
Pyrogallol from Spirogyra neglecta Inhibits Proliferation and Promotes Apoptosis in Castration-Resistant Prostate Cancer Cells via Modulating Akt/GSK-3β/β-catenin Signaling Pathway
 , ,
, ,  , , and
, , and 
Abstract
:1. Introduction
2. Results
2.1. Extraction and Phenolic Compound Identification of S. neglecta Ethyl Acetate Fraction (SnEA)
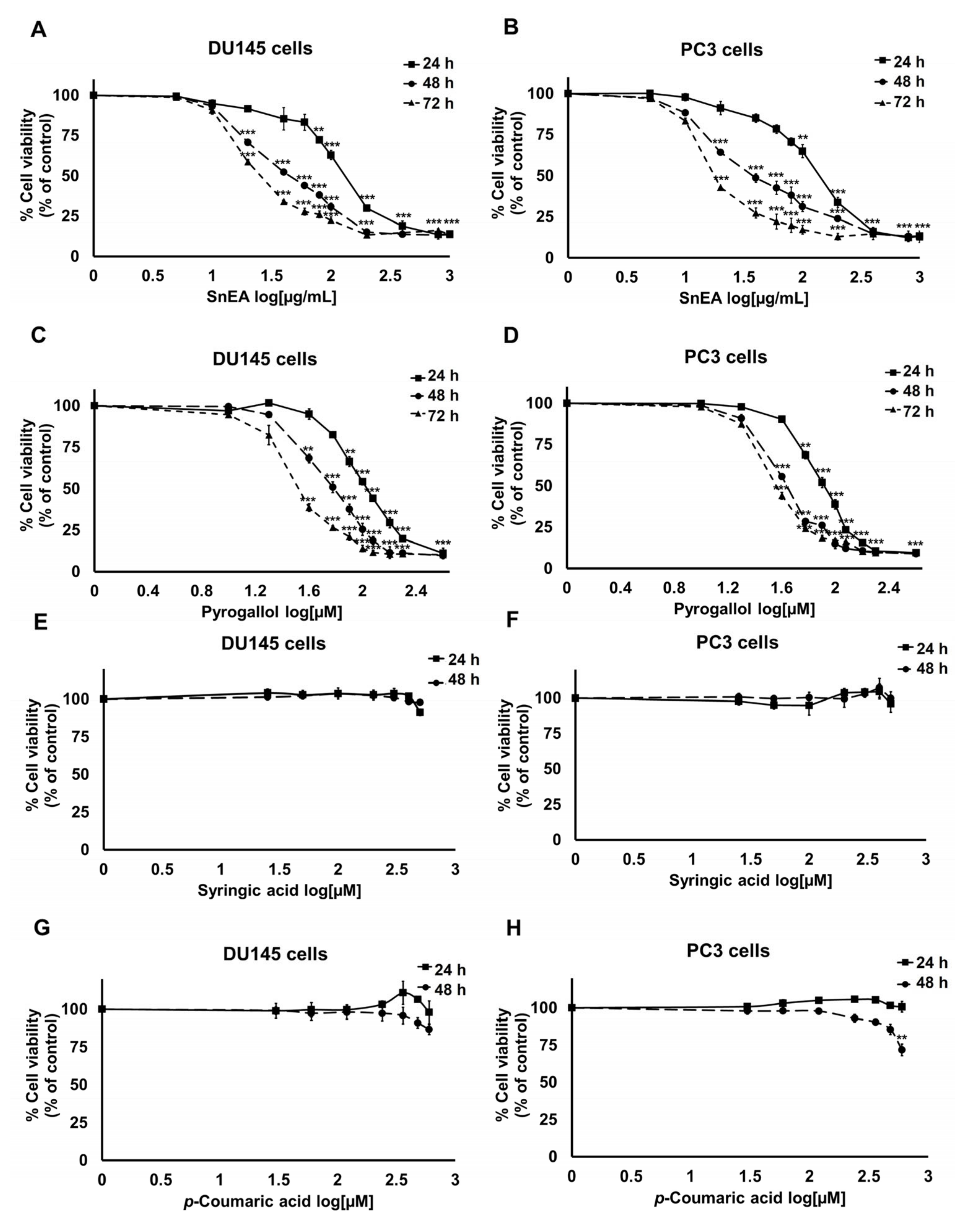
2.2. Effect of SnEA and Its Major Phenolic Compounds on DU145 and PC3 Cells Viability
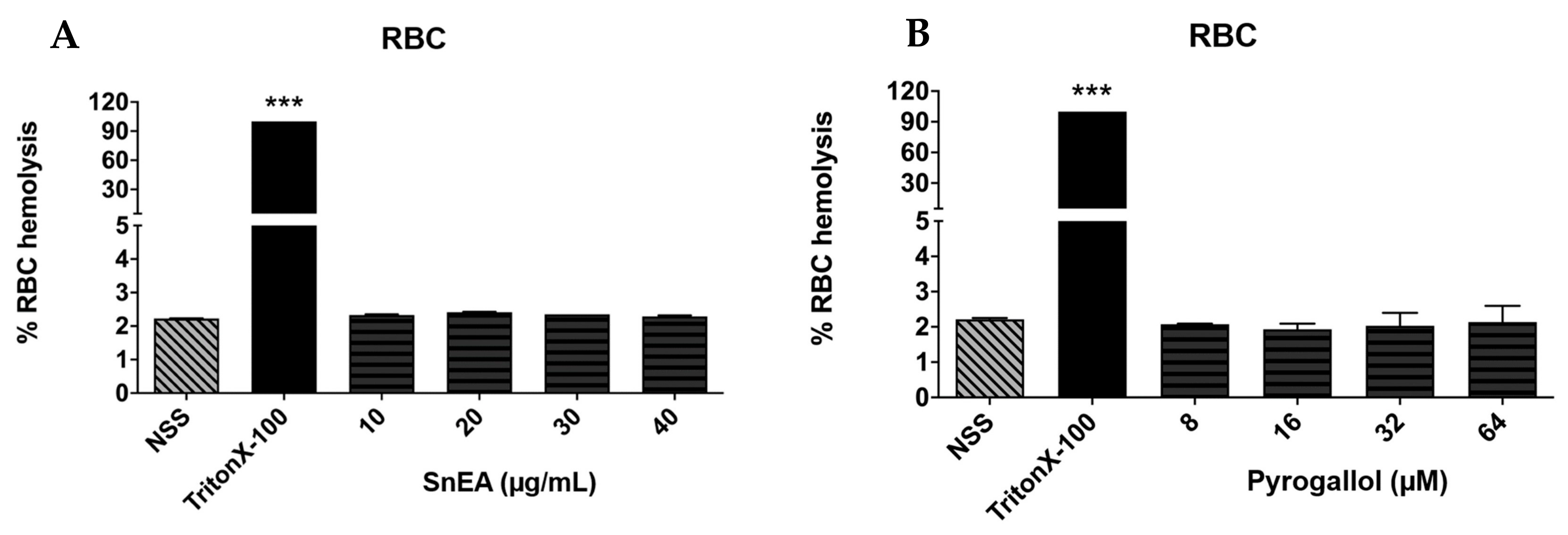
2.3. Evaluation of SnEA and Pyrogallol-Induced Toxicity in Normal Cells
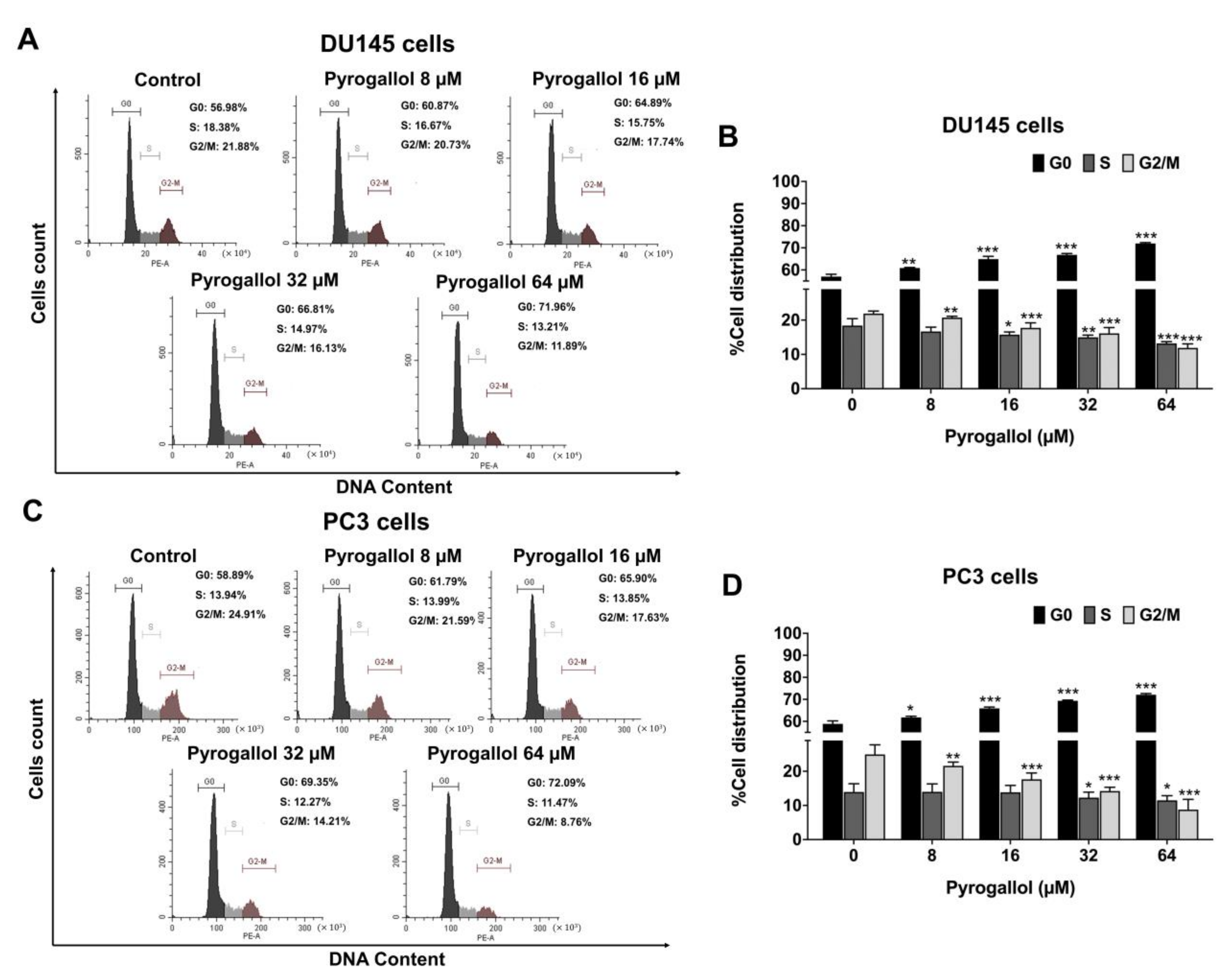
2.4. Effects of SnEA and Pyrogallol on Cell Cycle Arrest in CRPC Cells
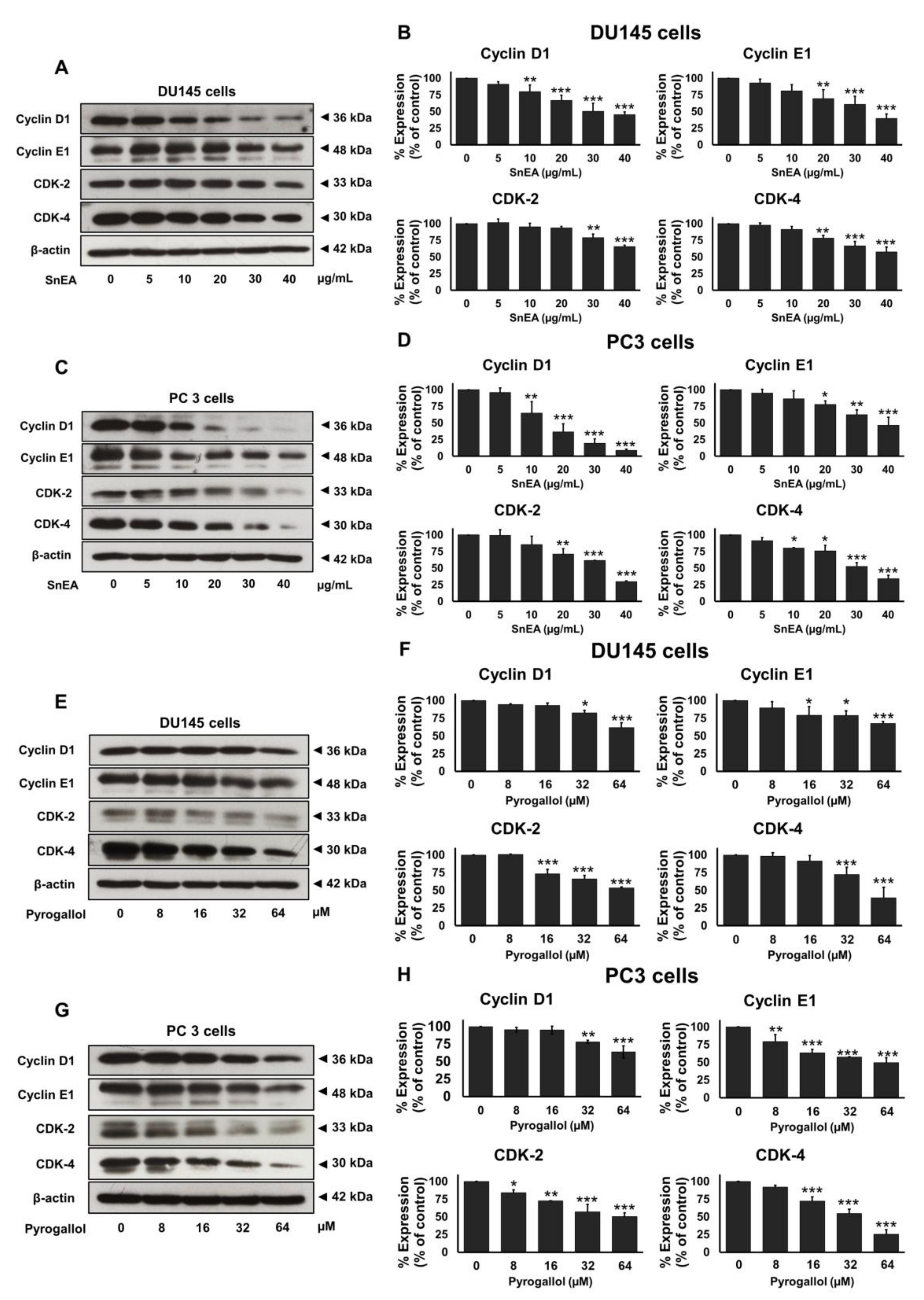
2.5. Effects of SnEA and Pyrogallol on Cell Cycle-Regulated Proteins’ Expression in CRPC Cells
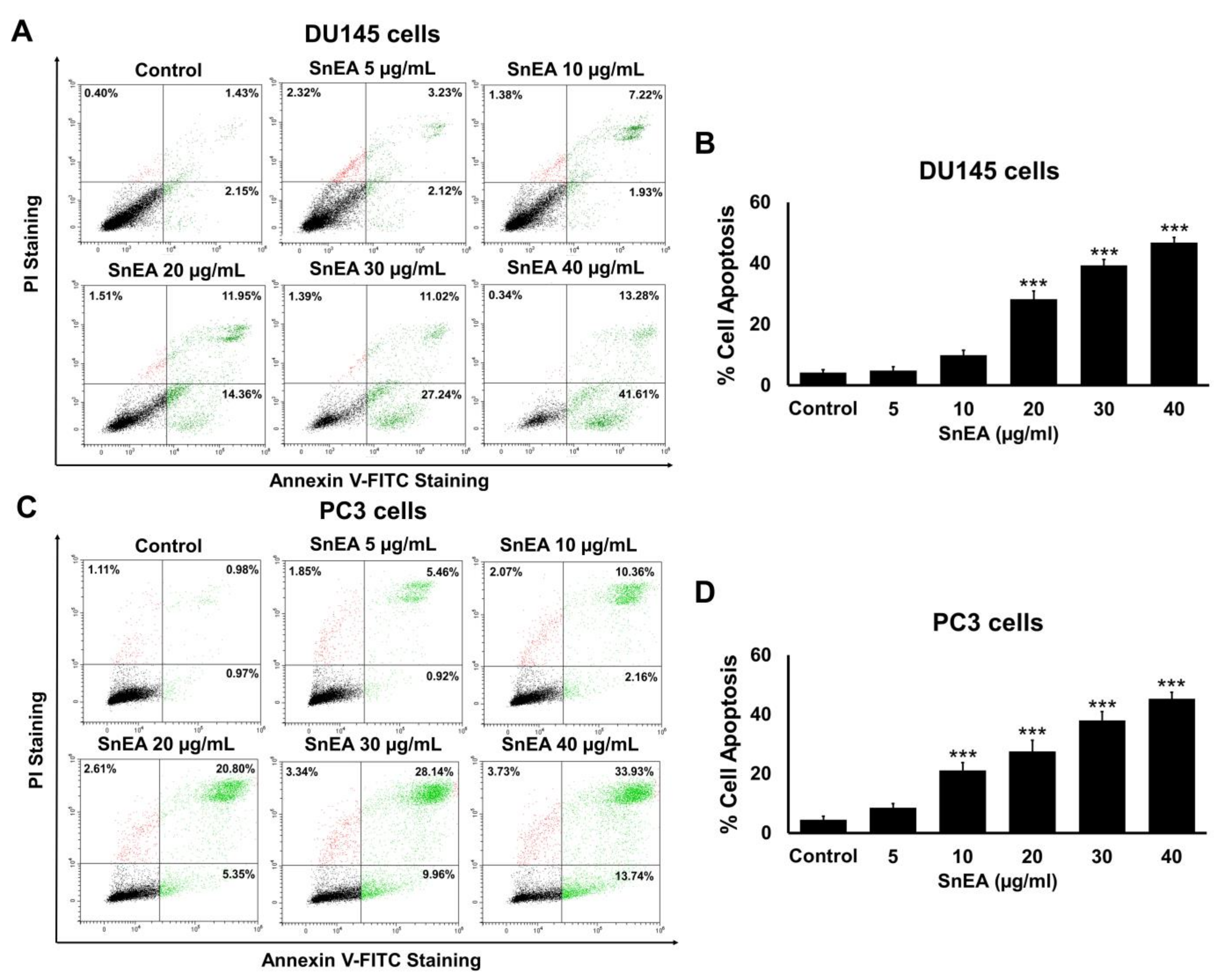
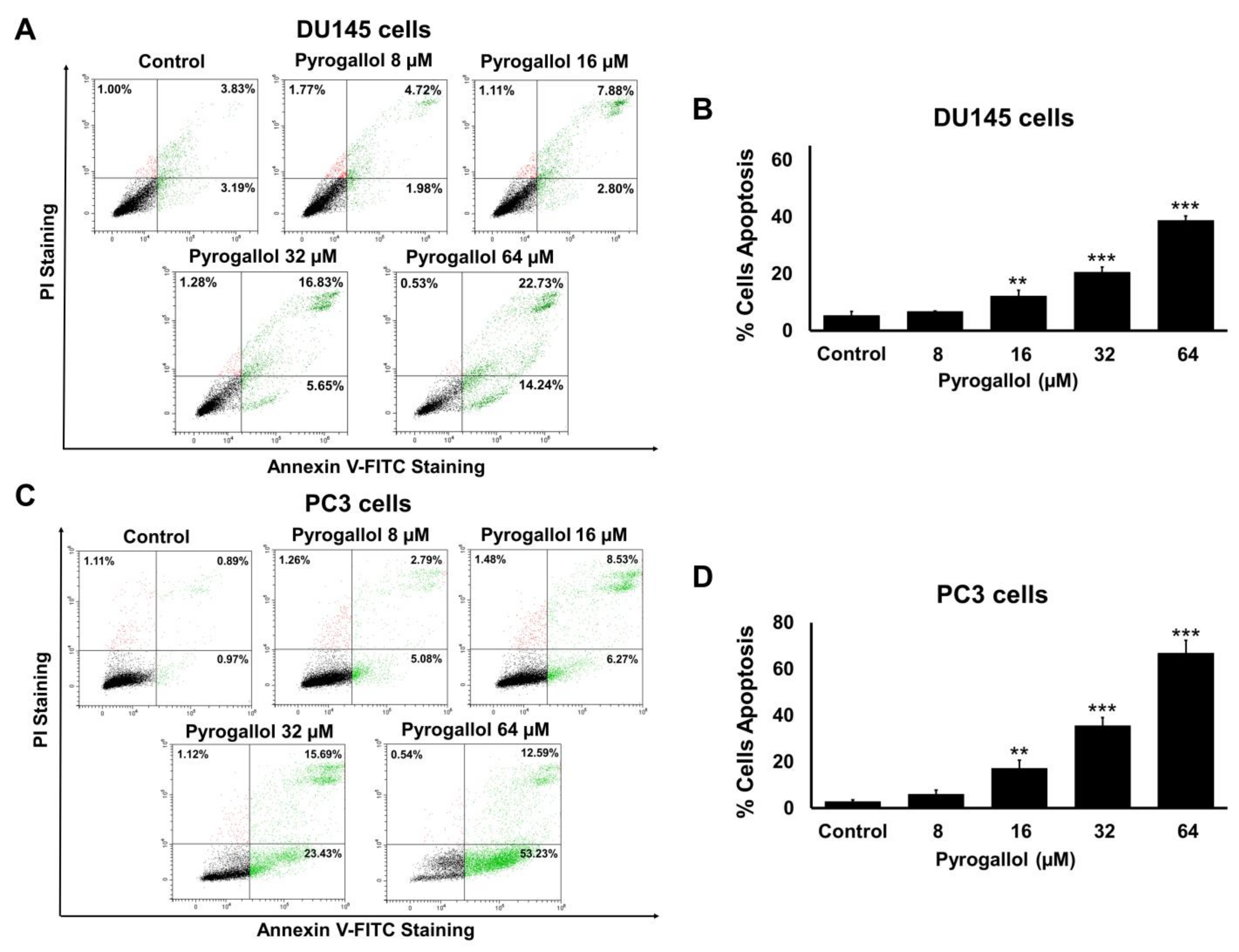
2.6. Effects of SnEA and Pyrogallol on CRPC Cells Apoptosis
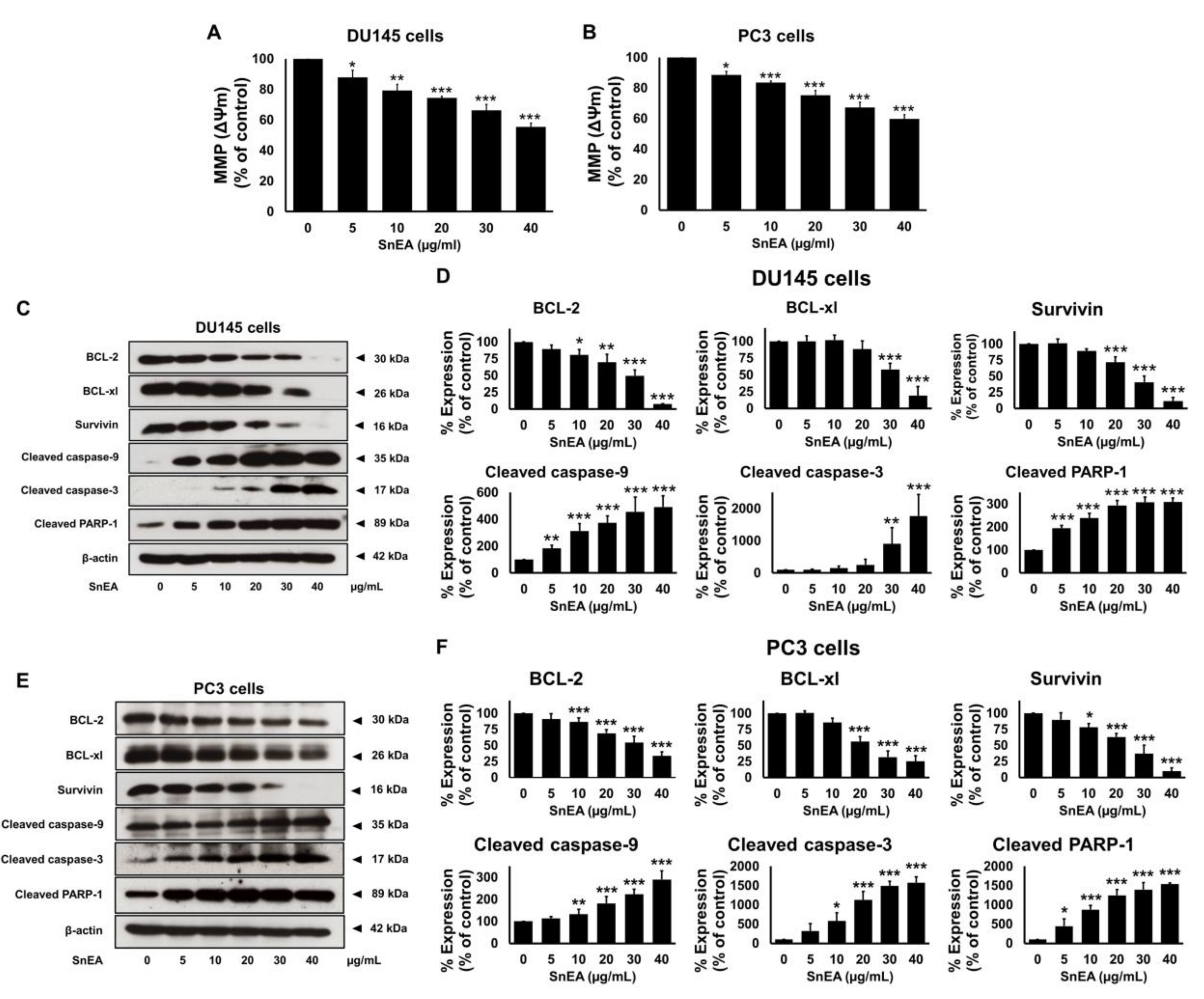
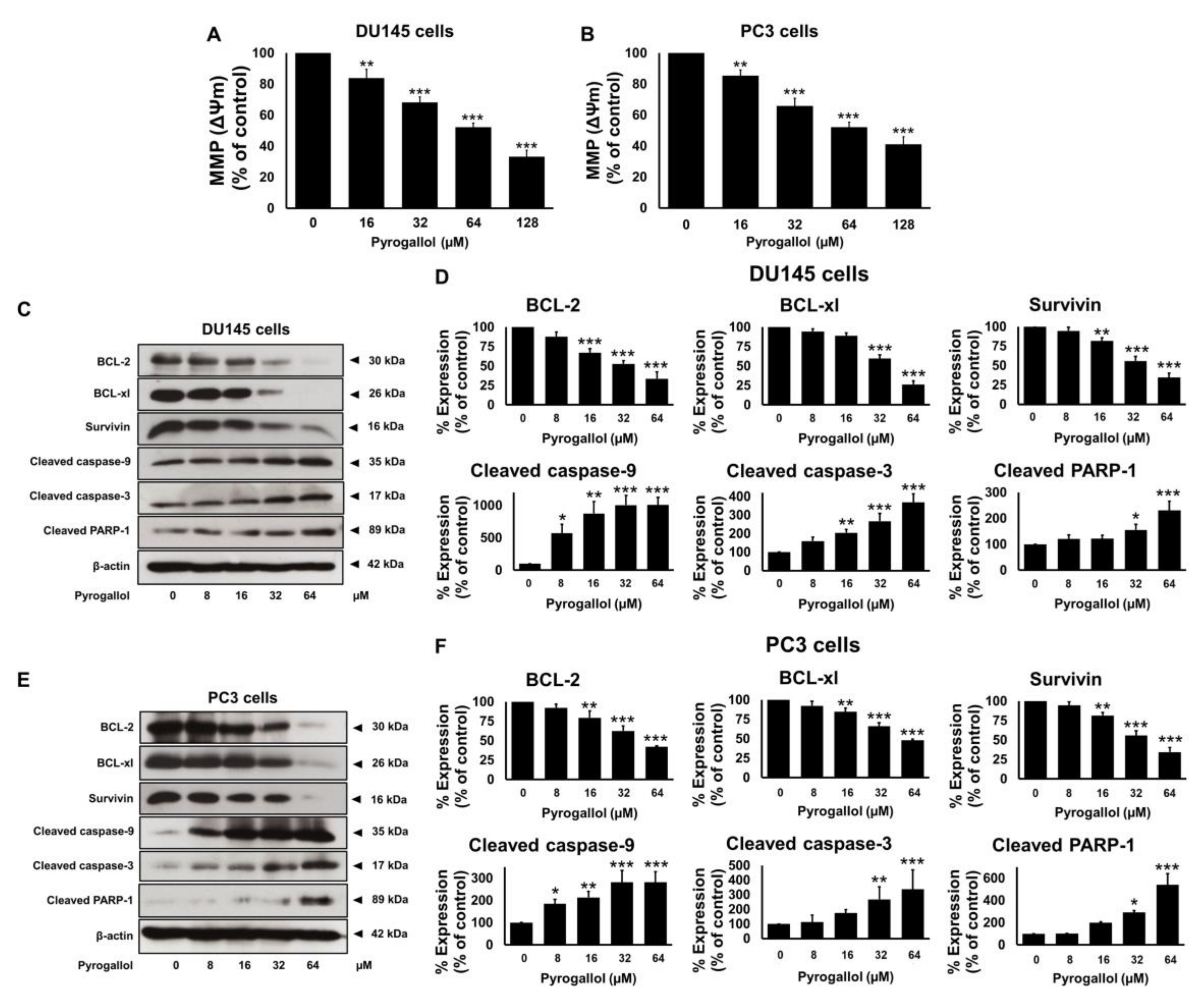
2.7. Effects of SnEA and Pyrogallol on Pro- and Anti-Apoptosis Protein Expression in CRPC Cells
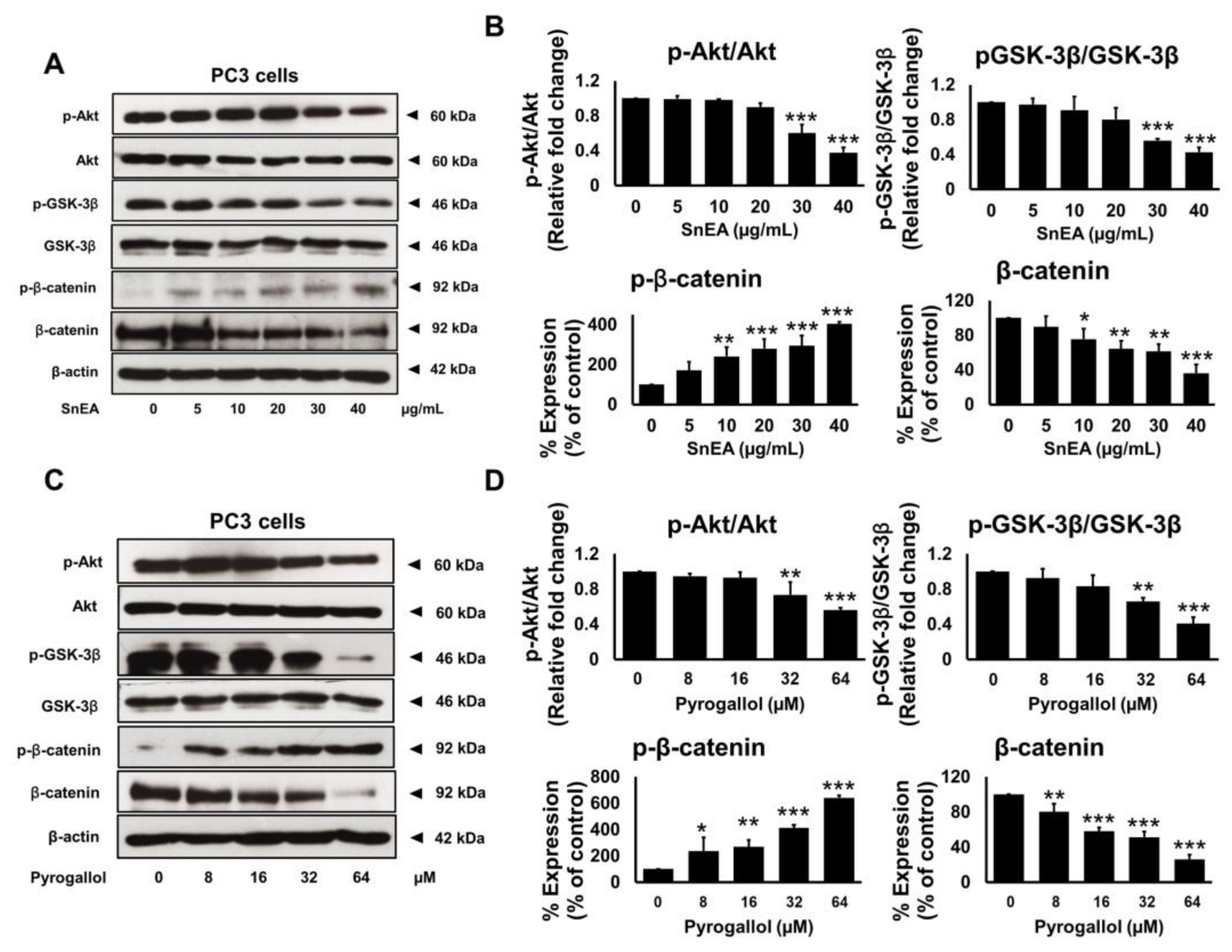
2.8. Effects of SnEA and Pyrogallol on the Akt/GSK-3β/β-Catenin Signaling Pathway in PC3 Cells
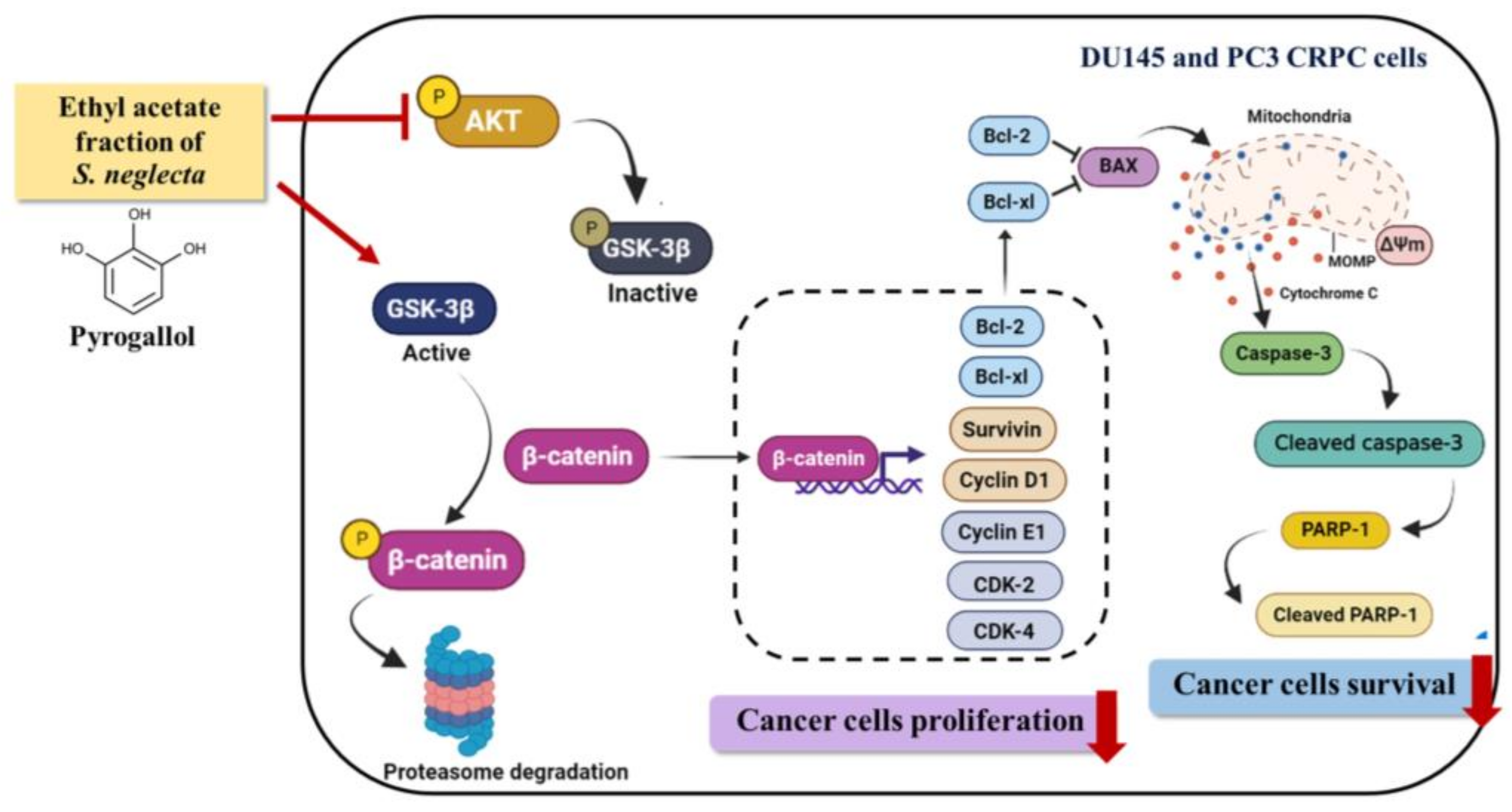
3. Discussion
4. Materials and Methods
4.1. Chemical and Reagents
4.2. Herb Materials
4.3. Preparation of Spirogyra Neglecta Extracts and Fractions
4.4. Total Phenolic Content
4.5. Phenolics Content Determination Using HPLC
4.6. Cell Cultures
4.7. The Cell Viability Assay (SRB Assay)
4.8. Red Blood Cells (RBCs) Hemolysis
4.9. Mitochondrial Membrane Potential Assay
4.10. Cell Cycle Assay
4.11. Apoptosis Assay
4.12. Western Blot Analysis
4.13. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Komura, K.; Sweeney, C.J.; Inamoto, T.; Ibuki, N.; Azuma, H.; Kantoff, P.W. Current treatment strategies for advanced prostate cancer. Int. J. Urol. 2018, 25, 220–231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Lu, B.; He, M.; Wang, Y.; Wang, Z.; Du, L. Prostate cancer incidence and mortality: Global status and temporal trends in 89 countries from 2000 to 2019. Front. Public Health 2022, 10, 811044. [Google Scholar] [CrossRef] [PubMed]
- Chandrasekar, T.; Yang, J.C.; Gao, A.C.; Evans, C.P. Mechanisms of resistance in castration-resistant prostate cancer (CRPC). Transl. Androl. Urol. 2015, 4, 365. [Google Scholar] [PubMed]
- Kirby, M.; Hirst, C.; Crawford, E. Characterising the castration-resistant prostate cancer population: A systematic review. Int. J. Clin. Pract. 2011, 65, 1180–1192. [Google Scholar] [CrossRef] [PubMed]
- Lin, T.-T.; Chen, Y.-H.; Wu, Y.-P.; Chen, S.-Z.; Li, X.-D.; Lin, Y.-Z.; Chen, S.-H.; Zheng, Q.-S.; Wei, Y.; Xu, N. Risk factors for progression to castration-resistant prostate cancer in metastatic prostate cancer patients. J. Cancer 2019, 10, 5608. [Google Scholar] [CrossRef]
- Vellky, J.E.; Ricke, W.A. Development and prevalence of castration-resistant prostate cancer subtypes. Neoplasia 2020, 22, 566–575. [Google Scholar] [CrossRef]
- Saad, F.; Hotte, S.J. Guidelines for the management of castrate-resistant prostate cancer. Can. Urol. Assoc. J 2010, 4, 380. [Google Scholar] [CrossRef] [Green Version]
- Bitting, R.L.; Armstrong, A.J. Targeting the PI3K/Akt/mTOR pathway in castration-resistant prostate cancer. Endocr.-Relat. Cancer 2013, 20, R83–R99. [Google Scholar] [CrossRef] [Green Version]
- Taylor, B.S.; Schultz, N.; Hieronymus, H.; Gopalan, A.; Xiao, Y.; Carver, B.S.; Arora, V.K.; Kaushik, P.; Cerami, E.; Reva, B. Integrative genomic profiling of human prostate cancer. Cancer Cell 2010, 18, 11–22. [Google Scholar] [CrossRef] [Green Version]
- Shorning, B.Y.; Dass, M.S.; Smalley, M.J.; Pearson, H.B. The PI3K-AKT-mTOR pathway and prostate cancer: At the crossroads of AR, MAPK, and WNT signaling. Int. J. Mol. Sci. 2020, 21, 4507. [Google Scholar] [CrossRef]
- Dong, J.; Xu, X.; Zhang, Q.; Yuan, Z.; Tan, B. The PI3K/AKT pathway promotes fracture healing through its crosstalk with Wnt/β-catenin. Exp. Cell Res. 2020, 394, 112137. [Google Scholar] [CrossRef]
- He, Y.; Xu, W.; Xiao, Y.-T.; Huang, H.; Gu, D.; Ren, S. Targeting signaling pathways in prostate cancer: Mechanisms and clinical trials. Signal Transduct. Target. Ther. 2022, 7, 198. [Google Scholar] [CrossRef]
- Satari, A.; Ghasemi, S.; Habtemariam, S.; Asgharian, S.; Lorigooini, Z. Rutin: A flavonoid as an effective sensitizer for anticancer therapy; insights into multifaceted mechanisms and applicability for combination therapy. Evid. Based Complement. Altern. Med. 2021, 2021, 9913179. [Google Scholar] [CrossRef]
- Maurya, S.K.; Fatma, H.; Maurya, A.K.; Mishra, N.; Siddique, H.R. Role of lupeol in chemosensitizing therapy-resistant prostate cancer cells by targeting MYC, β-catenin and c-FLIP: In silico and in vitro studies. Silico Pharm. 2022, 10, 16. [Google Scholar] [CrossRef]
- Yap, W.; Zaiden, N.; Luk, S.; Lee, D.; Ling, M.; Wong, Y.; Yap, Y. In vivo evidence of γ-tocotrienol as a chemosensitizer in the treatment of hormone-refractory prostate cancer. Pharmacology 2010, 85, 248–258. [Google Scholar] [CrossRef]
- Belyagoubi, L.; Belyagoubi-Benhammou, N.; Atik-Bekkara, F.; Abdelouahid, D.E. Influence of harvest season and different polarity solvents on biological activities, phenolic compounds and lipid-soluble pigment contents of Spirogyra sp. from Algeria. Adv. Trad. Med. 2022, 22, 359–369. [Google Scholar] [CrossRef]
- Kumar, J.; Dhar, P.; Tayade, A.B.; Gupta, D.; Chaurasia, O.P.; Upreti, D.K.; Toppo, K.; Arora, R.; Suseela, M.; Srivastava, R.B. Chemical composition and biological activities of trans-Himalayan alga Spirogyra porticalis (Muell.) Cleve. PLoS ONE 2015, 10, e0118255. [Google Scholar] [CrossRef]
- Mesbahzadeh, B.; Rajaei, S.A.; Tarahomi, P.; Seyedinia, S.A.; Rahmani, M.; Rezamohamadi, F.; Kakar, M.A.; Moradi-Kor, N. Beneficial effects of Spirogyra Neglecta Extract on antioxidant and anti-inflammatory factors in streptozotocin-induced diabetic rats. Biomol. Concepts 2018, 9, 184–189. [Google Scholar] [CrossRef] [Green Version]
- Junthip, R.; Amornlerdpison, D.; Chimsook, T. Phytochemical screening, antioxidant activity and total phenolic content of Spirogyra spp. In Advanced Materials Research; Trans Tech Publications Ltd.: Chiang Mai, Thailand, 2013; pp. 693–697. [Google Scholar]
- Jerez-Martel, I.; García-Poza, S.; Rodríguez-Martel, G.; Rico, M.; Afonso-Olivares, C.; Gómez-Pinchetti, J.L. Phenolic profile and antioxidant activity of crude extracts from microalgae and cyanobacteria strains. J. Food Qual. 2017, 2017, 2924508. [Google Scholar] [CrossRef] [Green Version]
- Shaaban, A.; Mansour, H. Comparative Phytochemical Evaluation Between Two of The Most Common Egyptian Fresh Water Green Algae. Egypt. J. Bot. 2012, 52, 455–463. [Google Scholar]
- Thumvijit, T.; Thuschana, W.; Amornlerdpison, D.; Peerapornpisal, Y.; Wongpoomchai, R. Evaluation of hepatic antioxidant capacities of Spirogyra neglecta (Hassall) Kützing in rats. Interdiscip. Toxicol. 2013, 6, 152–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ontawong, A.; Saowakon, N.; Vivithanaporn, P.; Pongchaidecha, A.; Lailerd, N.; Amornlerdpison, D.; Lungkaphin, A.; Srimaroeng, C. Antioxidant and renoprotective effects of Spirogyra neglecta (Hassall) Kützing extract in experimental type 2 diabetic rats. Biomed Res. Int. 2013, 2013, 820786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adrien, R.; Gitu, L.M.; Oyaro, N. Mineral composition, Antioxidants and Antimicrobial activities of freshwater algae (spirogyra genus). In Proceedings of the Jomo Kenyatta University of Agriculture and Technology (JKUAT) Conference, Juja, Kenya, 13–14 November 2014; Jomo Kenyatta University of Agriculture and Technology: Nairobi, Kenya, 2014. [Google Scholar]
- Thumvijit, T.; Taya, S.; Punvittayagul, C.; Peerapornpisal, Y.; Wongpoomchai, R. Cancer chemopreventive effect of Spirogyra neglecta (Hassall) Kützing on diethylnitrosamine-induced hepatocarcinogenesis in rats. Asian Pac. J. Cancer Prev. 2014, 15, 1611–1616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taya, S.; Thumvijit, T.; Chewonarin, T.; Punvittayagul, C.; Wongpoomchai, R. Effect of Spirogyra neglecta on the early stages of 1, 2-dimethylhydrazine-induced colon carcinogenesis in rats. Eur. J. Cancer Prev. 2018, 27, 110–117. [Google Scholar] [CrossRef]
- Arjsri, P.; Phetcharaburanin, J.; Suksawat, M.; Mapoung, S.; Subkamkaew, C.; Semmarath, W.; Yodkeeree, S.; Limtrakul, P. Spirogyra neglecta (Hassall) Kützing attenuates metastasis of castration-resistant human prostate cancer via the blockage of AKT signaling pathway. S. Afr. J. Bot 2021, 139, 26–37. [Google Scholar] [CrossRef]
- Kang, M.; Lee, K.H.; Lee, H.S.; Park, Y.H.; Jeong, C.W.; Ku, J.H.; Kim, H.H.; Kwak, C. PDLIM2 suppression efficiently reduces tumor growth and invasiveness of human castration-resistant prostate cancer-like cells. Prostate 2016, 76, 273–285. [Google Scholar] [CrossRef]
- Wang, Q.; He, W.-Y.; Zeng, Y.-Z.; Hossain, A.; Gou, X. Inhibiting autophagy overcomes docetaxel resistance in castration-resistant prostate cancer cells. Int. J. Nephrol. 2018, 50, 675–686. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.H.; Choo, C.; Watawana, M.I.; Jayawardena, N.; Waisundara, V.Y. An appraisal of eighteen commonly consumed edible plants as functional food based on their antioxidant and starch hydrolase inhibitory activities. J. Sci. Food Agric. 2015, 95, 2956–2964. [Google Scholar] [CrossRef]
- Noreen, H.; Semmar, N.; Farman, M.; McCullagh, J.S. Measurement of total phenolic content and antioxidant activity of aerial parts of medicinal plant Coronopus didymus. Asian Pac. J. Trop. Med. 2017, 10, 792–801. [Google Scholar] [CrossRef]
- Morin, P.J. β-catenin signaling and cancer. BioEssays 1999, 21, 1021–1030. [Google Scholar] [CrossRef]
- Sharma, M.; Chuang, W.W.; Sun, Z. Phosphatidylinositol 3-kinase/Akt stimulates androgen pathway through GSK3β inhibition and nuclear β-catenin accumulation. J. Biol. Chem. 2002, 277, 30935–30941. [Google Scholar] [CrossRef] [Green Version]
- Aly, M.; Leval, A.; Schain, F.; Liwing, J.; Lawson, J.; Vágó, E.; Nordström, T.; Andersson, T.M.-L.; Sjöland, E.; Wang, C. Survival in patients diagnosed with castration-resistant prostate cancer: A population-based observational study in Sweden. Scand. J. Urol. 2020, 54, 115–121. [Google Scholar] [CrossRef] [Green Version]
- Madan, R.A.; Gulley, J.L.; Schlom, J.; Steinberg, S.M.; Liewehr, D.J.; Dahut, W.L.; Arlen, P.M. Analysis of overall survival in patients with nonmetastatic castration-resistant prostate cancer treated with vaccine, nilutamide, and combination therapy. Clin. Cancer Res. 2008, 14, 4526–4531. [Google Scholar] [CrossRef] [Green Version]
- Duangjai, A.; Limpeanchob, N.; Trisat, K.; Amornlerdpison, D. Spirogyra neglecta inhibits the absorption and synthesis of cholesterol in vitro. Integr. Med. Res 2016, 5, 301–308. [Google Scholar] [CrossRef] [Green Version]
- Yongkhamcha, B.; Buddhakala, N. Phytochemical Compositions, Nutritional Contents, Cytotoxicity and Anti-Inflammatory Activity of Different Extracts from Spirogyra neglecta (Hassall) Kützing. Trends Sci. 2023, 20, 6528. [Google Scholar] [CrossRef]
- Tai, S.; Sun, Y.; Squires, J.M.; Zhang, H.; Oh, W.K.; Liang, C.Z.; Huang, J. PC3 is a cell line characteristic of prostatic small cell carcinoma. Prostate 2011, 71, 1668–1679. [Google Scholar] [CrossRef] [Green Version]
- Guo, J.; Ma, K.; Xia, H.M.; Chen, Q.K.; Li, L.; Deng, J.; Sheng, J.; Hong, Y.Y.; Hu, J.P. Androgen receptor reverts dexamethasone-induced inhibition of prostate cancer cell proliferation and migration. Mol. Med. Rep. 2018, 17, 5887–5893. [Google Scholar] [CrossRef] [Green Version]
- Saeed, B.; Zhang, H.; Ng, S.C. Apoptotic program is initiated but not completed in LNCaP cells in response to growth in charcoal-stripped media. Prostate 1997, 31, 145–152. [Google Scholar] [CrossRef]
- Nicolis, E.; Lampronti, I.; Dechecchi, M.C.; Borgatti, M.; Tamanini, A.; Bianchi, N.; Bezzerri, V.; Mancini, I.; Giri, M.G.; Rizzotti, P. Pyrogallol, an active compound from the medicinal plant Emblica officinalis, regulates expression of pro-inflammatory genes in bronchial epithelial cells. Int. Immunopharmacol. 2008, 8, 1672–1680. [Google Scholar] [CrossRef]
- Han, Y.H.; Kim, S.Z.; Kim, S.H.; Park, W.H. Pyrogallol inhibits the growth of lung cancer Calu-6 cells via caspase-dependent apoptosis. Chem.-Biol. Interact. 2009, 177, 107–114. [Google Scholar] [CrossRef]
- Park, W.H.; Park, M.N.; Han, Y.H.; Kim, S.W. Pyrogallol inhibits the growth of gastric cancer SNU-484 cells via induction of apoptosis. Int. J. Mol. Med. 2008, 22, 263–268. [Google Scholar] [PubMed]
- Mitsuhashi, S.; Saito, A.; Nakajima, N.; Shima, H.; Ubukata, M. Pyrogallol structure in polyphenols is involved in apoptosis-induction on HEK293T and K562 cells. Molecules 2008, 13, 2998–3006. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.T.; Lampronti, I.; Martello, D.; Bianchi, N.; Jabbar, S.; Choudhuri, M.S.K.; Datta, B.K.; Gambari, R. Identification of pyrogallol as an antiproliferative compound present in extracts from the medicinal plant Emblica officinalis: Effects on in vitro cell growth of human tumor cell lines. Int. J. Oncol. 2002, 21, 187–192. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.W.; Han, Y.W.; Lee, S.T.; Jeong, H.J.; Kim, S.H.; Kim, I.H.; Lee, S.O.; Kim, D.G.; Kim, S.H.; Kim, S.Z. A superoxide anion generator, pyrogallol, inhibits the growth of HeLa cells via cell cycle arrest and apoptosis. Mol. Carcinog. 2008, 47, 114–125. [Google Scholar] [CrossRef]
- Liu, S.; Zhao, B.; Shi, D.; Xin, W. Auto-oxidation mechanism of pyrogallol and its free radical effects studied by means of ESR, UV and NMR. Chem. Res. Chin. Univ. 1999, 15, 146–151. [Google Scholar]
- Liu, B.; Zhou, Z.; Zhou, W.; Liu, J.; Zhang, Q.; Xia, J.; Liu, J.; Chen, N.; Li, M.; Zhu, R. Resveratrol inhibits proliferation in human colorectal carcinoma cells by inducing G1/S-phase cell cycle arrest and apoptosis through caspase/cyclin-CDK pathways. Mol. Med. Rep. 2014, 10, 1697–1702. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Martínez, C.; Lallena, M.J.; Sanfeliciano, S.G.; de Dios, A. Cyclin dependent kinase (CDK) inhibitors as anticancer drugs: Recent advances (2015–2019). Bioorg. Med. Chem. Lett. 2019, 29, 126637. [Google Scholar] [CrossRef]
- Otto, T.; Sicinski, P. Cell cycle proteins as promising targets in cancer therapy. Nat. Rev. Cancer 2017, 17, 93–115. [Google Scholar] [CrossRef] [Green Version]
- Vaux, D.L.; Korsmeyer, S.J. Cell death in development. Cell 1999, 96, 245–254. [Google Scholar] [CrossRef] [Green Version]
- Ly, J.D.; Grubb, D.R.; Lawen, A. The mitochondrial membrane potential (Δψ m) in apoptosis; an update. Apoptosis Int. J. Program. Cell Death 2003, 8, 115–128. [Google Scholar] [CrossRef]
- Gottlieb, E.; Armour, S.; Harris, M.; Thompson, C. Mitochondrial membrane potential regulates matrix configuration and cytochrome c release during apoptosis. Cell Death Differ. 2003, 10, 709–717. [Google Scholar] [CrossRef] [Green Version]
- Giridharan, M.; Rupani, V.; Banerjee, S. Signaling pathways and targeted therapies for stem cells in prostate cancer. ACS Pharmacol. Transl. Sci. 2022, 5, 193–206. [Google Scholar] [CrossRef]
- Edlind, M.P.; Hsieh, A.C. PI3K-AKT-mTOR signaling in prostate cancer progression and androgen deprivation therapy resistance. Asian J. Androl. 2014, 16, 378. [Google Scholar]
- Morgan, T.M.; Koreckij, T.D.; Corey, E. Targeted therapy for advanced prostate cancer: Inhibition of the PI3K/Akt/mTOR pathway. Curr. Cancer Drug Targets 2009, 9, 237–249. [Google Scholar] [CrossRef] [Green Version]
- Schweizer, M.T.; Yu, E.Y. Persistent androgen receptor addiction in castration-resistant prostate cancer. J. Hematol. Oncol. 2015, 8, 128. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.H.; Johnson, D.; Luong, R.; Sun, Z. Crosstalking between androgen and PI3K/AKT signaling pathways in prostate cancer cells. J. Biol. Chem. 2015, 290, 2759–2768. [Google Scholar] [CrossRef] [Green Version]
- Guo, H.; Luo, H.; Yuan, H.; Xia, Y.; Shu, P.; Huang, X.; Lu, Y.; Liu, X.; Keller, E.T.; Sun, D. Litchi seed extracts diminish prostate cancer progression via induction of apoptosis and attenuation of EMT through Akt/GSK-3β signaling. Sci. Rep. 2017, 7, 41656. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Qin, X.; Li, P.; Zhang, H.; Lin, T.; Miao, Z.; Ma, S. Isobavachalcone isolated from Psoralea corylifolia inhibits cell proliferation and induces apoptosis via inhibiting the AKT/GSK-3β/β-catenin pathway in colorectal cancer cells. Drug Des. Devel. Ther. 2019, 13, 1449–1460. [Google Scholar] [CrossRef] [Green Version]
- Tang, Z.; Yang, G.; Wang, X.; Chen, F.; Liao, Z.; Zhang, Z.; Liu, Z.; Zeng, W.; Fang, M.; Wang, W. AKT/GSK-3β/β-catenin signaling pathway participates in erythropoietin-promoted glioma proliferation. J. Neuro-Oncol. 2020, 149, 231–242. [Google Scholar] [CrossRef]
- Wang, G.; Feng, C.-C.; Chu, S.-J.; Zhang, R.; Lu, Y.-M.; Zhu, J.-S.; Zhang, J. Toosendanin inhibits growth and induces apoptosis in colorectal cancer cells through suppression of AKT/GSK-3β/β-catenin pathway. Int. J. Oncol. 2015, 47, 1767–1774. [Google Scholar] [CrossRef] [Green Version]
- Jia, L.; Lv, D.; Zhang, S.; Wang, Z.; Zhou, B. Astragaloside IV inhibits the progression of non-small cell lung cancer through the Akt/GSK-3β/β-catenin pathway. Oncol. Res. 2019, 27, 503–508. [Google Scholar] [CrossRef] [PubMed]
- Choudhury, A.D.; Higano, C.S.; de Bono, J.S.; Cook, N.; Rathkopf, D.E.; Wisinski, K.B.; Martin-Liberal, J.; Linch, M.; Heath, E.I.; Baird, R.D.; et al. A Phase I Study Investigating AZD8186, a Potent and Selective Inhibitor of PI3Kβ/δ, in Patients with Advanced Solid Tumors. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2022, 28, 2257–2269. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.-H.; Tseng, J.-C.; Lin, C.-Y.; Kuo, Y.-Y.; Wang, B.-J.; Kao, Y.-H.; Muller, C.J.; Joubert, E.; Chuu, C.-P. Rooibos suppresses proliferation of castration-resistant prostate cancer cells via inhibition of Akt signaling. Phytomedicine 2019, 64, 153068. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Yue, Y.; Gu, Y.; Wang, Q.; Liu, T.; Li, L.; Wang, X.; Chang, L.S.; He, D.; Wu, K. MUC15 loss facilitates epithelial-mesenchymal transition and cancer stemness for prostate cancer metastasis through GSK3β/β-catenin signaling. Cell. Signal. 2021, 84, 110015. [Google Scholar] [CrossRef]
- Mapoung, S.; Arjsri, P.; Thippraphan, P.; Semmarath, W.; Yodkeeree, S.; Chiewchanvit, S.; Piyamongkol, W.; Limtrakul, P. Photochemoprotective effects of Spirulina platensis extract against UVB irradiated human skin fibroblasts. S. Afr. J. Bot 2020, 130, 198–207. [Google Scholar] [CrossRef]
- Mapoung, S.; Umsumarng, S.; Semmarath, W.; Arjsri, P.; Srisawad, K.; Thippraphan, P.; Yodkeeree, S.; Dejkriengkraikul, P. Photoprotective effects of a hyperoside-enriched fraction prepared from houttuynia cordata thunb. On ultraviolet B-induced skin aging in human fibroblasts through the MAPK signaling pathway. Plants 2021, 10, 2628. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Spirogyra neglecta Extracts | Total Phenolic Content (mg GA/g Extract) |
|---|---|
| SnEE | 245.73 ± 6.58 |
| SnDM | 37.68 ± 2.52 |
| SnEA | 735.42 ± 4.55 *** |
| Sn-H2O | 27.33 ± 0.99 |
| Ethyl Acetate Fraction of S. neglecta (SnEA) | mg/g Extract (Mean ± S.D.) |
|---|---|
| Syringic acid | 122.28 ± 11.72 |
| Pyrogallol | 107.53 ± 4.48 |
| p-coumaric acid | 93.22 ± 8.23 |
| Catechin | 10.11 ± 2.13 |
| Gallic acid | 9.96 ± 2.80 |
| Caffeic acid | ND |
| Chlorogenic acid | ND |
| Ellagic acid | ND |
| Ferulic acid | ND |
| Protocatechuic acid | ND |
| Vanillic acid | ND |
| IC50 of SnEA (µg/mL) at 48 h | IC50 of Pyrogallol (µM) at 48 h | |
|---|---|---|
| DU145 cells | 47.94 ± 0.70 | 58.89 ± 3.06 |
| PC3 cells | 42.97 ± 4.24 | 45.79 ± 1.82 |
| HDF cells | 122.63 ± 2.25 | 201.68 ± 28.72 |
| Selectivity Index (SI) * of SnEA and Pyrogallol Compounds | ||
| HDF/DU145 | 2.55 | 3.42 |
| HDF/PC3 | 2.85 | 4.40 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arjsri, P.; Mapoung, S.; Semmarath, W.; Srisawad, K.; Tuntiwechapikul, W.; Yodkeeree, S.; Dejkriengkraikul, P. Pyrogallol from Spirogyra neglecta Inhibits Proliferation and Promotes Apoptosis in Castration-Resistant Prostate Cancer Cells via Modulating Akt/GSK-3β/β-catenin Signaling Pathway. Int. J. Mol. Sci. 2023, 24, 6452. https://doi.org/10.3390/ijms24076452
Arjsri P, Mapoung S, Semmarath W, Srisawad K, Tuntiwechapikul W, Yodkeeree S, Dejkriengkraikul P. Pyrogallol from Spirogyra neglecta Inhibits Proliferation and Promotes Apoptosis in Castration-Resistant Prostate Cancer Cells via Modulating Akt/GSK-3β/β-catenin Signaling Pathway. International Journal of Molecular Sciences. 2023; 24(7):6452. https://doi.org/10.3390/ijms24076452
Chicago/Turabian StyleArjsri, Punnida, Sariya Mapoung, Warathit Semmarath, Kamonwan Srisawad, Wirote Tuntiwechapikul, Supachai Yodkeeree, and Pornngarm Dejkriengkraikul. 2023. "Pyrogallol from Spirogyra neglecta Inhibits Proliferation and Promotes Apoptosis in Castration-Resistant Prostate Cancer Cells via Modulating Akt/GSK-3β/β-catenin Signaling Pathway" International Journal of Molecular Sciences 24, no. 7: 6452. https://doi.org/10.3390/ijms24076452