
Interaction of Terminal Oxidases with Amphipathic Molecules

 ,
,  ,
, 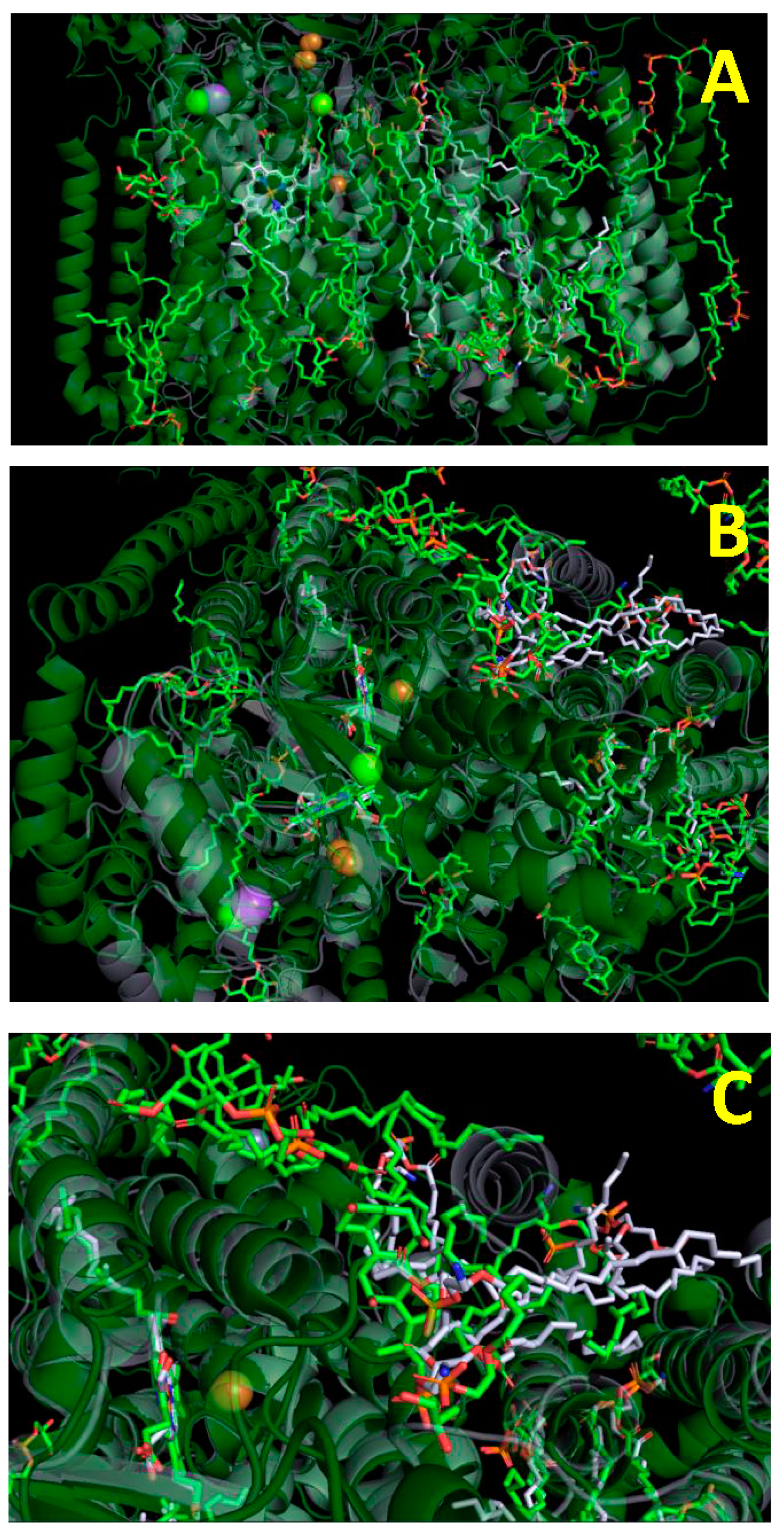{kind=link}
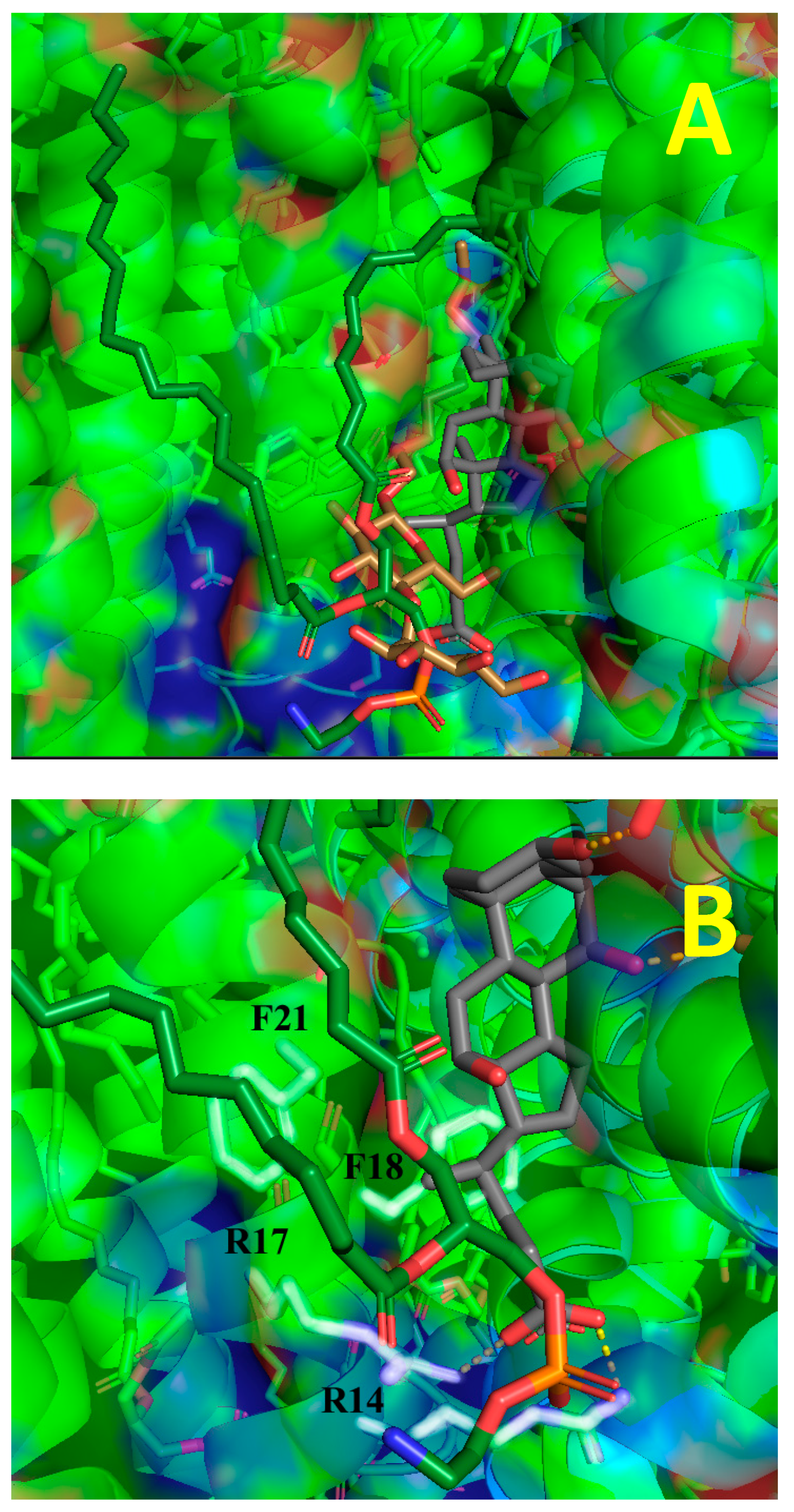{kind=link}
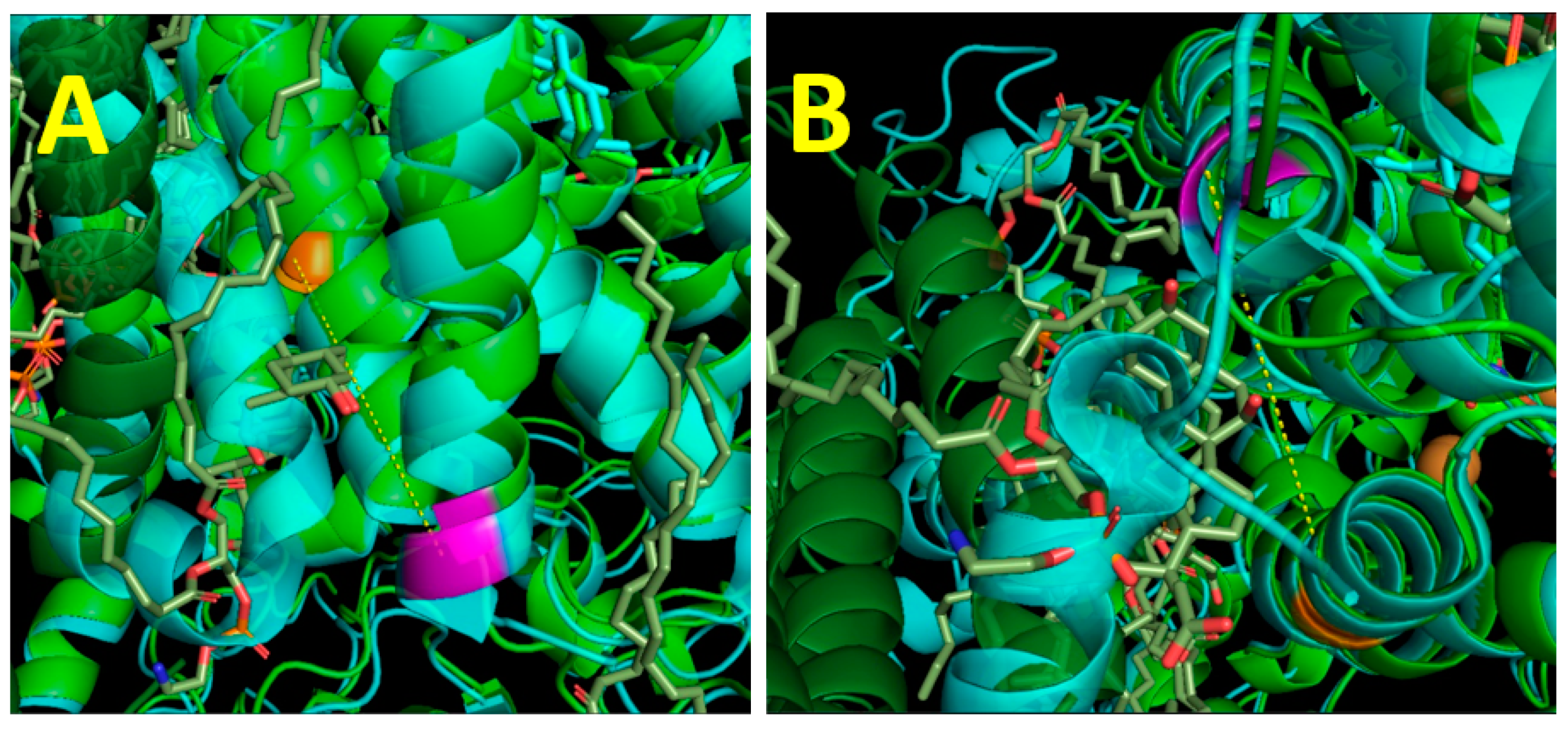{kind=link}
Abstract
:1. Introduction
- -
- Lipids.
- -
- Detergents.
- -
- Relatively small amphipathic molecules.
2. Lipids
3. Detergents
4. Potential Ligands of the Regulatory Sites
5. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Siletsky, S.A.; Borisov, V.B. Proton pumping and non-pumping terminal respiratory oxidases: Active sites intermediates of these molecular machines and their derivatives. Int. J. Mol. Sci. 2021, 22, 10852. [Google Scholar] [CrossRef]
- van der Oost, J.; De Boer, A.P.; de Gier, J.W.L.; Zumft, W.G.; Stouthamer, A.H.; van Spanning, R.J. The heme-copper oxidase family consists of three distinct types of terminal oxidases and is related to nitric oxide reductase. FEMS Microb. Lett. 1994, 121, 1–9. [Google Scholar] [CrossRef]
- Garcia-Horseman, J.A.; Barquera, B.; Rumbley, J.; Ma, J.; Gennis, R.B. The superfamily of heme-copper respiratory oxidases. J. Bacteriol. 1994, 176, 5587–5600. [Google Scholar] [CrossRef] [Green Version]
- Pereira, M.M.; Santana, M.; Teixeira, M. A novel scenario for the evolution of haem–copper oxygen reductases. Biochim. Biophys. Acta (BBA)-Bioenerg. 2001, 1505, 185–208. [Google Scholar] [CrossRef] [Green Version]
- Sousa, F.L.; Alves, R.J.; Ribeiro, M.A.; Pereira-Leal, J.B.; Teixeira, M.; Pereira, M.M. The superfamily of heme–copper oxygen reductases: Types and evolutionary considerations. Biochim. Biophys. Acta (BBA)-Bioenerg. 2012, 1817, 629–637. [Google Scholar] [CrossRef] [Green Version]
- Maréchal, A.; Xu, J.Y.; Genko, N.; Hartley, A.M.; Haraux, F.; Meunier, B.; Rich, P.R. A common coupling mechanism for A-type heme-copper oxidases from bacteria to mitochondria. Proc. Natl. Acad. Sci. USA 2020, 117, 9349–9355. [Google Scholar] [CrossRef] [Green Version]
- Rich, P.R. Mitochondrial cytochrome c oxidase: Catalysis, coupling and controversies. Biochem. Soc. Trans. 2017, 45, 813–829. [Google Scholar] [CrossRef]
- Wikström, M.; Sharma, V. Proton pumping by cytochrome c oxidase—A 40 year anniversary. Biochim. Biophys. Acta (BBA)-Bioenerg. 2018, 1859, 692–698. [Google Scholar] [CrossRef]
- von Ballmoos, C.; Ädelroth, P.; Gennis, R.B.; Brzezinski, P. Proton transfer in ba3 cytochrome c oxidase from Thermus thermophilus. Biochim. Biophys. Acta (BBA)-Bioenerg. 2012, 1817, 650–657. [Google Scholar] [CrossRef] [Green Version]
- Buschmann, S.; Warkentin, E.; Xie, H.; Langer, J.D.; Ermler, U.; Michel, H. The structure of cbb3 cytochrome oxidase provides insights into proton pumping. Science 2010, 329, 327–330. [Google Scholar] [CrossRef]
- Tsukihara, T.; Aoyama, H.; Yamashita, E.; Tomizaki, T.; Yamaguchi, H.; Shinzawa-Itoh, K.; Nakashima, R.; Yaono, R.; Yoshikawa, S. The whole structure of the 13-subunit oxidized cytochrome c oxidase at 2.8 Å. Science 1996, 272, 1136–1144. [Google Scholar] [CrossRef]
- Yoshikawa, S.; Shinzawa-Itoh, K.; Nakashima, R.; Yaono, R.; Yamashita, E.; Inoue, N.; Yao, M.; Fei, M.J.; Libeu, C.P.; Mizushima, T.; et al. Redox-coupled crystal structural changes in bovine heart cytochrome c oxidase. Science 1998, 280, 1723–1729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shinzawa-Itoh, K.; Sugimura, T.; Misaki, T.; Tadehara, Y.; Yamamoto, S.; Hanada, M.; Yano, N.; Nakagawa, T.; Uene, S.; Yamada, T.; et al. Monomeric structure of an active form of bovine cytochrome c oxidase. Proc. Natl. Acad. Sci. USA 2019, 116, 19945–19951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iwata, S.; Ostermeier, C.; Ludwig, B.; Michel, H. Structure at 2.8 Å resolution of cytochrome c oxidase from Paracoccus Denitrificans. Nature 1995, 376, 660–669. [Google Scholar]
- Koepke, J.; Olkhova, E.; Angerer, H.; Muller, H.; Peng, G.; Michel, H. High resolution crystal structure of Paracoccus denitrificans cytochrome c oxidase: New insights into the active site and the proton transfer pathways. Biochim. Biophys. Acta (BBA)-Bioenerg. 2009, 1787, 635–645. [Google Scholar] [CrossRef] [Green Version]
- Svensson-Ek, M.; Abramson, J.; Larsson, G.; Törnroth, S.; Brzezinski, P.; Iwata, S. The X-ray crystal structures of wild-type and EQ (I-286) mutant cytochrome c oxidases from Rhodobacter sphaeroides. J. Mol. Biol. 2002, 321, 329–339. [Google Scholar] [CrossRef]
- Qin, L.; Hiser, C.; Mulichak, A.; Garavito, R.M.; Ferguson-Miller, S. Identification of conserved lipid/detergent-binding sites in a high-resolution structure of the membrane protein cytochrome c oxidase. Proc. Natl. Acad. Sci. USA 2006, 103, 16117–16122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rumbley, J.N.; Nickels, E.F.; Gennis, R.B. One-step purification of histidine-tagged cytochrome bo3 from Escherichia coli and demonstration that associated quinone is not required for the structural integrity of the oxidase. Biochim. Biophys. Acta Protein Struct. Mol. Enzymol. 1997, 1340, 131–142. [Google Scholar] [CrossRef]
- Melin, F.; Meyer, T.; Lankiang, S.; Choi, S.K.; Gennis, R.B.; Blanck, C.; Schmutz, M.; Hellwig, P. Direct electrochemistry of cytochrome bo3 oxidase at a series of gold nanoparticles-modified electrodes. Electrochem. Commun. 2013, 26, 105–108. [Google Scholar] [CrossRef] [Green Version]
- Soulimane, T.; Buse, G.; Bourenkov, G.P.; Bartunik, H.D.; Huber, R.; Than, M.E. Structure and mechanism of the aberrant ba3-cytochrome c oxidase from Thermus thermophilus. EMBO J. 2000, 19, 1766–1776. [Google Scholar] [CrossRef] [Green Version]
- Hunsicker-Wang, L.M.; Pacoma, R.L.; Chen, Y.; Fee, J.A.; Stout, C.D. A novel cryoprotection scheme for enhancing the diffraction of crystals of recombinant cytochrome ba3 oxidase from Thermus thermophilus. Acta Crystallogr. D Biol. Cryst. 2005, 61, 340–343. [Google Scholar] [CrossRef] [PubMed]
- Tiefenbrunn, T.; Liu, W.; Chen, Y.; Katritch, V.; Stout, C.D.; Fee, J.A.; Cherezov, V. High resolution structure of the ba3 cytochrome c oxidase from Thermus thermophilus in a lipidic environment. PLoS ONE 2011, 6, e22348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pitcher, R.S.; Watmough, N.J. The bacterial cytochrome cbb3 oxidases. Biochim. Biophys. Acta (BBA)-Bioenerg. 2004, 1655, 388–399. [Google Scholar] [CrossRef] [PubMed]
- Hamada, M.; Toyofuku, M.; Miyano, T.; Nomura, N. cbb3-type cytochrome c oxidases, aerobic respiratory enzymes, impact the anaerobic life of Pseudomonas aeruginosa PAO1. J. Bacteriol. 2014, 196, 3881–3889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forte, E.; Borisov, V.B.; Vicente, J.B.; Giuffre, A. Cytochrome bd and gaseous ligands in bacterial physiology. Adv. Microb. Physiol. 2017, 71, 171–234. [Google Scholar]
- Borisov, V.B.; Gennis, R.B.; Hemp, J.; Verkhovsky, M.I. The cytochrome bd respiratory oxygen reductases. Biochim. Biophys. Acta (BBA)-Bioenerg. 2011, 1807, 1398–1413. [Google Scholar] [CrossRef] [Green Version]
- Safarian, S.; Rajendran, C.; Müller, H.; Preu, J.; Langer, J.D.; Ovchinnikov, S.; Hirose, T.; Kusumoto, T.; Sakamoto, J.; Michel, H. Structure of a bd oxidase indicates similar mechanisms for membrane-integrated oxygen reductases. Science 2016, 352, 583–586. [Google Scholar] [CrossRef] [Green Version]
- Safarian, S.; Hahn, A.; Mills, D.J.; Radloff, M.; Eisinger, M.L.; Nikolaev, A.; Meier-Credo, J.; Melin, F.; Miyoshi, H.; Gennis, R.B.; et al. Active site rearrangement and structural divergence in prokaryotic respiratory oxidases. Science 2019, 366, 100–104. [Google Scholar] [CrossRef]
- Friedrich, T.; Wohlwend, D.; Borisov, V.B. Recent advances in structural studies of cytochrome bd and its potential application as a drug target. Int. J. Mol. Sci. 2022, 23, 3166. [Google Scholar] [CrossRef]
- Borisov, V.B.; Siletsky, S.A.; Paiardini, A.; Hoogewijs, D.; Forte, E.; Giuffre, A.; Poole, R.K. Bacterial oxidases of the cytochrome bd family: Redox enzymes of unique structure, function and utility as drug targets. Antioxid. Redox Signal. 2021, 34, 1280–1318. [Google Scholar] [CrossRef]
- Forte, E.; Siletsky, S.A.; Borisov, V.B. In Escherichia coli ammonia inhibits cytochrome bo3 but activates cytochrome bd-I. Antioxidants 2021, 10, 13. [Google Scholar] [CrossRef] [PubMed]
- Kadenbach, B.; Hüttemann, M. The subunit composition and function of mammalian cytochrome c oxidase. Mitochondrion 2015, 24, 64–76. [Google Scholar] [CrossRef] [PubMed]
- Thesseling, A.; Rasmussen, T.; Burschel, S.; Wohlwend, D.; Kagi, J.; Muller, R.; Bottcher, B.; Friedrich, T. Homologous bd oxidases share the same architecture but differ in mechanism. Nat. Commun. 2019, 10, 5138. [Google Scholar] [CrossRef] [Green Version]
- Liko, I.; Degiacomi, M.T.; Mohammed, S.; Yoshikawa, S.; Schmidt, C.; Robinson, C.V. Dimer interface of bovine cytochrome c oxidase is influenced by local posttranslational modifications and lipid binding. Proc. Natl. Acad. Sci. USA 2016, 113, 8230–8235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Genova, M.L.; Lenaz, G. Functional role of mitochondrial respiratory supercomplexes. Biochim. Biophys. Acta (BBA)-Bioenerg. 2014, 1837, 427–443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shinzawa-Itoh, K.; Muramoto, K. Biochemical and crystallographic studies of monomeric and dimeric bovine cytochrome c oxidase. Biophys. Phys. 2021, 18, 186–195. [Google Scholar] [CrossRef]
- Helling, S.; Hüttemann, M.; Ramzan, R.; Kim, S.H.; Lee, I.; Müller, T.; Langenfeld, E.; Meyer, H.E.; Kadenbach, B.; Vogt, S.; et al. Multiple phosphorylations of cytochrome c oxidase and their functions. Proteomics 2012, 12, 950–959. [Google Scholar] [CrossRef] [Green Version]
- Kadenbach, B. Regulation of Mammalian 13-Subunit Cytochrome c Oxidase and Binding of other Proteins: Role of NDUFA4. Trends Endocrinol. Metab. 2017, 28, 761–770. [Google Scholar] [CrossRef]
- Cooper, C.E.; Brown, G.C. The inhibition of mitochondrial cytochrome oxidase by the gases carbon monoxide, nitric oxide, hydrogen cyanide and hydrogen sulfide: Chemical mechanism and physiological significance. J. Bioenerg. Biomembr. 2008, 40, 533–539. [Google Scholar] [CrossRef]
- Vygodina, T.V.; Mukhaleva, E.; Azarkina, N.V.; Konstantinov, A.A. Cytochrome c oxidase inhibition by calcium at physiological ionic composition of the medium: Implications for physiological significance of the effect. Biochim. Biophys. Acta (BBA)-Bioenerg. 2017, 1858, 982–990. [Google Scholar] [CrossRef]
- Qin, L.; Mills, D.A.; Buhrow, L.; Hiser, C.; Ferguson-Miller, S. A Conserved steroid binding site in cytochrome c oxidase. Biochemistry 2008, 47, 9931–9933. [Google Scholar] [CrossRef] [Green Version]
- Hiser, C.; Buhrow, L.; Liu, J.; Kuhn, L.; Ferguson-Miller, S. A conserved amphipathic ligand binding region influences K-path-dependent activity of cytochrome c oxidase. Biochemistry 2013, 52, 1385–1396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buhrow, L.; Hiser, C.; van Voorst, J.R.; Ferguson-Miller, S.; Kuhn, L.A. Computational prediction and in vitro analysis of potential physiological ligands of the bile acid binding site in cytochrome c oxidase. Biochemistry 2013, 52, 6995–7006. [Google Scholar] [CrossRef] [Green Version]
- Di Trani, J.M.; Moe, A.; Riepl, D.; Saura, P.; Kaila, V.R.; Brzezinski, P.; Rubinstein, J.L. Structural basis of mammalian complex IV inhibition by steroids. Proc. Natl. Acad. Sci. USA 2022, 119, e2205228119. [Google Scholar] [CrossRef] [PubMed]
- Brierley, G.P.; Merola, A.J.; Fleischer, S. Studies of the electron-transfer system XLIX. Sites of phospholipid involvement in the electron-transfer chain. Biochim. Biophys. Acta 1962, 64, 218–228. [Google Scholar] [CrossRef] [PubMed]
- Awasthi, Y.C.; Chuang, T.F.; Keenan, T.W.; Crane, F.L. Tightly bound cardiolipin in cytochrome oxidase. Biochim. Biophys. Acta (BBA)-Bioenerg. 1971, 226, 42–52. [Google Scholar] [CrossRef]
- Yu, C.; Yu, L.; King, T.E. Studies on cytochrome oxidase. Interactions of the cytochrome oxidase protein with phospholipids and cytochrome c. J. Biol. Chem. 1975, 250, 1383–1392. [Google Scholar] [CrossRef]
- Robinson, N.C.; Capaldi, R.A. Components of the mitochondrial inner membrane. 5. Interaction of detergents with cytochrome c oxidase. Biochemistry 1977, 16, 375–381. [Google Scholar] [CrossRef]
- Robinson, N.C.; Zborowski, J.; Talbert, L.H. Cardiolipin-depleted bovine heart cytochrome c oxidase: Binding stoichiometry and affinity for cardiolipin derivatives. Biochemistry 1990, 29, 8962–8969. [Google Scholar] [CrossRef]
- Sedlák, E.; Robinson, N.C. Phospholipase A2 digestion of cardiolipin bound to bovine cytochrome c oxidase alters both activity and quaternary structure. Biochemistry 1999, 38, 14966–14972. [Google Scholar] [CrossRef]
- Lee, S.J.; Yamashita, E.; Abe, T.; Fukumoto, Y.; Tsukihara, T.; Shinzawa-Itoh, K.; Ueda, H.; Yoshikawa, S. Intermonomer interactions in dimer of bovine heart cytochrome c oxidase. Acta Crystallogr. Sect. D Biol. Crystallogr. 2001, 57, 941–947. [Google Scholar] [CrossRef]
- Musatov, A.; Robinson, N.C. Bound cardiolipin is essential for cytochrome c oxidase proton translocation. Biochimie 2014, 105, 159–164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arnarez, C.; Marrink, S.J.; Periole, X. Identification of cardiolipin binding sites on cytochrome c oxidase at the entrance of proton channels. Sci. Rep. 2013, 3, 1263. [Google Scholar] [CrossRef] [Green Version]
- Arnold, S.; Kadenbach, B. Cell respiration is controlled by ATP, an allosteric inhibitor of cytochrome-c oxidase. Eur. J. Biochem. 1997, 249, 350–354. [Google Scholar] [CrossRef] [PubMed]
- Sedlak, E.; Kozar, T.; Musatov, A. The interplay among subunit composition, cardiolipin content, and aggregation state of bovine heart cytochrome c oxidase. Cells 2020, 9, 2588. [Google Scholar] [CrossRef] [PubMed]
- Ramzan, R.; Kadenbach, B.; Vogt, S. Multiple mechanisms regulate eukaryotic cytochrome c oxidase. Cells 2021, 10, 514. [Google Scholar] [CrossRef]
- Arias-Cartin, R.; Grimaldi, S.; Arnoux, P.; Guigliarelli, B.; Magalon, A. Cardiolipin binding in bacterial respiratory complexes: Structural and functional implications. Biochim. Biophys. Acta (BBA)-Bioenerg. 2012, 1817, 1937–1949. [Google Scholar] [CrossRef] [Green Version]
- Asseri, A.H.; Godoy-Hernandez, A.; Goojani, H.G.; Lill, H.; Sakamoto, J.; McMillan, D.G.G.; Bald, D. Cardiolipin enhances the enzymatic activity of cytochrome bd and cytochrome bo3 solubilized in dodecyl-maltoside. Sci. Rep. 2021, 11, 8006. [Google Scholar] [CrossRef]
- Arias-Cartin, R.; Grimaldi, S.; Pommier, J.; Lanciano, P.; Schaefer, C.; Arnoux, P.; Giordano, G.; Guigliarelli, B.; Magalon, A. Cardiolipin-based respiratory complex activation in bacteria. Proc. Natl. Acad. Sci. USA 2011, 108, 7781–7786. [Google Scholar] [CrossRef] [Green Version]
- Palsdottir, H.; Hunte, C. Lipids in membrane protein structures. Biochim. Biophys. Acta (BBA)-Bioenerg. 2004, 1666, 2–18. [Google Scholar] [CrossRef] [Green Version]
- Jormakka, M.; Tornroth, S.; Byrne, B.; Iwata, S. Molecular basis of proton motive force generation: Structure of formate dehydrogenase-N. Science 2002, 295, 1863–1868. [Google Scholar] [CrossRef] [Green Version]
- Shinzawa-Itoh, K.; Aoyama, H.; Muramoto, K.; Terada, H.; Kurauchi, T.; Tadehara, Y.; Yamasaki, A.; Sugimura, T.; Kurono, S.; Tsujimoto, K.; et al. Structures and physiological roles of 13 integral lipids of bovine heart cytochrome c oxidase. EMBO J. 2007, 26, 1713–1725. [Google Scholar] [CrossRef] [PubMed]
- Harrenga, A.; Michel, H. The cytochrome c oxidase from Paracoccus denitrificans does not change the metal center ligation upon reduction. J. Biol. Chem. 1999, 274, 33296–33299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, L.; Sharpe, M.A.; Garavito, R.M.; Ferguson-Miller, S. Conserved lipidbinding sites in membrane proteins: A focus on cytochrome c oxidase. Curr. Opin. Struct. Biol. 2007, 17, 444–450. [Google Scholar] [CrossRef] [Green Version]
- Aygun-Sunar, S.; Mandaci, S.; Koch, H.G.; Murray, I.V.; Goldfine, H.; Daldal, F. Ornithine lipid is required for optimal steady-state amounts of c-type cytochromes in Rhodobacter capsulatus. Mol. Microbiol. 2006, 61, 418–435. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Reimann, J.; Singh, L.M.; Adelroth, P. Substrate binding and the catalytic reactions in cbb3-type oxidases: The lipid membrane modulates ligand binding. Biochim. Biophys. Acta (BBA)-Bioenerg. 2010, 1797, 724–731. [Google Scholar] [CrossRef] [Green Version]
- Ekici, S.; Pawlik, G.; Lohmeyer, E.; Koch, H.G.; Daldal, F. Biogenesis of cbb3-type cytochrome c oxidase in Rhodobacter capsulatus. Biochim. Biophys. Acta (BBA)-Bioenerg. 2012, 1817, 898–910. [Google Scholar] [CrossRef] [Green Version]
- Okunaki, K.; Sekuzu, I.; Yonetani, T.; Takemori, S. Studies on cytochrome A. I. Extraction, purification and some properties of cytochrome A. J. Biochem. 1958, 45, 847–854. [Google Scholar]
- Yonetani, T.; Takemori, S.; Sekuzu, I.; Okunuki, K. Activation of cytochrome oxidase by synthetic detergents. Nature 1958, 181, 1339–1340. [Google Scholar] [CrossRef]
- Horie, S.; Morrison, M.; Bright, J. Cytochrome c Oxidase Components: I. Purification and properties. J. Biol. Chem. 1963, 238, 1855–1860. [Google Scholar] [CrossRef]
- Van Buuren, K.J.H.; Van Gelder, B.F. Biochemical and biophysical studies on cytochrome c oxidase. XIII. Effect of cholate on the enzymic activity. Biochim. Biophys. Acta (BBA)-Bioenerg. 1974, 333, 209–217. [Google Scholar] [CrossRef]
- Chuang, T.F.; Crane, F.L. Phospholipids in the cytochrome oxidase reaction. J. Bioenergetics 1973, 4, 563–578. [Google Scholar] [CrossRef]
- Rosevear, P.; VanAken, T.; Baxter, J.; Ferguson-Miller, S. Alkyl glycoside detergents: A simpler synthesis and their effects on kinetic and physical properties of cytochrome c oxidase. Biochemistry 1980, 19, 4108–4115. [Google Scholar] [CrossRef]
- Napiwotzki, J.; Shinzawa-Itoh, K.; Yoshikawa, S.; Kadenbach, B. ATP and ADP bind to cytochrome c oxidase and regulate its activity. Biol. Chem. 1997, 378, 1013–1021. [Google Scholar] [CrossRef]
- Musatov, A.; Robinson, N.C. Cholate-induced dimerization of detergent- or phospholipid-solubilized bovine cytochrome c oxidase. Biochemistry 2002, 41, 4371–4376. [Google Scholar] [CrossRef]
- Oleynikov, I.P.; Sudakov, R.V.; Radyukhin, V.A.; Arutyunyan, A.M.; Azarkina, N.V.; Vygodina, T.V. Interaction of Amphipathic Peptide from Influenza Virus M1 Protein with Mitochondrial Cytochrome Oxidase. Int. J. Mol. Sci. 2023, 24, 4119. [Google Scholar] [CrossRef]
- Soulimane, T.; Buse, G. Integral cytochrome-c oxidase. Preparation and progress towards a three-dimensional crystallization. Eur. J. Biochem. 1995, 227, 588–595. [Google Scholar] [CrossRef]
- Georgevich, G.; Darley-Usmar, V.M.; Malatesta, F.; Capaldi, R.A. Electron transfer in monomeric forms of beef and shark heart cytochrome c oxidase. Biochemistry 1983, 22, 1317–1322. [Google Scholar] [CrossRef]
- Robinson, N.C.; Talbert, L. Triton-X-100 induced dissociation of beef heart cytochrome c oxidase into monomers. Biochemistry 1986, 25, 2328–2335. [Google Scholar] [CrossRef]
- Sadoski, R.C.; Zaslavsky, D.; Gennis, R.B.; Durham, B.; Millett, F. Exposure of bovine cytochrome c oxidase to high triton X-100 or to alkaline conditions causes a dramatic change in the rate of reduction of compound F. J. Biol. Chem. 2001, 276, 33616–33620. [Google Scholar] [CrossRef] [Green Version]
- Oleynikov, I.P.; Azarkina, N.V.; Vygodina, T.V.; Konstantinov, A.A. Mechanism of inhibition of cytochrome c oxidase by Triton X-100. Biochemistry (Moscow) 2021, 86, 44–58. [Google Scholar] [CrossRef] [PubMed]
- Oleynikov, I.P.; Azarkina, N.V.; Vygodina, T.V.; Konstantinov, A.A. Interaction of cytochrome c oxidase with steroid hormones. Cells 2020, 9, 2211. [Google Scholar] [CrossRef] [PubMed]
- Oleynikov, I.P.; Sudakov, R.V.; Azarkina, N.V.; Vygodina, T.V. Direct Interaction of Mitochondrial Cytochrome c Oxidase with Thyroid Hormones: Evidence for Two Binding Sites. Cells 2022, 11, 908. [Google Scholar] [CrossRef] [PubMed]
- Giuffrè, A.; Forte, E.; Antonini, G.; D’Itri, E.; Brunori, M.; Soulimane, T.; Buse, G. Kinetic properties of ba3 oxidase from Thermus thermophilus: Effect of temperature. Biochemistry 1999, 38, 1057–1065. [Google Scholar] [CrossRef] [PubMed]
- Urbani, A.; Gemeinhardt, S.; Warne, A.; Saraste, M. Properties of the detergent solubilised cytochrome c oxidase (cytochrome cbb3) purified from Pseudomonas stutzeri. FEBS Lett. 2001, 508, 29–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, M.J.; Gennis, R.B. Purification and reconstitution of the cytochrome d terminal oxidase complex from Escherichia coli. Meth. Enzymol. 1986, 126, 87–94. [Google Scholar]
- Borisov, V.B. Effect of membrane environment on ligand-binding properties of the terminal oxidase cytochrome bd-I from Escherichia coli. Biochemistry 2020, 85, 1603–1612. [Google Scholar] [CrossRef]
- Varhac, R.; Tomaskova, N.; Fabian, M.; Sedlak, E. Kinetics of cyanide binding as a probe of local stability/flexibility of cytochrome c. Biophys. Chem. 2009, 144, 21–26. [Google Scholar] [CrossRef]
- George, P.; Schejter, A. The reactivity of ferrocytochrome c with iron-binding ligands. J. Biol. Chem. 1964, 239, 1504–1508. [Google Scholar] [CrossRef]
- Ferguson-Miller, S.; Hiser, C.; Liu, J. Gating and regulation of the cytochrome c oxidase proton pump. Biochim. Biophys. Acta (BBA)-Bioenerg. 2012, 1817, 489–494. [Google Scholar] [CrossRef] [Green Version]
- Hiser, C.; Liu, J.; Ferguson-Miller, S. The K-path entrance in cytochrome c oxidase is defined by mutation of E101 and controlled by an adjacent ligand binding domain. Biochim. Biophys. Acta (BBA)-Bioenerg. 2018, 1859, 725–733. [Google Scholar] [CrossRef] [PubMed]
- Qin, L.; Mills, D.A.; Hiser, C.; Murphree, A.; Garavito, R.M.; Ferguson-Miller, S.; Hosler, J. Crystallographic location and mutational analysis of Zn and Cd inhibitory sites and role of lipidic carboxylates in rescuing proton path mutants in cytochrome c oxidase. Biochemistry 2007, 46, 6239–6248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fetter, J.; Sharpe, M.; Qian, J.; Mills, D.; Ferguson-Miller, S.; Nicholls, P. Fatty acids stimulate activity and restore respiratory control in a proton channel mutant of cytochrome c oxidase. FEBS Lett. 1996, 393, 155–160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smirnova, I.A.; Adelroth, P.; Gennis, R.B.; Brzezinski, P. Aspartate-132 in cytochrome c oxidase from Rhodobacter sphaeroides is involved in a two-step proton transfer during oxo-ferryl formation. Biochemistry 1999, 38, 6826–6833. [Google Scholar] [CrossRef] [PubMed]
- Checover, S.; Nachliel, E.; Dencher, N.A.; Gutman, M. Mechanism of proton entry into the cytoplasmic section of the proton-conducting channel of bacteriorhodopsin. Biochemistry 1997, 36, 13919–13928. [Google Scholar] [CrossRef] [PubMed]
- Vygodina, T.V.; Pecoraro, C.; Mitchell, D.; Gennis, R.; Konstantinov, A.A. Mechanism of inhibition of electron transfer by amino acid replacement K362M in a proton channel of Rhodobacter sphaeroides cytochrome c oxidase. Biochemistry 1998, 37, 3053–3061. [Google Scholar] [CrossRef] [PubMed]
- Konstantinov, A. Cytochrome c oxidase as a proton-pumping peroxidase: Reaction cycle and electrogenic mechanism. J. Bioenerg. Biomembr. 1998, 30, 121–130. [Google Scholar] [CrossRef] [PubMed]
- Goglia, F.; Lanni, A.; Barth, J.; Kadenbach, B. Interaction of diiodothyronines with isolated cytochromecoxidase. FEBS Lett. 1994, 346, 295–298. [Google Scholar] [PubMed] [Green Version]
- Arnold, S.; Goglia, F.; Kadenbach, B. 3,5-Diiodothyronine binds to subunit Va of cytochrome-c oxidase and abolishes the allosteric inhibition of respiration by ATP. JBIC J. Biol. Inorg. Chem. 1998, 252, 325–330. [Google Scholar] [CrossRef] [PubMed]
- Antalik, M.; Jancura, D.; Palmer, G.; Fabian, M. A role for the protein in internal electron transfer to the catalytic center of cytochrome c oxidase. Biochemistry 2005, 44, 14881–14889. [Google Scholar] [CrossRef] [PubMed]
- Madsen, L.L.; Korsgaard, B.; Bjerregaard, P. Estrogenic effects in flounder Platichthys flesus orally exposed to 4-tert-octylphenol. Aquatic Toxicol. 2003, 64, 393–405. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, R.H.; Pelletier, J.H.; Tupper, E.J.; Hutchinson, L.M.; Gosse, J.A. Estrogen mimetic 4-tert-octylphenol enhances IgE-mediated degranulation of RBL-2H3 mast cells. J. Toxicol. Envir. Health 2012, 75, 1451–1455. [Google Scholar] [CrossRef] [PubMed]
- Landolt-Marticorena, C.; Williams, K.A.; Deber, C.M.; Reithmeier, R.A. Non-random distribution of amino acids in the transmembrane segments of human type I single span membrane proteins. J. Mol. Biol. 1993, 229, 602–608. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Azarkina, N.V.; Borisov, V.B.; Oleynikov, I.P.; Sudakov, R.V.; Vygodina, T.V. Interaction of Terminal Oxidases with Amphipathic Molecules. Int. J. Mol. Sci. 2023, 24, 6428. https://doi.org/10.3390/ijms24076428
Azarkina NV, Borisov VB, Oleynikov IP, Sudakov RV, Vygodina TV. Interaction of Terminal Oxidases with Amphipathic Molecules. International Journal of Molecular Sciences. 2023; 24(7):6428. https://doi.org/10.3390/ijms24076428
Chicago/Turabian StyleAzarkina, Natalia V., Vitaliy B. Borisov, Ilya P. Oleynikov, Roman V. Sudakov, and Tatiana V. Vygodina. 2023. "Interaction of Terminal Oxidases with Amphipathic Molecules" International Journal of Molecular Sciences 24, no. 7: 6428. https://doi.org/10.3390/ijms24076428