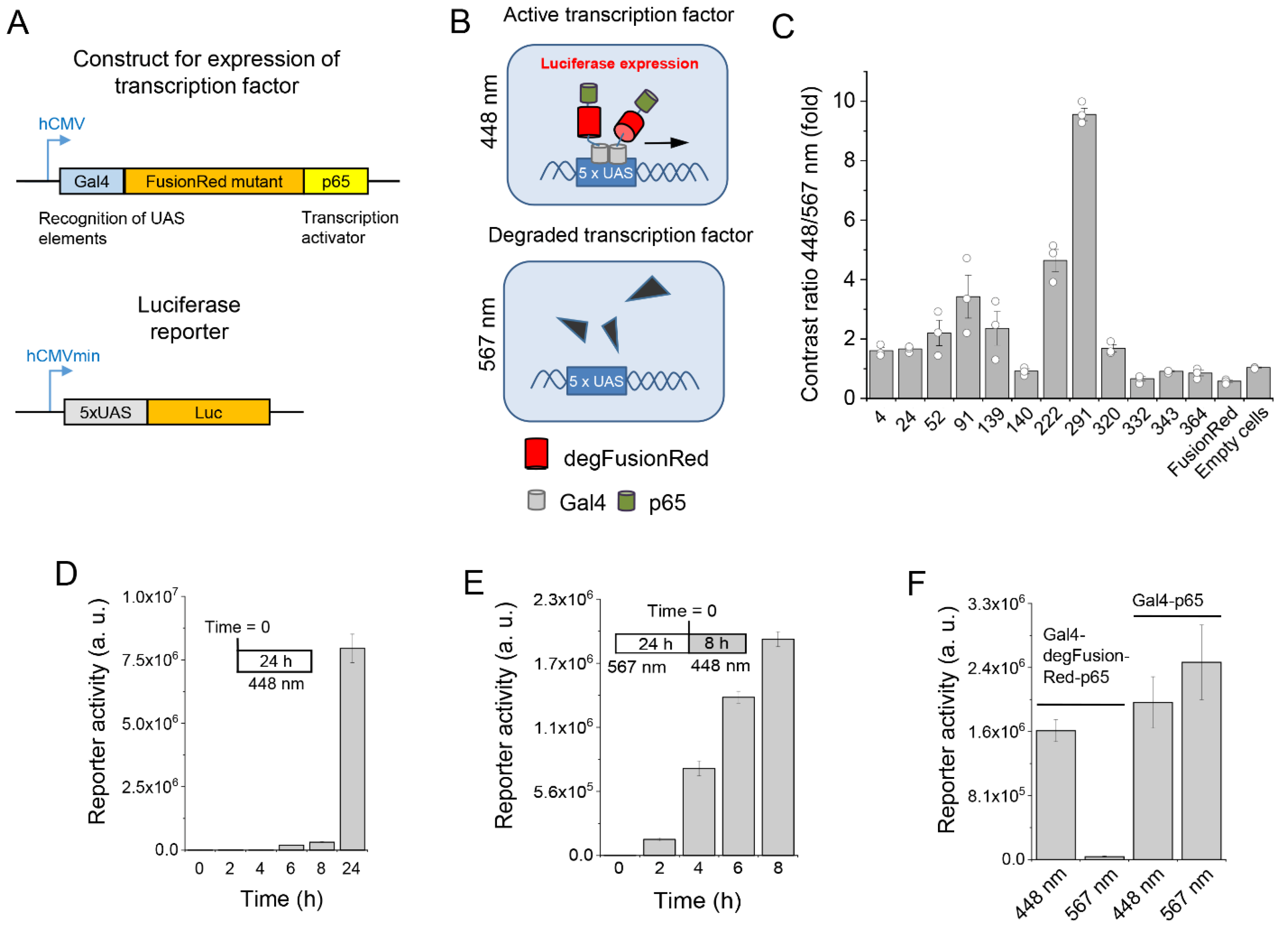
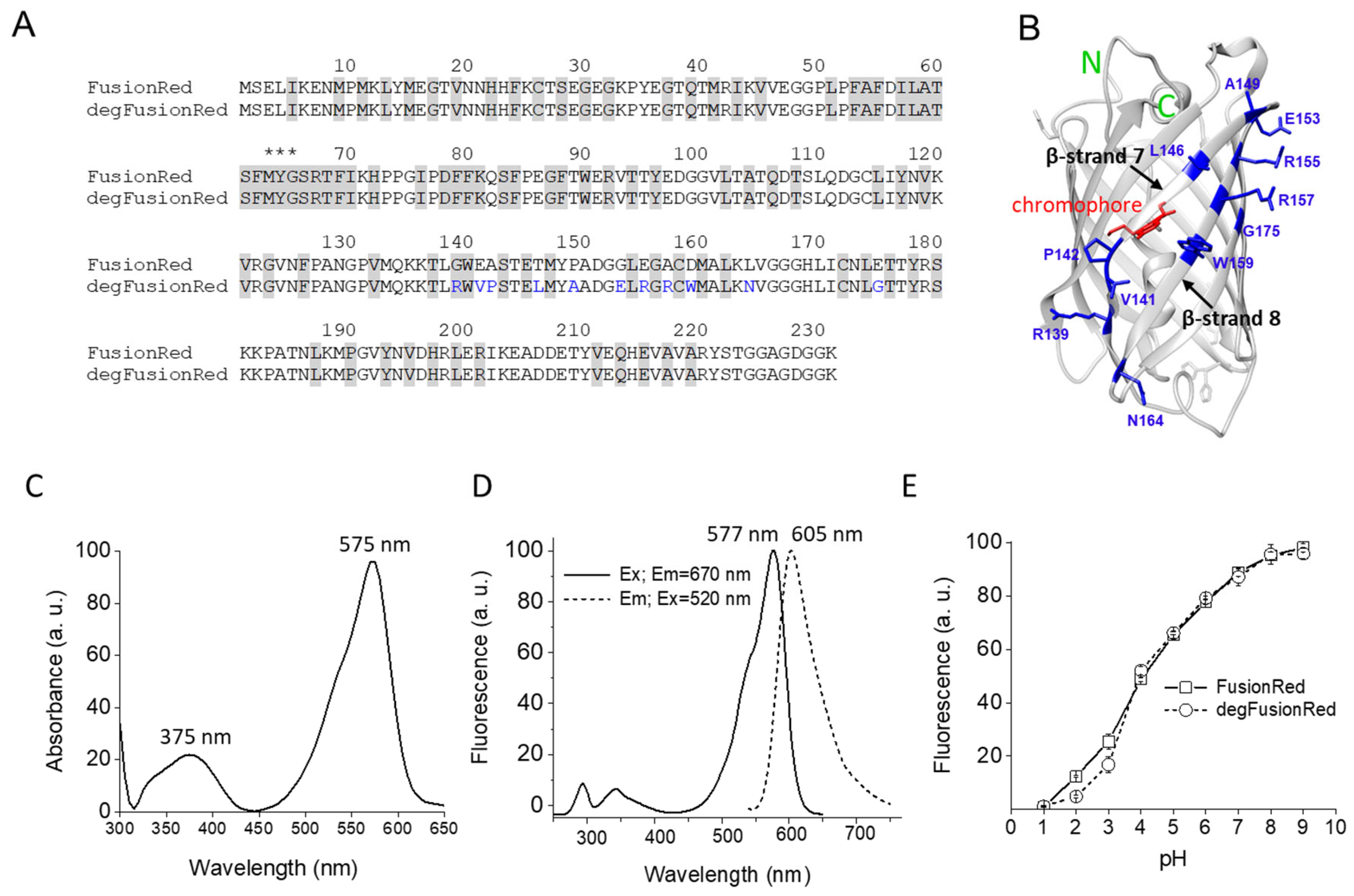
2.1. Mutagenesis and Screening of FusionRed Mutants
2.2. Optimization of Transcription with degFusionRed
2.3. Biochemical and Spectral Characterization of degFusionRed
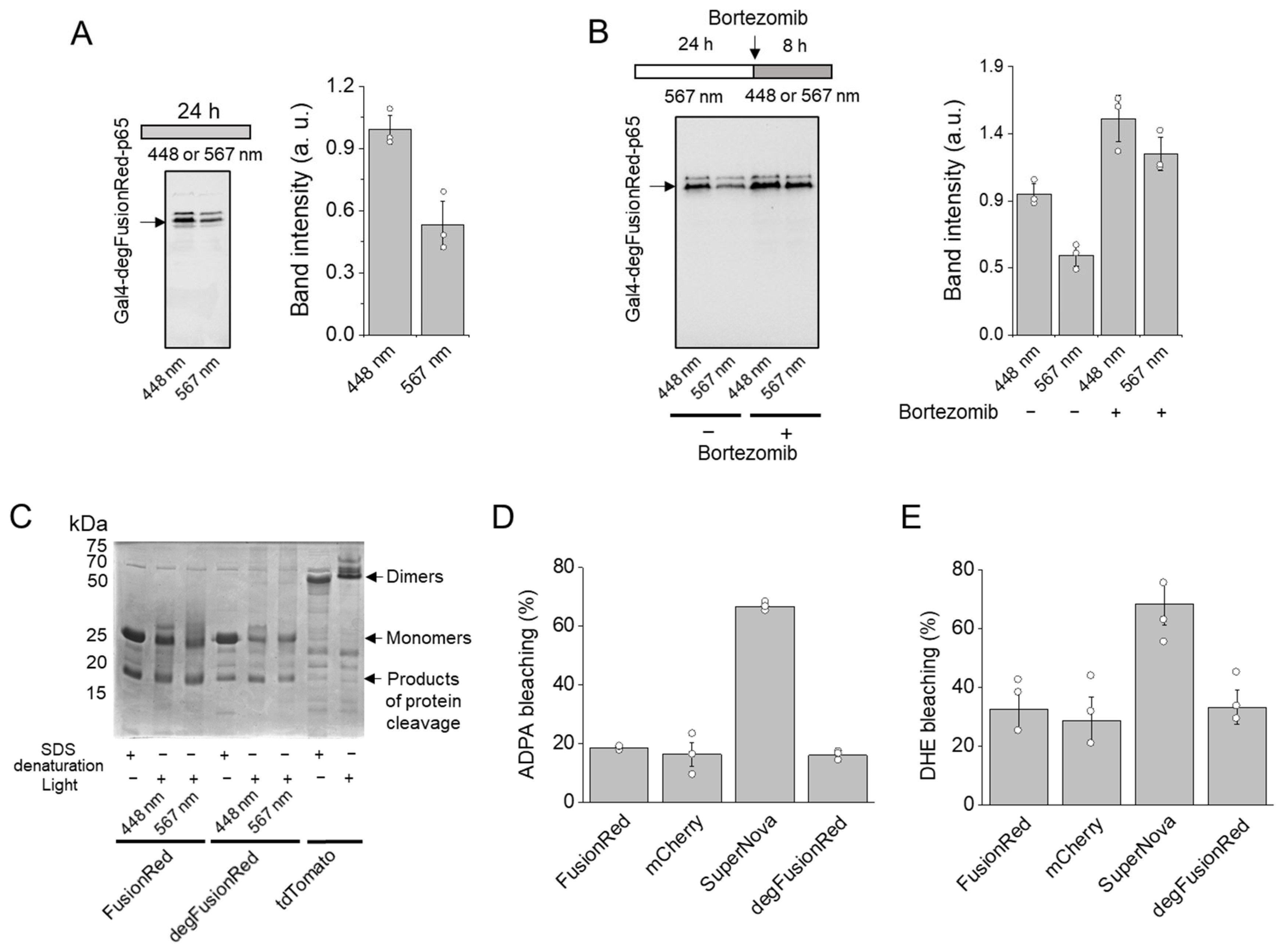
2.4. Mode of Action of degFusionRed
2.5. Analysis of degFusionRed Spectral Compatibility
2.6. Nanobody-Coupled degFusionRed for Protein Control
3. Discussion
4. Materials and Methods
4.1. Random Mutagenesis
4.2. Mammalian Cell Culture and Transfection
4.3. Screening of Mutants in Mammalian Cells
4.4. Protein Expression and Purification
4.5. Absorbance and Fluorescence Spectroscopy
4.6. Western Blot Analysis
4.7. Semi-Native PAGE Gel
4.8. Photobleaching Assay
4.9. Spectral Multiplexing with RpBphP1-QPAS1
4.10. Light Regulation of Gal4-ALFA-Tag-VP16 via Nanobody-degFusionRed
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Müller, K.; Engesser, R.; Schulz, S.; Steinberg, T.; Tomakidi, P.; Weber, C.C.; Ulm, R.; Timmer, J.; Zurbriggen, M.D.; Weber, W. Multi-chromatic control of mammalian gene expression and signaling. Nucleic Acids Res. 2013, 41, e124. [Google Scholar] [CrossRef]
- Redchuk, T.A.; Omelina, E.S.; Chernov, K.G.; Verkhusha, V.V. Near-infrared optogenetic pair for protein regulation and spectral multiplexing. Nat. Chem. Biol. 2017, 13, 633–639. [Google Scholar] [CrossRef] [PubMed]
- Redchuk, T.A.; Karasev, M.M.; Verkhusha, P.V.; Donnelly, S.K.; Hülsemann, M.; Virtanen, J.; Moore, H.M.; Vartiainen, M.K.; Hodgson, L.; Verkhusha, V.V. Optogenetic regulation of endogenous proteins. Nat. Commun. 2020, 11, 605. [Google Scholar] [CrossRef] [PubMed]
- Chernov, K.G.; Redchuk, T.A.; Omelina, E.S.; Verkhusha, V.V. Near-Infrared Fluorescent Proteins, Biosensors, and Optogenetic Tools Engineered from Phytochromes. Chem. Rev. 2017, 117, 6423–6446. [Google Scholar] [CrossRef]
- Zhang, K.; Cui, B. Optogenetic control of intracellular signaling pathways. Trends Biotechnol. 2015, 33, 92–100. [Google Scholar] [CrossRef]
- Zhang, W.; Lohman, A.W.; Zhuravlova, Y.; Lu, X.; Wiens, M.D.; Hoi, H.; Yaganoglu, S.; Mohr, M.A.; Kitova, E.N.; Klassen, J.S.; et al. Optogenetic control with a photocleavable protein, PhoCl. Nat. Methods 2017, 14, 391–394. [Google Scholar] [CrossRef] [PubMed]
- Losi, A.; Gardner, K.H.; Möglich, A. Blue-Light Receptors for Optogenetics. Chem. Rev. 2018, 118, 10659–10709. [Google Scholar] [CrossRef]
- Polstein, L.R.; Gersbach, C.A. Light-inducible spatiotemporal control of gene activation by customizable zinc finger transcription factors. J. Am. Chem. Soc. 2012, 134, 16480–16483. [Google Scholar] [CrossRef]
- Salinas, F.; Rojas, V.; Delgado, V.; López, J.; Agosin, E.; Larrondo, L.F. Fungal Light-Oxygen-Voltage Domains for Optogenetic Control of Gene Expression and Flocculation in Yeast. mBio 2018, 9, e00626-18. [Google Scholar] [CrossRef]
- Benedetti, L.; Marvin, J.S.; Falahati, H.; Guillén-Samander, A.; Looger, L.L.; De Camilli, P. Optimized Vivid-derived Magnets photodimerizers for subcellular optogenetics in mammalian cells. eLife 2020, 9, e63230. [Google Scholar] [CrossRef]
- Strickland, D.; Lin, Y.; Wagner, E.; Hope, C.M.; Zayner, J.; Antoniou, C.; Sosnick, T.R.; Weiss, E.L.; Glotzer, M. TULIPs: Tunable, light-controlled interacting protein tags for cell biology. Nat. Methods 2012, 9, 379–384. [Google Scholar] [CrossRef]
- Bugaj, L.J.; Choksi, A.T.; Mesuda, C.K.; Kane, R.S.; Schaffer, D.V. Optogenetic protein clustering and signaling activation in mammalian cells. Nat. Methods 2013, 10, 249–252. [Google Scholar] [CrossRef]
- Chen, Z.; Kang, X.W.; Zhou, Y.; Zhou, Z.; Tang, S.; Zou, S.; Wang, K.; Huang, J.; Ding, B.; Zhong, D. Dissecting the Ultrafast Stepwise Bidirectional Proton Relay in a Blue-Light Photoreceptor. J. Am. Chem. Soc. 2023, 145, 3394–3400. [Google Scholar] [CrossRef]
- Ando, R.; Mizuno, H.; Miyawaki, A. Regulated fast nucleocytoplasmic shuttling observed by reversible protein highlighting. Science 2004, 306, 1370–1373. [Google Scholar] [CrossRef]
- Zhou, X.X.; Fan, L.L.Z.; Li, P.P.; Shen, K.; Lin, M.Z. Optical control of cell signaling by single-chain photoswitchable kinases. Science 2017, 355, 836–841. [Google Scholar] [CrossRef]
- Schneider, N.; Chatelle, C.V.; Ochoa-Fernandez, R.; Zurbriggen, M.D.; Weber, W. Green Light-Controlled Gene Switch for Mammalian and Plant Cells. Methods Mol. Biol. 2021, 2312, 89–107. [Google Scholar] [PubMed]
- Jang, J.; McDonald, S.; Uppalapati, M.; Woolley, G.A. Green, orange, red, and far-red optogenetic tools derived from cyanobacteriochromes. bioRxiv 2019. [Google Scholar] [CrossRef]
- Song, P.; Yang, Z.; Guo, C.; Han, R.; Wang, H.; Dong, J.; Kang, D.; Guo, Y.; Yang, S.; Li, J. 14-3-3 proteins regulate photomorphogenesis by facilitating light-induced degradation of PIF3. New Phytol. 2023, 237, 140–159. [Google Scholar] [CrossRef] [PubMed]
- Ochoa-Fernandez, R.; Abel, N.B.; Wieland, F.G.; Schlegel, J.; Koch, L.A.; Miller, J.B.; Engesser, R.; Giuriani, G.; Brandl, S.M.; Timmer, J.; et al. Optogenetic control of gene expression in plants in the presence of ambient white light. Nat. Methods 2020, 17, 717–725. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Wu, X.; Guan, N.; Shao, J.; Li, H.; Chen, Y.; Ping, Y.; Li, D.; Ye, H. Engineering a far-red light-activated split-Cas9 system for remote-controlled genome editing of internal organs and tumors. Sci. Adv. 2020, 6, eabb1777. [Google Scholar] [CrossRef]
- Kuwasaki, Y.; Suzuki, K.; Yu, G.; Yamamoto, S.; Otabe, T.; Kakihara, Y.; Nishiwaki, M.; Miyake, K.; Fushimi, K.; Bekdash, R.; et al. A red light–responsive photoswitch for deep tissue optogenetics. Nat. Biotechnol. 2022, 40, 1672–1679. [Google Scholar] [CrossRef] [PubMed]
- Kaberniuk, A.A.; Baloban, M.; Monakhov, M.V.; Shcherbakova, D.M.; Verkhusha, V.V. Single-component near-infrared optogenetic systems for gene transcription regulation. Nat. Commun. 2021, 12, 3859. [Google Scholar] [CrossRef] [PubMed]
- Kaberniuk, A.A.; Shemetov, A.A.; Verkhusha, V.V. A bacterial phytochrome-based optogenetic system controllable with near-infrared light. Nat. Methods 2016, 13, 591–597. [Google Scholar] [CrossRef]
- Kciuk, M.; Marciniak, B.; Mojzych, M.; Kontek, R. Focus on UV-Induced DNA Damage and Repair-Disease Relevance and Protective Strategies. Int. J. Mol. Sci. 2020, 21, 7264. [Google Scholar] [CrossRef] [PubMed]
- Bauer, R.; Hoenes, K.; Meurle, T.; Hessling, M.; Spellerberg, B. The effects of violet and blue light irradiation on ESKAPE pathogens and human cells in presence of cell culture media. Sci. Rep. 2021, 11, 24473. [Google Scholar] [CrossRef]
- Andresen, M.; Stiel, A.C.; Trowitzsch, S.; Weber, G.; Eggeling, C.; Wahl, M.C.; Hell, S.W.; Jakobs, S. Structural basis for reversible photoswitching in Dronpa. Proc. Natl. Acad. Sci. USA 2007, 104, 13005–13009. [Google Scholar] [CrossRef]
- Mizuno, H.; Mal, T.K.; Walchli, M.; Kikuchi, A.; Fukano, T.; Ando, R.; Jeyakanthan, J.; Taka, J.; Shiro, Y.; Ikura, M.; et al. Light-dependent regulation of structural flexibility in a photochromic fluorescent protein. Proc. Natl. Acad. Sci. USA 2008, 105, 9227–9232. [Google Scholar] [CrossRef] [PubMed]
- Hermann, A.; Liewald, J.F.; Gottschalk, A. A photosensitive degron enables acute light-induced protein degradation in the nervous system. Curr. Biol. 2015, 25, R749–R750. [Google Scholar] [CrossRef] [PubMed]
- Takemoto, K.; Matsuda, T.; Sakai, N.; Fu, D.; Noda, M.; Uchiyama, S.; Kotera, I.; Arai, Y.; Horiuchi, M.; Fukui, K.; et al. SuperNova, a monomeric photosensitizing fluorescent protein for chromophore-assisted light inactivation. Sci. Rep. 2013, 3, 2629. [Google Scholar] [CrossRef]
- Bulina, M.E.; Chudakov, D.M.; Britanova, O.V.; Yanushevich, Y.G.; Staroverov, D.B.; Chepurnykh, T.V.; Merzlyak, E.M.; Shkrob, M.A.; Lukyanov, S.; Lukyanov, K.A. A genetically encoded photosensitizer. Nat. Biotechnol. 2006, 24, 95–99. [Google Scholar] [CrossRef]
- Torra, J.; Lafaye, C.; Signor, L.; Aumonier, S.; Flors, C.; Shu, X.; Nonell, S.; Gotthard, G.; Royant, A. Tailing miniSOG: Structural bases of the complex photophysics of a flavin-binding singlet oxygen photosensitizing protein. Sci. Rep. 2019, 9, 2428. [Google Scholar] [CrossRef] [PubMed]
- Shemiakina, I.I.; Ermakova, G.V.; Cranfill, P.J.; Baird, M.A.; Evans, R.A.; Souslova, E.A.; Staroverov, D.B.; Gorokhovatsky, A.Y.; Putintseva, E.V.; Gorodnicheva, T.V.; et al. A monomeric red fluorescent protein with low cytotoxicity. Nat. Commun. 2012, 3, 1204. [Google Scholar] [CrossRef] [PubMed]
- Shaner, N.C.; Campbell, R.E.; Steinbach, P.A.; Giepmans, B.N.; Palmer, A.E.; Tsien, R.Y. Improved monomeric red, orange and yellow fluorescent proteins derived from Discosoma sp. red fluorescent protein. Nat. Biotechnol. 2004, 22, 1567–1572. [Google Scholar] [CrossRef] [PubMed]
- Onukwufor, J.O.; Trewin, A.J.; Baran, T.M.; Almast, A.; Foster, T.H.; Wojtovich, A.P. Quantification of reactive oxygen species production by the red fluorescent proteins KillerRed, SuperNova and mCherry. Free Radic. Biol. Med. 2020, 147, 1–7. [Google Scholar] [CrossRef]
- Götzke, H.; Kilisch, M.; Martínez-Carranza, M.; Sograte-Idrissi, S.; Rajavel, A.; Schlichthaerle, T.; Engels, N.; Jungmann, R.; Stenmark, P.; Opazo, F.; et al. The ALFA-tag is a highly versatile tool for nanobody-based bioscience applications. Nat. Commun. 2019, 10, 4403. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Zhao, Y.; Guo, C.; Gan, Y.; Huang, H. The role of proline substitutions within flexible regions on thermostability of luciferase. Biochim. Biophys. Acta 2015, 1854, 65–72. [Google Scholar] [CrossRef]
- Muslinkina, L.; Pletnev, V.Z.; Pletneva, N.V.; Ruchkin, D.A.; Kolesov, D.V.; Bogdanov, A.M.; Kost, L.A.; Rakitina, T.V.; Agapova, Y.K.; Shemyakina, I.I.; et al. Two independent routes of post-translational chemistry in fluorescent protein FusionRed. Int. J. Biol. Macromol. 2020, 155, 551–559. [Google Scholar] [CrossRef]
- Marschall, A.L.; Dübel, S. Antibodies inside of a cell can change its outside: Can intrabodies provide a new therapeutic paradigm? Comput. Struct. Biotechnol. J. 2016, 14, 304–308. [Google Scholar] [CrossRef]
- Dong, G.; Ding, Y.; He, S.; Sheng, C. Molecular Glues for Targeted Protein Degradation: From Serendipity to Rational Discovery. J. Med. Chem. 2021, 64, 10606–10620. [Google Scholar] [CrossRef]
- Carmony, K.C.; Kim, K.B. PROTAC-induced proteolytic targeting. Methods Mol. Biol. 2012, 832, 627–638. [Google Scholar]
- Liu, J.; Chen, H.; Ma, L.; He, Z.; Wang, D.; Liu, Y.; Lin, Q.; Zhang, T.; Gray, N.; Kaniskan, H.; et al. Light-induced control of protein destruction by opto-PROTAC. Sci. Adv. 2020, 6, eaay5154. [Google Scholar] [CrossRef]
- Yesbolatova, A.; Saito, Y.; Kitamoto, N.; Makino-Itou, H.; Ajima, R.; Nakano, R.; Nakaoka, H.; Fukui, K.; Gamo, K.; Tominari, Y.; et al. The auxin-inducible degron 2 technology provides sharp degradation control in yeast, mammalian cells, and mice. Nat. Commun. 2020, 11, 5701. [Google Scholar] [CrossRef]
- Macdonald, L.; Taylor, G.C.; Brisbane, J.M.; Christodoulou, E.; Scott, L.; von Kriegsheim, A.; Rossant, J.; Gu, B.; Wood, A.J. Rapid and specific degradation of endogenous proteins in mouse models using auxin-inducible degrons. eLife 2022, 11, e77987. [Google Scholar] [CrossRef]
- Zeng, J.; Santos, A.F.; Mukadam, A.S.; Osswald, M.; Jacques, D.A.; Dickson, C.F.; McLaughlin, S.H.; Johnson, C.M.; Kiss, L.; Luptak, J.; et al. Target-induced clustering activates Trim-Away of pathogens and proteins. Nat. Struct. Mol. Biol. 2021, 28, 278–289. [Google Scholar] [CrossRef]
- Taxis, C.; Stier, G.; Spadaccini, R.; Knop, M. Efficient protein depletion by genetically controlled deprotection of a dormant N-degron. Mol. Syst. Biol. 2009, 5, 267. [Google Scholar] [CrossRef] [PubMed]
- Izert, M.A.; Klimecka, M.M.; Górna, M.W. Applications of Bacterial Degrons and Degraders—Toward Targeted Protein Degradation in Bacteria. Front. Mol. Biosci. 2021, 8, 669762. [Google Scholar] [CrossRef]
- Bonger, K.M.; Rakhit, R.; Payumo, A.Y.; Chen, J.K.; Wandless, T.J. General method for regulating protein stability with light. ACS Chem. Biol. 2014, 9, 111–115. [Google Scholar] [CrossRef]
- Sun, W.; Zhang, W.; Zhang, C.; Mao, M.; Zhao, Y.; Chen, X.; Yang, Y. Light-induced protein degradation in human-derived cells. Biochem. Biophys. Res. Commun. 2017, 487, 241–246. [Google Scholar] [CrossRef] [PubMed]
- Mondal, P.; Krishnamurthy, V.V.; Sharum, S.R.; Haack, N.; Zhou, H.; Cheng, J.; Yang, J.; Zhang, K. Repurposing Protein Degradation for Optogenetic Modulation of Protein Activities. ACS Synth. Biol. 2019, 8, 2585–2592. [Google Scholar] [CrossRef]
- Pathak, G.P.; Spiltoir, J.I.; Höglund, C.; Polstein, L.R.; Heine-Koskinen, S.; Gersbach, C.A.; Rossi, J.; Tucker, C.L. Bidirectional approaches for optogenetic regulation of gene expression in mammalian cells using Arabidopsis cryptochrome 2. Nucleic Acids Res. 2017, 45, e167. [Google Scholar] [CrossRef] [PubMed]
- Longo, P.A.; Kavran, J.M.; Kim, M.S.; Leahy, D.J. Transient mammalian cell transfection with polyethylenimine (PEI). Methods Enzymol. 2013, 529, 227–240. [Google Scholar]
- Piatkevich, K.D.; Subach, F.V.; Verkhusha, V.V. Far-red light photoactivatable near-infrared fluorescent proteins engineered from a bacterial phytochrome. Nat. Commun. 2013, 4, 2153. [Google Scholar] [CrossRef] [PubMed]
- Mruk, D.D.; Cheng, C.Y. Enhanced chemiluminescence (ECL) for routine immunoblotting: An inexpensive alternative to commercially available kits. Spermatogenesis 2011, 1, 121–122. [Google Scholar] [CrossRef] [PubMed]
- Oliinyk, O.S.; Baloban, M.; Clark, C.L.; Carey, E.; Pletnev, S.; Nimmerjahn, A.; Verkhusha, V.V. Single-domain near-infrared protein provides a scaffold for antigen-dependent fluorescent nanobodies. Nat. Methods 2022, 19, 740–750. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chernov, K.G.; Manoilov, K.Y.; Oliinyk, O.S.; Shcherbakova, D.M.; Verkhusha, V.V. Photodegradable by Yellow-Orange Light degFusionRed Optogenetic Module with Autocatalytically Formed Chromophore. Int. J. Mol. Sci. 2023, 24, 6526. https://doi.org/10.3390/ijms24076526
Chernov KG, Manoilov KY, Oliinyk OS, Shcherbakova DM, Verkhusha VV. Photodegradable by Yellow-Orange Light degFusionRed Optogenetic Module with Autocatalytically Formed Chromophore. International Journal of Molecular Sciences. 2023; 24(7):6526. https://doi.org/10.3390/ijms24076526
Chicago/Turabian StyleChernov, Konstantin G., Kyrylo Yu. Manoilov, Olena S. Oliinyk, Daria M. Shcherbakova, and Vladislav V. Verkhusha. 2023. "Photodegradable by Yellow-Orange Light degFusionRed Optogenetic Module with Autocatalytically Formed Chromophore" International Journal of Molecular Sciences 24, no. 7: 6526. https://doi.org/10.3390/ijms24076526