
Short, Lipidated Dendrimeric γ-AApeptides as New Antimicrobial Peptidomimetics
 ,
,
Abstract
:1. Introduction
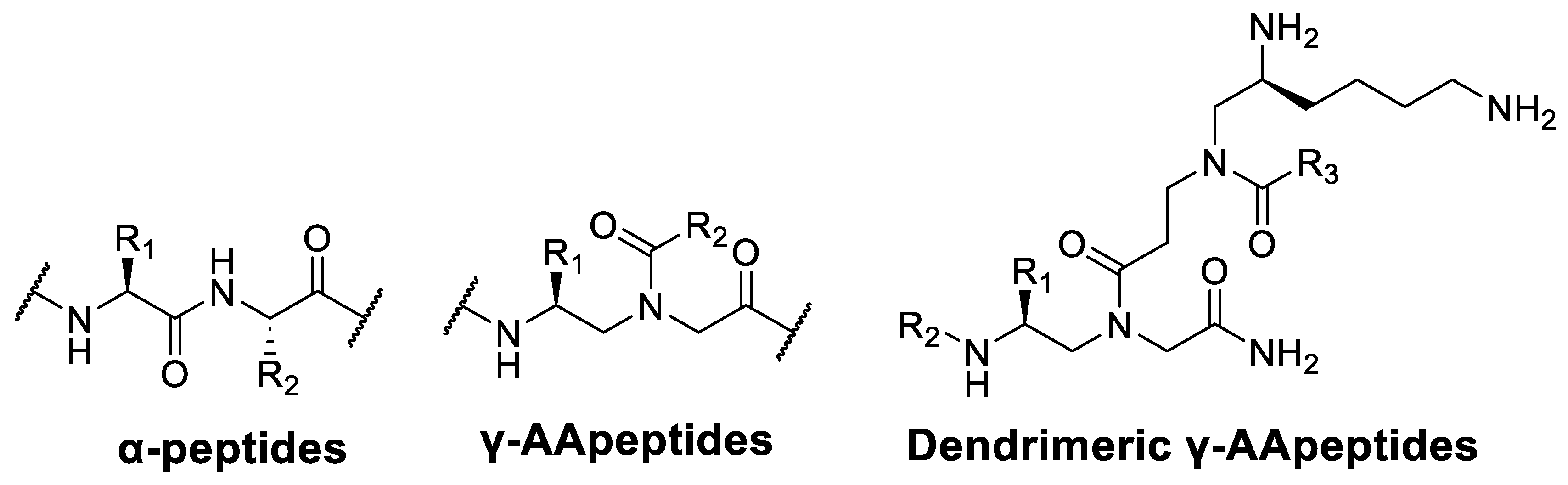
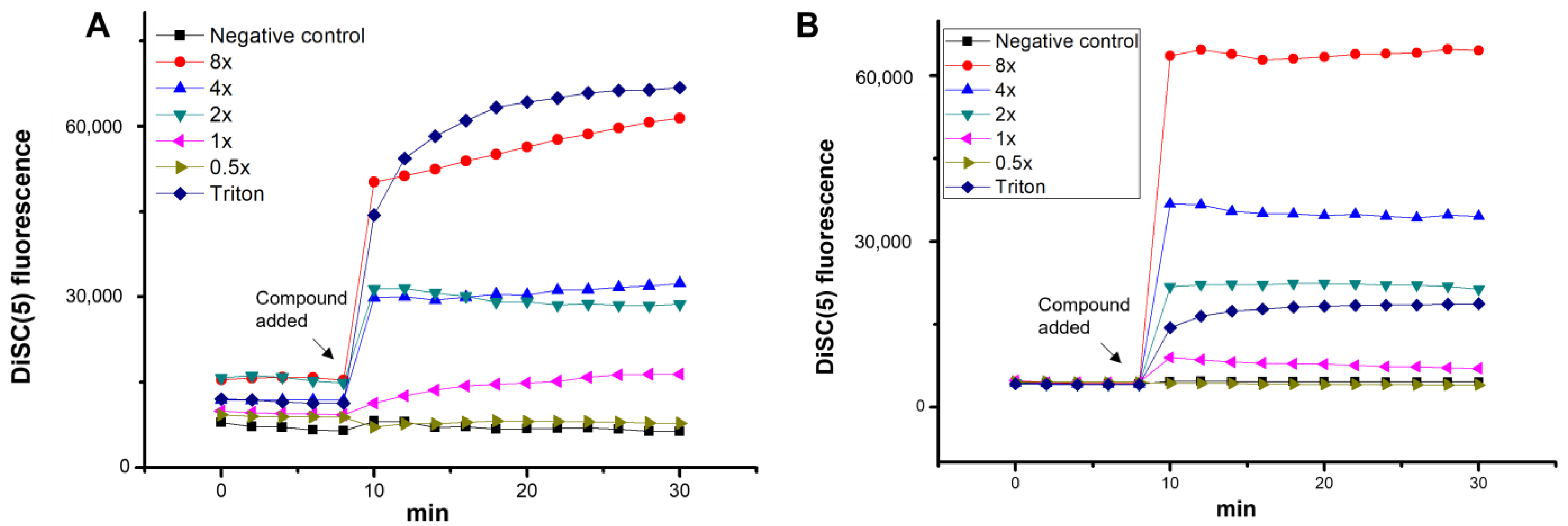
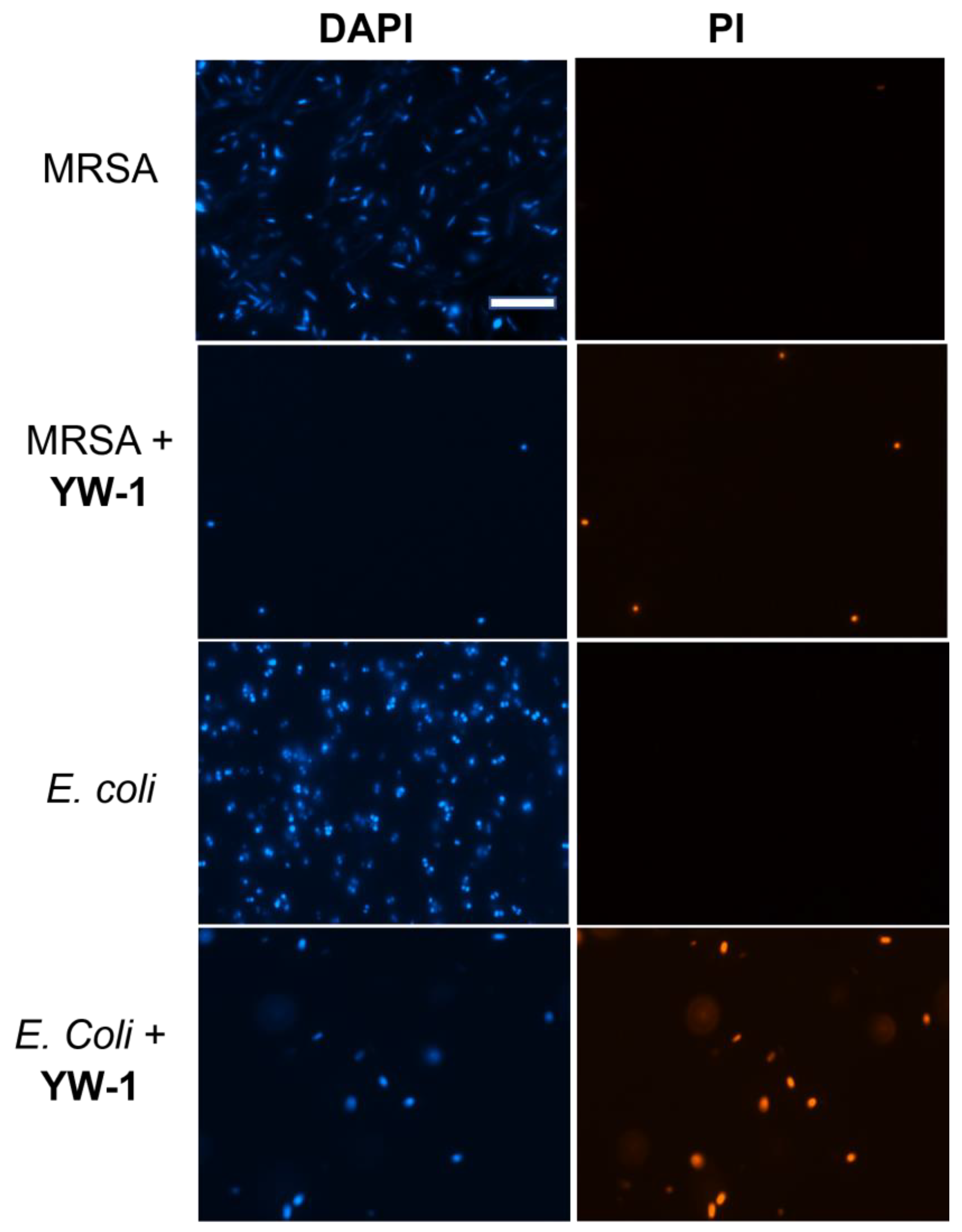
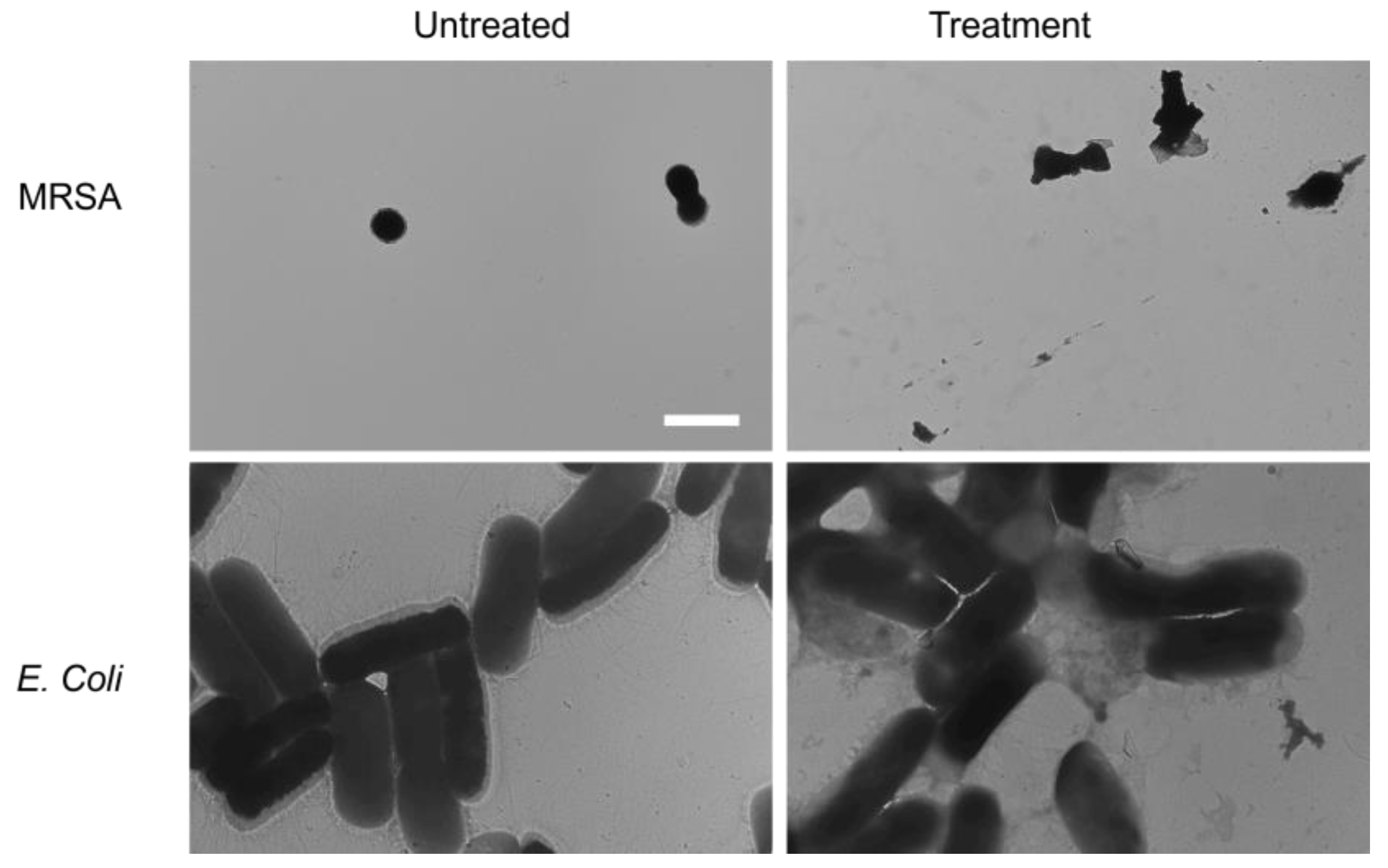
2. Results and Discussion
3. Material and Methods
3.1. Membrane Depolarization Study
3.2. Outer Membrane (OM) Permeabilization
3.3. Inner Membrane (IM) Permeabilization
3.4. Fluorescence Microscopy
3.5. Transmission Electron Microscopy (TEM)
3.6. Bacterial Killing Efficiency
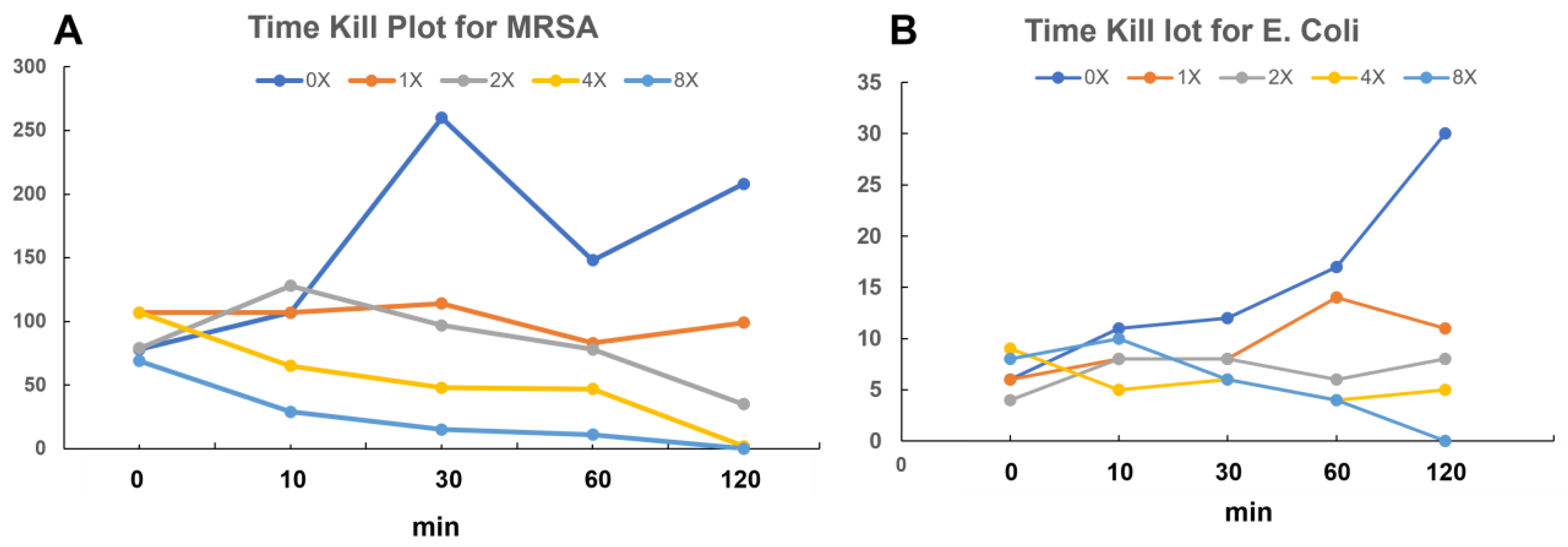
3.6.1. Time-Kill Kinetics Study
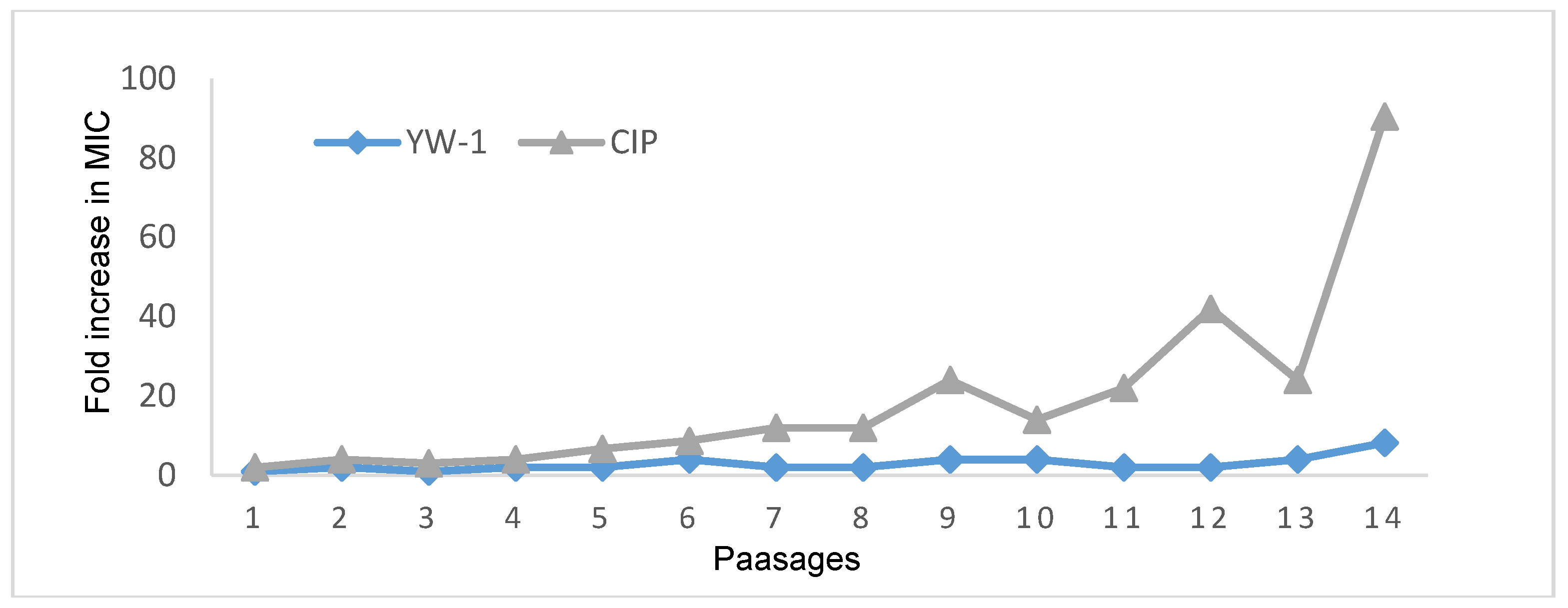
3.6.2. Drug Resistance Test
3.7. Experiment Procedure
3.7.1. General Experiment Methods
3.7.2. MIC (Minimum Inhibitory Concentration)
3.7.3. Hemolytic Activity
3.7.4. Drug Resistance Study
3.7.5. TEM
3.7.6. Inner Membrane Permeability
3.7.7. Fluorescence Microscopy
3.7.8. Time-Kill Kinetics Study
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- US Centers for Disease Control. Antibiotic Resistance Threats in the United States; US Department of Health and Human Services: Washington, DC, USA, 2019. [Google Scholar] [CrossRef] [Green Version]
- Hancock, R.E.W.; Sahl, H.G. Antimicrobial and Host-Defense Peptides as New Anti-Infective Therapeutic Strategies. Nat. Biotechnol. 2006, 24, 1551–1557. [Google Scholar] [CrossRef]
- Niu, Y.; Padhee, S.; Wu, H.; Bai, G.; Qiao, Q.; Hu, Y.; Harrington, L.; Burda, W.N.; Shaw, L.N.; Cao, C.; et al. Peptides as the next Generation of Anti-Infectives. Future Med. Chem. 2013, 5, 315–337. [Google Scholar] [CrossRef]
- Chongsiriwatana, N.P.; Patch, J.A.; Czyzewski, A.M.; Dohm, M.T.; Ivankin, A.; Gidalevitz, D.; Zuckermann, R.N.; Barron, A.E. Peptoids That Mimic the Structure, Function, and Mechanism of Helical Antimicrobial Peptides. Proc. Natl. Acad. Sci. USA 2008, 105, 2794–2799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Porter, E.A.; Weisblum, B.; Gellman, S.H. Mimicry of Host-Defense Peptides by Unnatural Oligomers: Antimicrobial β-Peptides. J. Am. Chem. Soc. 2002, 124, 7324–7330. [Google Scholar] [CrossRef] [PubMed]
- Niu, Y.; Padhee, S.; Wu, H.; Bai, G.; Qiao, Q.; Hu, Y.; Harrington, L.; Burda, W.N.; Shaw, L.N.; Cao, C.; et al. Lipo-γ-AApeptides as a New Class of Potent and Broad-Spectrum Antimicrobial Agents. J. Med. Chem. 2012, 55, 4003–4009. [Google Scholar] [CrossRef] [PubMed]
- Claudon, P.; Violette, A.; Lamour, K.; Decossas, M.; Fournel, S.; Heurtault, B.; Godet, J.; Mély, Y.; Jamart-Grégoire, B.; Averlant-Petit, M. Consequences of Isostructural Main-chain Modifications for the Design of Antimicrobial Foldamers: Helical Mimics of Host-defense Peptides Based on a Heterogeneous Amide/Urea Backbone. Angew. Chem. Int. Ed. 2010, 49, 333–336. [Google Scholar] [CrossRef] [PubMed]
- Violette, A.; Fournel, S.; Lamour, K.; Chaloin, O.; Frisch, B.; Briand, J.-P.; Monteil, H.; Guichard, G. Mimicking Helical Antibacterial Peptides with Nonpeptidic Folding Oligomers. Chem. Biol. 2006, 13, 531–538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsubery, H.; Ofek, I.; Cohen, S.; Fridkin, M. N-Terminal Modifications of Polymyxin B Nonapeptide and Their Effect on Antibacterial Activity. Peptides 2001, 22, 1675–1681. [Google Scholar] [CrossRef]
- Steenbergen, J.N.; Alder, J.; Thorne, G.M.; Tally, F.P. Daptomycin: A Lipopeptide Antibiotic for the Treatment of Serious Gram-Positive Infections. J. Antimicrob. Chemother. 2005, 55, 283–288. [Google Scholar] [CrossRef] [Green Version]
- Lockwood, N.A.; Haseman, J.R.; Tirrell, M.V.; Mayo, K.H.; Mak, P.; Pohl, J.; Dubin, A.; Reed, M.S.; Bowers, S.E.; Fallon, M.T.; et al. Acylation of SC4 Dodecapeptide Increases Bactericidal Potency against Gram-Positive Bacteria, Including Drug-Resistant Strains. Int. J. Antimicrob. Agents. 2003, 42, 14946–14956. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Amin, M.N.; Padhee, S.; Wang, R.E.; Qiao, Q.; Bai, G.; Li, Y.; Mathew, A.; Cao, C.; Cai, J. Lipidated Peptidomimetics with Improved Antimicrobial Activity. ACS. Med. Chem. Lett. 2012, 3, 683–686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Z.; He, S.; Wang, J.; Yang, Y.; Zhang, L.; Li, Y.; Shan, A. Rational Design of Short Peptide Variants by Using Kunitzin-RE, an Amphibian-Derived Bioactivity Peptide, for Acquired Potent Broad-Spectrum Antimicrobial and Improved Therapeutic Potential of Commensalism Coinfection of Pathogens. J. Med. Chem. 2019, 62, 4586–4605. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Gao, R.; Zheng, M.; Sang, P.; Li, C.; Zhang, E.; Li, Q.; Cai, J. Development of Bis-Cyclic Imidazolidine-4-One Derivatives as Potent Antibacterial Agents. J. Med. Chem. 2020, 63, 15591–15602. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | MIC (μg/mL) | HC50 (μg/mL) | Selectivity Index (HC50/MICMRSA) | |||||
|---|---|---|---|---|---|---|---|---|
| Gram-Positive (+) | Gram-Negative (−) | |||||||
| MRSA | E.F. | MRSE | P.A. | K.P. | E. coli | |||
| YW-1 | 1.5–3 | 1.5–3 | 1.5–3 | 6–12.5 | 12.5–25 | 3.0–6.0 | >125 | >41.7 |
| YW-2 | 1.5–3 | 1.5–3 | 1.5–3 | 12.5–25 | 6–12.5 | 3.0–6.0 | >125 | >41.7 |
| YW-3 | 0.75–1.5 | 3.0–6.0 | 6–12.5 | 6–12.5 | >25 | 3.0–6.0 | >125 | >83.3 |
| YW-4 | 0.75–1.5 | 3.0–6.0 | 3.0–6.0 | >25 | >25 | 3.0–6.0 | >125 | >83.3 |
| YW-5 | 1.5–3 | 6–12.5 | >125 | >41.7 | ||||
| YW-6 | 0.75–1.5 | 6–12.5 | >125 | >83.3 | ||||
| YW-7 | 0.75–1.5 | 3.0–6.0 | 3.0–6.0 | 12.5–25 | >25 | 3.0–6.0 | >125 | >83.3 |
| YW-8 | 12.5–25 | >25 | ||||||
| YW-9 | >25 | >25 | ||||||
| YW-10 | >25 | >25 | ||||||
| YW-11 | >25 | --- | --- | --- | --- | >25 | --- | --- |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Xue, M.; Gao, R.; Chakraborty, S.; Wang, S.; Zhao, X.; Gu, M.; Cao, C.; Sun, X.; Cai, J. Short, Lipidated Dendrimeric γ-AApeptides as New Antimicrobial Peptidomimetics. Int. J. Mol. Sci. 2023, 24, 6407. https://doi.org/10.3390/ijms24076407
Wang Y, Xue M, Gao R, Chakraborty S, Wang S, Zhao X, Gu M, Cao C, Sun X, Cai J. Short, Lipidated Dendrimeric γ-AApeptides as New Antimicrobial Peptidomimetics. International Journal of Molecular Sciences. 2023; 24(7):6407. https://doi.org/10.3390/ijms24076407
Chicago/Turabian StyleWang, Yafeng, Menglin Xue, Ruixuan Gao, Soumyadeep Chakraborty, Shaohui Wang, Xue Zhao, Meng Gu, Chuanhai Cao, Xingmin Sun, and Jianfeng Cai. 2023. "Short, Lipidated Dendrimeric γ-AApeptides as New Antimicrobial Peptidomimetics" International Journal of Molecular Sciences 24, no. 7: 6407. https://doi.org/10.3390/ijms24076407