
The Role of SCL Isoforms in Embryonic Hematopoiesis
 , and
, and
Abstract
:1. Introduction
2. Results
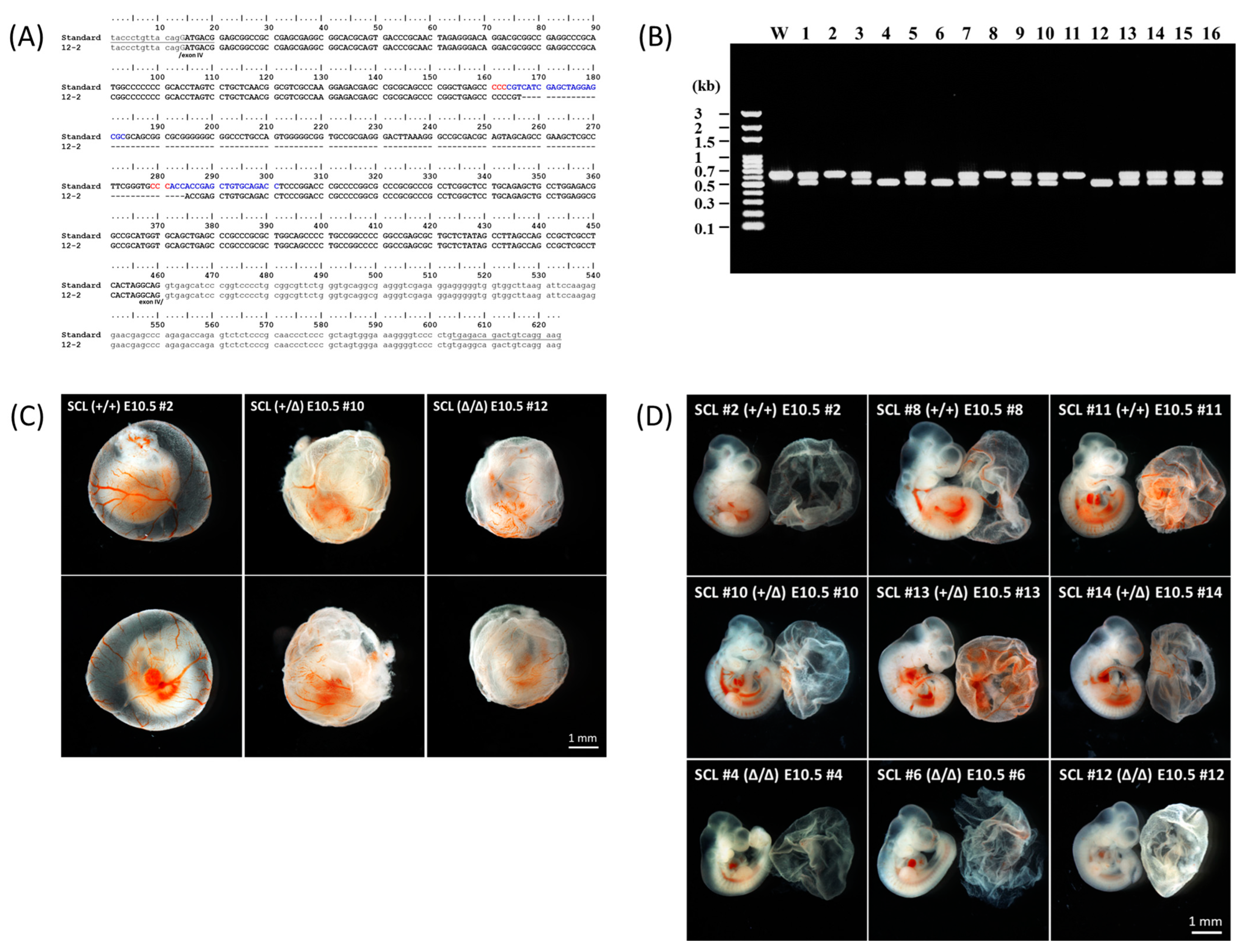
2.1. Preparation of the Gene Edited Mice
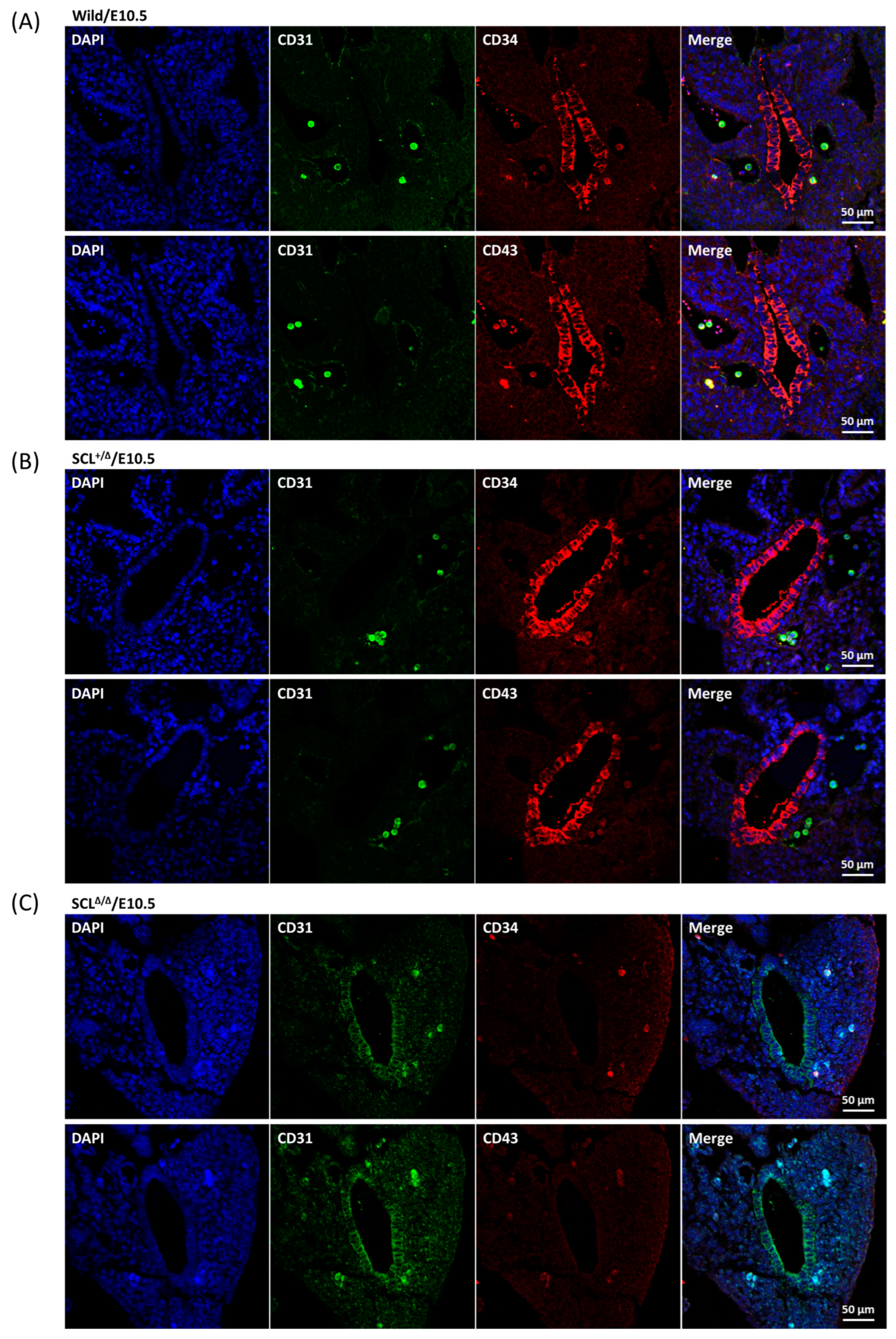
2.2. Characterization of the Gene Edited Murine Fetuses
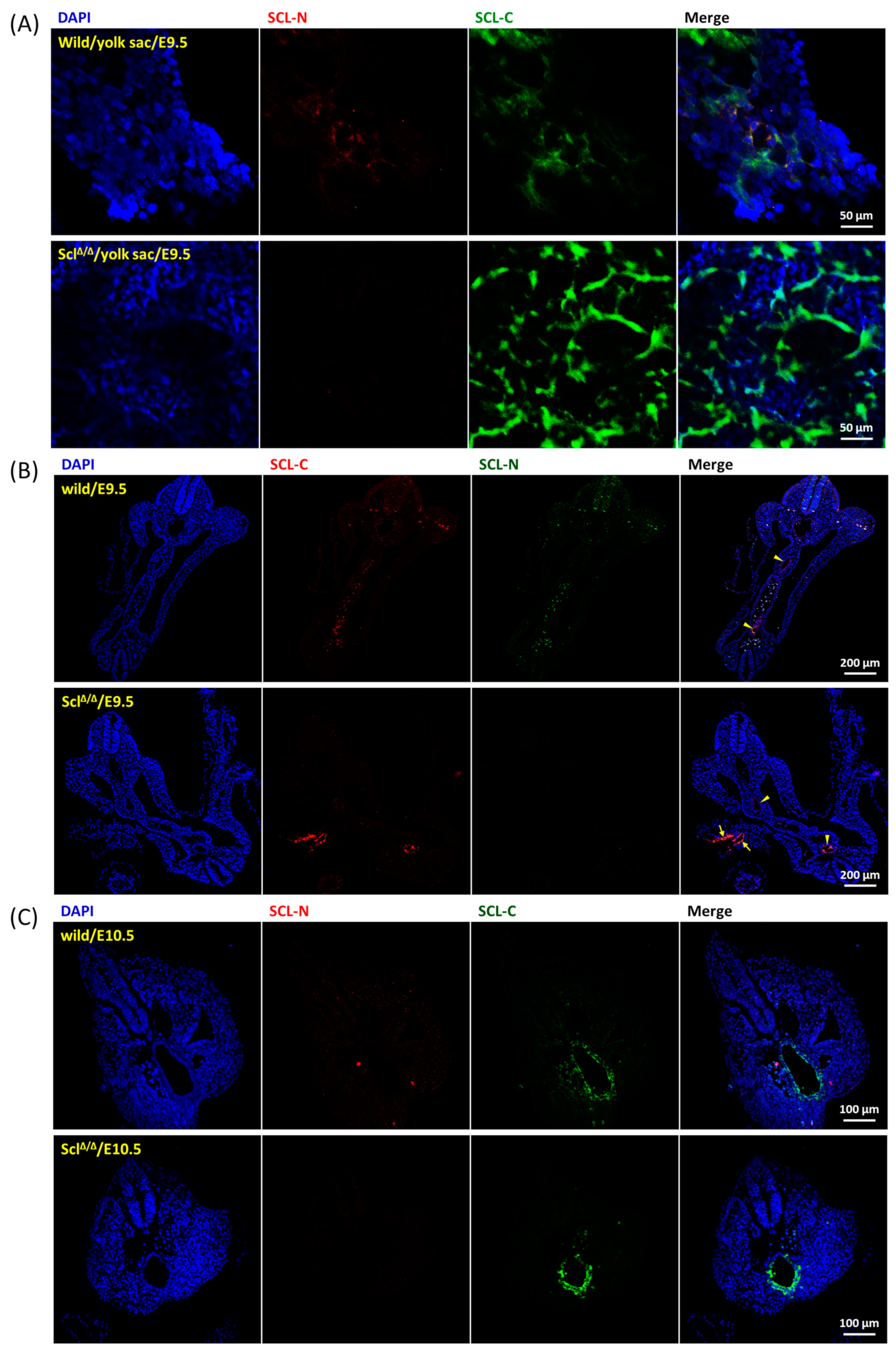
2.3. Expression Pattern of SCL Isoforms
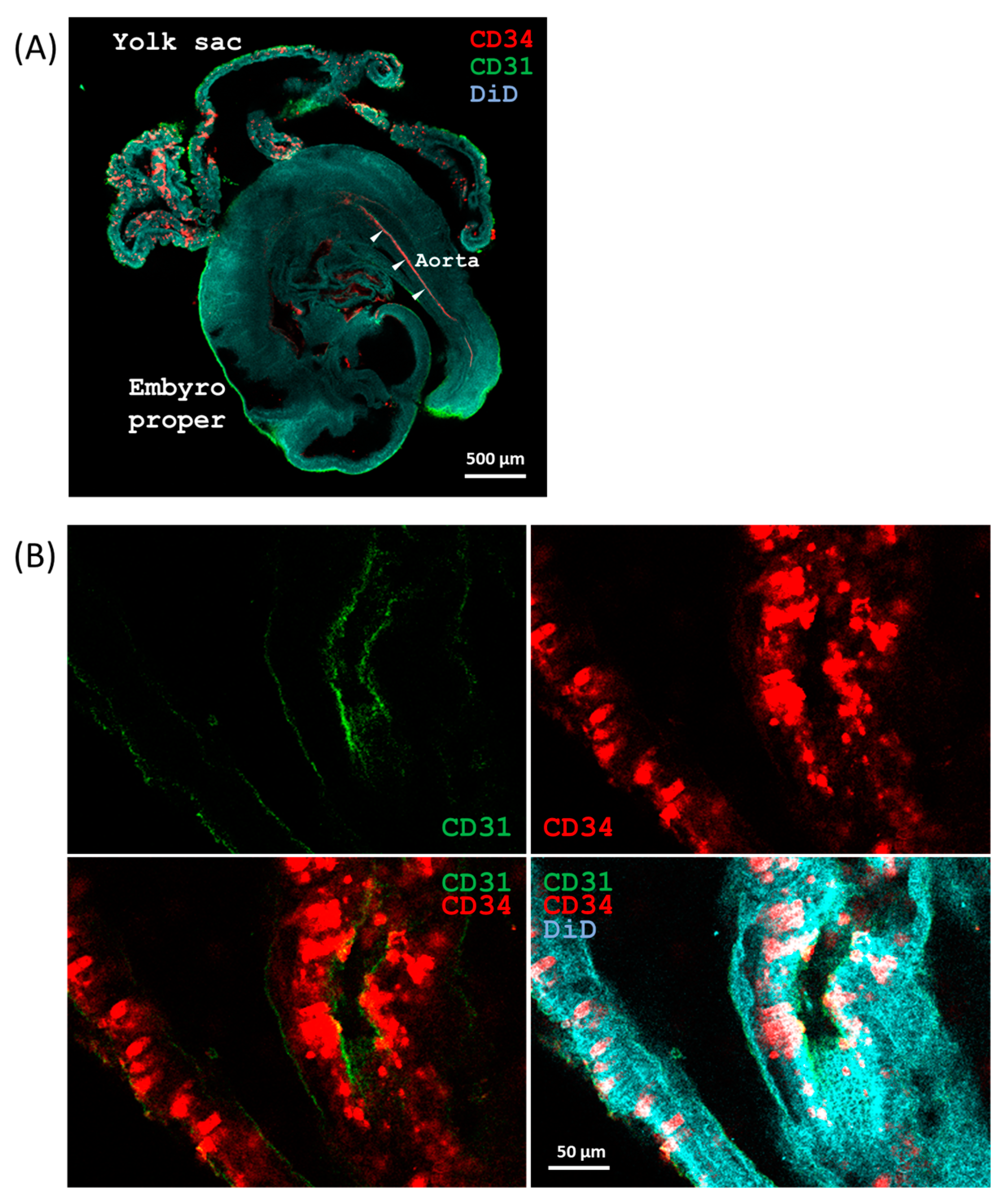
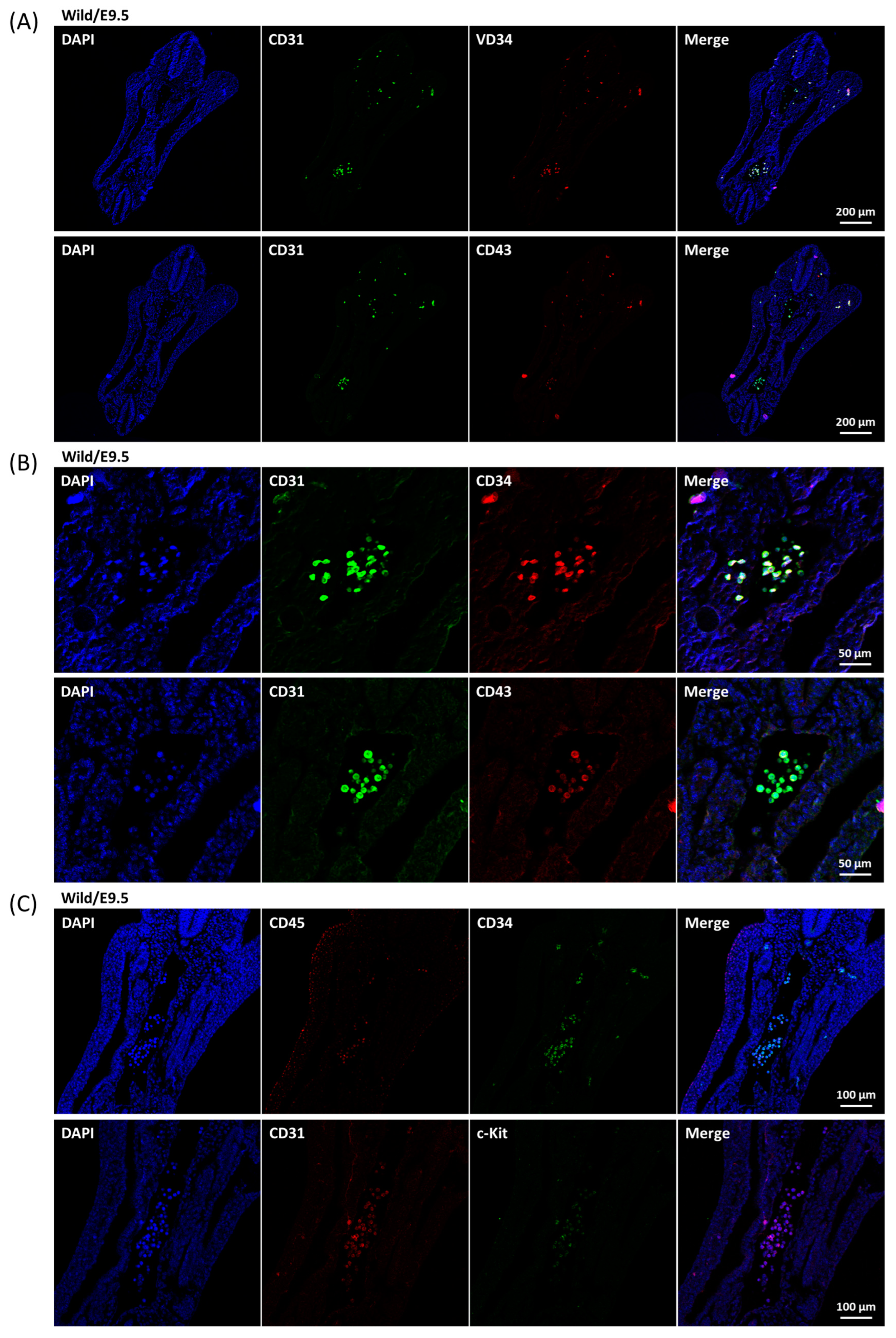
2.4. Development of Aortic HECs
3. Discussion
4. Materials and Methods
4.1. Generation of SCL Gene Edited Mice by RNA Micro-Injection
4.2. Genotyping
4.3. Whole-Mount Immunofluorescence Staining
4.4. Embryo Section and Immunofluorescence Staining
5. Concluding Remarks
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Palis, J.; Malik, J.; Mcgrath, K.E.; Kingsley, P.D. Primitive erythropoiesis in the mammalian embryo. Int. J. Dev. Biol. 2010, 54, 1011–1018. [Google Scholar] [CrossRef] [Green Version]
- Yamane, T. Mouse yolk sac hematopoiesis. Front. Cell Dev. Biol. 2018, 6, 80. [Google Scholar] [CrossRef] [Green Version]
- McGrath, K.E.; Koniski, A.D.; Malik, J.; Palis, J. Circulation is established in a stepwise pattern in the mammalian embryo. Blood J. Am. Soc. Hematol. 2003, 101, 1669–1675. [Google Scholar] [CrossRef] [Green Version]
- Garcia, M.D.; Larina, I.V. Vascular development and hemodynamic force in the mouse yolk sac. Front. Physiol. 2014, 5, 308. [Google Scholar] [CrossRef] [Green Version]
- Ji, R.P.; Phoon, C.K.; Aristizábal, O.; McGrath, K.E.; Palis, J.; Turnbull, D.H. Onset of cardiac function during early mouse embryogenesis coincides with entry of primitive erythroblasts into the embryo proper. Circ. Res. 2003, 92, 133–135. [Google Scholar] [CrossRef]
- Lucitti, J.L.; Jones, E.A.; Huang, C.; Chen, J.; Fraser, S.E.; Dickinson, M.E. Vascular remodeling of the mouse yolk sac requires hemodynamic force. Development 2007, 134, 3317–3326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boisset, J.-C.; van Cappellen, W.; Andrieu-Soler, C.; Galjart, N.; Dzierzak, E.; Robin, C. In vivo imaging of haematopoietic cells emerging from the mouse aortic endothelium. Nature 2010, 464, 116–120. [Google Scholar] [CrossRef]
- Boisset, J.-C.; Andrieu-Soler, C.; Van Cappellen, W.A.; Clapes, T.; Robin, C. Ex vivo time-lapse confocal imaging of the mouse embryo aorta. Nat. Protoc. 2011, 6, 1792–1805. [Google Scholar] [CrossRef] [PubMed]
- de Bruijn, M.F.; Speck, N.A.; Peeters, M.C.; Dzierzak, E. Definitive hematopoietic stem cells first develop within the major arterial regions of the mouse embryo. EMBO J. 2000, 19, 2465–2474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Bruijn, M.F.; Ma, X.; Robin, C.; Ottersbach, K.; Sanchez, M.-J.; Dzierzak, E. Hematopoietic stem cells localize to the endothelial cell layer in the midgestation mouse aorta. Immunity 2002, 16, 673–683. [Google Scholar] [CrossRef] [Green Version]
- Taylor, E.; Taoudi, S.; Medvinsky, A. Hematopoietic stem cell activity in the aorta-gonad-mesonephros region enhances after mid-day 11 of mouse development. Int. J. Dev. Biol. 2010, 54, 1055–1060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ema, H.; Nakauchi, H. Expansion of hematopoietic stem cells in the developing liver of a mouse embryo. Blood J. Am. Soc. Hematol. 2000, 95, 2284–2288. [Google Scholar]
- Christensen, J.L.; Wright, D.E.; Wagers, A.J.; Weissman, I.L. Circulation and chemotaxis of fetal hematopoietic stem cells. PLoS Biol. 2004, 2, e75. [Google Scholar] [CrossRef] [PubMed]
- Ferdous, A.; Caprioli, A.; Iacovino, M.; Martin, C.M.; Morris, J.; Richardson, J.A.; Latif, S.; Hammer, R.E.; Harvey, R.P.; Olson, E.N. Nkx2–5 transactivates the Ets-related protein 71 gene and specifies an endothelial/endocardial fate in the developing embryo. Proc. Natl. Acad. Sci. USA 2009, 106, 814–819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robb, L.; Lyons, I.; Li, R.; Hartley, L.; Köntgen, F.; Harvey, R.P.; Metcalf, D.; Begley, C.G. Absence of yolk sac hematopoiesis from mice with a targeted disruption of the scl gene. Proc. Natl. Acad. Sci. USA 1995, 92, 7075–7079. [Google Scholar] [CrossRef] [Green Version]
- Shivdasani, R.A.; Mayer, E.L.; Orkin, S.H. Absence of blood formation in mice lacking the T-cell leukaemia oncoprotein tal-1/SCL. Nature 1995, 373, 432–434. [Google Scholar] [CrossRef]
- Qian, F.; Zhen, F.; Xu, J.; Huang, M.; Li, W.; Wen, Z. Distinct functions for different scl isoforms in zebrafish primitive and definitive hematopoiesis. PLoS Biol. 2007, 5, e132. [Google Scholar] [CrossRef]
- Ren, X.; Gomez, G.A.; Zhang, B.; Lin, S. Scl isoforms act downstream of etsrp to specify angioblasts and definitive hematopoietic stem cells. Blood J. Am. Soc. Hematol. 2010, 115, 5338–5346. [Google Scholar] [CrossRef] [Green Version]
- Zhen, F.; Lan, Y.; Yan, B.; Zhang, W.; Wen, Z. Hemogenic endothelium specification and hematopoietic stem cell maintenance employ distinct Scl isoforms. Development 2013, 140, 3977–3985. [Google Scholar] [CrossRef] [Green Version]
- Galloway, J.L.; Zon, L.I. 3 ontogeny of hematopoiesis: Examining the emergence of hematopoietic cells in the vertebrate embryo. In Current Topics in Developmental Biology; Academic Press: Cambridge, MA, USA, 2003. [Google Scholar]
- Begley, C.; Robb, L.; Rockman, S.; Visvader, J.; Bockamp, E.; Chan, Y.; Green, A. Structure of the gene encoding the murine SCL protein. Gene 1994, 138, 93–99. [Google Scholar] [CrossRef]
- Bockamp, E.; McLaughlin, F.; Murrell, A.; Gottgens, B.; Robb, L.; Begley, C.; Green, A. Lineage-restricted regulation of the murine SCL/TAL-1 promoter. Blood 1995, 86, 1502–1514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bockamp, E.-O.; McLaughlin, F.; Göttgens, B.; Murrell, A.M.; Elefanty, A.G.; Green, A.R. Distinct mechanisms direct SCL/tal-1 expression in erythroid cells and CD34 positive primitive myeloid cells. J. Biol. Chem. 1997, 272, 8781–8790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tu, C.-F.; Chuang, C.-k.; Hsiao, K.-H.; Chen, C.-H.; Chen, C.-M.; Peng, S.-H.; Su, Y.-H.; Chiou, M.-T.; Yen, C.-H.; Hung, S.-W. Lessening of porcine epidemic diarrhoea virus susceptibility in piglets after editing of the CMP-N-glycolylneuraminic acid hydroxylase gene with CRISPR/Cas9 to nullify N-glycolylneuraminic acid expression. PLoS ONE 2019, 14, e0217236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chuang, C.-K.; Lin, W.-M. Points of View on the Tools for Genome/Gene Editing. Int. J. Mol. Sci. 2021, 22, 9872. [Google Scholar] [CrossRef] [PubMed]
- Asmamaw, M.; Zawdie, B. Mechanism and applications of CRISPR/Cas-9-mediated genome editing. Biol. Targets Ther. 2021, 15, 353–361. [Google Scholar]
- Hung, S.-W.; Chuang, C.-k.; Wong, C.-H.; Yen, C.-H.; Peng, S.-H.; Yang, C.; Chen, M.-C.; Yang, T.-S.; Tu, C.-F. Activated macrophages of CD 163 gene edited pigs generated by direct cytoplasmic microinjection with CRISPR gRNA/Cas9 mRNA are resistant to PRRS virus assault. Anim. Biotechnol. 2022, 1–14. [Google Scholar] [CrossRef]
- Canu, G.; Ruhrberg, C. First blood: The endothelial origins of hematopoietic progenitors. Angiogenesis 2021, 24, 199–211. [Google Scholar] [CrossRef]
- Wu, Y.; Hirschi, K.K. Regulation of hemogenic endothelial cell development and function. Annu. Rev. Physiol. 2021, 83, 17–37. [Google Scholar] [CrossRef]
- Lange, L.; Morgan, M.; Schambach, A. The hemogenic endothelium: A critical source for the generation of PSC-derived hematopoietic stem and progenitor cells. Cell. Mol. Life Sci. 2021, 78, 4143–4160. [Google Scholar] [CrossRef]
- Vodyanik, M.A.; Thomson, J.A.; Slukvin, I.I. Leukosialin (CD43) defines hematopoietic progenitors in human embryonic stem cell differentiation cultures. Blood 2006, 108, 2095–2105. [Google Scholar] [CrossRef] [Green Version]
- Demirci, S.; Haro-Mora, J.J.; Leonard, A.; Drysdale, C.; Malide, D.; Keyvanfar, K.; Essawi, K.; Vizcardo, R.; Tamaoki, N.; Restifo, N.P. Definitive hematopoietic stem/progenitor cells from human embryonic stem cells through serum/feeder-free organoid-induced differentiation. Stem Cell Res. Ther. 2020, 11, 1–14. [Google Scholar]
- Batsivari, A.; Rybtsov, S.; Souilhol, C.; Binagui-Casas, A.; Hills, D.; Zhao, S.; Travers, P.; Medvinsky, A. Understanding hematopoietic stem cell development through functional correlation of their proliferative status with the intra-aortic cluster architecture. Stem Cell Rep. 2017, 8, 1549–1562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plein, A.; Fantin, A.; Denti, L.; Pollard, J.W.; Ruhrberg, C. Erythro-myeloid progenitors contribute endothelial cells to blood vessels. Nature 2018, 562, 223–228. [Google Scholar] [CrossRef]
- Lahlil, R.; Lécuyer, E.; Herblot, S.; Hoang, T. SCL assembles a multifactorial complex that determines glycophorin A expression. Mol. Cell. Biol. 2004, 24, 1439–1452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El Omari, K.; Hoosdally, S.J.; Tuladhar, K.; Karia, D.; Vyas, P.; Patient, R.; Porcher, C.; Mancini, E.J. Structure of the leukemia oncogene LMO2: Implications for the assembly of a hematopoietic transcription factor complex. Blood J. Am. Soc. Hematol. 2011, 117, 2146–2156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uversky, V.N. Intrinsically disordered proteins and their “mysterious” (meta)physics. Front. Phys. 2019, 7, 10. [Google Scholar] [CrossRef] [Green Version]
- Trivedi, R.; Nagarajaram, H.A. Intrinsically Disordered Proteins: An Overview. Int. J. Mol. Sci. 2022, 23, 14050. [Google Scholar] [CrossRef]
- Liu, J.; Perumal, N.B.; Oldfield, C.J.; Su, E.W.; Uversky, V.N.; Dunker, A.K. Intrinsic disorder in transcription factors. Biochemistry 2006, 45, 6873–6888. [Google Scholar] [CrossRef] [Green Version]
- Minezaki, Y.; Homma, K.; Kinjo, A.R.; Nishikawa, K. Human transcription factors contain a high fraction of intrinsically disordered regions essential for transcriptional regulation. J. Mol. Biol. 2006, 359, 1137–1149. [Google Scholar] [CrossRef]
- Staby, L.; O’Shea, C.; Willemoës, M.; Theisen, F.; Kragelund, B.B.; Skriver, K. Eukaryotic transcription factors: Paradigms of protein intrinsic disorder. Biochem. J. 2017, 474, 2509–2532. [Google Scholar] [CrossRef] [Green Version]
- Bondos, S.E.; Dunker, A.K.; Uversky, V.N. On the roles of intrinsically disordered proteins and regions in cell communication and signaling. Cell Commun. Signal. 2021, 19, 88. [Google Scholar] [CrossRef] [PubMed]
- Morris, O.M.; Torpey, J.H.; Isaacson, R.L. Intrinsically disordered proteins: Modes of binding with emphasis on disordered domains. Open Biol. 2021, 11, 210222. [Google Scholar] [CrossRef]
- Chakrabarti, P.; Chakravarty, D. Intrinsically disordered proteins/regions and insight into their biomolecular interactions. Biophys. Chem. 2022, 283, 106769. [Google Scholar] [CrossRef] [PubMed]
- Brodsky, S.; Jana, T.; Mittelman, K.; Chapal, M.; Kumar, D.K.; Carmi, M.; Barkai, N. Intrinsically disordered regions direct transcription factor in vivo binding specificity. Mol. Cell 2020, 79, 459–471.e454. [Google Scholar] [CrossRef]
- Brodsky, S.; Jana, T.; Barkai, N. Order through disorder: The role of intrinsically disordered regions in transcription factor binding specificity. Curr. Opin. Struct. Biol. 2021, 71, 110–115. [Google Scholar] [CrossRef] [PubMed]
- Chiang, A.-S.; Lin, W.-Y.; Liu, H.-P.; Pszczolkowski, M.A.; Fu, T.-F.; Chiu, S.-L.; Holbrook, G.L. Insect NMDA receptors mediate juvenile hormone biosynthesis. Proc. Natl. Acad. Sci. USA 2002, 99, 37–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | DNA Sequence |
|---|---|
| SclF1 | TACCCTGTTACAGGATGACG |
| SclR1 | CTTCCTGACAGTCTGTCTCA |
| SclSpF1 | CGTC GCGCTCCTAGCTCGATGACG GTTTTAGAGCTAGAAAT |
| SclSpR1 | TGCTATTTCTAGCTCTAAAAC CGTCATCGAGCTAGGAGCGC |
| SclSpF2 | CGTC GGTCTGCACAGCTCGGTGGT GTTTTAGAGCTAGAAAT |
| SclSpR2 | TGCTATTTCTAGCTCTAAAAC ACCACCGAGCTGTGCAGACC |
| Antibodies | Species/Isotype | Brand | Cat. No. |
|---|---|---|---|
| Anti-Scl (N-terminus) | Rabbit polyclonal antibody | LSBio | LS-B14706 |
| Anti-Scl (C-terminus) | Mouse monoclonal antibody (IgG1, κ) | Santa Cruz | sc-393287 |
| CD31 | Rat monoclonal antibody (IgG2a, κ) | BioCare Medical | CM 303 |
| CD31 | Rabbit polyclonal antibody | GeneTex | GTX130274 |
| CD34 | Mouse monoclonal antibody (IgG1, κ) | Santa Cruz | sc-74499 |
| CD43 | Mouse monoclonal antibody (IgG1, κ) [DF-T1] | GeneTex | GTX73633 |
| c-Kit | Rat monoclonal antibody (IgG2b) | Santa Cruz | sc-19619 |
| CD45 (H-230) | Rabbit polyclonal antibody | Santa Cruz | sc-25590 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chuang, C.-K.; Chen, S.-F.; Su, Y.-H.; Chen, W.-H.; Lin, W.-M.; Wang, I.-C.; Shyue, S.-K. The Role of SCL Isoforms in Embryonic Hematopoiesis. Int. J. Mol. Sci. 2023, 24, 6427. https://doi.org/10.3390/ijms24076427
Chuang C-K, Chen S-F, Su Y-H, Chen W-H, Lin W-M, Wang I-C, Shyue S-K. The Role of SCL Isoforms in Embryonic Hematopoiesis. International Journal of Molecular Sciences. 2023; 24(7):6427. https://doi.org/10.3390/ijms24076427
Chicago/Turabian StyleChuang, Chin-Kai, Su-Fen Chen, Yu-Hsiu Su, Wei-Hsin Chen, Wei-Ming Lin, I-Ching Wang, and Song-Kun Shyue. 2023. "The Role of SCL Isoforms in Embryonic Hematopoiesis" International Journal of Molecular Sciences 24, no. 7: 6427. https://doi.org/10.3390/ijms24076427