
Sex Difference in Cardioprotection against Acute Myocardial Infarction in MAO-B Knockout Mice In Vivo
 , ,
, ,
Abstract
:1. Introduction
2. Results
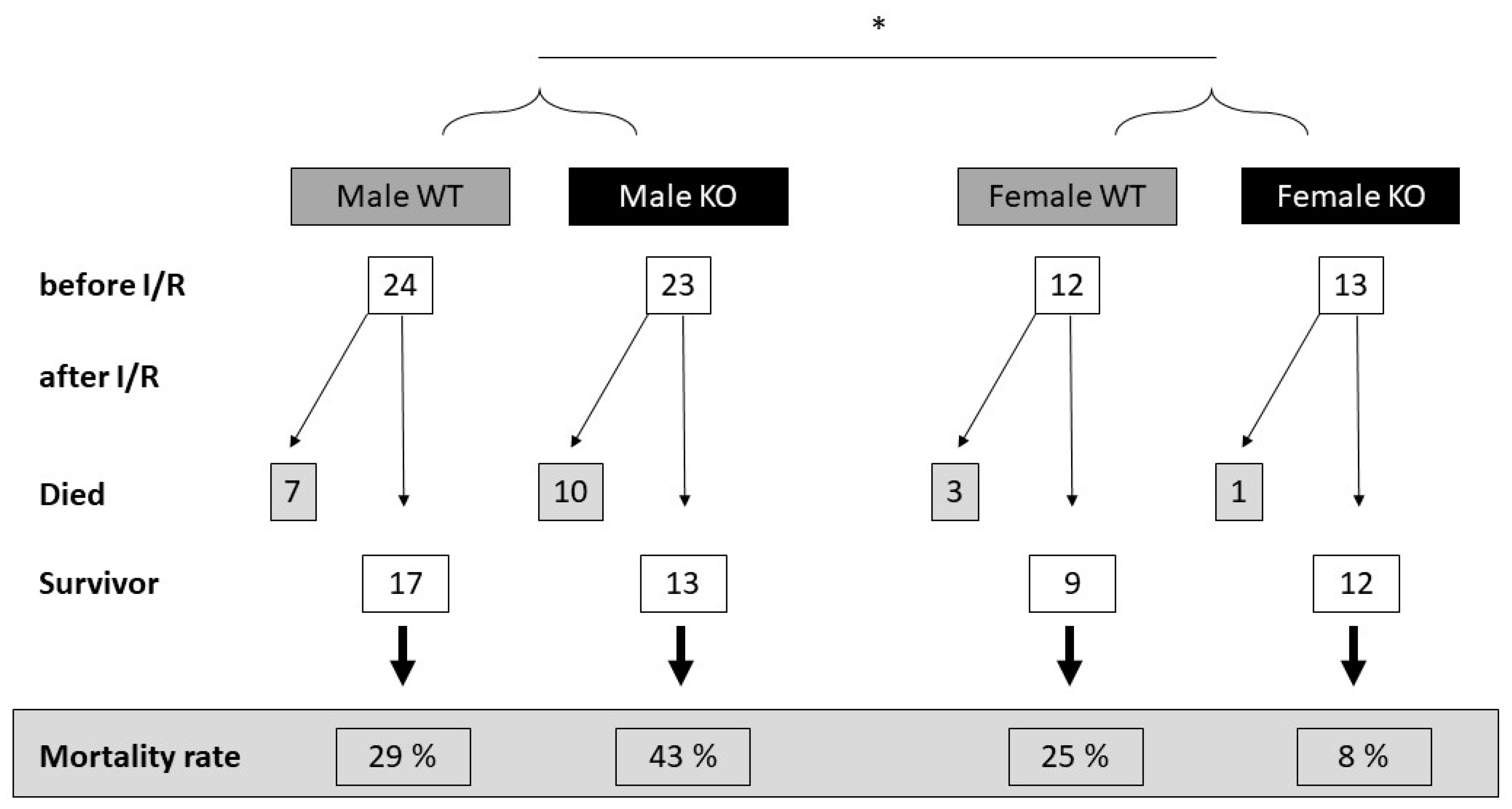
2.1. Cardiomyocyte-Specific MAO-B Deletion and Mortality Rate after I/R
2.2. Cardiomyocyte-Specific MAO-B Deletion and Infarct Size
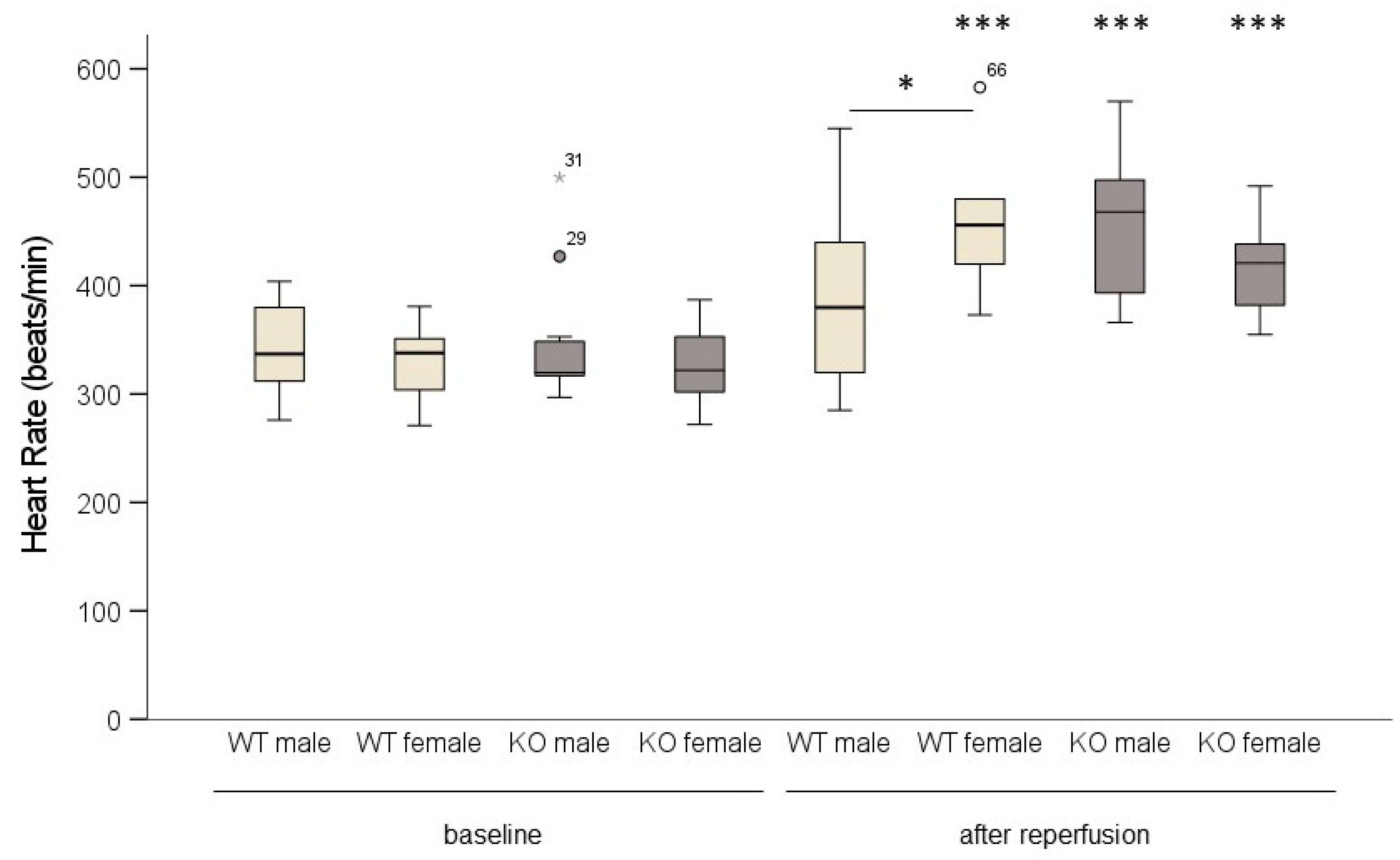
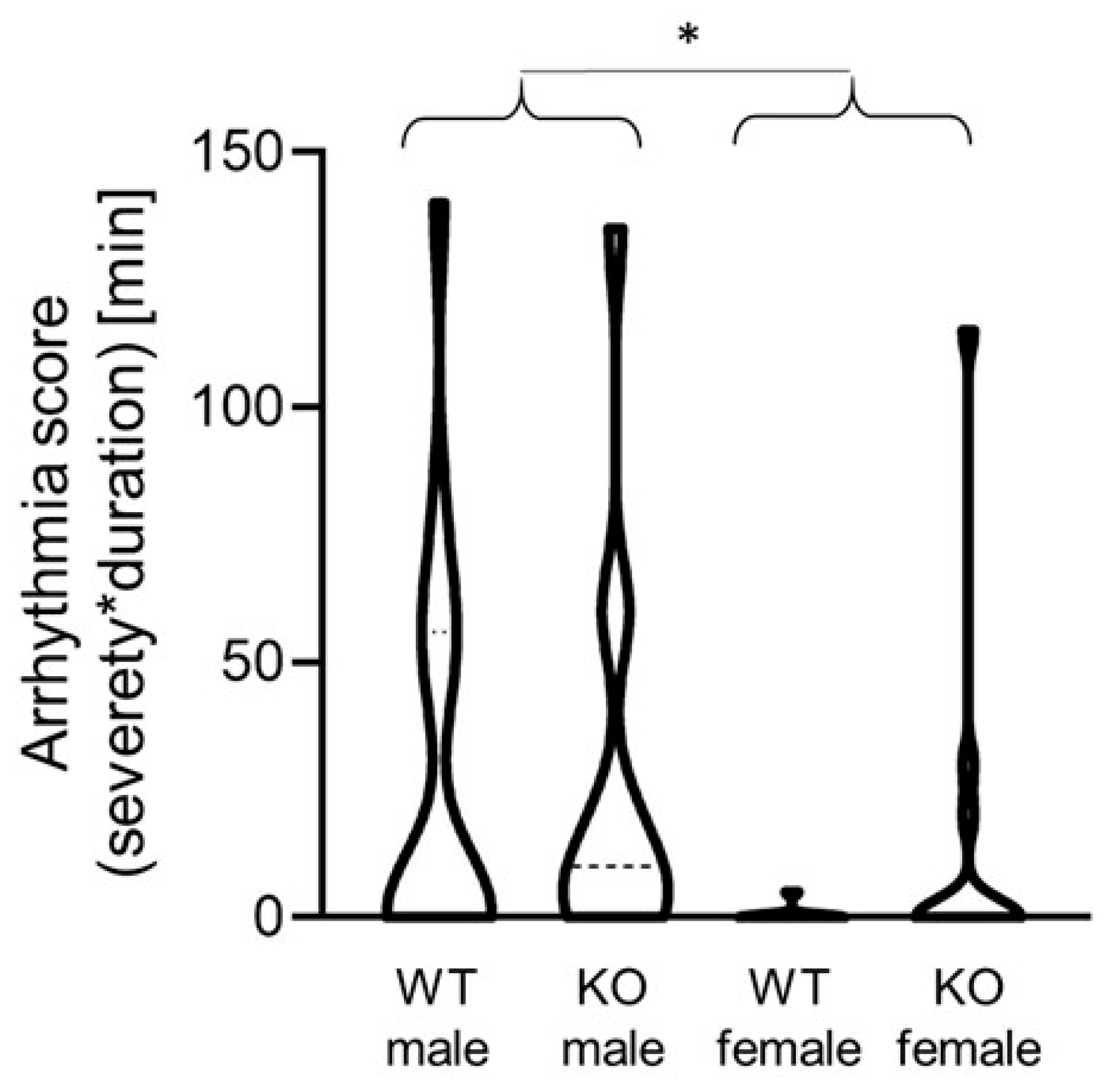
2.3. Cardiomyocyte-Specific MAO-B Deletion and Heart Rate
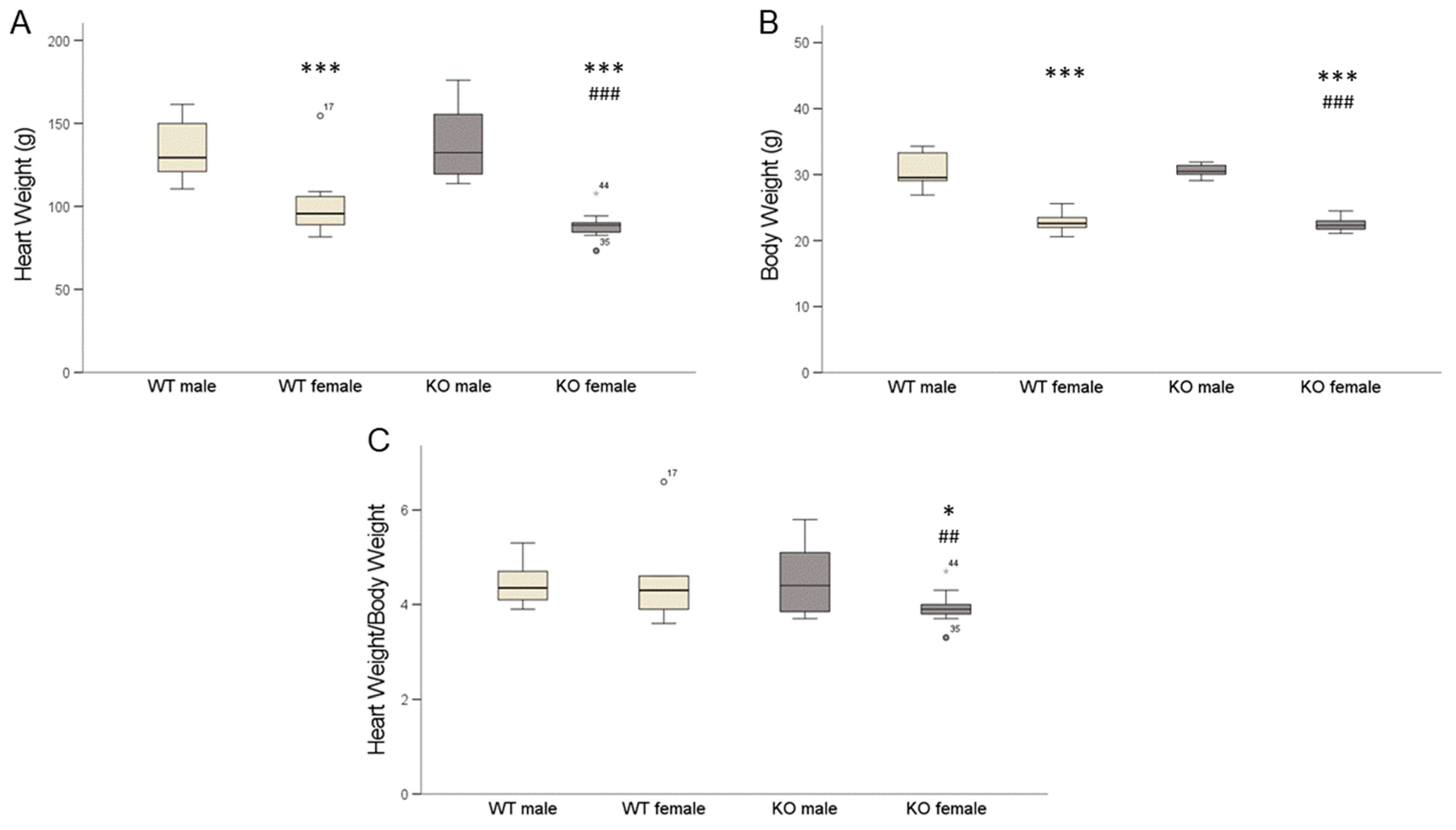
2.4. Cardiomyocyte-Specific MAO-B Knockout and Heart Weight
2.5. Cardiomyocyte-Specific MAO-B Knockout and 17β Estradiol
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Genotyping
| MAO-Bfl/fl forward | 5′-GCC CAC GAG TAA GTA AAT ACG TGG A-3′ |
| MAO-Bfl/fl reverse | 5′ GGT CTC TGT TTC TGG GAC AGT CTG-3 |
| Myh6-MCreM forward | 5′-GAC CAG GTT CGT TCA CTC ATG G-3′ |
| Myh6-MCreM revererse | 5′-AGG CTA AGT GCC TTC TCT ACA C-3′ |
4.3. Determination of Infarct Size In Vivo
4.4. Determination of 17 β Estradiol
4.5. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Di Lisa, F.; Kaludercic, N.; Carpi, A.; Menabò, R.; Giorgio, M. Mitochondrial pathways for ROS formation and myocardial injury: The relevance of p66Shc and monoamine oxidase. Basic Res. Cardiol. 2009, 104, 131–139. [Google Scholar] [CrossRef]
- Zhou, B.P.; Lewis, D.A.; Kwan, S.-W.; Abell, C.W. Flavinylation of Monoamine Oxidase B. J. Biol. Chem. 1995, 270, 23653–23660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murphy, M.P. How mitochondria produce reactive oxygen species. Biochem. J. 2009, 417, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edmondson, D.E.; Binda, C.; Mattevi, A. Structural insights into the mechanism of amine oxidation by monoamine oxidases A and B. Arch. Biochem. Biophys. 2007, 464, 269–276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Youdim, M.B.; Bakhle, Y.S. Monoamine oxidase: Isoforms and inhibitors in Parkinson’s disease and depressive illness. Br. J. Pharmacol. 2006, 147 (Suppl. 1), S287–S296. [Google Scholar] [CrossRef] [Green Version]
- Kirksey, T.J.; Kwan, S.W.; Abell, C.W. Arginine-42 and Threonine-45 are required for FAD incorporation and catalytic activity in human monoamine oxidase B. Biochemistry 1998, 37, 12360–12366. [Google Scholar] [CrossRef]
- Binda, C.; Mattevi, A.; Edmondson, D.E. Structure-Function Relationships in Flavoenzyme-dependent Amine Oxidations. J. Biol. Chem. 2002, 277, 23973–23976. [Google Scholar] [CrossRef] [Green Version]
- van Rhijn, J.-R.; Shi, Y.; Bormann, M.; Mossink, B.; Frega, M.; Recaioglu, H.; Hakobjan, M.; Gunnewiek, T.K.; Schoenmaker, C.; Palmer, E.; et al. Brunner syndrome associated MAOA mutations result in NMDAR hyperfunction and increased network activity in human dopaminergic neurons. Neurobiol. Dis. 2022, 163, 105587. [Google Scholar] [CrossRef]
- Schömig, A.; Richardt, G. Cardiac Sympathetic Activity in Myocardial Ischemia: Release and Effects of Noradrenaline. Basic Res. Cradiol. 1990, 85, 9–30. [Google Scholar] [CrossRef]
- Schömig, A.; Richardt, G.; Kurz, T. Sympatho-adrenergic activation of the ischemic myocardium and its arrhythmogenic impact. Herz 1995, 20, 169–186. [Google Scholar]
- Rodríguez, M.J.; Saura, J.; Billett, E.E.; Finch, C.C.; Mahy, N. Cellular localization of monoamine oxidase A and B in human tissues outside of the central nervous system. Cell Tissue Res. 2001, 304, 215–220. [Google Scholar] [CrossRef] [PubMed]
- Saura, J.; Nadal, E.; Berg, B.V.D.; Vila, M.; Bombi, J.; Mahy, N. Localization of monoamine oxidases in human peripheral tissues. Life Sci. 1996, 59, 1341–1349. [Google Scholar] [CrossRef] [PubMed]
- Grimsby, J.; Toth, M.; Chen, K.; Kumazawa, T.; Klaidman, L.; Adams, J.D.; Karoum, F.; Gal, J.; Shih, J.C. Increased stress response and beta-phenylethylamine in MAOB-deficient mice. Nat. Genet. 1997, 17, 206–210. [Google Scholar] [CrossRef] [PubMed]
- Maggiorani, D.; Manzella, N.; Edmondson, D.E.; Mattevi, A.; Parini, A.; Binda, C.; Mialet-Perez, J. Monoamine Oxidases, Oxidative Stress, and Altered Mitochondrial Dynamics in Cardiac Ageing. Oxidative Med. Cell. Longev. 2017, 2017, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heusch, G. Cardioprotection: Chances and challenges of its translation to the clinic. Lancet 2013, 381, 166–175. [Google Scholar] [CrossRef]
- Ferdinandy, P.; Andreadou, I.; Baxter, G.F.; Bøtker, H.E.; Davidson, S.M.; Dobrev, D.; Gersh, B.J.; Heusch, G.; Lecour, S.; Ruiz-Meana, M.; et al. Interaction of Cardiovascular Nonmodifiable Risk Factors, Comorbidities and Comedications With Ischemia/Reperfusion Injury and Cardioprotection by Pharmacological Treatments and Ischemic Conditioning. Pharmacol. Rev. 2023, 75, 159–216. [Google Scholar] [CrossRef]
- Andreadou, I.; Schulz, R.; Papapetropoulos, A.; Turan, B.; Ytrehus, K.; Ferdinandy, P.; Daiber, A.; Di Lisa, F. The role of mitochondrial reactive oxygen species, NO and H2S in ischemia/reperfusion injury and cardioprotection. J. Cell. Mol. Med. 2020, 24, 6510–6522. [Google Scholar] [CrossRef]
- Kaludercic, N.; Carpi, A.; Menabò, R.; Di Lisa, F.; Paolocci, N. Monoamine oxidases (MAO) in the pathogenesis of heart failure and ischemia/reperfusion injury. Biochim. Biophys. Acta 2011, 1813, 1323–1332. [Google Scholar] [CrossRef] [Green Version]
- Du, C.-K.; Zhan, D.-Y.; Akiyama, T.; Inagaki, T.; Shishido, T.; Shirai, M.; Pearson, J.T. Myocardial interstitial levels of serotonin and its major metabolite 5-hydroxyindole acetic acid during ischemia-reperfusion. Am. J. Physiol.-Heart Circ. Physiol. 2017, 312, H60–H67. [Google Scholar] [CrossRef] [Green Version]
- He, G.-H.; Hu, J.; Li, T.; Ma, X.; Meng, J.; Jia, M.; Lu, J.; Ohtsu, H.; Chen, Z.; Luo, X. Arrhythmogenic Effect of Sympathetic Histamine in Mouse Hearts Subjected to Acute Ischemia. Mol. Med. 2012, 18, 1–9. [Google Scholar] [CrossRef]
- Genovese, A.; Spadaro, G. Highlights in cardiovascular effects of histamine and H1-receptor antagonists. Allergy 1997, 52, 67–78. [Google Scholar] [CrossRef] [PubMed]
- He, Z.; Ma, C.; Yu, T.; Song, J.; Leng, J.; Gu, X.; Li, J. Activation mechanisms and multifaceted effects of mast cells in ischemia reperfusion injury. Exp. Cell Res. 2019, 376, 227–235. [Google Scholar] [CrossRef]
- Valen, G.; Kaszaki, J.; Szabo, I.; Nagy, S.; Vaage, J. Histamine release and its effects in ischaemia-reperfusion injury of the isolated rat heart. Acta Physiol. Scand. 1994, 150, 413–424. [Google Scholar] [CrossRef]
- Costiniti, V.; Spera, I.; Menabò, R.; Palmieri, E.M.; Menga, A.; Scarcia, P.; Porcelli, V.; Gissi, R.; Castegna, A.; Canton, M. Monoamine oxidase-dependent histamine catabolism accounts for post-ischemic cardiac redox imbalance and injury. Biochim. Biophys. Acta 2018, 1864, 3050–3059. [Google Scholar] [CrossRef]
- Kaludercic, N.; Mialet-Perez, J.; Paolocci, N.; Parini, A.; Di Lisa, F. Monoamine oxidases as sources of oxidants in the heart. J. Mol. Cell. Cardiol. 2014, 73, 34–42. [Google Scholar] [CrossRef] [Green Version]
- Yeung, A.W.K.; Georgieva, M.G.; Atanasov, A.G.; Tzvetkov, N.T. Monoamine Oxidases (MAOs) as Privileged Molecular Targets in Neuroscience: Research Literature Analysis. Front. Mol. Neurosci. 2019, 12, 143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finberg, J.; Youdim, M.B. Modification of blood pressure and nictitating membrane response to sympathetic amines by selective monoamine oxidase inhibitors, types A and B, in the cat. Br. J. Pharmacol. 1985, 85, 541–546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scotton, W.J.; Hill, L.J.; Williams, A.C.; Barnes, N.M. Serotonin Syndrome: Pathophysiology, Clinical Features, Management, and Potential Future Directions. Int. J. Tryptophan Res. 2019, 12, 1178646919873925. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antonucci, S.; Di Sante, M.; Tonolo, F.; Pontarollo, L.; Scalcon, V.; Alanova, P.; Menabò, R.; Carpi, A.; Bindoli, A.; Rigobello, M.P.; et al. The Determining Role of Mitochondrial Reactive Oxygen Species Generation and Monoamine Oxidase Activity in Doxorubicin-Induced Cardiotoxicity. Antioxid. Redox Signal. 2021, 34, 531–550. [Google Scholar] [CrossRef]
- Cases, O.; Seif, I.; Grimsby, J.; Gaspar, P.; Chen, K.; Pournin, S.; Müller, U.; Aguet, M.; Babinet, C.; Shih, J.C.; et al. Aggressive Behavior and Altered Amounts of Brain Serotonin and Norepinephrine in Mice Lacking MAOA. Science 1995, 268, 1763–1766. [Google Scholar] [CrossRef] [Green Version]
- Chen, K.; Holschneider, D.P.; Wu, W.; Rebrin, I.; Shih, J.C. A Spontaneous Point Mutation Produces Monoamine Oxidase A/B Knock-out Mice with Greatly Elevated Monoamines and Anxiety-like Behavior. J. Biol. Chem. 2004, 279, 39645–39652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heger, J.; Hirschhäuser, C.; Bornbaum, J.; Sydykov, A.; Dempfle, A.; Schneider, A.; Braun, T.; Schlüter, K.D.; Schulz, R. Cardiomyocytes-specific deletion of monoamine oxidase B reduces irreversible myocardial ischemia/reperfusion injury. Free Radic. Biol. Med. 2021, 165, 14–23. [Google Scholar] [CrossRef] [PubMed]
- Murphy, E.; Steenbergen, C. Gender-based differences in mechanisms of protection in myocardial ischemia–reperfusion injury. Cardiovasc. Res. 2007, 75, 478–486. [Google Scholar] [CrossRef] [Green Version]
- Ostadal, B.; Ostadal, P. Sex-based differences in cardiac ischaemic injury and protection: Therapeutic implications. Br. J. Pharmacol. 2014, 171, 541–554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruiz-Meana, M.; Boengler, K.; Garcia-Dorado, D.; Hausenloy, D.J.; Kaambre, T.; Kararigas, G.; Perrino, C.; Schulz, R.; Ytrehus, K. Ageing, sex, and cardioprotection. Br. J. Pharmacol. 2020, 177, 5270–5286. [Google Scholar] [CrossRef]
- Perrino, C.; Ferdinandy, P.; Bøtker, H.E.; Brundel, B.J.J.M.; Collins, P.; Davidson, S.M.; Ruijter, H.M.D.; Engel, F.B.; Gerdts, E.; Girao, H.; et al. Improving translational research in sex-specific effects of comorbidities and risk factors in ischaemic heart disease and cardioprotection: Position paper and recommendations of the ESC Working Group on Cellular Biology of the Heart. Cardiovasc. Res. 2020, 117, 367–385. [Google Scholar] [CrossRef]
- Lagranha, C.J.; Deschamps, A.; Aponte, A.; Steenbergen, C.; Murphy, E. Sex Differences in the Phosphorylation of Mitochondrial Proteins Result in Reduced Production of Reactive Oxygen Species and Cardioprotection in Females. Circ. Res. 2010, 106, 1681–1691. [Google Scholar] [CrossRef] [Green Version]
- Johnson, M.S.; Moore, R.L.; Brown, D.A. Sex differences in myocardial infarct size are abolished by sarcolemmal KATP channel blockade in rat. Am. J. Physiol. Circ. Physiol. 2006, 290, H2644–H2647. [Google Scholar] [CrossRef] [Green Version]
- Robinson, D.S.; Davis, J.M.; Nies, A.; Ravaris, C.L.; Sylwester, D. Relation of Sex and Aging to Monoamine Oxidase Activity of Human Brain, Plasma, and Platelets. Arch. Gen. Psychiatry 1971, 24, 536–539. [Google Scholar] [CrossRef]
- Zhang, Z.; Chen, K.; Shih, J.C.; Teng, C.T. Estrogen-Related Receptors-Stimulated Monoamine Oxidase B Promoter Activity Is Down-Regulated by Estrogen Receptors. Mol. Endocrinol. 2006, 20, 1547–1561. [Google Scholar] [CrossRef] [Green Version]
- Sramek, J.J.; Murphy, M.F.; Cutler, N.R. Sex differences in the psychopharmacological treatment of depression. Dialogues Clin. Neurosci. 2016, 18, 447–457. [Google Scholar] [CrossRef] [PubMed]
- Quitkin, F.M.; Stewart, J.W.; McGrath, P.J.; Taylor, B.P.; Tisminetzky, M.S.; Petkova, E.; Chen, Y.; Ma, G.; Klein, D.F. Are there differences between women’s and men’s antidepressant responses? Am. J. Psychiatry 2002, 159, 1848–1854. [Google Scholar] [CrossRef] [PubMed]
- Curtis, M.J.; Hancox, J.C.; Farkas, A.; Wainwright, C.L.; Stables, C.L.; Saint, D.A.; Clements-Jewery, H.; Lambiase, P.D.; Billman, G.E.; Janse, M.J.; et al. The Lambeth Conventions (II): Guidelines for the study of animal and human ventricular and supraventricular arrhythmias. Pharmacol. Ther. 2013, 139, 213–248. [Google Scholar] [CrossRef]
- Morkuniene, R.; Arandarcikaite, O.; Ivanoviene, L.; Borutaite, V. Estradiol-induced protection against ischemia-induced heart mitochondrial damage and caspase activation is mediated by protein kinase G. Biochim. Biophys. Acta 2010, 1797, 1012–1017. [Google Scholar] [CrossRef] [Green Version]
- Pavón, N.; Martínez-Abundis, E.; Hernández, L.; Gallardo-Pérez, J.C.; Alvarez-Delgado, C.; Cerbón, M.; Pérez-Torres, I.; Aranda, A.; Chávez, E. Sexual hormones: Effects on cardiac and mitochondrial activity after ischemia-reperfusion in adult rats. Gender difference. J. Steroid Biochem. Mol. Biol. 2012, 132, 135–146. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Hernández, O.T.; Martínez-Mota, L.; Herrera-Pérez, J.J.; Jiménez-Rubio, G. Role of Estradiol in the Expression of Genes Involved in Serotonin Neurotransmission: Implications for Female Depression. Curr. Neuropharmacol. 2019, 17, 459–471. [Google Scholar] [CrossRef]
- Querio, G.; Geddo, F.; Antoniotti, S.; Gallo, M.P.; Penna, C. Sex and Response to Cardioprotective Conditioning Maneuvers. Front. Physiol. 2021, 12, 667961. [Google Scholar] [CrossRef]
- Nilsson, S.; Mäkelä, S.; Treuter, E.; Tujague, M.; Thomsen, J.; Andersson, G.; Enmark, E.; Pettersson, K.; Warner, M.; Gustafsson, J.A. Mechanisms of Estrogen Action. Physiol. Rev. 2001, 81, 1535–1565. [Google Scholar] [CrossRef]
- Yang, S.H.; Liu, R.; Perez, E.J.; Wen, Y.; Stevens, S.M.; Valencia, T.; Brun-Zinkernagel, A.M.; Prokai, L.; Will, Y.; Dykens, J. Mitochondrial localization of estrogen receptor beta. Proc. Natl. Acad. Sci. USA 2004, 101, 4130–4135. [Google Scholar] [CrossRef] [Green Version]
- Klaiber, E.L.; Broverman, D.M.; Haffajee, C.I.; Hochman, J.S.; Sacks, G.M.; Dalen, J.E. Serum estrogen levels in men with acute myocardial infarction. Am. J. Med. 1982, 73, 872–881. [Google Scholar] [CrossRef]
- Purves-Tyson, T.D.; Handelsman, D.J.; Double, K.L.; Owens, S.J.; Bustamante, S.; Weickert, C.S. Testosterone regulation of sex steroid-related mRNAs and dopamine-related mRNAs in adolescent male rat substantia nigra. BMC Neurosci. 2012, 13, 95. [Google Scholar] [CrossRef] [Green Version]
- Le, T.Y.L.; Ashton, A.W.; Mardini, M.; Stanton, P.G.; Funder, J.W.; Handelsman, D.J.; Mihailidou, A.S. Role of Androgens in Sex Differences in Cardiac Damage During Myocardial Infarction. Endocrinology 2014, 155, 568–575. [Google Scholar] [CrossRef]
- Alnouti, Y.; Klaassen, C.D. Tissue Distribution, Ontogeny, and Regulation of Aldehyde Dehydrogenase (Aldh) Enzymes mRNA by Prototypical Microsomal Enzyme Inducers in Mice. Toxicol. Sci. 2008, 101, 51–64. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.-H.; Budas, G.R.; Churchill, E.N.; Disatnik, M.-H.; Hurley, T.D.; Mochly-Rosen, D. Activation of Aldehyde Dehydrogenase-2 Reduces Ischemic Damage to the Heart. Science 2008, 321, 1493–1495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eaton, P.; Li, J.-M.; Hearse, D.J.; Shattock, M.J. Formation of 4-hydroxy-2-nonenal-modified proteins in ischemic rat heart. Am. J. Physiol.-Heart Circ. Physiol. 1999, 276 Pt 2, H935–H943. [Google Scholar] [CrossRef] [Green Version]
- Santin, Y.; Fazal, L.; Sainte-Marie, Y.; Sicard, P.; Maggiorani, D.; Tortosa, F.; Yücel, Y.Y.; Teyssedre, L.; Rouquette, J.; Marcellin, M.; et al. Mitochondrial 4-HNE derived from MAO-A promotes mitoCa2+ overload in chronic postischemic cardiac remodeling. Cell Death Differ. 2020, 27, 1907–1923. [Google Scholar] [CrossRef]
- Kaludercic, N.; Carpi, A.; Nagayama, T.; Sivakumaran, V.; Zhu, G.; Lai, E.W.; Bedja, D.; De Mario, A.; Chen, K.; Gabrielson, K.L.; et al. Monoamine Oxidase B Prompts Mitochondrial and Cardiac Dysfunction in Pressure Overloaded Hearts. Antioxid. Redox Signal. 2014, 20, 267–280. [Google Scholar] [CrossRef] [Green Version]
- Ma, H.; Guo, R.; Yu, L.; Zhang, Y.; Ren, J. Aldehyde dehydrogenase 2 (ALDH2) rescues myocardial ischaemia/reperfusion injury: Role of autophagy paradox and toxic aldehyde. Eur. Heart J. 2010, 32, 1025–1038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Zhao, M.; Shi, Q.; Xu, B.; Zhu, C.; Li, M.; Mir, V.; Bers, D.M.; Xiang, Y.K. Monoamine Oxidases Desensitize Intracellular β1AR Signaling in Heart Failure. Circ. Res. 2021, 129, 965–967. [Google Scholar] [CrossRef] [PubMed]
- Hahnova, K.; Brabcova, I.; Neckar, J.; Weissova, R.; Svatonova, A.; Novakova, O.; Zurmanova, J.; Kalous, M.; Silhavy, J.; Pravenec, M.; et al. β-Adrenergic signaling, monoamine oxidase A and antioxidant defence in the myocardium of SHR and SHR-mtBN conplastic rat strains: The effect of chronic hypoxia. J. Physiol. Sci. 2018, 68, 441–454. [Google Scholar] [CrossRef] [Green Version]
- Csonka, C.; Kupai, K.; Kocsis, G.F.; Novák, G.; Fekete, V.; Bencsik, P.; Csont, T.; Ferdinandy, P. Measurement of myokardial infarct size in preclinical studies. J. Pharmacol. Toxicol. Methods 2010, 61, 163–170. [Google Scholar] [CrossRef] [PubMed]


 ). * p < 0.05, *** p < 0.001 baseline vs. after reperfusion, Student’s t-test. Two-way ANOVA: p < 0.001, baseline vs. after reperfusion p < 0.001, genotype * sex p = 0.02.
). * p < 0.05, *** p < 0.001 baseline vs. after reperfusion, Student’s t-test. Two-way ANOVA: p < 0.001, baseline vs. after reperfusion p < 0.001, genotype * sex p = 0.02.
). * p < 0.05, *** p < 0.001 baseline vs. after reperfusion, Student’s t-test. Two-way ANOVA: p < 0.001, baseline vs. after reperfusion p < 0.001, genotype * sex p = 0.02.
). * p < 0.05, *** p < 0.001 baseline vs. after reperfusion, Student’s t-test. Two-way ANOVA: p < 0.001, baseline vs. after reperfusion p < 0.001, genotype * sex p = 0.02.
 ).
).
).
).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| WT | MAO-B KO | |||
|---|---|---|---|---|
| Male | Female | Male | Female | |
| Mean | 27.9 *,## | 25.1 | 23.7 | 23.9 |
| SEM | 0.81 | 0.96 | 1.01 | 0.81 |
| N | 9 | 9 | 8 | 11 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Heger, J.; Szabados, T.; Brosinsky, P.; Bencsik, P.; Ferdinandy, P.; Schulz, R. Sex Difference in Cardioprotection against Acute Myocardial Infarction in MAO-B Knockout Mice In Vivo. Int. J. Mol. Sci. 2023, 24, 6443. https://doi.org/10.3390/ijms24076443
Heger J, Szabados T, Brosinsky P, Bencsik P, Ferdinandy P, Schulz R. Sex Difference in Cardioprotection against Acute Myocardial Infarction in MAO-B Knockout Mice In Vivo. International Journal of Molecular Sciences. 2023; 24(7):6443. https://doi.org/10.3390/ijms24076443
Chicago/Turabian StyleHeger, Jacqueline, Tamara Szabados, Paulin Brosinsky, Péter Bencsik, Péter Ferdinandy, and Rainer Schulz. 2023. "Sex Difference in Cardioprotection against Acute Myocardial Infarction in MAO-B Knockout Mice In Vivo" International Journal of Molecular Sciences 24, no. 7: 6443. https://doi.org/10.3390/ijms24076443