
Reprogramming the Circadian Dynamics of Epileptic Genes in Mouse Temporal Lobe Epilepsy

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
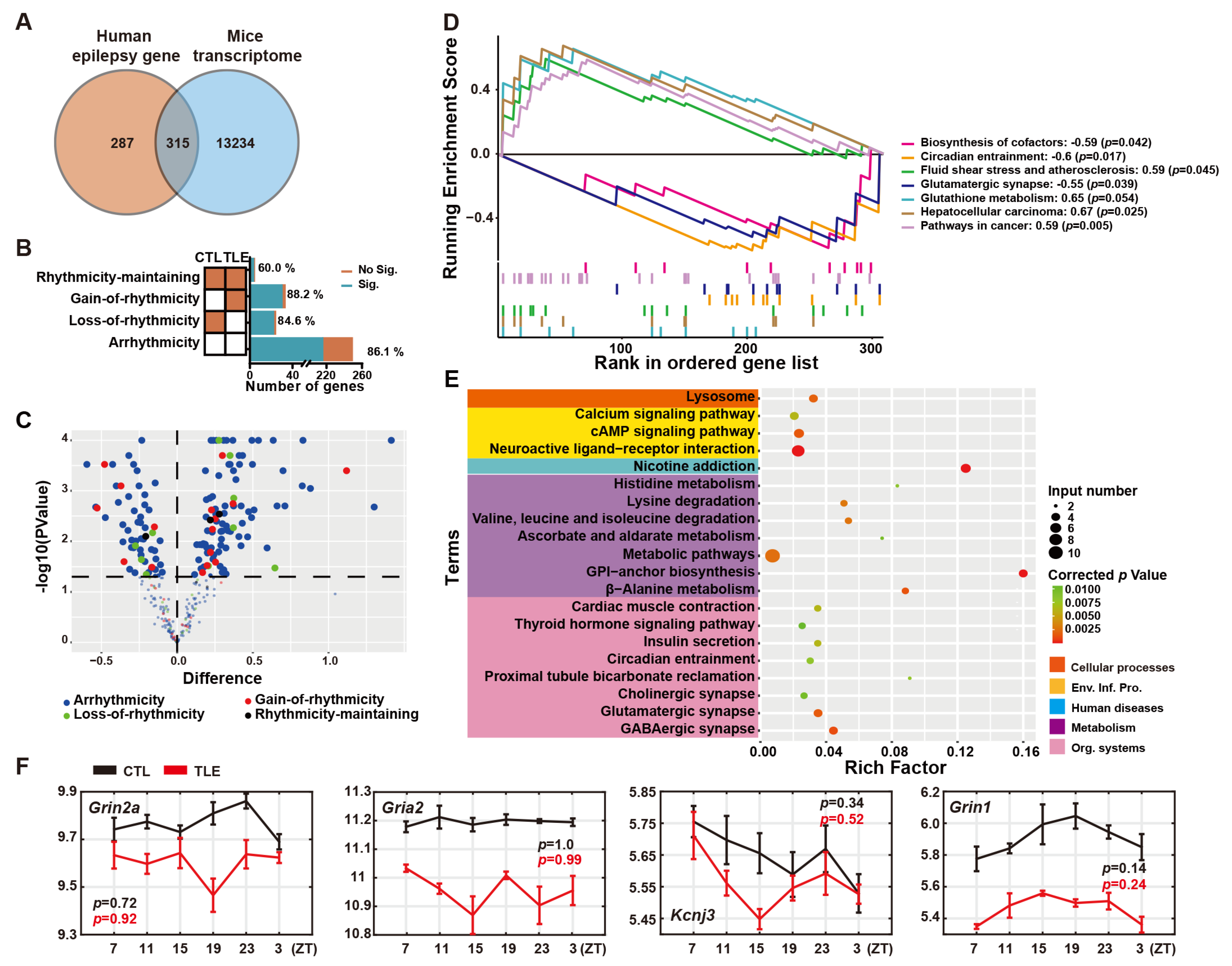
2.1. Epilepsy-Related Genes in the Ventral Hippocampal Transcriptome of Mice
2.2. Arrhythmic Epilepsy-Related Genes in TLE Mice
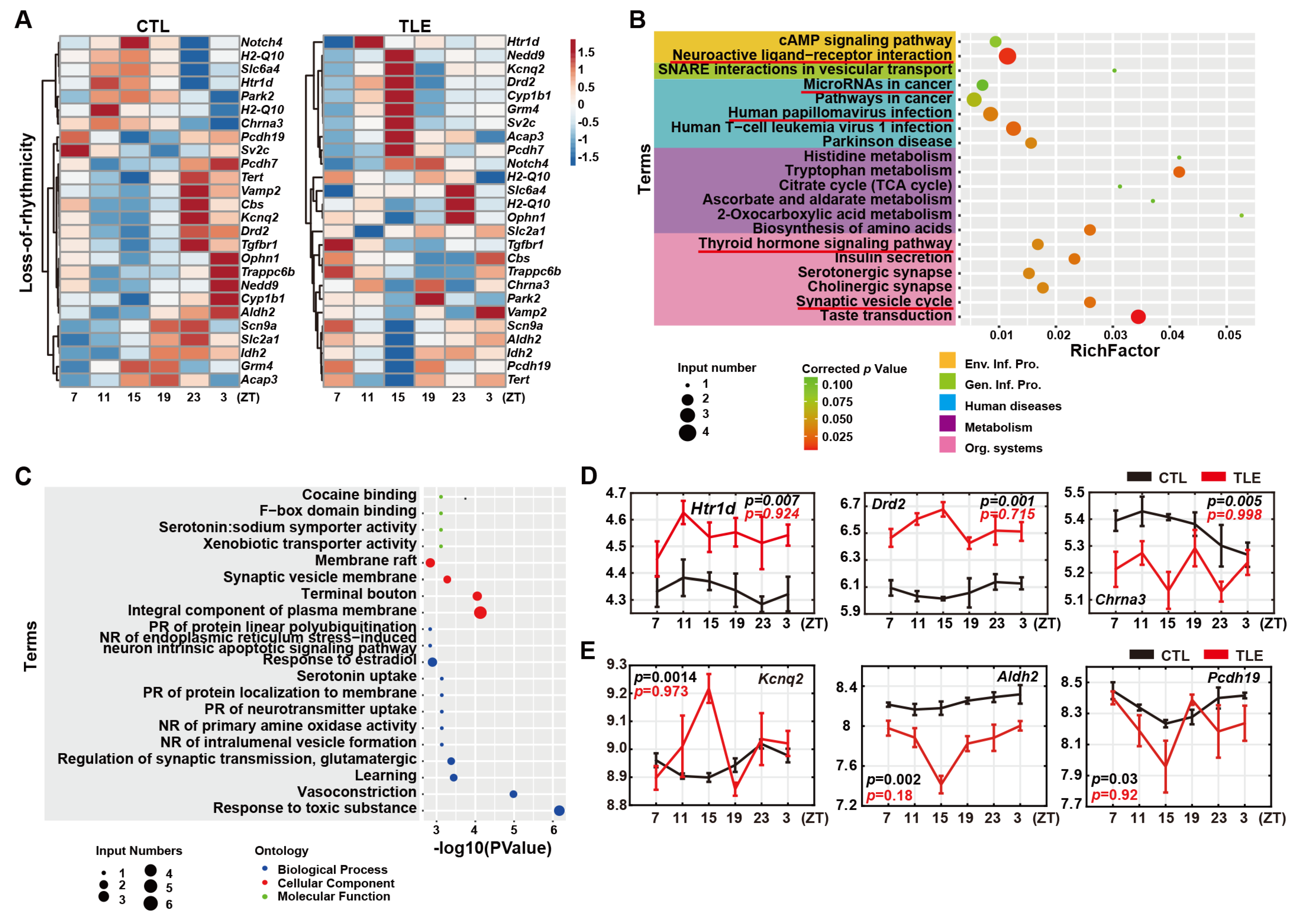
2.3. Loss-Of-Rhythmicity Epileptic Genes in TLE Mice
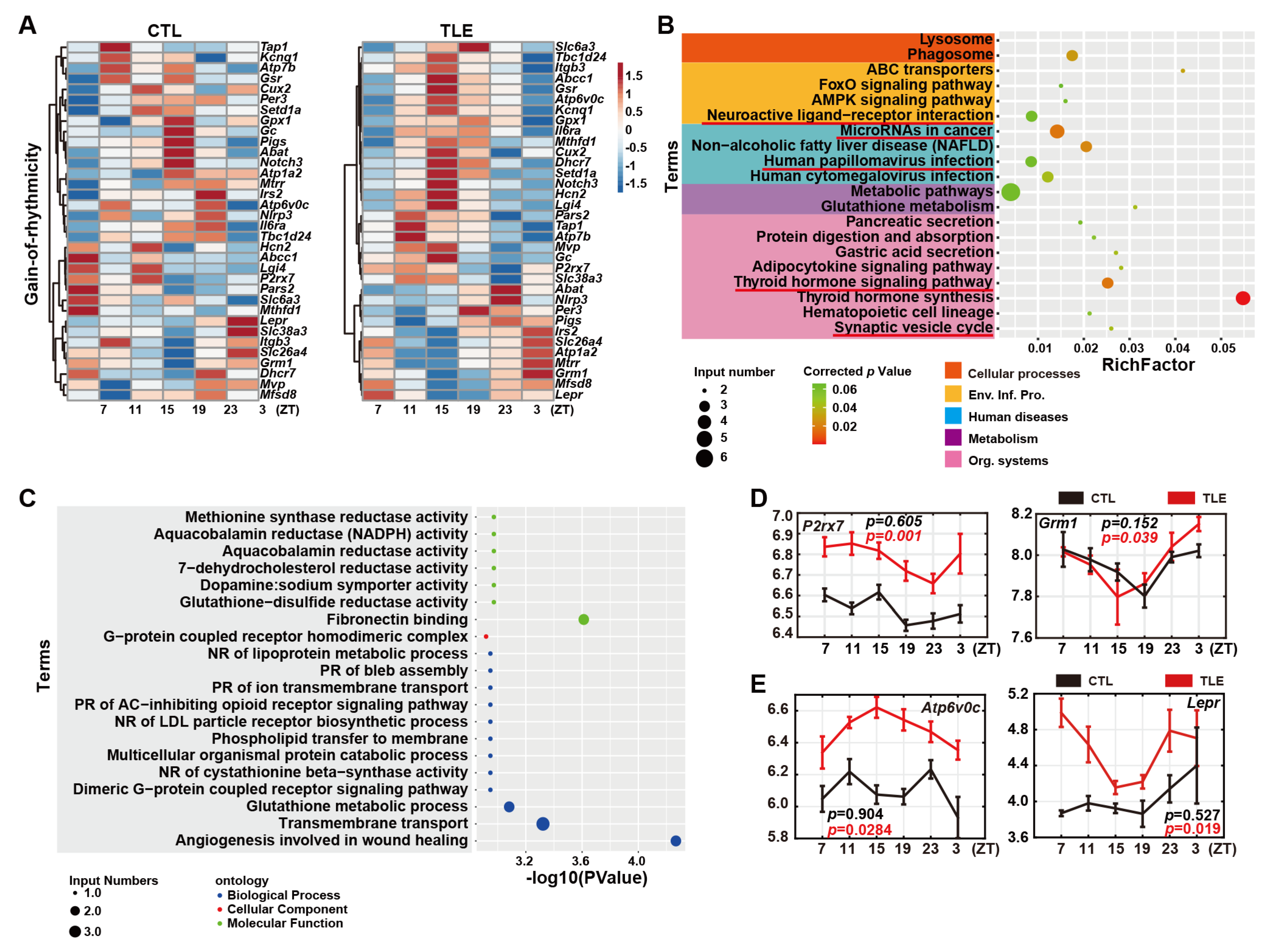
2.4. Gain-Of-Rhythmicity Epileptic Genes in TLE Mice
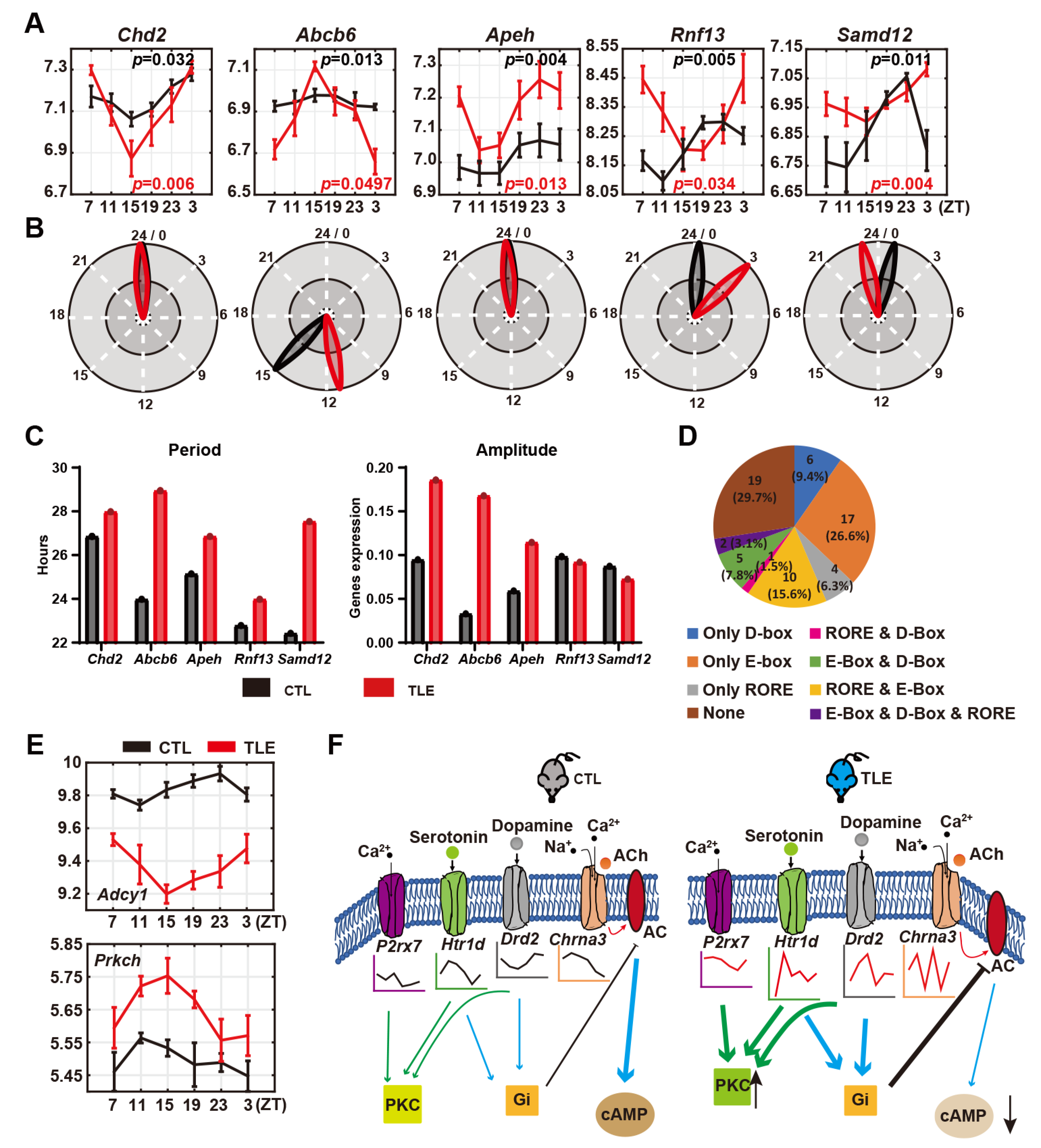
2.5. Rhythmicity-Maintaining Epileptic Genes in TLE and CTL Mice
2.6. Possible Circadian Regulation of Rhythmically Expressed Epileptic Genes in Mice
3. Discussion
4. Materials and Methods
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Engel, J., Jr. A proposed diagnostic scheme for people with epileptic seizures and with epilepsy: Report of the ILAE Task Force on Classification and Terminology. Epilepsia 2001, 42, 796–803. [Google Scholar] [CrossRef]
- Mirzoev, A.; Bercovici, E.; Stewart, L.S.; Cortez, M.A.; Snead, O.C., 3rd; Desrocher, M. Circadian profiles of focal epileptic seizures: A need for reappraisal. Seizure 2012, 21, 412–416. [Google Scholar] [CrossRef] [Green Version]
- Dębski, K.; Ceglia, N.; Ghestem, A.; Ivanov, A.; Brancati, G.; Bröer, S.; Bot, A.; Müller, J.; Becker, A.; Löscher, W.; et al. The circadian dynamics of the hippocampal transcriptome and proteome is altered in experimental temporal lobe epilepsy. Sci. Adv. 2020, 6, eaat5979. [Google Scholar] [CrossRef]
- Hofstra, W.A.; Spetgens, W.P.; Leijten, F.S.; van Rijen, P.C.; Gosselaar, P.; van der Palen, J.; de Weerd, A.W. Diurnal rhythms in seizures detected by intracranial electrocorticographic monitoring: An observational study. Epilepsy Behav. 2009, 14, 617–621. [Google Scholar] [CrossRef]
- Zhang, T.; Yu, F.; Xu, H.; Chen, M.; Chen, X.; Guo, L.; Zhou, C.; Xu, Y.; Wang, F.; Yu, J.; et al. Dysregulation of REV-ERBα impairs GABAergic function and promotes epileptic seizures in preclinical models. Nat. Commun. 2021, 12, 1216. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Fu, X.; Smith, N.A.; Ziobro, J.; Curiel, J.; Tenga, M.J.; Martin, B.; Freedman, S.; Cea-Del Rio, C.A.; Oboti, L.; et al. Loss of CLOCK Results in Dysfunction of Brain Circuits Underlying Focal Epilepsy. Neuron 2017, 96, 387–401.e386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, S.; Wang, H. Clocking Epilepsies: A Chronomodulated Strategy-Based Therapy for Rhythmic Seizures. Int. J. Mol. Sci. 2023, 24, 4223. [Google Scholar] [CrossRef] [PubMed]
- Zeng, H.; Jia, J.; Liu, A. Coordinated translocation of mammalian Gli proteins and suppressor of fused to the primary cilium. PLoS ONE 2010, 5, e15900. [Google Scholar] [CrossRef] [Green Version]
- Masson, J.; Emerit, M.B.; Hamon, M.; Darmon, M. Serotonergic signaling: Multiple effectors and pleiotropic effects. Wiley Interdiscip. Rev. Membr. Transp. Signal. 2012, 1, 685–713. [Google Scholar] [CrossRef]
- Brandon, N.J.; Jovanovic, J.N.; Colledge, M.; Kittler, J.T.; Brandon, J.M.; Scott, J.D.; Moss, S.J. A-kinase anchoring protein 79/150 facilitates the phosphorylation of GABAA receptors by cAMP-dependent protein kinase via selective interaction with receptor β subunits. Mol. Cell. Neurosci. 2003, 22, 87–97. [Google Scholar] [CrossRef]
- Chen, C.; Yang, J.M.; Hu, T.T.; Xu, T.J.; Xu, W.P.; Wei, W. Elevated dopamine D2 receptor in prefrontal cortex of CUMS rats is associated with downregulated cAMP-independent signaling pathway. Can. J. Physiol. Pharmacol. 2013, 91, 750–758. [Google Scholar] [CrossRef]
- Jijón-Lorenzo, R.; Caballero-Florán, I.H.; Recillas-Morales, S.; Cortés, H.; Avalos-Fuentes, J.A.; Paz-Bermúdez, F.J.; Erlij, D.; Florán, B. Presynaptic Dopamine D2 Receptors Modulate [(3)H]GABA Release at StriatoPallidal Terminals via Activation of PLC → IP3 → Calcineurin and Inhibition of AC → cAMP → PKA Signaling Cascades. Neuroscience 2018, 372, 74–86. [Google Scholar] [CrossRef] [PubMed]
- Carlson, A.B.; Kraus, G.P. Physiology, Cholinergic Receptors. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2022. [Google Scholar]
- Bläsche, R.; Ebeling, G.; Perike, S.; Weinhold, K.; Kasper, M.; Barth, K. Activation of P2X7R and downstream effects in bleomycin treated lung epithelial cells. Int. J. Biochem. Cell Biol. 2012, 44, 514–524. [Google Scholar] [CrossRef]
- Ayoub, M.A.; Angelicheva, D.; Vile, D.; Chandler, D.; Morar, B.; Cavanaugh, J.A.; Visscher, P.M.; Jablensky, A.; Pfleger, K.D.; Kalaydjieva, L. Deleterious GRM1 mutations in schizophrenia. PLoS ONE 2012, 7, e32849. [Google Scholar] [CrossRef] [Green Version]
- Chu, H.; Sun, P.; Yin, J.; Liu, G.; Wang, Y.; Zhao, P.; Zhu, Y.; Yang, X.; Zheng, T.; Zhou, X.; et al. Integrated network analysis reveals potentially novel molecular mechanisms and therapeutic targets of refractory epilepsies. PLoS ONE 2017, 12, e0174964. [Google Scholar] [CrossRef] [Green Version]
- Frank, M.; Kemler, R. Protocadherins. Curr. Opin. Cell Biol. 2002, 14, 557–562. [Google Scholar] [CrossRef]
- Dibbens, L.M.; Tarpey, P.S.; Hynes, K.; Bayly, M.A.; Scheffer, I.E.; Smith, R.; Bomar, J.; Sutton, E.; Vandeleur, L.; Shoubridge, C.; et al. X-linked protocadherin 19 mutations cause female-limited epilepsy and cognitive impairment. Nat. Genet. 2008, 40, 776–781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoshina, N.; Johnson-Venkatesh, E.M.; Hoshina, M.; Umemori, H. Female-specific synaptic dysfunction and cognitive impairment in a mouse model of PCDH19 disorder. Science 2021, 372, eaaz3893. [Google Scholar] [CrossRef] [PubMed]
- Mattison, K.A.; Tossing, G.; Mulroe, F.; Simmons, C.; Butler, K.M.; Schreiber, A.; Alsadah, A.; Neilson, D.E.; Naess, K.; Wedell, A.; et al. ATP6V0C variants impair vacuolar V-ATPase causing a neurodevelopmental disorder often associated with epilepsy. Brain A J. Neurol. 2022, awac33. [Google Scholar] [CrossRef]
- Anttila, V.; Bulik-Sullivan, B.; Finucane, H.K.; Walters, R.K.; Bras, J.; Duncan, L.; Escott-Price, V.; Falcone, G.J.; Gormley, P.; Malik, R.; et al. Analysis of shared heritability in common disorders of the brain. Science 2018, 360, eaap8757. [Google Scholar] [CrossRef] [Green Version]
- Bonsall, D.R.; Lall, G.S. Protein kinase C differentially regulates entrainment of the mammalian circadian clock. Chronobiol. Int. 2013, 30, 460–469. [Google Scholar] [CrossRef] [PubMed]
- Lemmer, B.; Barmeier, H.; Schmidt, S.; Lang, P.H. On the daily variation in the beta-receptor-adenylate cyclase-cAMP-phosphodiesterase system in rat forebrain. Chronobiol. Int. 1987, 4, 469–475. [Google Scholar] [CrossRef]
- Osonoe, K.; Ogata, S.; Iwata, Y.; Mori, N. Kindled amygdaloid seizures in rats cause immediate and transient increase in protein kinase C activity followed by transient suppression of the activity. Epilepsia 1994, 35, 850–854. [Google Scholar] [CrossRef]
- Ping, X.; Qin, S.K.; Liu, S.N.; Lu, Y.; Zhao, Y.N.; Cao, Y.F.; Zhang, Y.H.; Zhang, S.D.; Chu, L.; Pei, L. Effects of Huazhuo Jiedu Shugan Decoction on Cognitive and Emotional Disorders in a Rat Model of Epilepsy: Possible Involvement of AC-cAMP-CREB Signaling and NPY Expression. Evid.-Based Complement. Altern. Med. Ecam 2019, 2019, 4352879. [Google Scholar] [CrossRef] [PubMed]
- Arshad, M.N.; Naegele, J.R. Induction of Temporal Lobe Epilepsy in Mice with Pilocarpine. Bio-protocol 2020, 10, e3533. [Google Scholar] [CrossRef]
- Hughes, M.E.; Abruzzi, K.C.; Allada, R.; Anafi, R.; Arpat, A.B.; Asher, G.; Baldi, P.; de Bekker, C.; Bell-Pedersen, D.; Blau, J.; et al. Guidelines for Genome-Scale Analysis of Biological Rhythms. J. Biol. Rhythm. 2017, 32, 380–393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bu, D.; Luo, H.; Huo, P.; Wang, Z.; Zhang, S.; He, Z.; Wu, Y.; Zhao, L.; Liu, J.; Guo, J.; et al. KOBAS-i: Intelligent prioritization and exploratory visualization of biological functions for gene enrichment analysis. Nucleic Acids Res. 2021, 49, W317–W325. [Google Scholar] [CrossRef]
- Wu, G.; Anafi, R.C.; Hughes, M.E.; Kornacker, K.; Hogenesch, J.B. MetaCycle: An integrated R package to evaluate periodicity in large scale data. Bioinformatics 2016, 32, 3351–3353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, T.; Hu, E.; Xu, S.; Chen, M.; Guo, P.; Dai, Z.; Feng, T.; Zhou, L.; Tang, W.; Zhan, L.; et al. clusterProfiler 4.0: A universal enrichment tool for interpreting omics data. Innovation 2021, 2, 100141. [Google Scholar] [CrossRef]
- Zielinski, T.; Moore, A.M.; Troup, E.; Halliday, K.J.; Millar, A.J. Strengths and limitations of period estimation methods for circadian data. PLoS ONE 2014, 9, e96462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raudvere, U.; Kolberg, L.; Kuzmin, I.; Arak, T.; Adler, P.; Peterson, H.; Vilo, J. g:Profiler: A web server for functional enrichment analysis and conversions of gene lists (2019 update). Nucleic Acids Res. 2019, 47, W191–W198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grant, C.E.; Bailey, T.L.; Noble, W.S. FIMO: Scanning for occurrences of a given motif. Bioinformatics 2011, 27, 1017–1018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castro-Mondragon, J.A.; Riudavets-Puig, R.; Rauluseviciute, I.; Lemma, R.B.; Turchi, L.; Blanc-Mathieu, R.; Lucas, J.; Boddie, P.; Khan, A.; Manosalva Pérez, N.; et al. JASPAR 2022: The 9th release of the open-access database of transcription factor binding profiles. Nucleic Acids Res. 2022, 50, D165–D173. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, S.; Wang, H. Reprogramming the Circadian Dynamics of Epileptic Genes in Mouse Temporal Lobe Epilepsy. Int. J. Mol. Sci. 2023, 24, 6400. https://doi.org/10.3390/ijms24076400
Sun S, Wang H. Reprogramming the Circadian Dynamics of Epileptic Genes in Mouse Temporal Lobe Epilepsy. International Journal of Molecular Sciences. 2023; 24(7):6400. https://doi.org/10.3390/ijms24076400
Chicago/Turabian StyleSun, Sha, and Han Wang. 2023. "Reprogramming the Circadian Dynamics of Epileptic Genes in Mouse Temporal Lobe Epilepsy" International Journal of Molecular Sciences 24, no. 7: 6400. https://doi.org/10.3390/ijms24076400