
Postbiotics against Obesity: Perception and Overview Based on Pre-Clinical and Clinical Studies

Abstract
:1. Introduction
2. Probiotics
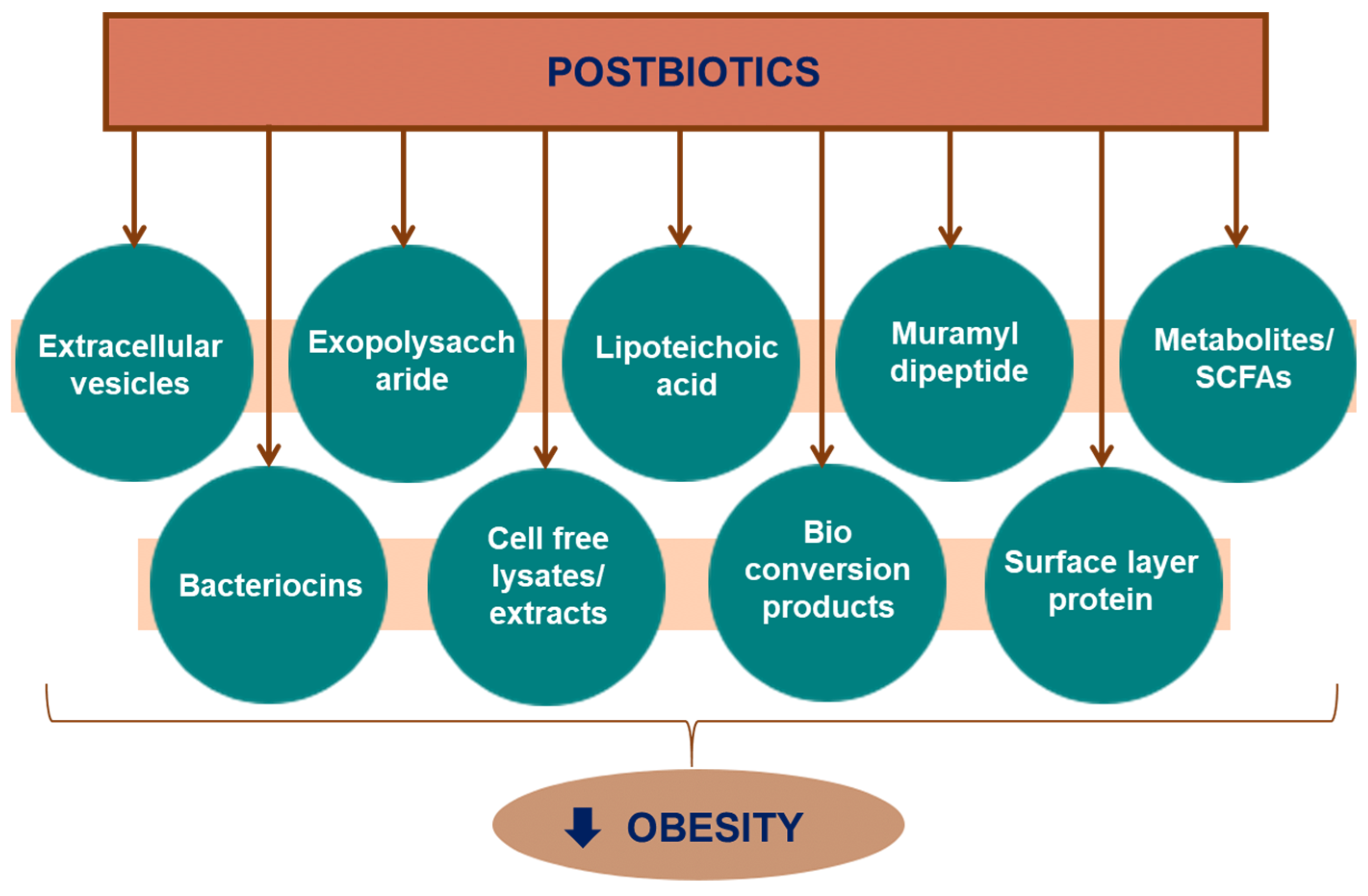
3. Postbiotics
Production and Characterisation Approaches
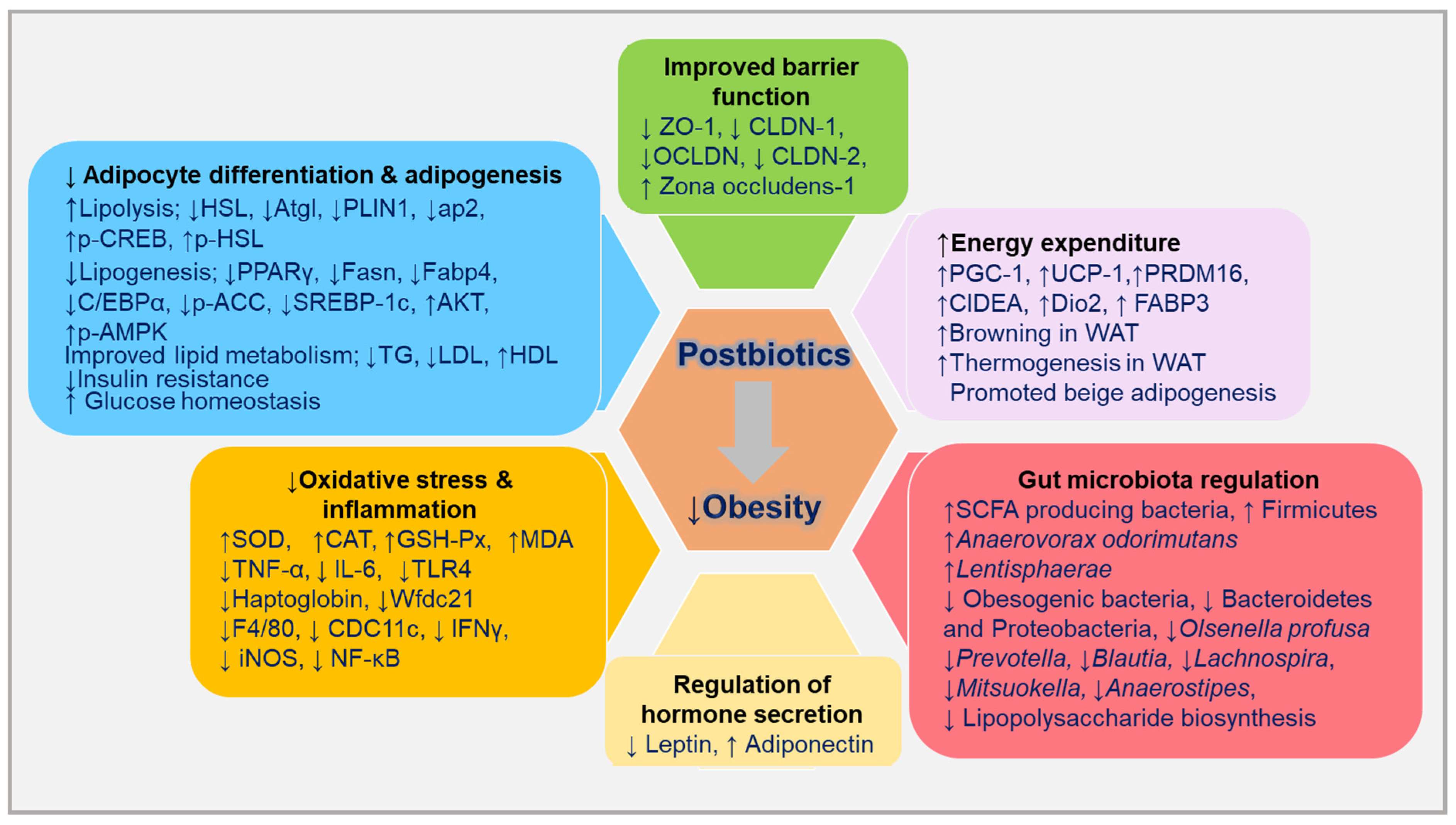
4. Postbiotics and Anti-Obesity Mechanisms
4.1. Cell and Animal Modal Studies
4.1.1. Cell Wall Components
Muramyl Dipeptide
Surface Layer Proteins
Lipoteichoic Acid
Exopolysaccharide
Surface Layer Protein, Exopolysaccharide, and Prebiotics
4.1.2. Biotransformation Products
4.1.3. Cell-Free Extracts
4.1.4. Extracellular Vesicles
4.1.5. Metabolites
4.1.6. Bacteriocins
4.1.7. Cell-Free Lysates

{kind=link}
{kind=link}
{kind=link}
| Source | Postbiotic | Model | Treatment/Duration | Biological Effects | Reference |
|---|---|---|---|---|---|
| Muramyl dipeptide | |||||
| Commercial | MDP | Mouse: wild type (WT) C57BL/6J, Irf4−/− mice, hepatocyte-specific NOD2−/− mice, and leptin-deficient (ob/ob) mice | (a) MDP 100 µg; 4 days/week, 5 weeks), + HFD. (b) HFD, 10 weeks + daily injection of MDP, 3 days + final MDP injection before 24 h of experiments | ↓Obesity, ↓hepatic insulin resistance, ↓fat inflammation Upregulated NOD2-IRF4 pathway | Cavallari et al. [63] |
| Commercial | MDP | Male and female: Adipocyte-specific Irf4−/− (IRF4 knockout mice (AdipoIRF4fl/fl) and control without adiponectin-cre transgene (WTfl/fl), 3 days before LPS injection + 6 h fasting | Acute endotoxaemia experiments: MDP 100 μg, 3 days, 4th day, LPS (i.p., ultrapure) 0.2 mg/kg, 6 h prior to GTT. Diet-induced obesity (a) Standard chow diet (b) Treatment group (60% kcal fat diet) + MDP (100 μg) 4 days/week, GTT performed At 5th week, eWAT collected | MDP: For blood glucose-lowering effects during endotoxaemia and HFD-fed male mice, adipocyte IRF4 was essential. No alteration in glucose level in endotoxaemia in female mice HFD-fed female mice had lower blood glucose level than the control. Inflammatory markers: ↓TNF, ↓CCL2, ↓CXCL10, ↓CD8, ↓IL-1β, ↓IL-10, ↓IL-6, ↓IFNγ, ↓CD4, ↓NOS2, and ↓NLRP3 in male and female mice in both groups | Duggan et al. [65] |
| S-layer protein | |||||
| Lentilactobacillus kefiri (DH1 and DH5 (SDH1 and SDH5) and Leuconostoc mesenteroides DH1606, DH1608, and DH1609 (LCM6, LCM8, and LCM9) | SLPs | RAW 264.7 cells | LPS (0.1 μg/mL), SLPs (10 or 20 μg/mL), 24 h at 37 °C | ↓IL-6 in LCM8, LCM9 and SDH5 ↓NF-κB p65 with SLCM9 | Kim E. et al. [68] |
| C57BL/6J (n = 10/group) | (a) Control group (saline) (b) Treatment HFD-SDH5 group (SLP from DH5 [SDH5] 120 mg/kg BW), and HFD-SLCM8 group (SLP from LCM8 120 mg/kg BW), 6 weeks | In SDH5 and SLCM8: ↓body weight gains and adipose tissue weight, ↓plasma TG, ↓insulin In SDH5: ↓TC, ↓LDL-C In SLCM8: adipocyte differentiation: ↓KLF8, acute phase response: ↓TRDN, ↓LBP, adipogenesis: ↓ADAM23, autophagy: ↓MAP1a, ↓Atp6v0d2, immune response: ↓ADAM8, ↓SLAMF7, ↓DCSTAMP, ↓MARC1, ↓UBD, ↓TREML4, ↓DOCK8, Inflammatory response: ↓EAR12, ↓HORMAD2, ↓NCAN, ↓Gpr50, ↓LIPF, ↓OXTR, ↓TREM2, ↓RGS1, ↓Tm4sf19, ↓Sfrp5 Upregulated gene expression ↑SH2B2, ↑MOGAT1, ↑FCNA, ↑EBF 2, ↑GPX3, ↑KLHL2, ↑CCR8, ↑CES1f/2c, ↑C2 | |||
| Lactobacillus curvatus (HKLC), and Lactobacillus plantarum (HKLP) | SLPs and heat-killed LAB | 3T3-L1 preadipocytes | SLPs (LC and LP) 7.5 μg/mL, 48 h | Inhibited lipid accumulation: LPSLP (19%), LCSLP (24%), and LPCSLP (24%), ↓FABP4, ↓PPARγ,. Induced apoptosis: ↓BCL-2, ↑caspase 3, HKLC, HKLP, and HKLPC: ↓FABP4 | Yoon et al. [69] |
| Lipoteichoic acid | |||||
| Bifidobacterium animalis subsp. lactis BPL1 (CECT 8145) | LTAs | Caenorhabditis elegans | LTA BPL1-0.1, 1.0, 10, 20, 50 μg/mL | ↓Fat content in nematodes ↓TG through involvement of IGF-1 pathway | Balaguer et al. [75] |
| EPS | |||||
| Lactobacillus rhamnosus GG | EPS | 3T3-L1 cells | 10.0 μg/mL, 6 days | ↓ Intracellular TAG, Genes of adipose differentiation and adipogenesis: ↓PPARγ, ↓SCD1, ↓ap2, ↓DGAT1 ↓FAS, ↓LPL | Zhang et al. [81] |
| C57BL/6J mice (n = 6) |
| Significant ↓fat mass: subcutaneous, gonadal, and pararenal, ↓serum TAG levels, ↓TAG and cholesterol ester in liver, ↓PPARγ, ↓ap2, ↓FAS, ↓SCD1, ↓LPL, ↓DGAT1. | |||
| Lactobacillus plantarum L-14 | EPS | 3T3-L1 cells and hBM-MSCs | 50 and 100 μg/mL | L-14 extract inhibited differentiation of 3T3-L1 cells and hBM-MSCs. ↓TAG storage in mature adipocytes ↓PPARγ, ↓C/EBPα, ↓FAS, ↓LPL, ↓CD36, ↓GPDH, EPS: ↑AMPK pathway ↓PPARγ, ↓C/EBPα, ↑adiponectin, ↓p-ACC, ↓SREBP-1c, ↑AKT, ↑p-AMPKα | Lee et al. [82] |
| Surface layer protein, exopolysaccharide, and prebiotics | |||||
| Kefir LAB: Leuconostoc mesenteroides DH 1606 (LCM6) and L. mesenteroides DH 1608 (LCM8) | EPS S-layer protein | C57BL/6 mice (n = 120) |
| ↓Body weight gain: SLP (20%) and all (23%) ↓Adipose tissue weight: SLP (22%) and all (24%). ↓Plasma TG EPS (16%), SLP (31%), 2% GSF (18%), and all (34%) EPS: ↓LDL-C (34%), ↓TC/HDL ratio (18%), and ↑HDL-C (10%) ↓ Area under the curve (AUC) for insulin response: SLP (22%), 2% GSF (14%), and all (23%) ↓Glucose response (AUC): all (14%) Acute-phase response: ↓TRDN, ↓LBP, Differentiation of adipocyte: ↓KLF8, Adipogenesis: ↓ADAM23, ↓BMP3, ↓POSTN, Autophagy: ↓CLEC4A3, ↓PLD4, ↑GSDMC, ↑HK2, Immune response: ↓S100A8, ↓DOCK8, ↓EAR10, ↓MPEG1, ↓UBD, ↓AKAP13, Lysosomal program: ↓LIPA, Inflammatory response: ↓PTAFR, ↓FGF13, ↓OXTR, ↓KNG1, ↓SERPINA3M, Lipid metabolism: ↓NCEH1, Anti-inflammation: ↑USP2, angiogenesis: ↓SERPINF1, ↓ANGPTL4, ↑SCFA-producing bacteria, ↓Obesogenic bacteria | Seo et al. [83] |
| Biotransformation products | |||||
| Lentilactobacillus kefiri DH5 (LKDH5) from Kefir | CPB (postbiotic) from whey (WHE) and citrus pomace extract (CPX) | C57BL/6J |
| ↑Hesperetin, CPB group: ↓adipose tissue weight/body weight ratio, ↓TG Adipose tissue: ↑UCP-1, ↑PGC-1α ↑Anaerovorax odorimutans ↓Olsenella profusa | Youn et al. [85] |
| Cell-free extract | |||||
| Lactobacillus paracasei | CFE | Wistar albino male rats |
| ↓Body weight gain, ↓serum lipid level CFE-1 (29%), and CFE-2 (34%), ↓TG level CFE-1 (32%), and CFE-2 (45%), ↓serum cholesterol CFE-1 (32%), CFE-2 (39%), ↓LDL-C CFE-1 (38%), CFE-2 (50%), ↑HDL-C CFE-1 (20%), CFE-2 (30%) ↓MDA, ↑SOD, ↑CAT, and ↑GSH-px. | Osman et al. [87] |
| Bifidobacterium longum DS0956 and Lactobacillus rhamnosus DS0508 28 = Bifidobacterium longum DS0950;13 = Bifidobacterium bifidum DS0908; 30 = B. longum DS0956; 51 = Lactobacillus rhamnosus DS0508 | CFE | 3T3-L1 preadipocytes | 1, 5, or 10 μL per 1 mL | Brown-adipocyte markers: ↑PGC-1α, ↑UCP-1, and ↑PRDM16, beige-cell genes: ↑CD137, ↑FGF21, ↑P2RX5, and ↑Tbx1, white markers: ↑ap2, ↑PPARγ. Lipolysis factor: ↑HSL, ↑ATGL Activated PKA pathway ↑p-CREB, ↑p-HSL Reversed downregulated expressions of UCP-1, PGC-1α, and PPARγ | Hossain et al. [88] |
| BSs = 30 = B. longum DS0956; 51 = Lactobacillus rhamnosus DS0508 | C57BL/6N mice (n = 56) |
| ↓Body weight: BS groups, ↑UCP-1, PGC-1α, PRDM16 (BS 30), ↑HSL, ↑PLIN1, ↑aP2. (BS 30) M1 polarisation marker: ↓IL-1β, ↓TNF-α (BS 30) M2 polarisation marker: ↑ARG1, ↑CD206 (BS 30) | ||
| B. bifidum DS0908, B. longum DS0950 | CFE | Male C57BL/6 mice (n = 56) |
| ↓weight gain ↓fat accumulation, improved insulin sensitivity and glucose metabolism. Lipid profile, Culture supernatant: ↓TG Both pellet and supernantant: ↓LDL, ↓cholesterol, ↑HDL | Rahman et al. [89] |
| C3H10T1/2 MSCs | 5 μL per 1 mL | ↑UCP-1, ↑PPARγ, ↑PGC-1α Beige adipocyte-specific markers: ↑P2RX5, ↑FGF21, Brown adipocyte-specific marker: ↑Cox2 ↑p-PKA,↑p-p38 MAPK, ↓AMP, ↓CREB S133 | |||
| EVs | |||||
| Akkermansia muciniphila | Evs | Male C57BL/6 mice | First group:
| Both treatments: low level of body weight gain and substantial reduction in food intake in HFD mice ↓Adipocyte size in both treatments but more noticeable in eVs. eVs group: ↓TC, ↑PPARα, ↑PPARγ, ↓TGF-β Alleviated inflammation: ↓TLR-4, ↓TNF-α, ↓IL-6. Improved barrier function: ↑CLDN-1, ↑ZO-1, ↑OCLDN, and ↓CLDN-2. Both treatments influenced body weight in the ND group. | Ashrafian et al. [96] |
| Caco-2 | 10 μg of EV, 24 h | EV: ↑ZO-1, ↑OCLDN, and ↑CLDN-1 | |||
| Akkermansia muciniphila Live and pasteurised (P) | eVs | Male C57BL/6 mice |
| ↓Food intake in pasteurised bacterium and its eVs EV: ↓adipocyte size and showed normal morphology ↓TG, ↓LDL, ↑HDL. ↓TNF-α, ↓IL-6, ↑IL-10 ↑ZO-1 and ↑OCLDN (highest in EV), ↑CLDN-1 (highest in P), ↓CLDN-2 (EV and P). Colonic inflammation markers: ↑TLR-2, ↑IL-10, ↓TNF-α, ↓TLR-4 (EV highest) Lipid metabolism: ↑Angptl4 (highest in EV) | Ashrafian et al. [97] |
| Metabolites | |||||
| Commercial | Urolithin A (Uro-A), urolithin B (Uro-B) | Wistar rats (n = 24) |
| Significant reduction in final body weight ↓Cholesterol, ↓LDL-C, ↓TG, ↑HDL-C Uro-A: ↑Bacteroidetes, ↑Proteobacteria, Restored Firmicutes Uro-B: ↑Firmicutes, ↓Proteobacteria | Abdulrahman et al. [101] |
| Pomegranate ellagitannin (eTs) | Uro-A | C57BL/6 mice, leptin-deficient ob/ob mice |
| UA prevented both diet-induced and genetic obesity ↓Adipocytes, ↓fat mass, ↓body weight, ↓Plasma TG ↑Energy expenditure, ↑Thermogenesis in BAT, ↑browning in WAT In liver: ↓TNF-α, ↓IL-6, ↓TG, ↓liver weight | Xia et al. [102] |
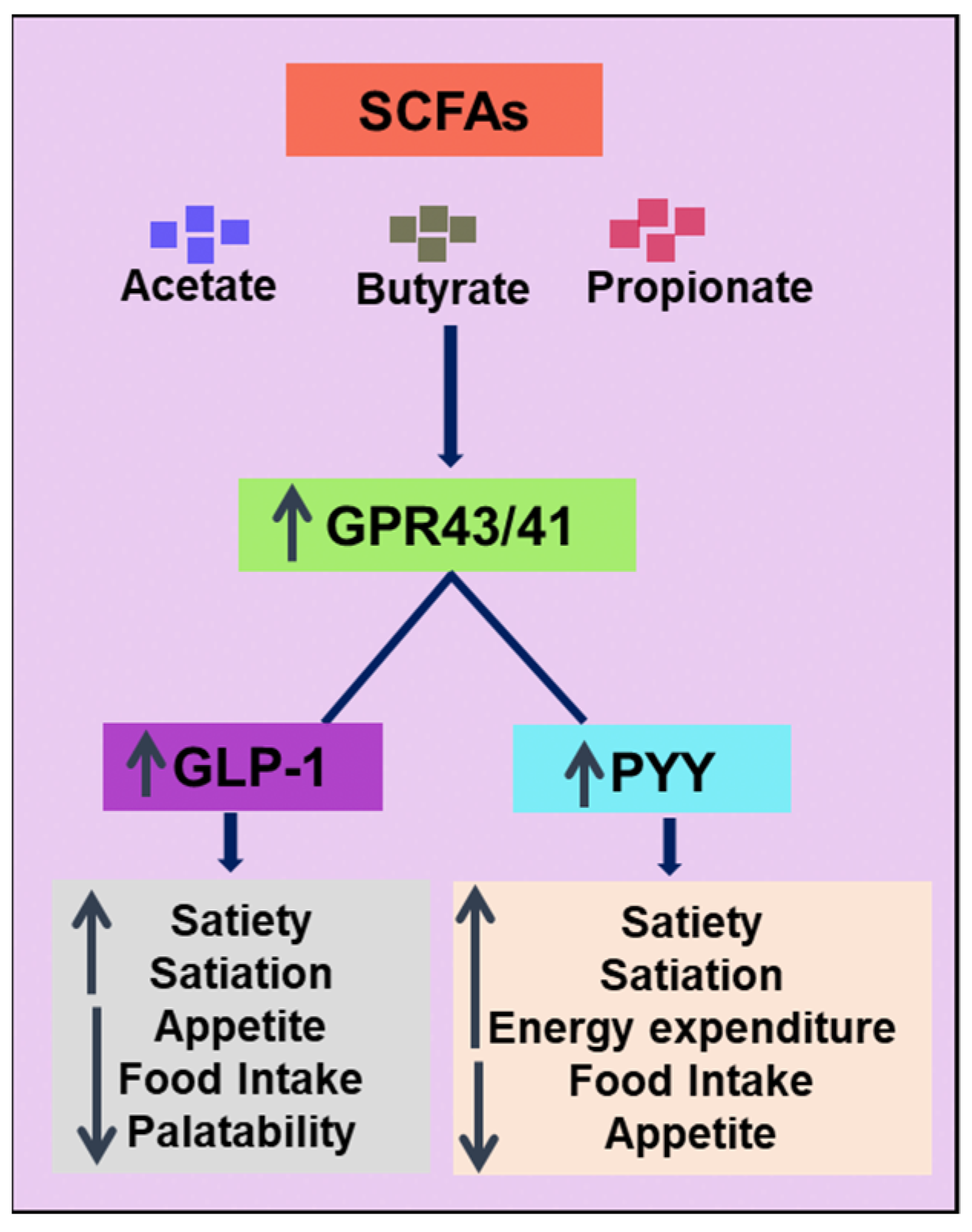
| SCFA | Butyrate | Male C57BL/6J mice |
| ↑Fatty acid oxidation, ↑thermogenic markers (PGC-1α and UCP-1) in BAT ↓adiposity, ↑mitochondrial biogenesis and function in BAT | Gao et al. [113] |
| SCFA | Butyrate, Acetate, and Propionate | Male C57BL/6N mice |
| Butyrate and propionate inhibited weight gain completely, acetate 40% suppression Butyrate and propionate reduced food intake and stimulated gut hormones | Lin et al. [114] |
| SCFA | Butyrate | Male APOE*3-Leiden.CETP (E3L.CETP) |
| Prevented body weight gain and weight of the g-WAT, ↑Fat oxidation, ↑thermogenic marker (UCP-1) in BAT | Li et al. [115] |
| SCFA | Acetate | 3T3-L1 cells | Na acetate (1 mM), 3 days | ↑UCP-1, ↑PGC-1α, ↑PRDM16, ↑PPARα, ↑DiO2, ↑CIDEA, ↑FABP3 Beige adipocyte-selective genes: ↑TMEM26, ↑TBX1 | Hanatani et al. [112] |
| Male KK-Ay mice (obese diabetic) |
| ↑Thermogenic markers in BAT ↑Browning markers in WAT | |||
| SCFA | Acetate | Immortalised BAT | Acetate (10 mM) or acute treatment 6 h (10 mM) | ↑PPARγ, ↑AP2, ↑PGC-1α, ↑UCP-1 ↑p-ERK1/2, ↑p-CREB, ↑GPR3 | Hu et al. [116] |
| Male C57BL/6J mice | Na acetate (150 mM), 6 weeks, in drinking water | ↑PGC-1α, ↑UCP-1 | |||
| SCFA | Acetate | Male C57BL/6JRj mice |
| Acetate suppressed hepatic lipogenesis ↑Body temperature ↑Browning markers’ expression in WAT | Weitkunat et al. [117] |
| SCFA | Acetate, Propionate, Butyrate, and their admixture | Male C57BL/6 J mice |
| Inhibited body weight gain Biochemical parameters: ↓TG, ↓cholesterol, ↓IL-1β, ↓MCP-1, ↓IL-6 ↑GPR43 expression in the adipose tissue and decreased expression in colon ↓Leptin expression by acetate or SCFA admixture ↑Expression of adiponectin and resistin by all SCFAs or their admixture Promoted beige adipogenesis | Lu et al. [118] |
| Metabolite of anthocyanins | Vanillic acid | Male C57BL/6N mice | HF/high sucrose diet + vanillic acid (0.5%), 16 weeks | ↑Expression of browning markers, and thermogenic markers, ↑cold tolerance, ↑mitochondriogeneis in BAT and WAT | Han et al. [119] |
| Linoleic acid metabolites | KetoA | Male C57BL/6N mice | HF diet + KetoA (0.;1%), 10 weeks | ↑UCP-1 in BAT and WAT ↑Expression of thermogenic markers and browning markers Activation of TRPV1 and SNS | Kim et al. [120] |
| Bacteriocins | |||||
| Bacteriocin | Plantaricin EF | Male C57BL6/J mice |
| Reduced food intake and weight gain. No change in gut microbiota (LP group). ↑Zonula Occludens-1 in LP group | Heeney et al. [122] |
| Bacteriocin | PJ4 by L. helveticus PJ4 | Male C57BL/6 J mice |
| Group: PJ4 more promising results: Significant decrease in body weight, ↓Adipocyte size, ↓TC, ↓TG, ↓LPS, ↓insulin, ↓IL-1β, ↓IL17, ↓IL-6, ↓IL-10, ↓IL14, ↓IFNγ, ↓TNFα, ↓MCP-1, ↓Adipokine and inflammasome. Modulated gut microflora: ↑Firmicutes, ↓Bacteroidetes and Proteobacteria | Bai et al. [123] |
| Cell-free lysates | |||||
| Ligilactobacillus salivarius strain 189 | Heat-killed (HK LS) | Pigs (n = 48) | Control group: a corn-soybean meal (CON), a basal diet Treatment group: a basal diet supplemented with HK LS 189 (0.2%), 4 weeks | Significant reduction in final body weight and daily weight gain. Significant difference in β-diversity between two groups. ↑Lentisphaerae, ↓Prevotella, ↓Blautia, ↓Lachnospira, YS2_ unclassified, ↓Mitsuokella, ↓Anaerostipes | Ryu et al. [125] |
| Lactobacillus plantarum L-137 | Heat-killed (HK L-137) | C57BL/6 J (n = 30–32/group) | Three groups by body weight: (a) Normal group (64% as carbohydrates, 20% as protein, and 6% of energy as fat) (b) HFD group (62% of energy as fat, 18% as protein, and 20% as carbohydrates) + HK L-137 (0.002%) (c) HFD group without treatment, 4 to 20 weeks | ↓Weight gain Plasma: ↓cholesterol, ↓glucose, ↓AST, ↓ALT level ↓LBP (a marker of endotoxaemia) plasma levels In eWAT epididymal adipose tissue: ↓CD11c, ↓IL-1β, ↓F4/80, ↓TNF-α, ↓MCP-1 | Yoshitake et al. [127] |
| Bifidobacterium longum BR-108 | Heat treated (IBL) | Male C57BL/6J mice |
| Reduced both weight gain and epididymal body fat mass, ↓TC, ↓cholesterol, ↓glucose, ↓LPS, ↓hepatic TG | Kikuchi et al. [128] |
| Bifidobacterium longum BR-108 | Heat treated (IBL) | Male Tsumura Suzuki obese diabetes (TSOD) mice, genetically obese mouse |
| Reduced the body weight gain 100 (6.5 g) and 150 mg/kg (7.2 g) Reduced adipose tissue buildup ↓blood glucose levels, ↓TC, ↓cholesterol, ↓FFA | Othman and Sakamoto, [129] |
| Kefir-derived lactic acid bacteria (LAB) + prebiotic LAB: Leuconostoc mesenteroides 4 (LMDH4), Lactobacillus kefiri DH5 (LKDH5) | Heat-killed HLAB | Male C57BL/6J |
| Reduced body weight gain and adipose tissue weight gain ↓Haptoglobin (HP) ↓WFDC21 ↓FABP4 ↓FAS | Seo et al. [130] |
| Lactobacillus plantarum K8 | LAB-P | 3T3-L1 preadipocytes |
| Suppressed lipid accumulation 50 (12%), 100 (42%), and 200 (58%) μg/mL. Reduced fat droplets ↓PPARγ, ↓C/EBPα, ↓FABP4 ↓p-JAK2, ↓p-STAT3 and ↓p-STAT5, ↑p-AMPKα | Kim H. et al. [131] |
| Lactiplantibacillus plantarum K8 | LAB-P | Male C57BL/6J mice |
| Reductions in weight gain Reduced HFD-induced hypertrophy: eWAT (36%), mWAT (20%), and iWAT (40%) ↓Hepatic fat accumulation, ↑p-AMPKα. Regulation of macrophages (adipose tissues): ↑CD206, ↓CD11c. ↓IL-1β, ↓IL-6, ↓NF-κB | Lim et al. [132] |
| LactiplantibacillusPlantarum (LP) K8 | Heat-treated LPK8 (K8HK) | Male C57BL/6 mice |
| ↓TG in both groups (live and heat-killed) ↓PPARγ, ↓C/EBPα, ↓FABP4 | Jang et al. [133] |
| 3T3-L1 cells | Live and heat-killed 109 CFU/mL | No cytotoxicity with heat-killed ↓TG, ↓PPARγ, ↓C/EBPα, ↓FABP4, ↓ACC, ↓FAS, ↓SCD1, ↑SOCS-1, ↓p-JAK2, ↓p-STAT3 compared to control | |||
| Lactobacillus brevis KB290 | Heat-killed KB290 (KB) | Male C57BL/6J mice |
| ↓Weights of epididymal and renal adipose tissue, ↓area of epididymal adipocytes ↑Adiponectin, ↑β3-adrenergic receptor In epididymal adipose tissue serum ↑FAA Altered microbiota composition | Watanabe et al. [134] |
4.2. Clinical Studies
| Source | Postbiotic/Paraprobiotic | Subjects Type and Count (n) | Design/Duration | Format/Dose | Outcomes | Country/Reference |
|---|---|---|---|---|---|---|
| SCFA | Propionate | Overweight adults, 60 | Randomised, double-blind, placebo-controlled, parallel design | Inulin-propionate ester (10 g/day) or inulin-control group (10 g/day), 24 weeks | ↓weight gain, ↓intrahepatocellular lipid content, ↓intra-abdominal adipose tissue distribution, Inhibited decline in insulin sensitivity ↑PYY, ↑GLP-1 | UK, Chambers et al. [135] |
| SCFA | Acetate | Overweight/obese men, 10 | Randomised, double-blind, crossover trial | Distal and proximal colonic sodium acetate infusions (one each) colonic acetate (100 or 180 mmol/L dissolved in saline 120 mL) Placebo: 120 mL, 3 days | Distal colonic acetate: ↑Fasting fat oxidation. ↑PYY, ↑fasting circulating acetate, ↑postprandial glucose, ↑insulin concentrations, ↓TNF-α Proximal colonic acetate: no significant difference | Netherlands, van der Beek et al. [136] |
| SCFA | Acetate, butyrate, propionate | Overweight/obese men, 13 | Randomised, double-blind, crossover study | HA: Na acetate (24 mmol 60%), Na propionate (8 mmol, 20%), Na butyrate (8 mmol, 20%). HP: Na acetate (18 mmol, 45%), Na propionate (14 mmol, 35%), Na butyrate (8 mmol, 20%). HB: Na acetate (18 mmol 45%), Na butyrate (14 mmol, 35%), Na propionate (8 mmol, 20%), all in 200 mL water. Placebo: 40 mmol sodium chloride in 200 mL water, 4 days | All treatments: ↑fasting fat oxidation. ↑PYY (fasting and postprandial plasma) ↓lipolysis HA and HP: ↑resting energy expenditure | Netherlands, Canfora et al. [137] |
| Pediococcus pentosaceus LP28 from longan fruit | Heat-killed LP28 | Overweight, 62: heat-killed LP28: n = 21, placebo: n = 20, LP28 (living): n = 21 | Randomised, double-blind, placebo-controlled, 12 weeks | LP28 (living) group: 10 mL spoon (1011 cells) Heat-killed LP28: 7.5 mL (1011 cells) Placebo: 7.5 mL | Heat-killed LP28: ↓body fat mass, ↓BMI, ↓waist circumference, ↓body fat percentages | Japan, Higashikawa et al. [139] |
| Lactobacillus amylovorus CP1563 from human fecal samples | Fragmented CP1563 | Overweight and mildly obese, 200 Test: n = 100, placebo: n = 100 | Double-blinded, placebo-controlled, randomised clinical trial (RCT), 12 weeks | Beverages with fragmented CP1563: 200 mg in a 500 mL bottle per patient per day | ↓Body fat percentage, ↓whole body fat, ↓visceral fat, ↓TG, ↓TC, ↓LDL-C, ↓diastolic blood pressure | Japan, Nakamura et al. [140] |
| Lactobacillus amylovorus CP1563 | Fragmented CP1563 with 10-HOA | Healthy subjects, 109 Test: n = 100, placebo: n = 100 | Randomised, double-blind, placebo-controlled, parallel study, 12 weeks | Beverages with fragmented CP1563: 200 mg (1.44 mg of 10-HOA) in a 500 mL bottle per patient per day | ↓Abdominal fat, ↓total fat, ↓visceral fat, ↓subcutaneous fat ↑genera Roseburia and Lachnospiraceae | Japan, Sugawara et al. [141] |
5. Conclusions and Future Implications
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Afshin, A.; Forouzanfar, M.; Reitsma, M.; Sur, P.; Estep, K.; Lee, A. Health effects of overweight and obesity in 195 countries over 25 years. N. Engl. J. Med. 2017, 377, 13–27. [Google Scholar] [PubMed]
- Charakida, M.; Khan, T.; Johnson, W.; Finer, N.; Woodside, J.; Whincup, P.H.; Sattar, N.; Kuh, D.; Hardy, R.; Deanfield, J. Lifelong patterns of BMI and cardiovascular phenotype in individuals aged 60–64 years in the 1946 British birth cohort study: An epidemiological study. Lancet Diabetes Endocrinol. 2014, 2, 648–654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thaker, V.V. Genetic and epigenetic causes of obesity. Adolesc. Med. State Art Rev. 2017, 28, 379–405. [Google Scholar] [PubMed]
- Bäckhed, F.; Ding, H.; Wang, T.; Hooper, L.V.; Koh, G.Y.; Nagy, A.; Semenkovich, C.F.; Gordon, J.I. The gut microbiota as an environmental factor that regulates fat storage. Proc. Natl. Acad. Sci. USA 2004, 101, 15718–15723. [Google Scholar] [CrossRef] [Green Version]
- Sheykhsaran, E.; Abbasi, A.; Leylabadlo, H.E.; Sadeghi, J.; Mehri, S.; Mazraeh, F.N.; Feizi, H.; Baghi, H.B. Gut microbiota and obesity: An overview of microbiota to microbial-based therapies. Postgrad. Med. J. 2022. [Google Scholar] [CrossRef]
- Dong, L.; Qin, C.; Li, Y.; Wu, Z.; Liu, L. Oat phenolic compounds regulate metabolic syndrome in high fat diet-fed mice via gut microbiota. Food Biosci. 2022, 50, 101946. [Google Scholar] [CrossRef]
- Hruby, A.; Hu, F.B. The epidemiology of obesity: A big picture. Pharmacoeconomics 2015, 33, 673–689. [Google Scholar] [CrossRef]
- Müller, T.D.; Blüher, M.; Tschöp, M.H.; DiMarchi, R.D. Anti-obesity drug discovery: Advances and challenges. Nat. Rev. Drug Discov. 2022, 21, 201–223. [Google Scholar] [CrossRef]
- Lin, X.; Li, H. Obesity: Epidemiology, pathophysiology, and therapeutics. Front. Endocrinol. 2021, 12, 706978. [Google Scholar] [CrossRef]
- Ahirwar, R.; Mondal, P.R. Prevalence of obesity in India: A systematic review. Diabetes Metab. Syndr. Clin. Res. Rev. 2019, 13, 318–321. [Google Scholar] [CrossRef]
- Collaboration, N.R.F. Trends in adult body-mass index in 200 countries from 1975 to 2014: A pooled analysis of 1698 population-based measurement studies with 19· 2 million participants. Lancet 2016, 387, 1377–1396. [Google Scholar]
- Fryar, C.D.; Carroll, M.D.; Afful, J. Prevalence of Overweight, Obesity, and Severe Obesity among Children and Adolescents Aged 2–19 Years: United States, 1963–1965 through 2017–2018. NCHS Health E-Stats. 2020. Available online: https://www.cdc.gov/nchs/data/hestat/obesity-child-17-18/obesity-child.htm (accessed on 18 December 2022).
- Development Initiatives. 2018 Global Nutrition Report: Shining a Light to Spur Action on Nutrition; Development Initiatives Poverty Research Ltd.: Bristol, UK, 2018; Available online: https://globalnutritionreport.org/ (accessed on 13 December 2022).
- Korea Disease Control and Prevention Agency. Korea Health Statistics 2021. The 8th Korea National Health and Nutrition Examination Survey 3rd Year (2021). Available online: https://www.index.go.kr/unify/idx-info.do?idxCd=8021 (accessed on 10 March 2023).
- Baik, I. Forecasting obesity prevalence in Korean adults for the years 2020 and 2030 by the analysis of contributing factors. Nutr. Res. Pract. 2018, 12, 251–257. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.S.; Han, B.-D.; Han, K.; Jung, J.-H.; Son, J.W. Obesity Fact Sheet in Korea, 2021: Trends in obesity prevalence and obesity-related comorbidity incidence stratified by age from 2009 to 2019. J. Obes. Metab. Syndr. 2022, 31, 169. [Google Scholar] [CrossRef] [PubMed]
- Unser, A.M.; Tian, Y.; Xie, Y. Opportunities and challenges in three-dimensional brown adipogenesis of stem cells. Biotechnol. Adv. 2015, 33, 962–979. [Google Scholar] [CrossRef] [Green Version]
- Vázquez-Vela, M.E.F.; Torres, N.; Tovar, A.R. White adipose tissue as endocrine organ and its role in obesity. Arch. Med. Res. 2008, 39, 715–728. [Google Scholar] [CrossRef]
- Bartelt, A.; Heeren, J. Adipose tissue browning and metabolic health. Nat. Rev. Endocrinol. 2014, 10, 24–36. [Google Scholar] [CrossRef]
- Betz, M.J.; Enerbäck, S. Targeting thermogenesis in brown fat and muscle to treat obesity and metabolic disease. Nat. Rev. Endocrinol. 2018, 14, 77–87. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Wang, Z.; Tan, B.K.; Christian, M. Dietary polyphenols turn fat “brown”: A narrative review of the possible mechanisms. Trends Food Sci. Technol. 2020, 97, 221–232. [Google Scholar] [CrossRef]
- Bartesaghi, S.; Hallen, S.; Huang, L.; Svensson, P.-A.; Momo, R.A.; Wallin, S.; Carlsson, E.K.; Forslöw, A.; Seale, P.; Peng, X.-R. Thermogenic activity of UCP1 in human white fat-derived beige adipocytes. Mol. Endocrinol. 2015, 29, 130–139. [Google Scholar] [CrossRef] [Green Version]
- Vargas-Castillo, A.; Fuentes-Romero, R.; Rodriguez-Lopez, L.A.; Torres, N.; Tovar, A.R. Understanding the biology of thermogenic fat: Is browning a new approach to the treatment of obesity? Arch. Med. Res. 2017, 48, 401–413. [Google Scholar] [CrossRef]
- Montanari, T.; Pošćić, N.; Colitti, M. Factors involved in white-to-brown adipose tissue conversion and in thermogenesis: A review. Obes. Rev. 2017, 18, 495–513. [Google Scholar] [CrossRef] [PubMed]
- Harms, M.; Seale, P. Brown and beige fat: Development, function and therapeutic potential. Nat. Med. 2013, 19, 1252–1263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madsen, M.S.; Siersbæk, R.; Boergesen, M.; Nielsen, R.; Mandrup, S. Peroxisome proliferator-activated receptor γ and C/EBPα synergistically activate key metabolic adipocyte genes by assisted loading. Mol. Cell. Biol. 2014, 34, 939–954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosen, E.D.; Walkey, C.J.; Puigserver, P.; Spiegelman, B.M. Transcriptional regulation of adipogenesis. Genes Dev. 2000, 14, 1293–1307. [Google Scholar] [CrossRef] [PubMed]
- Tang, C.; Lu, Z. Health promoting activities of probiotics. J. Food Biochem. 2019, 43, e12944. [Google Scholar] [CrossRef]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S. Expert consensus document: The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef] [Green Version]
- Adams, C.A. The probiotic paradox: Live and dead cells are biological response modifiers. Nutr. Res. Rev. 2010, 23, 37–46. [Google Scholar] [CrossRef] [Green Version]
- Homayouni Rad, A.; Aghebati Maleki, L.; Samadi Kafil, H.; Abbasi, A. Postbiotics: A novel strategy in food allergy treatment. Crit. Rev. Food Sci. Nutr. 2021, 61, 492–499. [Google Scholar] [CrossRef]
- Tang, C.; Kong, L.; Shan, M.; Lu, Z.; Lu, Y. Protective and ameliorating effects of probiotics against diet-induced obesity: A review. Food Res. Int. 2021, 147, 110490. [Google Scholar] [CrossRef]
- Ejtahed, H.-S.; Angoorani, P.; Soroush, A.-R.; Atlasi, R.; Hasani-Ranjbar, S.; Mortazavian, A.M.; Larijani, B. Probiotics supplementation for the obesity management; A systematic review of animal studies and clinical trials. J. Funct. Foods 2019, 52, 228–242. [Google Scholar] [CrossRef]
- Kobyliak, N.; Falalyeyeva, T.; Beregova, T.; Spivak, M. Probiotics for experimental obesity prevention: Focus on strain dependence and viability of composition. Endokrynol. Pol. 2017, 68, 659–667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pradhan, D.; Mallappa, R.H.; Grover, S. Comprehensive approaches for assessing the safety of probiotic bacteria. Food Control 2020, 108, 106872. [Google Scholar] [CrossRef]
- Ayichew, T.; Belete, A.; Alebachew, T.; Tsehaye, H.; Berhanu, H.; Minwuyelet, A. Bacterial probiotics their importances and limitations: A review. J. Nutr. Health Sci. 2017, 4, 202. [Google Scholar]
- Evivie, S.E.; Huo, G.-C.; Igene, J.O.; Bian, X. Some current applications, limitations and future perspectives of lactic acid bacteria as probiotics. Food Nutr. Res. 2017, 61, 1318034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohen, P.A. Probiotic safety—No guarantees. JAMA Intern. Med. 2018, 178, 1577–1578. [Google Scholar] [CrossRef]
- Sender, R.; Fuchs, S.; Milo, R. Are we really vastly outnumbered? Revisiting the ratio of bacterial to host cells in humans. Cell 2016, 164, 337–340. [Google Scholar] [CrossRef] [Green Version]
- Rajakovich, L.J.; Balskus, E.P. Metabolic functions of the human gut microbiota: The role of metalloenzymes. Nat. Prod. Rep. 2019, 36, 593–625. [Google Scholar] [CrossRef] [Green Version]
- Gensollen, T.; Iyer, S.S.; Kasper, D.L.; Blumberg, R.S. How colonization by microbiota in early life shapes the immune system. Science 2016, 352, 539–544. [Google Scholar] [CrossRef] [Green Version]
- Aguilar-Toalá, J.; Garcia-Varela, R.; Garcia, H.; Mata-Haro, V.; González-Córdova, A.; Vallejo-Cordoba, B.; Hernández-Mendoza, A. Postbiotics: An evolving term within the functional foods field. Trends Food Sci. Technol. 2018, 75, 105–114. [Google Scholar] [CrossRef]
- Tomar, S.K.; Anand, S.; Sharma, P.; Sangwan, V.; Mandal, S. Role of probiotics, prebiotics, synbiotics and postbiotics in inhibition of pathogens. In The Battle against Microbial Pathogens: Basic Science, Technological Advances and Educational Programs; Méndez-Vilas, A., Ed.; Formatex Research Center: Badajoz, Spain, 2015; pp. 717–732. [Google Scholar]
- Zhang, J.; Du, G.-C.; Zhang, Y.; Liao, X.-Y.; Wang, M.; Li, Y.; Chen, J. Glutathione protects Lactobacillus sanfranciscensis against freeze-thawing, freeze-drying, and cold treatment. Appl. Environ. Microbiol. 2010, 76, 2989–2996. [Google Scholar] [CrossRef] [Green Version]
- Netzker, T.; Fischer, J.; Weber, J.; Mattern, D.J.; König, C.C.; Valiante, V.; Schroeckh, V.; Brakhage, A.A. Microbial communication leading to the activation of silent fungal secondary metabolite gene clusters. Front. Microbiol. 2015, 6, 299. [Google Scholar] [CrossRef] [PubMed]
- Salminen, S.; Collado, M.C.; Endo, A.; Hill, C.; Lebeer, S.; Quigley, E.M.; Sanders, M.E.; Shamir, R.; Swann, J.R.; Szajewska, H. The International Scientific Association of Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of postbiotics. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 649–667. [Google Scholar] [CrossRef] [PubMed]
- Cerdó, T.; García-Santos, J.A.; Bermúdez, M.G.; Campoy, C. The role of probiotics and prebiotics in the prevention and treatment of obesity. Nutrients 2019, 11, 635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Li, Y.; Ren, X.; Zhang, X.; Wu, Z.; Liu, L. The positive correlation of antioxidant activity and prebiotic effect about oat phenolic compounds. Food Chem. 2023, 402, 134231. [Google Scholar] [CrossRef]
- Li, Y.; Qin, C.; Dong, L.; Zhang, X.; Wu, Z.; Liu, L.; Yang, J.; Liu, L. Whole grain benefit: Synergistic effect of oat phenolic compounds and β-glucan on hyperlipidemia via gut microbiota in high-fat-diet mice. Food Funct. 2022, 13, 12686–12696. [Google Scholar] [CrossRef]
- Xu, Y.; Zang, J.; Regenstein, J.M.; Xia, W. Technological roles of microorganisms in fish fermentation: A review. Crit. Rev. Food Sci. Nutr. 2021, 61, 1000–1012. [Google Scholar] [CrossRef]
- de Almada, C.N.; Almada, C.N.; Martinez, R.C.; Sant’Ana, A.S. Paraprobiotics: Evidences on their ability to modify biological responses, inactivation methods and perspectives on their application in foods. Trends Food Sci. Technol. 2016, 58, 96–114. [Google Scholar] [CrossRef]
- Vallejo-Cordoba, B.; Castro-López, C.; García, H.S.; González-Córdova, A.F.; Hernández-Mendoza, A. Postbiotics and paraprobiotics: A review of current evidence and emerging trends. Adv. Food Nutr. Res. 2020, 94, 1–34. [Google Scholar]
- Guimarães, J.T.; Balthazar, C.F.; Scudino, H.; Pimentel, T.C.; Esmerino, E.A.; Ashokkumar, M.; Freitas, M.Q.; Cruz, A.G. High-intensity ultrasound: A novel technology for the development of probiotic and prebiotic dairy products. Ultrason. Sonochem. 2019, 57, 12–21. [Google Scholar] [CrossRef]
- Manassi, C.F.; de Souza, S.S.; de Souza Hassemer, G.; Sartor, S.; Lima, C.M.G.; Miotto, M.; Lindner, J.D.D.; Rezzadori, K.; Pimentel, T.C.; Ramos, G.L.d.P.A. Functional meat products: Trends in pro-, pre-, syn-, para- and post-biotic use. Food Res. Int. 2022, 154, 111035. [Google Scholar] [CrossRef]
- Bourebaba, Y.; Marycz, K.; Mularczyk, M.; Bourebaba, L. Postbiotics as potential new therapeutic agents for metabolic disorders management. Biomed. Pharmacother. 2022, 153, 113138. [Google Scholar] [CrossRef] [PubMed]
- Chan, M.Z.A.; Liu, S.-Q. Fortifying foods with synbiotic and postbiotic preparations of the probiotic yeast, Saccharomyces boulardii. Curr. Opin. Food Sci. 2022, 43, 216–224. [Google Scholar] [CrossRef]
- Patel, R.M.; Denning, P.W. Therapeutic use of prebiotics, probiotics, and postbiotics to prevent necrotizing enterocolitis: What is the current evidence? Clin. Perinatol. 2013, 40, 11–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imperial, I.C.; Ibana, J.A. Addressing the antibiotic resistance problem with probiotics: Reducing the risk of its double-edged sword effect. Front. Microbiol. 2016, 7, 1983. [Google Scholar] [CrossRef] [Green Version]
- Tuladhar, S.; Kanneganti, T.-D. NLRP12 in innate immunity and inflammation. Mol. Asp. Med. 2020, 76, 100887. [Google Scholar] [CrossRef]
- Purohit, J.S.; Hu, P.; Burke, S.J.; Collier, J.J.; Chen, J.; Zhao, L. The effects of NOD activation on adipocyte differentiation. Obesity 2013, 21, 737–747. [Google Scholar] [CrossRef]
- Anhê, F.F.; Jensen, B.A.; Perazza, L.R.; Tchernof, A.; Schertzer, J.D.; Marette, A. Bacterial postbiotics as promising tools to mitigate cardiometabolic diseases. J. Lipid Atheroscler. 2021, 10, 123–129. [Google Scholar] [CrossRef]
- Cavallari, J.F.; Fullerton, M.D.; Duggan, B.M.; Foley, K.P.; Denou, E.; Smith, B.K.; Desjardins, E.M.; Henriksbo, B.D.; Kim, K.J.; Tuinema, B.R. Muramyl dipeptide-based postbiotics mitigate obesity-induced insulin resistance via IRF4. Cell Metab. 2017, 25, 1063–1074.e1063. [Google Scholar] [CrossRef]
- Cavallari, J.F.; Barra, N.G.; Foley, K.P.; Lee, A.; Duggan, B.M.; Henriksbo, B.D.; Anhê, F.F.; Ashkar, A.A.; Schertzer, J.D. Postbiotics for NOD2 require nonhematopoietic RIPK2 to improve blood glucose and metabolic inflammation in mice. Am. J. Physiol. Endocrinol. Metab. 2020, 318, E579–E585. [Google Scholar] [CrossRef]
- Philpott, D.J.; Sorbara, M.T.; Robertson, S.J.; Croitoru, K.; Girardin, S.E. NOD proteins: Regulators of inflammation in health and disease. Nat. Rev. Immunol. 2014, 14, 9–23. [Google Scholar] [CrossRef]
- Duggan, B.M.; Singh, A.M.; Chan, D.Y.; Schertzer, J.D. Postbiotics engage IRF4 in adipocytes to promote sex-dependent changes in blood glucose during obesity. Physiol. Rep. 2022, 10, e15439. [Google Scholar] [CrossRef] [PubMed]
- Nataraj, B.H.; Ali, S.A.; Behare, P.V.; Yadav, H. Postbiotics-parabiotics: The new horizons in microbial biotherapy and functional foods. Microb. Cell Fact. 2020, 19, 168. [Google Scholar] [CrossRef] [PubMed]
- Fagan, R.P.; Fairweather, N.F. Biogenesis and functions of bacterial S-layers. Nat. Rev. Microbiol. 2014, 12, 211–222. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.; Lee, H.G.; Han, S.; Seo, K.-H.; Kim, H. Effect of surface layer proteins derived from paraprobiotic kefir lactic acid bacteria on inflammation and high-fat diet-induced obesity. J. Agric. Food Chem. 2021, 69, 15157–15164. [Google Scholar] [CrossRef] [PubMed]
- Yoon, D.H.; Kim, J.Y.; Keon, H.; Lee, H.G.; Seo, K.; Shim, J.-J.; Lee, J.-l.; Kim, H. The anti-adipogenic effects of postbiotics derived from plant-based lactic acid bacteria. Res. Sq. 2022; preprint. [Google Scholar] [CrossRef]
- Lebeer, S.; Vanderleyden, J.; De Keersmaecker, S.C. Host interactions of probiotic bacterial surface molecules: Comparison with commensals and pathogens. Nat. Rev. Microbiol. 2010, 8, 171–184. [Google Scholar] [CrossRef]
- Lebeer, S.; Claes, I.J.; Vanderleyden, J. Anti-inflammatory potential of probiotics: Lipoteichoic acid makes a difference. Trends Microbiol. 2012, 20, 5–10. [Google Scholar] [CrossRef]
- Yi, Z.-J.; Fu, Y.-R.; Li, M.; Gao, K.-S.; Zhang, X.-G. Effect of LTA isolated from bifidobacteria on D-galactose-induced aging. Exp. Gerontol. 2009, 44, 760–765. [Google Scholar] [CrossRef]
- Noh, S.Y.; Kang, S.-S.; Yun, C.-H.; Han, S.H. Lipoteichoic acid from Lactobacillus plantarum inhibits Pam2CSK4-induced IL-8 production in human intestinal epithelial cells. Mol. Immunol. 2015, 64, 183–189. [Google Scholar] [CrossRef]
- Mizuno, H.; Arce, L.; Tomotsune, K.; Albarracin, L.; Funabashi, R.; Vera, D.; Islam, M.A.; Vizoso-Pinto, M.G.; Takahashi, H.; Sasaki, Y. Lipoteichoic acid is involved in the ability of the immunobiotic strain Lactobacillus plantarum CRL1506 to modulate the intestinal antiviral innate immunity triggered by TLR3 activation. Front. Immunol. 2020, 11, 571. [Google Scholar] [CrossRef]
- Balaguer, F.; Enrique, M.; Llopis, S.; Barrena, M.; Navarro, V.; Álvarez, B.; Chenoll, E.; Ramon, D.; Tortajada, M.; Martorell, P. Lipoteichoic acid from Bifidobacterium animalis subsp. lactis BPL1: A novel postbiotic that reduces fat deposition via IGF-1 pathway. Microb. Biotechnol. 2022, 15, 805–816. [Google Scholar] [CrossRef]
- Yue, Y.; Li, S.; Shen, P.; Park, Y. Caenorhabditis elegans as a model for obesity research. Curr. Res. Food Sci. 2021, 4, 692–697. [Google Scholar] [CrossRef] [PubMed]
- Garsin, D.A.; Villanueva, J.M.; Begun, J.; Kim, D.H.; Sifri, C.D.; Calderwood, S.B.; Ruvkun, G.; Ausubel, F.M. Long-lived C. elegans daf-2 mutants are resistant to bacterial pathogens. Science 2003, 300, 1921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whitfield, C.; Wear, S.S.; Sande, C. Assembly of bacterial capsular polysaccharides and exopolysaccharides. Annu. Rev. Microbiol. 2020, 74, 521–543. [Google Scholar] [CrossRef] [PubMed]
- Angelin, J.; Kavitha, M. Exopolysaccharides from probiotic bacteria and their health potential. Int. J. Biol. Macromol. 2020, 162, 853–865. [Google Scholar] [CrossRef]
- Ryan, P.; Ross, R.; Fitzgerald, G.; Caplice, N.; Stanton, C. Sugar-coated: Exopolysaccharide producing lactic acid bacteria for food and human health applications. Food Funct. 2015, 6, 679–693. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhou, Z.; Li, Y.; Zhou, L.; Ding, Q.; Xu, L. Isolated exopolysaccharides from Lactobacillus rhamnosus GG alleviated adipogenesis mediated by TLR2 in mice. Sci. Rep. 2016, 6, 36083. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Park, S.; Oh, N.; Park, J.; Kwon, M.; Seo, J.; Roh, S. Oral intake of Lactobacillus plantarum L-14 extract alleviates TLR2- and AMPK-mediated obesity-associated disorders in high-fat-diet-induced obese C57BL/6J mice. Cell Prolif. 2021, 54, e13039. [Google Scholar] [CrossRef]
- Seo, K.-H.; Lee, H.G.; Eor, J.Y.; Jeon, H.J.; Yokoyama, W.; Kim, H. Effects of kefir lactic acid bacteria-derived postbiotic components on high fat diet-induced gut microbiota and obesity. Food Res. J. 2022, 157, 111445. [Google Scholar] [CrossRef]
- Kim, D.H.; Jeong, D.; Kang, I.B.; Kim, H.; Song, K.Y.; Seo, K.H. Dual function of Lactobacillus kefiri DH5 in preventing high-fat-diet-induced obesity: Direct reduction of cholesterol and upregulation of PPAR-α in adipose tissue. Mol. Nutr. Food Res. 2017, 61, 1700252. [Google Scholar] [CrossRef]
- Youn, H.-Y.; Seo, K.-H.; Kim, H.-J.; Kim, Y.-S.; Kim, H. Effect of postbiotics derived from kefir lactic acid bacteria-mediated bioconversion of citrus pomace extract and whey on high-fat diet-induced obesity and gut dysbiosis. Food Res. Int. 2022, 162, 111930. [Google Scholar] [CrossRef]
- Saadatzadeh, A.; Fazeli, M.R.; Jamalifar, H.; Dinarvand, R. Probiotic properties of lyophilized cell free extract of Lactobacillus casei. Jundishapur J. Nat. Pharm. Prod. 2013, 8, 131–137. [Google Scholar] [CrossRef] [Green Version]
- Osman, A.; El-Gazzar, N.; Almanaa, T.N.; El-Hadary, A.; Sitohy, M. Lipolytic postbiotic from Lactobacillus paracasei manages metabolic syndrome in albino wistar rats. Molecules 2021, 26, 472. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.; Park, D.-S.; Rahman, M.; Ki, S.-J.; Lee, Y.R.; Imran, K.; Yoon, D.; Heo, J.; Lee, T.-J.; Kim, Y.-S. Bifidobacterium longum DS0956 and Lactobacillus rhamnosus DS0508 culture-supernatant ameliorate obesity by inducing thermogenesis in obese-mice. Benef. Microbes 2020, 11, 361–373. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.S.; Lee, Y.; Park, D.-S.; Kim, Y.-S. Bifidobacterium bifidum DS0908 and Bifidobacterium longum DS0950 Culture-Supernatants Ameliorate Obesity-Related Characteristics in Mice with High-Fat Diet-Induced Obesity. J. Microbiol. Biotechnol. 2023, 33, 96–105. [Google Scholar] [CrossRef] [PubMed]
- Brown, L.; Wolf, J.M.; Prados-Rosales, R.; Casadevall, A. Through the wall: Extracellular vesicles in Gram-positive bacteria, mycobacteria and fungi. Nat. Rev. Microbiol. 2015, 13, 620–630. [Google Scholar] [CrossRef] [Green Version]
- Raposo, G.; Stahl, P.D. Extracellular vesicles: A new communication paradigm? Nat. Rev. Mol. Cell Biol. 2019, 20, 509–510. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Yang, J.; Wang, W.; Sanidad, K.Z.; Cinelli, M.A.; Wan, D.; Hwang, S.H.; Kim, D.; Lee, K.S.S.; Xiao, H. Soluble epoxide hydrolase is an endogenous regulator of obesity-induced intestinal barrier dysfunction and bacterial translocation. Proc. Natl. Acad. Sci. USA 2020, 117, 8431–8436. [Google Scholar] [CrossRef]
- Macchione, I.; Lopetuso, L.; Ianiro, G.; Napoli, M.; Gibiino, G.; Rizzatti, G.; Petito, V.; Gasbarrini, A.; Scaldaferri, F. Akkermansia muciniphila: Key player in metabolic and gastrointestinal disorders. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 8075–8083. [Google Scholar]
- Dao, M.C.; Everard, A.; Aron-Wisnewsky, J.; Sokolovska, N.; Prifti, E.; Verger, E.O.; Kayser, B.D.; Levenez, F.; Chilloux, J.; Hoyles, L. Akkermansia muciniphila and improved metabolic health during a dietary intervention in obesity: Relationship with gut microbiome richness and ecology. Gut 2016, 65, 426–436. [Google Scholar] [CrossRef] [Green Version]
- Plovier, H.; Everard, A.; Druart, C.; Depommier, C.; Van Hul, M.; Geurts, L.; Chilloux, J.; Ottman, N.; Duparc, T.; Lichtenstein, L. A purified membrane protein from Akkermansia muciniphila or the pasteurized bacterium improves metabolism in obese and diabetic mice. Nat. Med. 2017, 23, 107–113. [Google Scholar] [CrossRef] [Green Version]
- Ashrafian, F.; Shahriary, A.; Behrouzi, A.; Moradi, H.R.; Keshavarz Azizi Raftar, S.; Lari, A.; Hadifar, S.; Yaghoubfar, R.; Ahmadi Badi, S.; Khatami, S. Akkermansia muciniphila-derived extracellular vesicles as a mucosal delivery vector for amelioration of obesity in mice. Front. Microbiol. 2019, 10, 2155. [Google Scholar] [CrossRef] [PubMed]
- Ashrafian, F.; Keshavarz Azizi Raftar, S.; Lari, A.; Shahryari, A.; Abdollahiyan, S.; Moradi, H.R.; Masoumi, M.; Davari, M.; Omrani, M.D.; Vaziri, F. Extracellular vesicles and pasteurized cells derived from Akkermansia muciniphila protect against high-fat induced obesity in mice. Microb. Cell Fact. 2021, 20, 219. [Google Scholar] [CrossRef] [PubMed]
- Al-Harbi, S.A.; Abdulrahman, A.O.; Zamzami, M.A.; Khan, M.I. Urolithins: The gut based polyphenol metabolites of ellagitannins in cancer prevention, a review. Front. Nutr. 2021, 8, 647582. [Google Scholar] [CrossRef]
- Zhao, W.; Wang, Y.; Hao, W.; Yang, H.; Song, X.; Zhao, M.; Peng, S. Preparative isolation and purification of urolithins from the intestinal metabolites of pomegranate ellagitannins by high-speed counter-current chromatography. J. Chromatogr. B 2015, 990, 111–117. [Google Scholar] [CrossRef] [PubMed]
- González-Sarrías, A.; Núñez-Sánchez, M.Á.; García-Villalba, R.; Tomás-Barberán, F.A.; Espín, J.C. Antiproliferative activity of the ellagic acid-derived gut microbiota isourolithin A and comparison with its urolithin A isomer: The role of cell metabolism. Eur. J. Nutr. 2017, 56, 831–841. [Google Scholar] [CrossRef]
- Abdulrahman, A.O.; Alzubaidi, M.Y.; Nadeem, M.S.; Khan, J.A.; Rather, I.A.; Khan, M.I. Effects of urolithins on obesity-associated gut dysbiosis in rats fed on a high-fat diet. Int. J. Food Sci. Nutr. 2021, 72, 923–934. [Google Scholar] [CrossRef] [PubMed]
- Xia, B.; Shi, X.C.; Xie, B.C.; Zhu, M.Q.; Chen, Y.; Chu, X.Y.; Cai, G.H.; Liu, M.; Yang, S.Z.; Mitchell, G.A. Urolithin A exerts antiobesity effects through enhancing adipose tissue thermogenesis in mice. PLoS Biol. 2020, 18, e3000688. [Google Scholar] [CrossRef] [Green Version]
- Rauf, A.; Khalil, A.A.; Rahman, U.-U.; Khalid, A.; Naz, S.; Shariati, M.A.; Rebezov, M.; Urtecho, E.Z.; de Albuquerque, R.D.D.G.; Anwar, S. Recent advances in the therapeutic application of short-chain fatty acids (SCFAs): An updated review. Crit. Rev. Food Sci. Nutr. 2022, 62, 6034–6054. [Google Scholar] [CrossRef]
- Yao, Y.; Cai, X.; Fei, W.; Ye, Y.; Zhao, M.; Zheng, C. The role of short-chain fatty acids in immunity, inflammation and metabolism. Crit. Rev. Food Sci. Nutr. 2022, 62, 1–12. [Google Scholar] [CrossRef]
- Cheng, Y.; Liu, J.; Ling, Z. Short-chain fatty acids-producing probiotics: A novel source of psychobiotics. Crit. Rev. Food Sci. Nutr. 2022, 62, 7929–7959. [Google Scholar] [CrossRef]
- Kim, C.H. Control of lymphocyte functions by gut microbiota-derived short-chain fatty acids. Cell Mol. Immunol. 2021, 18, 1161–1171. [Google Scholar] [CrossRef] [PubMed]
- Reynés, B.; Palou, M.; Rodriguez, A.M.; Palou, A. Regulation of adaptive thermogenesis and browning by prebiotics and postbiotics. Front. Physiol. 2019, 9, 1908. [Google Scholar] [CrossRef] [PubMed]
- Pascale, A.; Marchesi, N.; Marelli, C.; Coppola, A.; Luzi, L.; Govoni, S.; Giustina, A.; Gazzaruso, C. Microbiota and metabolic diseases. Endocrine 2018, 61, 357–371. [Google Scholar] [CrossRef] [PubMed]
- Den Besten, G.; Van Eunen, K.; Groen, A.K.; Venema, K.; Reijngoud, D.-J.; Bakker, B.M. The role of short-chain fatty acids in the interplay between diet, gut microbiota, and host energy metabolism. J. Lipid Res. 2013, 54, 2325–2340. [Google Scholar] [CrossRef] [Green Version]
- Jung, R.; Shetty, P.; James, W.; Barrand, M.; Callingham, B. Reduced thermogenesis in obesity. Nature 1979, 279, 322–323. [Google Scholar] [CrossRef]
- Hu, J.; Lin, S.; Zheng, B.; Cheung, P.C. Short-chain fatty acids in control of energy metabolism. Crit. Rev. Food Sci. Nutr. 2018, 58, 1243–1249. [Google Scholar] [CrossRef]
- Hanatani, S.; Motoshima, H.; Takaki, Y.; Kawasaki, S.; Igata, M.; Matsumura, T.; Kondo, T.; Senokuchi, T.; Ishii, N.; Kawashima, J. Acetate alters expression of genes involved in beige adipogenesis in 3T3-L1 cells and obese KK-Ay mice. J. Clin. Biochem. Nutr. 2016, 59, 16–23. [Google Scholar] [CrossRef] [Green Version]
- Gao, Z.; Yin, J.; Zhang, J.; Ward, R.E.; Martin, R.J.; Lefevre, M.; Cefalu, W.T.; Ye, J. Butyrate improves insulin sensitivity and increases energy expenditure in mice. Diabetes 2009, 58, 1509–1517. [Google Scholar] [CrossRef] [Green Version]
- Lin, H.V.; Frassetto, A.; Kowalik, E.J., Jr.; Nawrocki, A.R.; Lu, M.M.; Kosinski, J.R.; Hubert, J.A.; Szeto, D.; Yao, X.; Forrest, G. Butyrate and propionate protect against diet-induced obesity and regulate gut hormones via free fatty acid receptor 3-independent mechanisms. PLoS ONE 2012, 7, e35240. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Yi, C.-X.; Katiraei, S.; Kooijman, S.; Zhou, E.; Chung, C.K.; Gao, Y.; van den Heuvel, J.K.; Meijer, O.C.; Berbée, J.F. Butyrate reduces appetite and activates brown adipose tissue via the gut-brain neural circuit. Gut 2018, 67, 1269–1279. [Google Scholar] [CrossRef] [Green Version]
- Hu, J.; Kyrou, I.; Tan, B.K.; Dimitriadis, G.K.; Ramanjaneya, M.; Tripathi, G.; Patel, V.; James, S.; Kawan, M.; Chen, J. Short-chain fatty acid acetate stimulates adipogenesis and mitochondrial biogenesis via GPR43 in brown adipocytes. Endocrinology 2016, 157, 1881–1894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weitkunat, K.; Stuhlmann, C.; Postel, A.; Rumberger, S.; Fankhänel, M.; Woting, A.; Petzke, K.J.; Gohlke, S.; Schulz, T.J.; Blaut, M. Short-chain fatty acids and inulin, but not guar gum, prevent diet-induced obesity and insulin resistance through differential mechanisms in mice. Sci. Rep. 2017, 7, 6109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, Y.; Fan, C.; Li, P.; Lu, Y.; Chang, X.; Qi, K. Short chain fatty acids prevent high-fat-diet-induced obesity in mice by regulating G protein-coupled receptors and gut microbiota. Sci. Rep. 2016, 6, 37589. [Google Scholar] [CrossRef] [Green Version]
- Han, X.; Guo, J.; You, Y.; Yin, M.; Liang, J.; Ren, C.; Zhan, J.; Huang, W. Vanillic acid activates thermogenesis in brown and white adipose tissue. Food Funct. 2018, 9, 4366–4375. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Furuzono, T.; Yamakuni, K.; Li, Y.; Kim, Y.I.; Takahashi, H.; Ohue-Kitano, R.; Jheng, H.F.; Takahashi, N.; Kano, Y. 10-oxo-12 (Z)-octadecenoic acid, a linoleic acid metabolite produced by gut lactic acid bacteria, enhances energy metabolism by activation of TRPV1. FASEB J. 2017, 31, 5036–5048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moradi, M.; Kousheh, S.A.; Almasi, H.; Alizadeh, A.; Guimarães, J.T.; Yılmaz, N.; Lotfi, A. Postbiotics produced by lactic acid bacteria: The next frontier in food safety. Compr. Rev. Food Sci. Food Saf. 2020, 19, 3390–3415. [Google Scholar] [CrossRef]
- Heeney, D.D.; Zhai, Z.; Bendiks, Z.; Barouei, J.; Martinic, A.; Slupsky, C.; Marco, M.L. Lactobacillus plantarum bacteriocin is associated with intestinal and systemic improvements in diet-induced obese mice and maintains epithelial barrier integrity in vitro. Gut Microbes 2019, 10, 382–397. [Google Scholar] [CrossRef] [Green Version]
- Bai, L.; Kumar, S.; Verma, S.; Seshadri, S. Bacteriocin PJ4 from probiotic lactobacillus reduced adipokine and inflammasome in high fat diet induced obesity. 3 Biotech 2020, 10, 355. [Google Scholar] [CrossRef]
- Cao, S.-Y.; Zhao, C.-N.; Xu, X.-Y.; Tang, G.-Y.; Corke, H.; Gan, R.-Y.; Li, H.-B. Dietary plants, gut microbiota, and obesity: Effects and mechanisms. Trends Food Sci. Technol. 2019, 92, 194–204. [Google Scholar] [CrossRef]
- Ryu, S.; Kyoung, H.; Park, K.I.; Oh, S.; Song, M.; Kim, Y. Postbiotic heat-killed lactobacilli modulates on body weight associated with gut microbiota in a pig model. AMB Express 2022, 12, 83. [Google Scholar] [CrossRef]
- Sakai, T.; Taki, T.; Nakamoto, A.; Shuto, E.; Tsutsumi, R.; Toshimitsu, T.; Makino, S.; Ikegami, S. Lactobacillus plantarum OLL2712 regulates glucose metabolism in C57BL/6 mice fed a high-fat diet. J. Nutr. Sci. Vitaminol. 2013, 59, 144–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshitake, R.; Hirose, Y.; Murosaki, S.; Matsuzaki, G. Heat-killed Lactobacillus plantarum L-137 attenuates obesity and associated metabolic abnormalities in C57BL/6 J mice on a high-fat diet. Biosci. Microbiota Food Health 2021, 40, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Kikuchi, K.; Ben Othman, M.; Sakamoto, K. Sterilized bifidobacteria suppressed fat accumulation and blood glucose level. Biochem. Biophys. Res. Commun. 2018, 501, 1041–1047. [Google Scholar] [CrossRef]
- Othman, M.B.; Sakamoto, K. Effect of inactivated Bifidobacterium longum intake on obese diabetes model mice (TSOD). Food Res. J. 2020, 129, 108792. [Google Scholar] [CrossRef]
- Seo, K.-H.; Jeong, J.; Kim, H. Synergistic effects of heat-killed kefir paraprobiotics and flavonoid-rich prebiotics on western diet-induced obesity. Nutrients 2020, 12, 2465. [Google Scholar] [CrossRef]
- Kim, H.; Lim, J.-J.; Shin, H.Y.; Suh, H.J.; Choi, H.-S. Lactobacillus plantarum K8-based paraprobiotics suppress lipid accumulation during adipogenesis by the regulation of JAK/STAT and AMPK signaling pathways. J. Funct. Foods 2021, 87, 104824. [Google Scholar] [CrossRef]
- Lim, J.-J.; Jung, A.-H.; Suh, H.J.; Choi, H.-S.; Kim, H. Lactiplantibacillus plantarum K8-based paraprobiotics prevents obesity and obesity-induced inflammatory responses in high fat diet-fed mice. Food Res. Int. 2022, 155, 111066. [Google Scholar] [CrossRef]
- Jang, K.O.; Choi, J.S.; Choi, K.H.; Kim, S.; Kim, H.; Chung, D.K. Anti-obesity potential of heat-killed Lactiplantibacillus plantarum K8 in 3T3-L1 cells and high-fat diet mice. Heliyon 2023, 9, e12926. [Google Scholar] [CrossRef]
- Watanabe, J.; Hashimoto, N.; Yin, T.; Sandagdorj, B.; Arakawa, C.; Inoue, T.; Suzuki, S. Heat-killed Lactobacillus brevis KB290 attenuates visceral fat accumulation induced by high-fat diet in mice. J. Appl. Microbiol. 2021, 131, 1998–2009. [Google Scholar] [CrossRef]
- Chambers, E.S.; Viardot, A.; Psichas, A.; Morrison, D.J.; Murphy, K.G.; Zac-Varghese, S.E.; MacDougall, K.; Preston, T.; Tedford, C.; Finlayson, G.S. Effects of targeted delivery of propionate to the human colon on appetite regulation, body weight maintenance and adiposity in overweight adults. Gut 2015, 64, 1744–1754. [Google Scholar] [CrossRef] [Green Version]
- van der Beek, C.M.; Canfora, E.E.; Lenaerts, K.; Troost, F.J.; Olde Damink, S.W.; Holst, J.J.; Masclee, A.A.; Dejong, C.H.; Blaak, E.E. Distal, not proximal, colonic acetate infusions promote fat oxidation and improve metabolic markers in overweight/obese men. Clin. Sci. 2016, 130, 2073–2082. [Google Scholar] [CrossRef] [PubMed]
- Canfora, E.E.; van der Beek, C.M.; Jocken, J.W.; Goossens, G.H.; Holst, J.J.; Olde Damink, S.W.; Lenaerts, K.; Dejong, C.H.; Blaak, E.E. Colonic infusions of short-chain fatty acid mixtures promote energy metabolism in overweight/obese men: A randomized crossover trial. Sci. Rep. 2017, 7, 2360. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Higashikawa, F.; Noda, M.; Kawamura, Y.; Matoba, Y.; Kumagai, T.; Sugiyama, M. The obesity and fatty liver are reduced by plant-derived Pediococcus pentosaceus LP28 in high fat diet-induced obese mice. PLoS ONE 2012, 7, e30696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Higashikawa, F.; Noda, M.; Awaya, T.; Danshiitsoodol, N.; Matoba, Y.; Kumagai, T.; Sugiyama, M. Antiobesity effect of Pediococcus pentosaceus LP28 on overweight subjects: A randomized, double-blind, placebo-controlled clinical trial. Eur. J. Clin. Nutr. 2016, 70, 582–587. [Google Scholar] [CrossRef]
- Nakamura, F.; Ishida, Y.; Aihara, K.; Sawada, D.; Ashida, N.; Sugawara, T.; Aoki, Y.; Takehara, I.; Takano, K.; Fujiwara, S. Effect of fragmented Lactobacillus amylovorus CP1563 on lipid metabolism in overweight and mildly obese individuals: A randomized controlled trial. Microb. Ecol. Health Dis. 2016, 27, 30312. [Google Scholar]
- Sugawara, T.; Sawada, D.; Yanagihara, S.; Aoki, Y.; Takehara, I.; Sugahara, H.; Hirota, T.; Nakamura, Y.; Ishikawa, S. Daily intake of paraprobiotic Lactobacillus amylovorus CP1563 improves pre-obese conditions and affects the gut microbial community in healthy pre-obese subjects: A double-blind, randomized, placebo-controlled study. Microorganisms 2020, 8, 304. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Chen, J.; Tan, Q.; Deng, X.; Tsai, P.-J.; Chen, P.-H.; Ye, M.; Guo, J.; Su, Z. Nondigestible oligosaccharides with anti-obesity effects. J. Agric. Food Chem. 2019, 68, 4–16. [Google Scholar] [CrossRef]
- Laurindo, L.F.; Barbalho, S.M.; Guiguer, E.L.; da Silva Soares de Souza, M.; de Souza, G.A.; Fidalgo, T.M.; Araújo, A.C.; de Souza Gonzaga, H.F.; de Bortoli Teixeira, D.; de Oliveira Silva Ullmann, T. GLP-1a: Going beyond traditional use. Int. J. Mol. Sci. 2022, 23, 739. [Google Scholar] [CrossRef]
- Kim, T.T.; Parajuli, N.; Sung, M.M.; Bairwa, S.C.; Levasseur, J.; Soltys, C.-L.M.; Wishart, D.S.; Madsen, K.; Schertzer, J.D.; Dyck, J.R. Fecal transplant from resveratrol-fed donors improves glycaemia and cardiovascular features of the metabolic syndrome in mice. Am. J. Physiol. Endocrinol. Metab. 2018, 315, E511–E519. [Google Scholar] [CrossRef]
- Aguilar-Toalá, J.E.; Hall, F.G.; Urbizo-Reyes, U.C.; Garcia, H.S.; Vallejo-Cordoba, B.; González-Córdova, A.F.; Hernández-Mendoza, A.; Liceaga, A.M. In silico prediction and in vitro assessment of multifunctional properties of postbiotics obtained from two probiotic bacteria. Probiotics Antimicrob. Proteins 2020, 12, 608–622. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, S.-J.; Sharma, A.; Lee, H.-J. Postbiotics against Obesity: Perception and Overview Based on Pre-Clinical and Clinical Studies. Int. J. Mol. Sci. 2023, 24, 6414. https://doi.org/10.3390/ijms24076414
Park S-J, Sharma A, Lee H-J. Postbiotics against Obesity: Perception and Overview Based on Pre-Clinical and Clinical Studies. International Journal of Molecular Sciences. 2023; 24(7):6414. https://doi.org/10.3390/ijms24076414
Chicago/Turabian StylePark, Seon-Joo, Anshul Sharma, and Hae-Jeung Lee. 2023. "Postbiotics against Obesity: Perception and Overview Based on Pre-Clinical and Clinical Studies" International Journal of Molecular Sciences 24, no. 7: 6414. https://doi.org/10.3390/ijms24076414