

Zebrafish: A Relevant Genetic Model for Human Primary Immunodeficiency (PID) Disorders?


1
School of Medicine, Deakin University, Geelong, VIC 3216, Australia
2
Institute for Mental and Physical Health and Clinical Translation (IMPACT), Deakin University, Geelong, VIC 3216, Australia
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2023, 24(7), 6468; https://doi.org/10.3390/ijms24076468
Submission received: 6 March 2023
/
Revised: 28 March 2023
/
Accepted: 28 March 2023
/
Published: 30 March 2023
(This article belongs to the Topic Animal Models of Human Disease)
Abstract
:Primary immunodeficiency (PID) disorders, also commonly referred to as inborn errors of immunity, are a heterogenous group of human genetic diseases characterized by defects in immune cell development and/or function. Since these disorders are generally uncommon and occur on a variable background profile of potential genetic and environmental modifiers, animal models are critical to provide mechanistic insights as well as to create platforms to underpin therapeutic development. This review aims to review the relevance of zebrafish as an alternative genetic model for PIDs. It provides an overview of the conservation of the zebrafish immune system and details specific examples of zebrafish models for a multitude of specific human PIDs across a range of distinct categories, including severe combined immunodeficiency (SCID), combined immunodeficiency (CID), multi-system immunodeficiency, autoinflammatory disorders, neutropenia and defects in leucocyte mobility and respiratory burst. It also describes some of the diverse applications of these models, particularly in the fields of microbiology, immunology, regenerative biology and oncology.
1. Introduction
The human immune system is composed of a complex network of specialist cells and organs that represents the body’s major protective mechanism against infections and cancer. Innate immunity, of which neutrophils and macrophages are the key components, represents the first line of defense against these pathogenic threats [1,2]. Adaptive immunity, which is dominated by cells of the lymphoid compartment, provides extended defense, including the immune memory that facilitates rapid and effective responses against subsequent encounters with infectious agents and tumor cells [3]. The development and function of the various immune cells throughout the life-course is precisely controlled by a large set of critical genes.
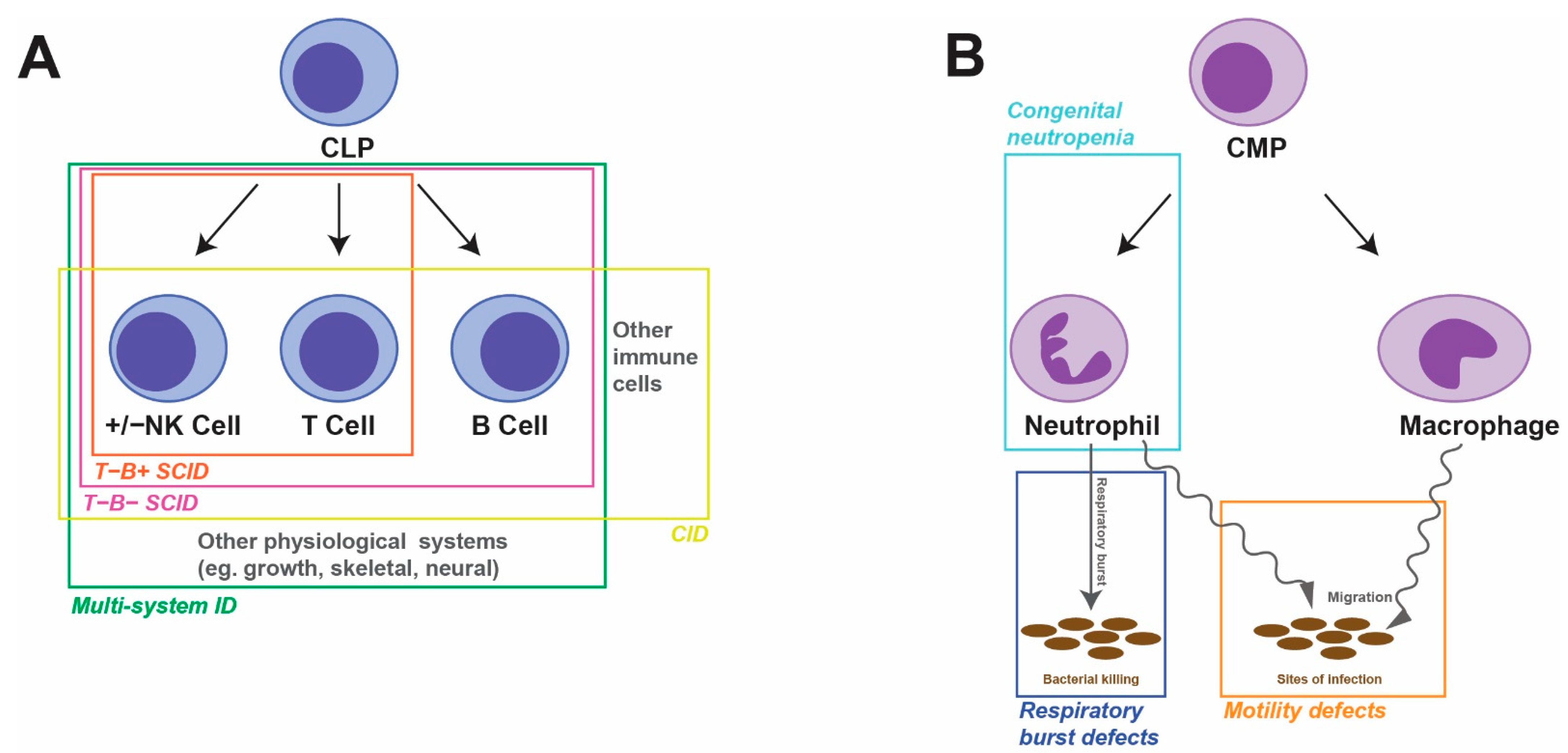
Human primary immunodeficiencies (PIDs), also known as inborn errors of immunity (IEIs), represent a diverse group of nearly 500 individual disorders [4]. Each is characterized by specific combinations of absent immune cell(s) and/or defective immune function(s) and may include syndromic features outside of the immune system [5]. The major forms typically impact either the lymphoid or myeloid lineages that are the focus of this review (Figure 1). The understanding of how particular genetic defects mediate the specific immunological phenotypes remains poor in many instances. Such information is critical to the development of new and tailored treatment options to overcome the high levels of morbidity and mortality experienced and to provide patient-specific ‘precision’ medical care.
Zebrafish represent an established genetic model for studying development and disease, including the cells and functions relevant to immunity [6]. The zebrafish immune system has both innate and adaptive arms, comprising immune cells that possess comparable functions to their human counterparts, concomitant with a high level of conservation of the specific genes that control their development and action [7]. Zebrafish are also easily manipulable genetically and physically, notably including the introduction of gene mutations using genome editing, as well as transgenesis and other sophisticated approaches [8]. They are also extremely accessible for imaging and other analyses [9] and exhibit favorable husbandry. Collectively, this has resulted in wide application of this organism to study blood and immune cell disorders [10,11], with over 100 publications describing immunodeficient zebrafish, including the gene disruptions associated with human PIDs.
2. The Zebrafish Immune System in Comparison to That of Humans
The cells comprising the zebrafish immune system are generated and maintained via distinct waves of hematopoiesis occurring at multiple anatomical sites, as is the case in humans [12]. Primitive macrophages are produced as part of the so-called ‘primitive’ wave of hematopoiesis, some of which populate the brain to form the microglial population, with a transient ‘intermediate’ wave of hematopoiesis in the posterior blood island, generating neutrophils [13]. Multipotent hematopoietic stem cells (HSCs) initiate the ‘definitive’ wave of hematopoiesis and sustain the process throughout the lifespan [14], with these HSCs initially homing to the caudal hematopoietic tissue (CHT), equivalent to the mammalian fetal liver, to produce multiple cell lineages, with lymphocyte precursors generated migrating to the thymus to commence the production of T cells [12,15]. HSCs ultimately populate the kidney marrow, equivalent to the mammalian bone marrow, allowing ongoing production of neutrophils, macrophages and other leukocytes, and later B cells, with full immunity including natural killer (NK) cell populations in place by 4–6 weeks post fertilization [12,15,16].
Like their human counterparts, zebrafish neutrophils expressing the enzymes myeloperoxidase [17] and gelatinase [18] are the initial innate immune cell responders to sites of infection or injury [19], with zebrafish macrophages expressing perforin 2 and l-plastin [20] arriving subsequently to phagocytose pathogens and cell debris [19]. Macrophages and dendritic cells present antigens derived from infecting microorganisms or cancer cells to lymphocytes, initiating an adaptive immune response [21]. Eosinophils and mast cells are also found in zebrafish, with eosinophils able to degranulate in response to antigenic exposure [22,23]. Various zebrafish T cell populations, including those expressing CD4 and CD8, induce appropriate cellular responses mediated via four T cell receptor (TCR) chains (α, β, γ, δ) [15,24], while B cells elicit humoral responses leading to the production of immunoglobin (Ig) molecules equivalent to those seen in mammals (IgM and IgD) as well as fish-specific forms (IgT, IgZ) [25]. Zebrafish also possess two distinct classes of NK cell populations, NK and NK-like, which express archetypal NK-lysin enzymes [26].
The control of immune cell development shows strong correlation between zebrafish and humans. This includes lineage-specific transcription factors important for the production of specific lymphoid and myeloid immune cell lineages. For example, the zebrafish IKZF1 gene orthologue has been shown to play a key role in early lymphoid development [27], while SPI1-related genes were demonstrated to be important in early myeloid cell development [13]. This correlation also extends to cytokine and chemokine cell signaling components that stimulate differentiation, proliferation, survival, migration and activation of these cells [28,29]. As a consequence, zebrafish have been widely employed as a relevant genetic model across multiple categories of PIDs (Table 1).
3. Zebrafish Models of Lymphoid Disorders
3.1. Severe Combined Immunodeficiency
Severe combined immunodeficiency (SCID) is a disorder that is characterized by a block in T cell differentiation (T−) along with variable impacts on the development of B cells (B+/−) and NK cells (NK+/−), thereby rendering patients highly susceptible to a range of life-threatening infections and cancers [63,64].
3.1.1. T−B− SCID
The first zebrafish PID model was generated by targeting the zebrafish orthologue of the human RAG1 gene. Zebrafish homozygous for a loss-of-function (LOF) rag1 mutation were viable, but exhibited decreased lymphocytes in both peripheral blood and kidney, with increased circulating neutrophils and increased myelomonocytes in the kidney. The mutants were defective in V(D)J recombination, with rearrangement of TCRβ and IgM significantly abrogated, indicating a paucity of mature T and B cells, although NK markers were found to be comparable to wild-type fish [30,31], and so reflective of the T−B−NK+ SCID observed in patients with RAG1 deficiency [65]. In contrast, adult zebrafish homozygous for a rag2 mutation displayed a significant reduction in circulating lymphocytes, reflecting a loss of T cells, but with variable decreases in B cell numbers, possibly due to the hypomorphic nature of the mutation [16,32]. Zebrafish LOF prkdc mutants also showed a significant reduction in both T and B cell lineages, with NK cells but not NK-like cells also affected, resulting in high susceptibility to infection and reduced anti-tumor immunity in these fish [16,26,33]. In contrast, LOF mutations and morpholino-mediated knockdown (KD) of the zebrafish ak2 gene both resulted in reduction of embryonic T cells and neutrophils [34]. These impacts are consistent with the reticular dysgenesis (aleukocytosis) observed in humans deficient in AK2, a severe form of T−B−NK− SCID also impacting neutrophils [66].
3.1.2. T−B+ SCID
Human T−B+ SCID is caused by a variety of genetic lesions, with a number of such mutations impacting cytokine signaling via interleukin-2 (IL-2) family members [67], some of which have been successfully modelled in zebrafish. For instance, LOF mutations in the zebrafish il2rga gene, which encodes the IL-2 receptor γ (IL-2Rγ) common chain, resulted in severely reduced numbers of embryonic T cells, impaired juvenile TCR rearrangement and significantly reduced adult T cells and NK cells; however, B cells and myeloid cells were not affected [26,35,68]. LOF mutations in jak3, encoding the IL-2Rγ-associated Janus kinase 3 (Jak3), shared similar phenotypes, with severely impaired T cell development and a reduction in the number of NK cells [16,36,37], but only minor impacts on B cell maturation and neutrophil development [37]. Both of these T−B+NK− SCID models exhibited perturbed immunity, with the il2rga mutants displaying dysregulated intestinal microbiota and defective tumor immunity [35] and the jak3 mutants being susceptible to the development of lymphoid malignancy [37]. Finally, LOF mutations in il7r, encoding a signaling chain utilizing IL-2Rγ, also displayed severely impaired embryonic T cell development [36].
3.2. Combined Immunodeficiency
Combined immunodeficiency (CID) disorders are generally less profound than SCID. For example, human ZAP70 (Zeta-chain (TCR)-associated protein kinase, 70 kDa) deficiency represents a rare form of CID, characterized by a loss of peripheral CD8+ T cells and non-functional CD4+ T cells [69]. Consistent with the human disorder, zebrafish zap70 LOF mutants also displayed severe defects within the T cell lineage, with a reduction in mature T cells in kidney marrow and impaired thymic T cell development observed [38]. Similarly, IKZF1 deficiency impacts memory T and B cell populations in humans [70], with LOF ikzf1 mutant zebrafish exhibiting a severe reduction in larval T cells, although juvenile and adult T cell development was less impacted [39].
3.3. Multi-System Immunodeficiency
Other human immunodeficiencies have concurrent associated or syndromic features and so are referred to as ‘multi-system’ disorders [4]. For example, growth hormone insensitivity syndrome with immune dysregulation 1 (GHISID1) is characterized by immune perturbation and postnatal growth failure due to LOF mutations in human STAT5B [71]. LOF mutations in the equivalent zebrafish gene, stat5.1, caused impaired lymphopoiesis with reduced T cells throughout the lifespan, along with broader disruption of the lymphoid compartment into adulthood, including evidence of increased T cell activation [40]. The mutants also showed reduced growth and increased adiposity with concomitant dysregulation of growth and lipid metabolism genes [40,72], faithfully recapitulating the human disease. Similarly, human hypomorphic EXTL3 mutations cause an immuno-osseus syndrome that involves both immune deficiency and skeletal dysplasia [1], with equivalent zebrafish LOF extl3 mutants showing decreased thymic T cells [41] and defective pectoral fin development [73]. In addition, knockdown of the zebrafish arpc1b gene resulted in reduced embryonic T cells and thrombocytes [42], recapitulating human ARPC1B deficiency [74]. Ataxia-telangiectasia (A-T) syndrome is a human disorder characterized by neurodegeneration, immune dysregulation, cancer susceptibility and premature aging and results from defects in the ATM gene [43]. Zebrafish LOF atm mutants exhibited similar phenotypes, with multi-lineage immunodeficiency along with motor disturbances and cancer predisposition [75]. Finally, human BCL11B gain-of-function (GOF) mutations have been identified in an autosomal-dominant syndrome characterized by reduced T cells, congenital craniofacial abnormalities and neurocognitive defects [76], with overexpression of a patient-derived BCL11B mutant in bcl11b KD zebrafish embryos able to model the T cell defects and dominant nature of the human disease [44]. Other mutations impact human thymus development and thereby cause immunodeficiency indirectly by abrogating T cell production at this site [77]. This has been recapitulated in zebrafish foxn1 LOF mutants [78], both chd7 LOF mutants and KD embryos [46], and zebrafish tbx1 LOF mutants [47].
3.4. Auto-Inflammatory Disorders
A variety of human PIDs result from enhanced inflammation, including several auto-inflammatory disorders and immunodeficiencies [4]. For example, NCKAP1L deficiency has been shown to result in immunodeficiency, lymphoproliferation and excessive inflammation, with zebrafish nckap1l KD shown to cause defective neutrophil migration [48].
4. Zebrafish Models of Myeloid Disorders
4.1. Congenital Neutropenia
A number of human PIDs are characterized by insufficient numbers of neutrophils, with several of these successfully modelled in zebrafish. For example, LOF mutations in human CSF3R, encoding the receptor for the cytokine granulocyte colony-stimulating factor (G-CSF), have been identified in a cohort of neutropenia patients unresponsive to G-CSF treatment [79]. Zebrafish csf3r LOF mutants also possess significantly decreased numbers of neutrophils from embryonic to adult stages [49,80]. The neutrophils were found to be functionally compromised, with mutants unable to respond to G-CSF and displaying enhanced susceptibility to bacterial infection [49]. LOF mutations in SMARCD2, encoding a chromatin remodeling factor, have also been associated with human neutropenia in combination with myelodysplasia and developmental abnormalities, with ablation of the zebrafish smarcd2 gene similarly leading to a reduction in neutrophil numbers in embryos [50]. Mutations in human Wiskott-Aldrich syndrome (WAS) protein have been implicated in the pathogenesis of X-linked neutropenia (XLN) [74]. Consistent with this, a LOF zebrafish was mutant showed defective migration of neutrophils as well as macrophages, which correlated with increased susceptibility to bacterial infection due to delayed pathogen clearance [51]. Similarly, human HAX1 mutations have been associated with autosomal-recessive severe congenital neutropenia (SCN) [81], with hax1 KD in zebrafish embryos resulting in impaired neutrophil development [52]. In addition, LOF mutations in human VPS45, encoding a regulator of endosomal membrane trafficking, have been associated with neutropenia, neutrophil dysfunction, nephromegaly and bone marrow fibrosis, with the reduced neutrophil count recapitulated in zebrafish vps45 KD embryos [53]. Finally, patients with Shwachman-Diamond syndrome present with neutropenia in combination with exocrine pancreas deficiency and skeletal muscle abnormalities associated with dominant mutations in the ribosome maturation factor SRP54 in some cases [82]. Equivalent zebrafish srp54 mutants exhibited decreased embryonic neutrophils, although this was only observed in homozygous animals, indicating a lack of dominance in zebrafish [54].
4.2. Motility Defects
Mutations in the human ITGB2 gene encoding the cell surface molecule CD18 have been associated with human leukocyte adhesion deficiency (LAD), a disorder characterized by recurrent infections and reduced neutrophil motility [83]. Consistent with this, zebrafish LOF itgb2 mutants showed decreased trafficking to sites of inflammation despite increased neutrophil numbers in circulation [55]. Similarly, mutations in the RAC2 gene, encoding a member of the small GTPase family, have been associated with immunological defects in humans, with dominant-negative mutations leading to granulocyte mobility defects [84]. Ablation of the zebrafish rac2 gene by mutation or KD resulted in reduced mobility of neutrophils and macrophages, but enhanced neutrophil mobilization, resulting in hypersensitivity to infection by Pseudomonas aeruginosa [56,57]. So-called ‘warts, hypogammaglobulinemia, infections, and myelokathexis’ (WHIM) syndrome is a PID disorder characterized by defective and decreased neutrophils and consequent recurrent infections, mediated by dominant mutations in the human CXCR4 gene that encodes a G-protein-coupled chemokine receptor [85]. Zebrafish embryos transgenically expressing the equivalent mutations in the context of the zebrafish CXCR4 equivalent, Cxcr4b, possessed reduced numbers of embryonic neutrophils with impaired neutrophil mobility and wounding recruitment, along with enhanced retention in the hematopoietic tissues [58], reminiscent of the human WHIM disorder.
4.3. Respiratory Burst Defects
The neutrophil respiratory burst is a key mechanism for mediating the killing of bacteria and other pathogens but also plays an important role in cell signaling. Human chronic granulomatous disease (CGD) is caused by mutations in a number of genes that encode components of the so-called ‘phagocyte NADPH oxidase’ (PHOX) complex, which mediates superoxide formation, including the CYBA gene encoding the p22Phox subunit [86]. Zebrafish LOF cyba mutants showed reduced neutrophil reverse migration and impaired macrophage wound attraction, leading to increased neutrophil infiltration, enhanced susceptibility to invasive fungal infection and neutrophil-mediated inflammation [59,60]. The myeloperoxidase (MPO) enzyme acts on H2O2 generated in injured tissues through the respiratory burst, with evidence that humans harboring MPO mutations show increased susceptibility to infections [87]. Zebrafish mpo LOF mutants had normal numbers of neutrophils that migrated faithfully to sites of infection [61,62], with similar survival following infection with the fungal pathogen Candida albicans; however, there was increased pathogen proliferation, neutrophil accumulation and elevated expression of inflammatory cytokines [62].
5. Other Zebrafish Immunodeficiency Mutants
Additional zebrafish immunodeficient models have been generated that resemble human PIDs. For example, the zebrafish earl gray mutant, which represents a LOF mutation in the gene encoding the p110/SART3 general splicing factor, exhibits disrupted thymic development, leading to thymic hypoplasia and a lack of T lymphocytes typical in forms of CID [88]. In addition, LOF runx1 mutations resulted in a reduction in B cells and a failure in B cell V(D)J rearrangement, a phenotype similar to that observed in human common variable immunodeficiency [89]. Furthermore, zebrafish tlx1 mutations were found to disrupt spleen development, resulting in partial impairment of mononuclear phagocytes and reduced levels of IgM [90]. Other mutants have identified novel regulators of immune cell development and/or function, with the underlying genes representing potential candidates for idiotypic human PIDs. For example, a missense mutation in the transcriptional regulator zbtb17 resulted in an early block in zebrafish intrathymic T cell development [91], while a cmyb mutant zebrafish displayed severely reduced lymphocytes, precursors and myelomonocytes in the kidney, with early lethality [92]. In addition, the zebrafish urb2 mutant was deficient in HSC within the CHT and early T cells in the thymus [93], whereas stn1 KD zebrafish displayed an arrest in T cell progenitors [94]. Zebrafish is also being used to identify immune defects as part of other complex human syndromic disorders. For example, a fatal syndrome characterized with severe autoinflammation and leukoencephalopathy due to glycogen-storage-associated mitochondriopathy was identified in eight sporadic families with C2ORF69 LOF mutations, with the immune perturbation verified in c2orf69 mutant zebrafish [95]. Zebrafish are additionally being employed to confirm the gene–phenotype correlation in emerging human PIDs, such as LOF defects in human SRP19 and SRPRA associated with SCN [96]. Finally, this model organism is being applied to the study of other key aspects of immunity, such as barrier immunity [97] and Toll-like receptors (TLRs) [98].
6. Applications of Zebrafish Immunodeficiency Models
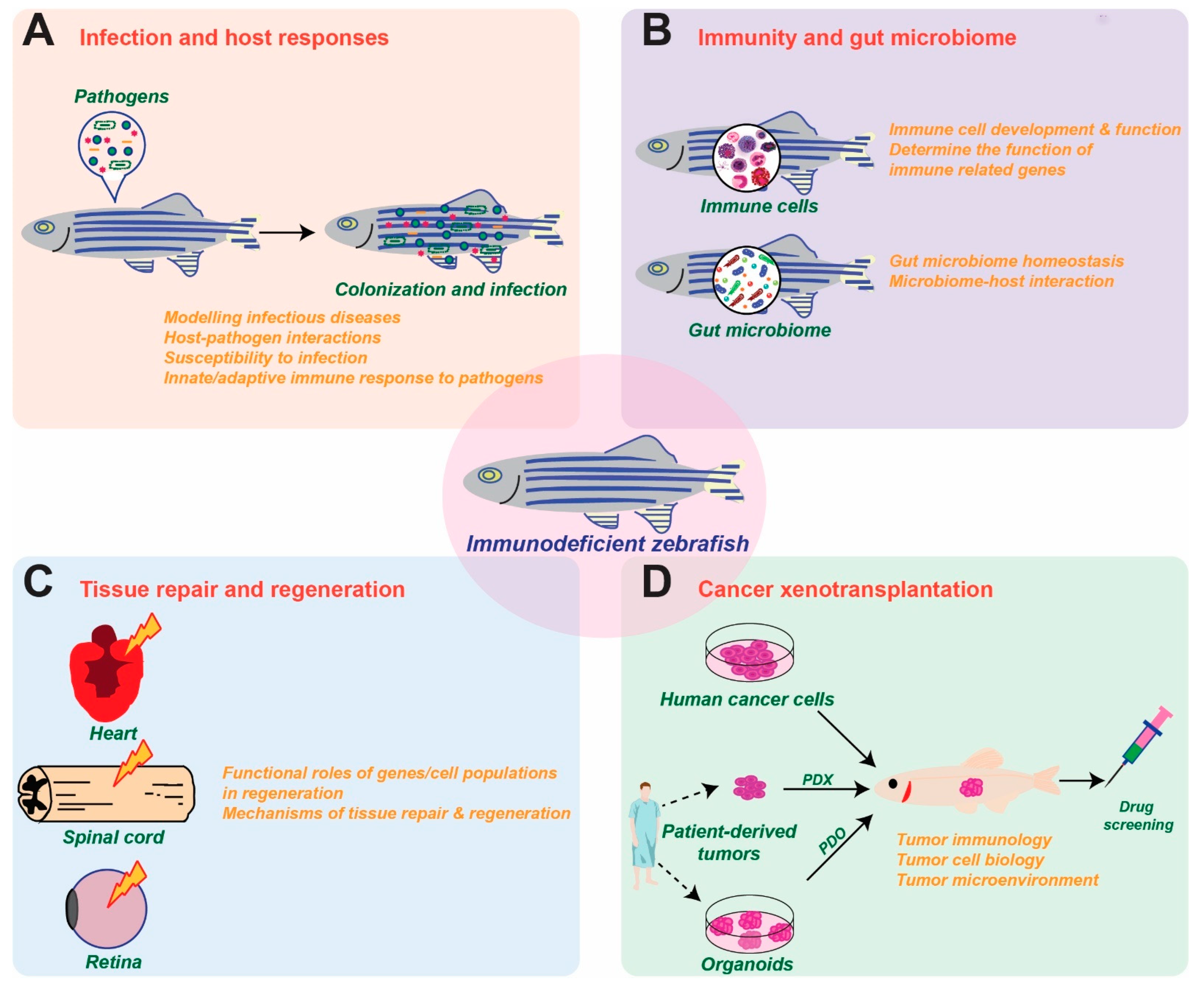
Immunodeficient zebrafish lines, including PID models, are being widely employed to offer new insights into the interplay of the immune system with pathogenic agents and normal microbiota, as well as in the fields of regenerative medicine and cancer research (Figure 2).
6.1. Infection and Host Responses
Immunodeficient zebrafish models have served as excellent in vivo platforms for studying host–pathogen interactions involving a wide variety of bacteria and other microorganisms. In many cases these studies take advantage of the optical transparency of zebrafish, often combined with fluorescent labeling of specific immune cell lineages and/or microbial agents to provide unprecedented levels of detail [99]. Utilization of various immune cell depletion strategies has allowed the role of specific lineages to be definitively determined [19]. For example, zebrafish rag1 mutants were found to be highly susceptible to Mycobacterium marinum infection, demonstrating a heavy reliance on adaptive immunity to defend against these pathogens [100]. However, reinfection with the same bacteria highlighted the ability of innate immune cells to mediate an ‘adaptive’ response, with vaccinated mutants displaying increased survival compared to naïve mutants [101]. Enhanced susceptibility to bacterial infection was demonstrated in both csf3r [49] and myd88 [102] LOF mutants, highlighting the essential functions of innate immunity in defense against bacteria. Increasingly, these studies are being directed toward human pathogens [103]. Thus, analysis of the adult T cell–deficient zebrafish lck mutant strain has revealed the significant role played by T cell–mediated immune response in immunity against the clinically relevant pathogen M. abscessus [104]. Other studies have investigated the role of specific elements of immunity in infection, such as the function of neutrophil calprotectin in immunity to Vibrio cholerae [105].
6.2. Immunity and Gut Microbiome
Immunodeficient zebrafish have also been used as important tools to study the immune system under normal and diseased conditions. This has provided new insights into immune development, but also inflammation and its resolution [106], allowing dissection of the signaling pathways that regulate recruitment and fate of phagocytes at inflammatory sites [107]. Crossing of PID models onto the optically transparent casper background has further enhanced the ability to characterize immune cells, allowing sophisticated approaches to be employed, including transcriptome analysis via RNAseq [16,26]. More recently, PID and other zebrafish models have been applied to study the interplay between immunity and the gut microbiome, typically analyzed by so-called ‘Next-gen’ sequencing of genomic DNA extracted from dissected gut tissues, complemented by the ability to manipulate the microbiome and also apply cutting-edge imaging approaches [108,109]. For example, zebrafish il2rga mutants displayed a dysregulated intestinal microbiota with significantly reduced alpha diversity and a concomitant increase in short chain fatty acid (SCFA)-producing bacteria [35], similar to human SCID patients [110]. In contrast, LOF rag1 mutants showed altered abundance of Vibrio species in the gut [111]. The contribution of innate immune cells to microbiota composition was revealed using LOF myd88 [112] and irf8 [113] mutants, both of which showed significantly altered gut microbiome composition.
6.3. Tissue Repair and Regeneration
The immune system plays a crucial role in tissue repair and regeneration. The regenerative mechanisms of a range of tissues, such as spinal cord, heart, retina, liver, pancreas, nervous system and skeletal elements, have been explored using zebrafish models, taking advantage of the ease of manipulation and imaging in this organism [114,115,116]. In particular, the use of zebrafish deficient in specific subsets of immune cells has aided in determining their functional roles in key aspects of regenerative biology. For example, the use of foxp3a mutant zebrafish has revealed an essential function for Treg cells in producing regenerative factors tailored to specific tissue repair/regeneration [117]. Zebrafish neutrophils and macrophages have also been shown to be critical in tissue repair and regeneration, which extends to microglial cells in the brain [118,119]. For example, granulin-deficient zebrafish lacking embryonic macrophages and embryonic and adult neutrophils showed impaired tissue repair and wound healing [120], whereas cftr deficiency resulted in excessive neutrophil recruitment, leading to increased tissue damage and abnormal repair [121]. In contrast, analysis of the neutrophil-defective runx1 mutant has revealed an unexpected inhibitory role for neutrophils during regeneration of the tail fin [118].
6.4. Cancer Xenotransplantation
Amongst the diverse applications of zebrafish immunodeficiency models, their use in cancer xenotransplantation studies continues to show great promise, from a handful of papers prior to 2009 to now over 200 publications. These are adding to the understanding of cancer biology and aiding the development of therapeutics [120,122]. A wide range of human cancer cell lines and tumors have been successfully grafted in zebrafish and analyzed using a variety of imaging-based strategies to investigate tumor cell proliferation, apoptosis, invasion, metastasis, angiogenesis and interactions with the host, including the tumor microenvironment, or for pre-clinical testing of potential therapeutic compounds by placing them directly in the water [123,124,125,126,127]. For example, the rag2 mutant line has been shown to be a useful model for human cancer xenotransplantation [32], while zebrafish prkdc mutants show 50–70% engraftment when injected intraperitoneally with tumor cells from human melanoma, leukemia, pancreatic cancer and bile duct cancer cell lines [33]. Comparison of zebrafish prkdc, rag2 and jak3 mutant models has revealed the prkdc mutants as the most efficient platform for tumor xenotransplantation [128]. Crossing of PID models onto the casper background has further enhanced the ability to image and characterize fluorescently labeled cancer cells [37] as well as immune cells and the interactions between the two in real time [129]. For example, a prkdc casper mutant line was used to develop a platform for drug testing of fluorescently labelled huma leukemia xenografts [130]. A prkdc il2rga casper mutant line was demonstrated to successfully engraft a wide range of human cancer cells that could be monitored in vivo at single-cell resolution [131]. Finally, a rag2 il2rga casper mutant line of zebrafish was suitable for long-term engraftment of human cancer and T cells, allowing it to be used as an anticancer screening platform to quantify responses to a range of cancer immunotherapy approaches, including those involving chimeric antigen receptor (CAR) T cell, bispecific T cell engagers (BiTE), and antigen-peptide epitope conjugate (APEC) approaches [132]. In addition to human cancer cell line engraftment, zebrafish SCID models have been successfully applied to the engraftment of primary patient-derived samples, such as patient-derived xenografts (PDXs) or patient-derived organoids (PDOs) [131,133,134]. These have been utilized for the assessment of tumor heterogeneity [135], chemosensitivity [136], radiosensitivity [137] and high-throughput therapeutic drug and immunotherapy testing [138].
6.5. Other Applications
Zebrafish immunodeficiency models have also been employed for additional applications. For example, zebrafish are being used to assess the relative impact of different human mutations, such as by transgenic expression of WAS protein mutants in a was knockout model [51]. Additionally, the zebrafish models are allowing for increased mechanistic understanding. This includes broader pathways involved in mediating the observed immunodeficiency. For example, both JAK1 and JAK3 have been implicated in IL-7R-mediated SCID [36], and IL-2R and JAK1 in JAK3-mediated SCID [37], with downstream STAT5 playing a role in the SCID phenotypes mediated by either IL-2Rγ [35] or JAK3 [37]. Similarly, analysis of hax1 KD zebrafish has identified a key role for decreased expression of target genes within the G-CSF pathway, which could be reversed by the addition of G-CSF [52]. This approach has been extended to understanding the complex genetic interactions across a suite of genes that cause T cell deficiency when mutated in zebrafish [139]. Zebrafish are further contributing to new biochemical understanding of these diseases, such as decreased lipid storage and ribosomal proteins within neutrophils following srp54 ablation [54], as well as an abnormally sustained H2O2 burst in mpo mutants, indicating a crucial role in H2O2 downregulation [61,62]. Such biochemical knowledge is leading to the development of new strategies for therapy. For example, localized activation of a photoactivable Rac-GTP has been identified as a novel therapeutic approach to overcome the defective neutrophil trafficking due to constitutive CXCR4/SDF-1 signaling [58].
7. Limitations and Future Prospects
Collectively the studies presented illustrate the strengths of zebrafish as a model for human PIDs. However, there are also a number of limitations that need to be considered. For example, many infection studies have concentrated on embryonic and larval stages, prior to full adaptive immunity being present, providing an incomplete picture of host responses [107]. Microbiome studies also need to be interpreted with some caution due to differences in intestinal anatomy and core microbiota [140], including the absence of strictly anaerobic bacteria in zebrafish [109], with longitudinal studies not possible [141]. Regeneration studies have been generally limited to zebrafish tissues [115,116], making the translation to human cells and tissues difficult. Finally, xenotransplantation studies typically employ compromised temperatures and other conditions to enable human cells to be propagated in zebrafish [120]. For some studies, mouse models may be superior, although they too have limitations [142].
However, the full prospects for zebrafish as a model remain untapped. Future infection studies have the potential to fully investigate host determinants of disease and, combined with chemical screens, to identify potential in patient-specific ‘precision’ therapeutics [107]. Microbiota studies can be extended to simultaneously manipulate the host, bacteria and environment to obtain unprecedented insights into the interplay between these [141]. Regeneration studies can also consider other immune populations, including for example, adaptive immune cells, dendritic cells, mast cells and M1 versus M2 macrophages [19,106]. Future cancer studies can further study the role of angiogenesis [122] and better mimic the human tumor microenvironment [129], including creating humanized models as achieved in mice [142]. Finally, so-called cancer ‘avatars’ have application in patient-specific ‘precision’ cancer therapy pipelines, with the potential to directly predict patient outcomes and thus influence individual patient care [143,144].
8. Conclusions
Zebrafish shows a strong correlation of immune cells and the genes controlling their development and function with humans. This, in combination with the genetic malleability and accessibility of this organism, has underpinned its broad application to modelling of a wide range of human PIDs, including multiple examples of both T−B− and T−B+ SCID, CID, multi-system ID, autoinflammatory disorders, congenital neutropenia as well as leucocyte mobility and respiratory burst defects. This has allowed precise and robust analysis of these genetic disorders without the limitations of human studies, providing exciting new insights into these diseases. In addition, a variety of zebrafish immunodeficiency models are being applied to the study of infection, immunity, the microbiome, tissue regeneration and repair, as well as tumor biology. This is leading to unprecedented new knowledge and novel possibilities for treatment. This includes true precision medicine, where an individual patient’s specific mutation(s) can be modeled—and various tailored therapies extensively tested—employing zebrafish models. There are limitations to the use of zebrafish but also unique opportunities unavailable in other experimental platforms. Collectively, this shows that zebrafish is indeed a relevant genetic model for human PIDs.
Author Contributions
Conceptualization, F.B., R.S. and A.C.W.; writing—original draft preparation, F.B., R.S. and A.C.W.; writing—review and editing, F.B., R.S., C.L. and A.C.W. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data was created or analyzed in this study.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Kraus, R.F.; Gruber, M.A. Neutrophils—From bone marrow to first-line defense of the innate immune system. Front. Immunol. 2021, 12, 767175. [Google Scholar] [CrossRef]
- Gharavi, A.T.; Hanjani, N.A.; Movahed, E.; Doroudian, M. The role of macrophage subtypes and exosomes in immunomodulation. Cell. Mol. Biol. Lett. 2022, 27, 83. [Google Scholar] [CrossRef]
- Marshall, J.S.; Warrington, R.; Watson, W.; Kim, H.L. An introduction to immunology and immunopathology. Allergy Asthma Clin. Immunol. 2018, 14, 49. [Google Scholar] [CrossRef] [Green Version]
- Tangye, S.G.; Al-Herz, W.; Bousfiha, A.; Cunningham-Rundles, C.; Franco, J.L.; Holland, S.M.; Klein, C.; Morio, T.; Oksenhendler, E.; Picard, C.; et al. Human inborn errors of immunity: 2022 update on the classification from the International Union of Immunological Societies Expert Committee. J. Clin. Immunol. 2022, 42, 1473–1507. [Google Scholar] [CrossRef] [PubMed]
- McCusker, C.; Upton, J.; Warrington, R. Primary immunodeficiency. Allergy Asthma Clin. Immunol. 2018, 14, 61. [Google Scholar] [CrossRef]
- Rasighaemi, P.; Basheer, F.; Liongue, C.; Ward, A.C. Zebrafish as a model for leukemia and other hematopoietic disorders. J. Hematol. Oncol. 2015, 8, 29. [Google Scholar] [CrossRef] [Green Version]
- Iwanami, N. Zebrafish as a model for understanding the evolution of the vertebrate immune system and human primary immunodeficiency. Exp. Hematol. 2014, 42, 697–706. [Google Scholar] [CrossRef] [PubMed]
- Rafferty, S.A.; Quinn, T.A. A beginner’s guide to understanding and implementing the genetic modification of zebrafish. Prog. Biophys. Mol. Biol. 2018, 138, 3–19. [Google Scholar] [CrossRef]
- Otterstrom, J.J.; Lubin, A.; Payne, E.M.; Paran, Y. Technologies bringing young Zebrafish from a niche field to the limelight. SLAS Technol. 2022, 27, 109–120. [Google Scholar] [CrossRef] [PubMed]
- Rissone, A.; Burgess, S.M. Rare genetic blood disease modeling in zebrafish. Front. Genet. 2018, 9, 348. [Google Scholar] [CrossRef] [Green Version]
- Konantz, M.; Schurch, C.; Hanns, P.; Muller, J.S.; Sauteur, L.; Lengerke, C. Modeling hematopoietic disorders in zebrafish. Dis. Model. Mech. 2019, 12, dmm040360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stosik, M.; Tokarz-Deptula, B.; Deptula, W. Haematopoiesis in zebrafish (Danio rerio). Front. Immunol. 2022, 13, 902941. [Google Scholar] [CrossRef]
- Hu, Y.X.; Jing, Q. Zebrafish: A convenient tool for myelopoiesis research. Cell Regen. 2023, 12, 2. [Google Scholar] [CrossRef]
- Faisal, M.; Hassan, M.; Kumar, A.; Zubair, M.; Jamal, M.; Menghwar, H.; Saad, M.; Kloczkowski, A. Hematopoietic stem and progenitor cells (HSPCs) and hematopoietic microenvironment: Molecular and bioinformatic studies of the zebrafish models. Int. J. Mol. Sci. 2022, 23, 7285. [Google Scholar] [CrossRef]
- Bajoghli, B.; Dick, A.M.; Claasen, A.; Doll, L.; Aghaallaei, N. Zebrafish and medaka: Two teleost models of T-cell and thymic development. Int. J. Mol. Sci. 2019, 20, 4179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moore, F.E.; Garcia, E.G.; Lobbardi, R.; Jain, E.; Tang, Q.; Moore, J.C.; Cortes, M.; Molodtsov, A.; Kasheta, M.; Luo, C.C. Single-cell transcriptional analysis of normal, aberrant, and malignant hematopoiesis in zebrafish. J. Exp. Med. 2016, 213, 979–992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lieschke, G.J.; Oates, A.C.; Crowhurst, M.O.; Ward, A.C.; Layton, J.E. Morphologic and functional characterization of granulocytes and macrophages in embryonic and adult zebrafish. Blood 2001, 98, 3087–3096. [Google Scholar] [CrossRef] [PubMed]
- Yoong, S.; O’Connell, B.; Soanes, A.; Crowhurst, M.O.; Lieschke, G.J.; Ward, A.C. Characterization of the zebrafish matrix metalloproteinase 9 gene and its developmental expression pattern. Gene Expr. Patterns 2007, 7, 39–46. [Google Scholar] [CrossRef]
- Rosowski, E.E. Determining macrophage versus neutrophil contributions to innate immunity using larval zebrafish. Dis. Model. Mech. 2020, 13, dmm041889. [Google Scholar] [CrossRef] [Green Version]
- Herbomel, P.; Thisse, B.; Thisse, C. Ontogeny and behaviour of early macrophages in the zebrafish embryo. Development 1999, 126, 3735–3745. [Google Scholar] [CrossRef]
- Lewis, K.L.; Del Cid, N.; Traver, D. Perspectives on antigen presenting cells in zebrafish. Dev. Comp. Immunol. 2014, 46, 63–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balla, K.M.; Lugo-Villarino, G.; Spitsbergen, J.M.; Stachura, D.L.; Hu, Y.; Bañuelos, K.; Romo-Fewell, O.; Aroian, R.V.; Traver, D. Eosinophils in the zebrafish: Prospective isolation, characterization, and eosinophilia induction by helminth determinants. Blood 2010, 116, 3944–3954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Da’as, S.; Teh, E.M.; Dobson, J.T.; Nasrallah, G.K.; McBride, E.R.; Wang, H.; Neuberg, D.S.; Marshall, J.S.; Lin, T.-J.; Berman, J.N. Zebrafish mast cells possess an FcɛRI-like receptor and participate in innate and adaptive immune responses. Dev. Comp. Immunol. 2011, 35, 125–134. [Google Scholar] [CrossRef] [PubMed]
- Wan, F.; Hu, C.-B.; Ma, J.-X.; Gao, K.; Xiang, L.-X.; Shao, J.-Z. Characterization of γδ T cells from zebrafish provides insights into their important role in adaptive humoral immunity. Front. Immunol. 2017, 7, 675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zimmerman, A.M.; Moustafa, F.M.; Romanowski, K.E.; Steiner, L.A. Zebrafish immunoglobulin IgD: Unusual exon usage and quantitative expression profiles with IgM and IgZ/T heavy chain isotypes. Mol. Immunol. 2011, 48, 2220–2223. [Google Scholar] [CrossRef] [Green Version]
- Tang, Q.; Iyer, S.; Lobbardi, R.; Moore, J.C.; Chen, H.; Lareau, C.; Hebert, C.; Shaw, M.L.; Neftel, C.; Suva, M.L. Dissecting hematopoietic and renal cell heterogeneity in adult zebrafish at single-cell resolution using RNA sequencing. J. Exp. Med. 2017, 214, 2875–2887. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Lu, Y.; He, Y.; Feng, Z.; Zhan, Y.; Huang, X.; Liu, Q.; Zhang, J.; Li, H.; Huang, H. Ikzf1 regulates embryonic T lymphopoiesis via Ccr9 and Irf4 in zebrafish. J. Biol. Chem. 2019, 294, 16152–16163. [Google Scholar] [CrossRef]
- Liongue, C.; Taznin, T.; Ward, A.C. Signaling via the CytoR/JAK/STAT/SOCS pathway: Emergence during evolution. Mol. Immunol. 2016, 71, 166–175. [Google Scholar] [CrossRef]
- Sommer, F.; Torraca, V.; Meijer, A.H. Chemokine receptors and phagocyte biology in zebrafish. Front. Immunol. 2020, 11, 325. [Google Scholar] [CrossRef] [Green Version]
- Wienholds, E.; Schulte-Merker, S.; Walderich, B.; Plasterk, R.H. Target-selected inactivation of the zebrafish rag1 gene. Science 2002, 297, 99–102. [Google Scholar] [CrossRef]
- Petrie-Hanson, L.; Hohn, C.; Hanson, L. Characterization of rag 1 mutant zebrafish leukocytes. BMC Immunol. 2009, 10, 8. [Google Scholar] [CrossRef] [Green Version]
- Tang, Q.; Abdelfattah, N.S.; Blackburn, J.S.; Moore, J.C.; Martinez, S.A.; Moore, F.E.; Lobbardi, R.; Tenente, I.M.; Ignatius, M.S.; Berman, J.N. Optimized cell transplantation using adult rag2 mutant zebrafish. Nat. Methods 2014, 11, 821–824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, I.H.; Chung, Y.-Y.; Jung, D.E.; Kim, Y.J.; Kim, K.-S.; Park, S.W. Impaired lymphocytes development and xenotransplantation of gastrointestinal tumor cells in prkdc-null SCID zebrafish model. Neoplasia 2016, 18, 468–479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rissone, A.; Weinacht, K.G.; la Marca, G.; Bishop, K.; Giocaliere, E.; Jagadeesh, J.; Felgentreff, K.; Dobbs, K.; Al-Herz, W.; Jones, M.; et al. Reticular dysgenesis-associated AK2 protects hematopoietic stem and progenitor cell development from oxidative stress. J. Exp. Med. 2015, 212, 1185–1202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sertori, R.; Jones, R.; Basheer, F.; Rivera, L.; Dawson, S.; Loke, S.; Heidary, S.; Dhillon, A.; Liongue, C.; Ward, A.C. Generation and characterization of a zebrafish IL-2Rgc SCID model. Int. J. Mol. Sci. 2022, 23, 2385. [Google Scholar] [CrossRef] [PubMed]
- Iwanami, N.; Mateos, F.; Hess, I.; Riffel, N.; Soza-Ried, C.; Schorpp, M.; Boehm, T. Genetic evidence for an evolutionarily conserved role of IL-7 signaling in T cell development of zebrafish. J. Immunol. 2011, 186, 7060–7066. [Google Scholar] [CrossRef] [Green Version]
- Basheer, F.; Lee, E.; Liongue, C.; Ward, A.C. Zebrafish model of severe combined immunodeficiency (SCID) due to JAK3 mutation. Biomolecules 2022, 12, 1521. [Google Scholar] [CrossRef]
- Moore, J.C.; Mulligan, T.S.; Yordán, N.T.; Castranova, D.; Pham, V.N.; Tang, Q.; Lobbardi, R.; Anselmo, A.; Liwski, R.S.; Berman, J.N. T cell immune deficiency in zap70 mutant zebrafish. Mol. Cell. Biol. 2016, 36, 2868–2876. [Google Scholar] [CrossRef] [Green Version]
- Schorpp, M.; Bialecki, M.; Diekhoff, D.; Walderich, B.; Odenthal, J.; Maischein, H.-M.; Zapata, A.G.; Boehm, T. Conserved functions of Ikaros in vertebrate lymphocyte development: Genetic evidence for distinct larval and adult phases of T cell development and two lineages of B cells in zebrafish. J. Immunol. 2006, 177, 2463–2476. [Google Scholar] [CrossRef] [Green Version]
- Heidary, S.; Awasthi, N.; Page, N.; Lewis, R.S.; Liongue, C.; Ward, A.C. A zebrafish model of growth hormone insensitivity syndrome with immune dysregulation 1 (GHISID1). Cell. Mol. Life Sci. 2023, in press. [Google Scholar]
- Volpi, S.; Yamazaki, Y.; Brauer, P.M.; van Rooijen, E.; Hayashida, A.; Slavotinek, A.; Sun Kuehn, H.; Di Rocco, M.; Rivolta, C.; Bortolomai, I.; et al. EXTL3 mutations cause skeletal dysplasia, immune deficiency, and developmental delay. J. Exp. Med. 2017, 214, 623–637. [Google Scholar] [CrossRef] [Green Version]
- Somech, R.; Lev, A.; Lee, Y.N.; Simon, A.J.; Barel, O.; Schiby, G.; Avivi, C.; Barshack, I.; Rhodes, M.; Yin, J.; et al. Disruption of thrombocyte and T lymphocyte development by a mutation in ARPC1B. J. Immunol. 2017, 199, 4036–4045. [Google Scholar] [CrossRef] [Green Version]
- Putti, S.; Giovinazzo, A.; Merolle, M.; Falchetti, M.L.; Pellegrini, M. ATM kinase dead: From ataxia telangiectasia syndrome to cancer. Cancers 2021, 13, 5498. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.Y.; Sertori, R.; Contreras, A.V.; Hamer, M.; Messing, M.; Del Bel, K.L.; Lopez-Rangel, E.; Chan, E.S.; Rehmus, W.; Milner, J.D.; et al. A novel germline heterozygous BCL11B variant causing severe atopic disease and immune dysregulation. Front. Immunol. 2021, 12, 788278. [Google Scholar] [CrossRef] [PubMed]
- Ma, D.; Wang, L.; Wang, S.; Gao, Y.; Wei, Y.; Liu, F. Foxn1 maintains thymic epithelial cells to support T-cell development via mcm2 in zebrafish. Proc. Natl. Acad. Sci. USA 2012, 109, 21040–21045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Z.-Z.; Wang, Z.-L.; Choi, T.-I.; Huang, W.-T.; Wang, H.-T.; Han, Y.-Y.; Zhu, L.-Y.; Kim, H.-T.; Choi, J.-H.; Lee, J.-S. Chd7 is critical for early T-cell development and thymus organogenesis in zebrafish. Am. J. Pathol. 2018, 188, 1043–1058. [Google Scholar] [CrossRef] [Green Version]
- Piotrowski, T.; Ahn, D.G.; Schilling, T.F.; Nair, S.; Ruvinsky, I.; Geisler, R.; Rauch, G.J.; Haffter, P.; Zon, L.I.; Zhou, Y.; et al. The zebrafish van gogh mutation disrupts tbx1, which is involved in the DiGeorge deletion syndrome in humans. Development 2003, 130, 5043–5052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castro, C.N.; Rosenzwajg, M.; Carapito, R.; Shahrooei, M.; Konantz, M.; Khan, A.; Miao, Z.; Gross, M.; Tranchant, T.; Radosavljevic, M.; et al. NCKAP1L defects lead to a novel syndrome combining immunodeficiency, lymphoproliferation, and hyperinflammation. J. Exp. Med. 2020, 217, e20192275. [Google Scholar] [CrossRef]
- Basheer, F.; Rasighaemi, P.; Liongue, C.; Ward, A.C. Zebrafish granulocyte colony-stimulating factor receptor maintains neutrophil number and function throughout the life span. Infect. Immun. 2019, 87, e00793-18. [Google Scholar] [CrossRef] [Green Version]
- Witzel, M.; Petersheim, D.; Fan, Y.; Bahrami, E.; Racek, T.; Rohlfs, M.; Puchalka, J.; Mertes, C.; Gagneur, J.; Ziegenhain, C.; et al. Chromatin-remodeling factor SMARCD2 regulates transcriptional networks controlling differentiation of neutrophil granulocytes. Nat. Genet. 2017, 49, 742–752. [Google Scholar] [CrossRef]
- Jones, R.A.; Feng, Y.; Worth, A.J.; Thrasher, A.J.; Burns, S.O.; Martin, P. Modelling of human Wiskott-Aldrich syndrome protein mutants in zebrafish larvae using in vivo live imaging. J. Cell Sci. 2013, 126, 4077–4084. [Google Scholar] [CrossRef] [Green Version]
- Doll, L.; Aghaallaei, N.; Dick, A.M.; Welte, K.; Skokowa, J.; Bajoghli, B. A zebrafish model for HAX1-associated congenital neutropenia. Haematologica 2021, 106, 1311–1320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vilboux, T.; Lev, A.; Malicdan, M.C.; Simon, A.J.; Jarvinen, P.; Racek, T.; Puchalka, J.; Sood, R.; Carrington, B.; Bishop, K.; et al. A congenital neutrophil defect syndrome associated with mutations in VPS45. N. Engl. J. Med. 2013, 369, 54–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oyarbide, U.; Shah, A.N.; Amaya-Mejia, W.; Snyderman, M.; Kell, M.J.; Allende, D.S.; Calo, E.; Topczewski, J.; Corey, S.J. Loss of Sbds in zebrafish leads to neutropenia and pancreas and liver atrophy. JCI Insight 2020, 5, e134309. [Google Scholar] [CrossRef] [PubMed]
- Bader, A.; Gao, J.; Riviere, T.; Schmid, B.; Walzog, B.; Maier-Begandt, D. Molecular insights into neutrophil biology from the zebrafish perspective: Lessons from CD18 deficiency. Front. Immunol. 2021, 12, 677994. [Google Scholar] [CrossRef]
- Deng, Q.; Yoo, S.K.; Cavnar, P.J.; Green, J.M.; Huttenlocher, A. Dual roles for Rac2 in neutrophil motility and active retention in zebrafish hematopoietic tissue. Dev. Cell 2011, 21, 735–745. [Google Scholar] [CrossRef] [Green Version]
- Rosowski, E.E.; Deng, Q.; Keller, N.P.; Huttenlocher, A. Rac2 functions in both neutrophils and macrophages to mediate motility and host defense in larval zebrafish. J. Immunol. 2016, 197, 4780–4790. [Google Scholar] [CrossRef] [Green Version]
- Walters, K.B.; Green, J.M.; Surfus, J.C.; Yoo, S.K.; Huttenlocher, A. Live imaging of neutrophil motility in a zebrafish model of WHIM syndrome. Blood 2010, 116, 2803–2811. [Google Scholar] [CrossRef] [Green Version]
- Tauzin, S.; Starnes, T.W.; Becker, F.B.; Lam, P.Y.; Huttenlocher, A. Redox and Src family kinase signaling control leukocyte wound attraction and neutrophil reverse migration. J. Cell Biol. 2014, 207, 589–598. [Google Scholar] [CrossRef]
- Schoen, T.J.; Rosowski, E.E.; Knox, B.P.; Bennin, D.; Keller, N.P.; Huttenlocher, A. Neutrophil phagocyte oxidase activity controls invasive fungal growth and inflammation in zebrafish. J. Cell Sci. 2019, 133, jcs236539. [Google Scholar] [CrossRef]
- Pase, L.; Layton, J.E.; Wittmann, C.; Ellett, F.; Nowell, C.J.; Reyes-Aldasoro, C.C.; Varma, S.; Rogers, K.L.; Hall, C.J.; Keightley, M.C. Neutrophil-delivered myeloperoxidase dampens the hydrogen peroxide burst after tissue wounding in zebrafish. Curr. Biol. 2012, 22, 1818–1824. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.; Fang, X.; Ma, N.; Lin, Q.; Huang, Z.; Liu, W.; Xu, M.; Chen, X.; Zhang, W.; Zhang, Y. Myeloperoxidase-deficient zebrafish show an augmented inflammatory response to challenge with Candida albicans. Fish Shellfish. Immunol. 2015, 44, 109–116. [Google Scholar] [CrossRef] [PubMed]
- Fischer, A. Severe combined immunodeficiencies (SCID). Clin. Exp. Immunol. 2000, 122, 143–149. [Google Scholar] [CrossRef] [PubMed]
- Michniacki, T.F.; Seth, D.; Secord, E. Severe Combined Immunodeficiency: A review for neonatal clinicians. Neoreviews 2019, 20, e326–e335. [Google Scholar] [CrossRef] [PubMed]
- Bosticardo, M.; Pala, F.; Notarangelo, L.D. RAG deficiencies: Recent advances in disease pathogenesis and novel therapeutic approaches. Eur. J. Immunol. 2021, 51, 1028–1038. [Google Scholar] [CrossRef]
- Hoenig, M.; Pannicke, U.; Gaspar, H.B.; Schwarz, K. Recent advances in understanding the pathogenesis and management of reticular dysgenesis. Br. J. Haematol. 2018, 180, 644–653. [Google Scholar] [CrossRef]
- Hernandez, J.D.; Hsieh, E.W.Y. A great disturbance in the force: IL-2 receptor defects disrupt immune homeostasis. Curr. Opin. Pediatr. 2022, 34, 580–588. [Google Scholar] [CrossRef]
- Sertori, R.; Liongue, C.; Basheer, F.; Lewis, K.L.; Rasighaemi, P.; de Coninck, D.; Traver, D.; Ward, A.C. Conserved IL-2Rgc signaling mediates lymphopoiesis in zebrafish. J. Immunol. 2016, 196, 135–143. [Google Scholar] [CrossRef] [Green Version]
- Ashouri, J.F.; Lo, W.L.; Nguyen, T.T.T.; Shen, L.; Weiss, A. ZAP70, too little, too much can lead to autoimmunity. Immunol. Rev. 2022, 307, 145–160. [Google Scholar] [CrossRef]
- Boast, B.; Nunes-Santos, C.J.; Kuehn, H.S.; Rosenzweig, S.D. Ikaros-associated diseases: From mice to humans and back again. Front. Pediatr. 2021, 9, 705497. [Google Scholar] [CrossRef]
- Smith, M.R.; Satter, L.R.F.; Vargas-Hernandez, A. STAT5b: A master regulator of key biological pathways. Front. Immunol. 2022, 13, 1025373. [Google Scholar] [CrossRef]
- Xiong, S.; Mei, J.; Huang, P.; Jing, J.; Li, Z.; Kang, J.; Gui, J.F. Essential roles of stat5.1/stat5b in controlling fish somatic growth. J. Genet. Genom. 2017, 44, 577–585. [Google Scholar] [CrossRef] [PubMed]
- Piotrowski, T.; Schilling, T.F.; Brand, M.; Jiang, Y.J.; Heisenberg, C.P.; Beuchle, D.; Grandel, H.; van Eeden, F.J.; Furutani-Seiki, M.; Granato, M.; et al. Jaw and branchial arch mutants in zebrafish II: Anterior arches and cartilage differentiation. Development 1996, 123, 345–356. [Google Scholar] [CrossRef] [PubMed]
- Dupre, L.; Prunier, G. Deciphering actin remodelling in immune cells through the prism of actin-related inborn errors of immunity. Eur. J. Cell Biol. 2023, 102, 151283. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Wang, P.; Chen, J.; Ying, Y.; Chen, Y.; Gilson, E.; Lu, Y.; Ye, J. Loss of atm in zebrafish as a model of ataxia-telangiectasia syndrome. Biomedicines 2022, 10, 392. [Google Scholar] [CrossRef]
- Punwani, D.; Zhang, Y.; Yu, J.; Cowan, M.J.; Rana, S.; Kwan, A.; Adhikari, A.N.; Lizama, C.O.; Mendelsohn, B.A.; Fahl, S.P. Multisystem anomalies in severe combined immunodeficiency with mutant BCL11B. N. Engl. J. Med. 2016, 375, 2165–2176. [Google Scholar] [CrossRef] [PubMed]
- Collins, C.; Sharpe, E.; Silber, A.; Kulke, S.; Hsieh, E.W.Y. Congenital athymia: Genetic etiologies, clinical manifestations, diagnosis, and treatment. J. Clin. Immunol. 2021, 41, 881–895. [Google Scholar] [CrossRef] [PubMed]
- Lv, P.; Ma, D.; Gao, S.; Zhang, Y.; Bae, Y.-K.; Liang, G.; Gao, S.; Choi, J.-H.; Kim, C.-H.; Wang, L. Generation of foxn1/casper mutant zebrafish for allograft and xenograft of normal and malignant cells. Stem Cell Rep. 2020, 15, 749–760. [Google Scholar] [CrossRef]
- Liongue, C.; Ward, A.C. Granulocyte colony-stimulating factor receptor mutations in myeloid malignancy. Front. Oncol. 2014, 4, 93. [Google Scholar] [CrossRef] [Green Version]
- Pazhakh, V.; Clark, S.; Keightley, M.C.; Lieschke, G.J. A GCSFR/CSF3R zebrafish mutant models the persistent basal neutrophil deficiency of severe congenital neutropenia. Sci. Rep. 2017, 7, 44455. [Google Scholar] [CrossRef] [Green Version]
- Klein, C. Kostmann’s disease and HCLS1-associated protein X-1 (HAX1). J. Clin. Immunol. 2017, 37, 117–122. [Google Scholar] [CrossRef]
- Bellanne-Chantelot, C.; Schmaltz-Panneau, B.; Marty, C.; Fenneteau, O.; Callebaut, I.; Clauin, S.; Docet, A.; Damaj, G.L.; Leblanc, T.; Pellier, I.; et al. Mutations in the SRP54 gene cause severe congenital neutropenia as well as Shwachman-Diamond-like syndrome. Blood 2018, 132, 1318–1331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fekadu, J.; Modlich, U.; Bader, P.; Bakhtiar, S. Understanding the role of LFA-1 in leukocyte adhesion deficiency type I (LAD I): Moving towards inflammation? Int. J. Mol. Sci. 2022, 23, 3578. [Google Scholar] [CrossRef] [PubMed]
- Lougaris, V.; Baronio, M.; Gazzurelli, L.; Benvenuto, A.; Plebani, A. RAC2 and primary human immune deficiencies. J. Leukoc. Biol. 2020, 108, 687–696. [Google Scholar] [CrossRef] [PubMed]
- McDermott, D.H.; Murphy, P.M. WHIM syndrome: Immunopathogenesis, treatment and cure strategies. Immunol. Rev. 2019, 287, 91–102. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.H.; Yang, Y.H.; Chiang, B.L. Chronic granulomatous disease: A comprehensive review. Clin. Rev. Allergy Immunol. 2021, 61, 101–113. [Google Scholar] [CrossRef]
- Van der Veen, B.S.; de Winther, M.P.; Heeringa, P. Myeloperoxidase: Molecular mechanisms of action and their relevance to human health and disease. Antioxid. Redox Signal. 2009, 11, 2899–2937. [Google Scholar] [CrossRef]
- Trede, N.S.; Medenbach, J.; Damianov, A.; Hung, L.H.; Weber, G.J.; Paw, B.H.; Zhou, Y.; Hersey, C.; Zapata, A.; Keefe, M.; et al. Network of coregulated spliceosome components revealed by zebrafish mutant in recycling factor p110. Proc. Natl. Acad. Sci. USA 2007, 104, 6608–6613. [Google Scholar] [CrossRef] [Green Version]
- Chi, Y.; Huang, Z.; Chen, Q.; Xiong, X.; Chen, K.; Xu, J.; Zhang, Y.; Zhang, W. Loss of runx1 function results in B cell immunodeficiency but not T cell in adult zebrafish. Open Biol. 2018, 8, 2046–2441. [Google Scholar] [CrossRef] [Green Version]
- Xie, L.; Tao, Y.; Wu, R.; Ye, Q.; Xu, H.; Li, Y. Congenital asplenia due to a tlx1 mutation reduces resistance to Aeromonas hydrophila infection in zebrafish. Fish Shellfish. Immunol. 2019, 95, 538–545. [Google Scholar] [CrossRef]
- Lawir, D.-F.; Iwanami, N.; Schorpp, M.; Boehm, T. A missense mutation in zbtb17 blocks the earliest steps of T cell differentiation in zebrafish. Sci. Rep. 2017, 7, 44145. [Google Scholar] [CrossRef] [Green Version]
- Hess, I.; Iwanami, N.; Schorpp, M.; Boehm, T. Zebrafish model for allogeneic hematopoietic cell transplantation not requiring preconditioning. Proc. Natl. Acad. Sci. USA 2013, 110, 4327–4332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, P.; Mao, X.; Zhao, J.; Luo, L. Ribosome biogenesis protein Urb2 regulates hematopoietic stem cells development via P53 pathway in zebrafish. Biochem. Biophys. Res. Commun. 2018, 497, 776–782. [Google Scholar] [CrossRef] [PubMed]
- Simon, A.J.; Lev, A.; Zhang, Y.; Weiss, B.; Rylova, A.; Eyal, E.; Kol, N.; Barel, O.; Cesarkas, K.; Soudack, M. Mutations in STN1 cause Coats plus syndrome and are associated with genomic and telomere defects. J. Exp. Med. 2016, 213, 1429–1440. [Google Scholar] [CrossRef] [Green Version]
- Wong, H.H.; Seet, S.H.; Maier, M.; Gurel, A.; Traspas, R.M.; Lee, C.; Zhang, S.; Talim, B.; Loh, A.Y.T.; Chia, C.Y.; et al. Loss of C2orf69 defines a fatal autoinflammatory syndrome in humans and zebrafish that evokes a glycogen-storage-associated mitochondriopathy. Am. J. Hum. Genet. 2021, 108, 1301–1317. [Google Scholar] [CrossRef]
- Linder, M.I.; Mizoguchi, Y.; Hesse, S.; Csaba, G.; Tatematsu, M.; Lyszkiewicz, M.; Zietara, N.; Jeske, T.; Hastreiter, M.; Rohlfs, M.; et al. Human genetic defects in SRP19 and SRPRA cause severe congenital neutropenia with distinctive proteome changes. Blood 2023, 141, 645–658. [Google Scholar] [CrossRef] [PubMed]
- Yue, Z.; Fan, Z.; Zhang, H.; Feng, B.; Wu, C.; Chen, S.; Ouyang, J.; Fan, H.; Weng, P.; Feng, H.; et al. Differential roles of the fish chitinous membrane in gut barrier immunity and digestive compartments. EMBO Rep. 2023, e56645. [Google Scholar] [CrossRef]
- Loes, A.N.; Hinman, M.N.; Farnsworth, D.R.; Miller, A.C.; Guillemin, K.; Harms, M.J. Identification and characterization of zebrafish Tlr4 coreceptor Md-2. J. Immunol. 2021, 206, 1046–1057. [Google Scholar] [CrossRef]
- Myllymaki, H.; Yu, P.P.; Feng, Y. Opportunities presented by zebrafish larval models to study neutrophil function in tissues. Int. J. Biochem. Cell Biol. 2022, 148, 106234. [Google Scholar] [CrossRef]
- Swaim, L.E.; Connolly, L.E.; Volkman, H.E.; Humbert, O.; Born, D.E.; Ramakrishnan, L. Mycobacterium marinum infection of adult zebrafish causes caseating granulomatous tuberculosis and is moderated by adaptive immunity. Infect. Immun. 2006, 74, 6108–6117. [Google Scholar] [CrossRef] [Green Version]
- Hohn, C.; Petrie-Hanson, L. Rag1−/− mutant zebrafish demonstrate specific protection following bacterial re-exposure. PLoS ONE 2012, 7, e44451. [Google Scholar] [CrossRef] [Green Version]
- van der Vaart, M.; van Soest, J.J.; Spaink, H.P.; Meijer, A.H. Functional analysis of a zebrafish myd88 mutant identifies key transcriptional components of the innate immune system. Dis. Model. Mech. 2013, 6, 841–854. [Google Scholar] [PubMed] [Green Version]
- Gomes, M.C.; Mostowy, S. The case for modeling human infection in zebrafish. Trends Microbiol. 2020, 28, 10–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kam, J.Y.; Hortle, E.; Krogman, E.; Warner, S.E.; Wright, K.; Luo, K.; Cheng, T.; Manuneedhi Cholan, P.; Kikuchi, K.; Triccas, J.A. Rough and smooth variants of Mycobacterium abscessus are differentially controlled by host immunity during chronic infection of adult zebrafish. Nat. Commun. 2022, 13, 952. [Google Scholar] [CrossRef]
- Farr, D.; Nag, D.; Chazin, W.J.; Harrison, S.; Thummel, R.; Luo, X.; Raychaudhuri, S.; Withey, J.H. Neutrophil-associated responses to Vibrio cholerae infection in a natural host model. Infect. Immun. 2022, 90, e0046621. [Google Scholar] [CrossRef] [PubMed]
- Robertson, T.F.; Huttenlocher, A. Real-time imaging of inflammation and its resolution: It’s apparent because it’s transparent. Immunol. Rev. 2022, 306, 258–270. [Google Scholar] [CrossRef] [PubMed]
- Linnerz, T.; Hall, C.J. The diverse roles of phagocytes during bacterial and fungal infections and sterile inflammation: Lessons from zebrafish. Front. Immunol. 2020, 11, 1094. [Google Scholar] [CrossRef]
- Stagaman, K.; Sharpton, T.J.; Guillemin, K. Zebrafish microbiome studies make waves. Lab. Anim. 2020, 49, 201–207. [Google Scholar] [CrossRef]
- Cornuault, J.K.; Byatt, G.; Paquet, M.E.; De Koninck, P.; Moineau, S. Zebrafish: A big fish in the study of the gut microbiota. Curr. Opin. Biotechnol. 2022, 73, 308–313. [Google Scholar] [CrossRef]
- Lane, J.P.; Stewart, C.J.; Cummings, S.P.; Gennery, A.R. Gut microbiome variations during hematopoietic stem cell transplant in severe combined immunodeficiency. J. Allergy Clin. Immunol. 2015, 135, 1654–1656.e1652. [Google Scholar] [CrossRef]
- Brugman, S.; Schneeberger, K.; Witte, M.; Klein, M.R.; van den Bogert, B.; Boekhorst, J.; Timmerman, H.M.; Boes, M.L.; Kleerebezem, M.; Nieuwenhuis, E.E. T lymphocytes control microbial composition by regulating the abundance of Vibrio in the zebrafish gut. Gut Microbes 2014, 5, 737–747. [Google Scholar] [CrossRef]
- Burns, A.R.; Miller, E.; Agarwal, M.; Rolig, A.S.; Milligan-Myhre, K.; Seredick, S.; Guillemin, K.; Bohannan, B.J. Interhost dispersal alters microbiome assembly and can overwhelm host innate immunity in an experimental zebrafish model. Proc. Natl. Acad. Sci. USA 2017, 114, 11181–11186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Earley, A.M.; Graves, C.L.; Shiau, C.E. Critical role for a subset of intestinal macrophages in shaping gut microbiota in adult zebrafish. Cell Rep. 2018, 25, 424–436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marques, I.J.; Lupi, E.; Mercader, N. Model systems for regeneration: Zebrafish. Development 2019, 146, dev167692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Francoeur, N.; Sen, R. Advances in cardiac development and regeneration using zebrafish as a model system for high-throughput research. J. Dev. Biol. 2021, 9, 40. [Google Scholar] [CrossRef]
- Henke, K.; Farmer, D.T.; Niu, X.; Kraus, J.M.; Galloway, J.L.; Youngstrom, D.W. Genetically engineered zebrafish as models of skeletal development and regeneration. Bone 2023, 167, 116611. [Google Scholar] [CrossRef]
- Kikuchi, K. New function of zebrafish regulatory T cells in organ regeneration. Curr. Opin. Immunol. 2020, 63, 7–13. [Google Scholar] [CrossRef]
- Li, L.; Yan, B.; Shi, Y.-Q.; Zhang, W.-Q.; Wen, Z.-L. Live imaging reveals differing roles of macrophages and neutrophils during zebrafish tail fin regeneration. J. Biol. Chem. 2012, 287, 25353–25360. [Google Scholar] [CrossRef] [Green Version]
- Var, S.R.; Byrd-Jacobs, C.A. Role of macrophages and microglia in zebrafish regeneration. Int. J. Mol. Sci. 2020, 21, 4768. [Google Scholar] [CrossRef]
- Campbell, C.A.; Fursova, O.; Cheng, X.; Snella, E.; McCune, A.; Li, L.; Solchenberger, B.; Schmid, B.; Sahoo, D.; Morton, M. A zebrafish model of granulin deficiency reveals essential roles in myeloid cell differentiation. Blood Adv. 2021, 5, 796–811. [Google Scholar] [CrossRef]
- Bernut, A.; Loynes, C.A.; Floto, R.A.; Renshaw, S.A. Deletion of cftr leads to an excessive neutrophilic response and defective tissue repair in a zebrafish model of sterile inflammation. Front. Immunol. 2020, 11, 1733. [Google Scholar] [CrossRef]
- Kwiatkowska, I.; Hermanowicz, J.M.; Iwinska, Z.; Kowalczuk, K.; Iwanowska, J.; Pawlak, D. Zebrafish—An optimal model in experimental oncology. Molecules 2022, 27, 4223. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Wang, X.; Zhao, Y.; Li, Z.; Lin, S.; Wei, Y.; Yang, H. A novel xenograft model in zebrafish for high-resolution investigating dynamics of neovascularization in tumors. PLoS ONE 2011, 6, e21768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, D.-W.; Oh, E.-S.; Park, S.-H.; Chang, Y.-T.; Kim, C.-H.; Choi, S.-Y.; Williams, D.R. A novel zebrafish human tumor xenograft model validated for anti-cancer drug screening. Mol. BioSystems 2012, 8, 1930–1939. [Google Scholar] [CrossRef]
- Drabsch, Y.; He, S.; Zhang, L.; Snaar-Jagalska, B.E.; ten Dijke, P. Transforming growth factor-β signalling controls human breast cancer metastasis in a zebrafish xenograft model. Breast Cancer Res. 2013, 15, R106. [Google Scholar] [CrossRef]
- Pudelko, L.; Edwards, S.; Balan, M.; Nyqvist, D.; Al-Saadi, J.; Dittmer, J.; Almlöf, I.; Helleday, T.; Bräutigam, L. An orthotopic glioblastoma animal model suitable for high-throughput screenings. Neuro. Oncol. 2018, 20, 1475–1484. [Google Scholar] [CrossRef]
- Vargas-Patron, L.A.; Agudelo-Dueñas, N.; Madrid-Wolff, J.; Venegas, J.A.; González, J.M.; Forero-Shelton, M.; Akle, V. Xenotransplantation of human glioblastoma in zebrafish larvae: In vivo imaging and proliferation assessment. Biol. Open 2019, 8, bio043257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moore, J.C.; Tang, Q.; Yordán, N.T.; Moore, F.E.; Garcia, E.G.; Lobbardi, R.; Ramakrishnan, A.; Marvin, D.L.; Anselmo, A.; Sadreyev, R.I. Single-cell imaging of normal and malignant cell engraftment into optically clear prkdc-null SCID zebrafish. J. Exp. Med. 2016, 213, 2575–2589. [Google Scholar] [CrossRef] [Green Version]
- Weiss, J.M.; Lumaquin-Yin, D.; Montal, E.; Suresh, S.; Leonhardt, C.S.; White, R.M. Shifting the focus of zebrafish toward a model of the tumor microenvironment. Elife 2022, 11, 69703. [Google Scholar] [CrossRef]
- Somasagara, R.R.; Huang, X.; Xu, C.; Haider, J.; Serody, J.S.; Armistead, P.M.; Leung, T. Targeted therapy of human leukemia xenografts in immunodeficient zebrafish. Sci. Rep. 2021, 11, 5715. [Google Scholar] [CrossRef]
- Yan, C.; Brunson, D.C.; Tang, Q.; Do, D.; Iftimia, N.A.; Moore, J.C.; Hayes, M.N.; Welker, A.M.; Garcia, E.G.; Dubash, T.D. Visualizing engrafted human cancer and therapy responses in immunodeficient zebrafish. Cell 2019, 177, 1903–1914.e14. [Google Scholar] [CrossRef]
- Yan, C.; Yang, Q.; Zhang, S.; Millar, D.G.; Alpert, E.J.; Do, D.; Veloso, A.; Brunson, D.C.; Drapkin, B.J.; Stanzione, M. Single-cell imaging of T cell immunotherapy responses in vivo. J. Exp. Med. 2021, 218, e20210314. [Google Scholar] [CrossRef] [PubMed]
- Zeng, M.; Pi, C.; Li, K.; Sheng, L.; Zuo, Y.; Yuan, J.; Zou, Y.; Zhang, X.; Zhao, W.; Lee, R.J. Patient-derived xenograft: A more standard “avatar” model in preclinical studies of gastric cancer. Front. Oncol. 2022, 12, 898563. [Google Scholar] [CrossRef] [PubMed]
- Gaudenzi, G.; Albertelli, M.; Dicitore, A.; Würth, R.; Gatto, F.; Barbieri, F.; Cotelli, F.; Florio, T.; Ferone, D.; Persani, L. Patient-derived xenograft in zebrafish embryos: A new platform for translational research in neuroendocrine tumors. Endocrine 2017, 57, 214–219. [Google Scholar] [CrossRef] [PubMed]
- Tang, Q.; Moore, J.C.; Ignatius, M.S.; Tenente, I.M.; Hayes, M.N.; Garcia, E.G.; Torres Yordán, N.; Bourque, C.; He, S.; Blackburn, J.S. Imaging tumour cell heterogeneity following cell transplantation into optically clear immune-deficient zebrafish. Nat. Commun. 2016, 7, 10358. [Google Scholar] [CrossRef] [Green Version]
- Fior, R.; Póvoa, V.; Mendes, R.V.; Carvalho, T.; Gomes, A.; Figueiredo, N.; Ferreira, M.G. Single-cell functional and chemosensitive profiling of combinatorial colorectal therapy in zebrafish xenografts. Proc. Natl. Acad. Sci. USA 2017, 114, E8234–E8243. [Google Scholar] [CrossRef] [Green Version]
- Costa, B.; Ferreira, S.; Póvoa, V.; Cardoso, M.J.; Vieira, S.; Stroom, J.; Fidalgo, P.; Rio-Tinto, R.; Figueiredo, N.; Parés, O. Developments in zebrafish avatars as radiotherapy sensitivity reporters—Towards personalized medicine. EBioMedicine 2020, 51, 102578. [Google Scholar] [CrossRef] [Green Version]
- Rebelo de Almeida, C.; Mendes, R.V.; Pezzarossa, A.; Gago, J.; Carvalho, C.; Alves, A.; Nunes, V.; Brito, M.J.; Cardoso, M.J.; Ribeiro, J. Zebrafish xenografts as a fast screening platform for bevacizumab cancer therapy. Commun. Biol. 2020, 3, 299. [Google Scholar] [CrossRef]
- O’Meara, C.P.; Guerri, L.; Lawir, D.F.; Mateos, F.; Iconomou, M.; Iwanami, N.; Soza-Ried, C.; Sikora, K.; Siamishi, I.; Giorgetti, O.; et al. Genetic landscape of T cells identifies synthetic lethality for T-ALL. Commun. Biol. 2021, 4, 1201. [Google Scholar] [CrossRef]
- Xia, H.; Chen, H.; Cheng, X.; Yin, M.; Yao, X.; Ma, J.; Huang, M.; Chen, G.; Liu, H. Zebrafish: An efficient vertebrate model for understanding role of gut microbiota. Mol. Med. 2022, 28, 161. [Google Scholar] [CrossRef]
- Murdoch, C.C.; Rawls, J.F. Commensal microbiota regulate vertebrate innate immunity-insights from the zebrafish. Front. Immunol. 2019, 10, 2100. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Liao, S.; Xiao, Z.; Pan, Q.; Wang, X.; Shen, K.; Wang, S.; Yang, L.; Guo, F.; Liu, H.F.; et al. The development and improvement of immunodeficient mice and humanized immune system mouse models. Front. Immunol. 2022, 13, 1007579. [Google Scholar] [CrossRef] [PubMed]
- Al-Hamaly, M.A.; Turner, L.T.; Rivera-Martinez, A.; Rodriguez, A.; Blackburn, J.S. Zebrafish cancer avatars: A translational platform for analyzing tumor heterogeneity and predicting patient outcomes. Int. J. Mol. Sci. 2023, 24, 2288. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Li, M. The application of zebrafish patient-derived xenograft tumor models in the development of antitumor agents. Med. Res. Rev. 2023, 43, 212–236. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Overview of human PIDs. Schematic of lymphopoiesis (A) and myelopoiesis (B), indicating the cells, developmental pathways and activities impacted by the indicated PIDs.
Figure 1.
Overview of human PIDs. Schematic of lymphopoiesis (A) and myelopoiesis (B), indicating the cells, developmental pathways and activities impacted by the indicated PIDs.

Figure 2.
Applications of immunodeficient zebrafish models. Schematic diagrams of how immunodeficient zebrafish are being utilized to study infection and host responses (A), immunity and the gut microbiome (B), tissue repair and regeneration (C) and cancer xenotransplantation (D).
Figure 2.
Applications of immunodeficient zebrafish models. Schematic diagrams of how immunodeficient zebrafish are being utilized to study infection and host responses (A), immunity and the gut microbiome (B), tissue repair and regeneration (C) and cancer xenotransplantation (D).


{kind=link}
{kind=link}
Table 1.
Zebrafish models of human PIDs.
| Human Defect † | Inheritance | OMIM | Zebrafish Model * | Immune Phenotype # | References |
|---|---|---|---|---|---|
| Lymphoid deficiencies | |||||
| T−B− severe combined immunodeficiency (SCID) | |||||
| RAG1 | AR | 179615 | rag1 LOF | ↓ T and B cells | [30,31] |
| RAG2 | AR | 179616 | rag2 hypomorph | ↓ T cells, variable B cell deficiency | [16,32] |
| PRKDC | AR | 615966 | prkdc LOF | ↓ T, B and NK (but not NK-like) cells | [33] |
| AK2 | AR | 103020 | ak2 LOF + KD | ↓ embryonic T cells (and neutrophils) | [34] |
| T−B+ severe combined immunodeficiency (SCID) | |||||
| IL2RG | XL | 308380 | il2rga LOF | ↓ T and NK cells | [26,35] |
| JAK3 | AR | 600173 | jak3 LOF | ↓ T and NK cells, minor defects in mature B cells and neutrophils | [16,36,37] |
| IL7R | AR | 146661 | il7r LOF | ↓ embryonic T cells | [36] |
| Combined immunodeficiencies (CID) | |||||
| ZAP70 | AR | 269840 | zap70 LOF | ↓ thymic and mature kidney T cells | [38] |
| IKZF1 | AD | 603023 | ikzf1 LOF | ↓ embryonic T cells, less severe defect in juveniles and adults | [39] |
| Multi-system immunodeficiencies | |||||
| STAT5B | AR | 245590 | stat5.1 LOF | ↓ T cells throughout lifespan, ↑ T cell activation | [40] |
| EXTL3 | AR | 617425 | extl3 LOF | ↓ embryonic T cells | [41] |
| ARPC1B | AR | 604223 | arpc1b KD | ↓ embryonic T cells | [42] |
| ATM | AR | 607585 | atm LOF | ↓ embryonic T cell and neutrophils, ↓ lymphocytes and ↑ precursors in adults | [43] |
| BCL11B | AD | 617237 | BCL11B GOF TG | ↓ embryonic T cells | [44] |
| FOXN1 | AR | 601705 | foxn1 LOF | ↓ embryonic T cells | [45] |
| CHD7 | AD | 608892 | chd7 LOF + KD | ↓ embryonic T cells | [46] |
| TBX1 | AD | 602054 | tbx1 LOF | athymic | [47] |
| Autoinflammatory disorders | |||||
| NCKAP1L | AR | 618982 | nckap1l LOF | defective neutrophil migration | [48] |
| Myeloid deficiencies | |||||
| Congenital neutropenias | |||||
| CSF3R | AR | 138971 | csf3r LOF | ↓ neutrophils throughout lifespan | [49] |
| SMARCD2 | AR | 601736 | smarcd2 LOF + KD | ↓ embryonic neutrophils | [50] |
| WAS | XL | 300392 | was LOF | defective neutrophil and macrophage migration | [51] |
| HAX1 | AR | 605998 | hax1 KD | ↓ embryonic neutrophils | [52] |
| VPS45 | AR | 615288 | vps45 KD | ↓ embryonic neutrophils | [53] |
| SRP54 | AD | 604857 | srp45 LOF | ↓ embryonic neutrophils | [54] |
| Motility defects | |||||
| ITGB2 | AR | 600065 | itgb2 LOF | defective neutrophil trafficking to inflammatory sites | [55] |
| RAC2 | AD | 608203 | rac2 LOF + KD | defective neutrophil and macrophage mobility | [56,57] |
| CXCR4 | AD | 162643 | cxcr4 GOF TG | ↓ embryonic neutrophils, defective neutrophil mobility and wound recruitment | [58] |
| Respiratory burst defects | |||||
| CYBA | AR | 608508 | cyba LOF | defective neutrophil reverse migration and macrophage wound attraction | [59,60] |
| MPO | AR | 254600 | mpo LOF | sustained hydrogen peroxidase burst | [61,62] |
† Limited to those listed in the 2022 update of human inborn errors of immunity by the IUISEC [4]. * GOF: gain-of-function; KD: knockdown; LOF: loss-of-function; TG: transgenic. # ↓: decreased; ↑: increased.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Basheer, F.; Sertori, R.; Liongue, C.; Ward, A.C. Zebrafish: A Relevant Genetic Model for Human Primary Immunodeficiency (PID) Disorders? Int. J. Mol. Sci. 2023, 24, 6468. https://doi.org/10.3390/ijms24076468
AMA Style
Basheer F, Sertori R, Liongue C, Ward AC. Zebrafish: A Relevant Genetic Model for Human Primary Immunodeficiency (PID) Disorders? International Journal of Molecular Sciences. 2023; 24(7):6468. https://doi.org/10.3390/ijms24076468
Chicago/Turabian StyleBasheer, Faiza, Robert Sertori, Clifford Liongue, and Alister C. Ward. 2023. "Zebrafish: A Relevant Genetic Model for Human Primary Immunodeficiency (PID) Disorders?" International Journal of Molecular Sciences 24, no. 7: 6468. https://doi.org/10.3390/ijms24076468
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.