
The Role of Potential Oxidative Biomarkers in the Prognosis of Acute Ischemic Stroke and the Exploration of Antioxidants as Possible Preventive and Treatment Options
 , ,
, ,
Abstract
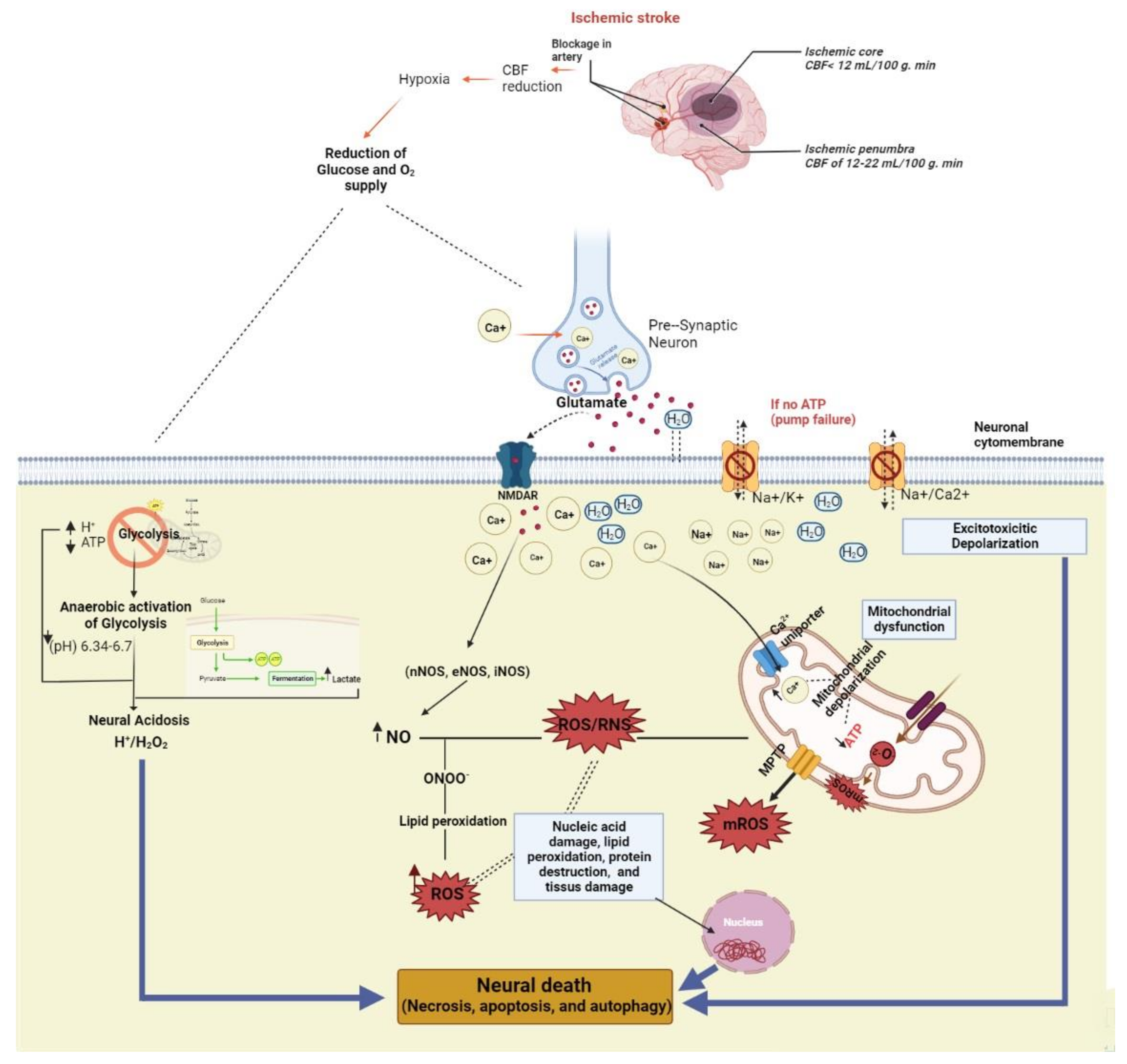
:1. Introduction
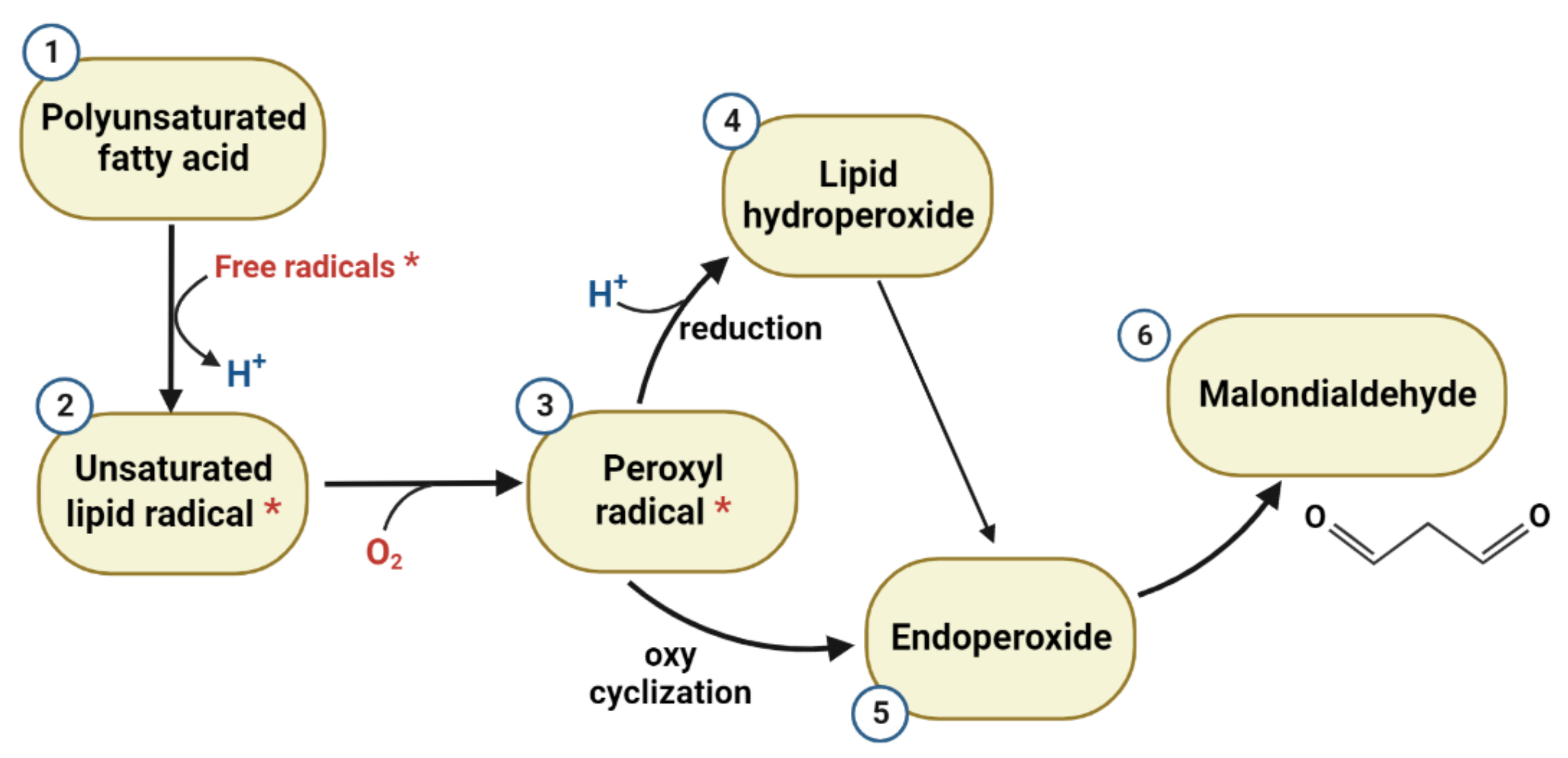
2. Lipid Peroxidation
2.1. Malondialdehyde
2.2. F2-Isoprostanes and 8-Iso-Prostaglandin F2 Alpha
2.3. 4-Hydroxy-2-nonenal
2.4. Oxidized LDL

{kind=link}
{kind=link}
{kind=link}
| Biomarkers | Source Type | Study Design | Concentration | Comment | References |
|---|---|---|---|---|---|
| Lipid peroxidation | |||||
| MDA (µmol/L) | Saliva | 150 individuals: | Salivary and serum MDA levels were significantly higher in the ischemic stroke group than the healthy control and risk group. | [65] | |
| Ischemic stroke, N = 50 | 0.64 ± 0.22 | ||||
| Healthy control, N = 25 | 0.23 ± 0.07 | ||||
| Risk group, N = 75: | |||||
| Hypertension | 0.75 ± 0.21 | ||||
| Type 2 diabetes | 0.65 ± 0.22 | ||||
| Ischemic heart disease | 0.48 ± 0.13 | ||||
| Serum | 150 individuals: | ||||
| Ischemic stroke, N = 50 | 2.51 ± 1.11 | ||||
| Healthy control, N = 25 | 1.12 ± 0.35 | ||||
| Risk group, N = 75: | |||||
| Hypertension | 2.19 ± 0.83 | ||||
| Type 2 diabetes | 2.39 ± 0.97 | ||||
| Ischemic heart disease | 2.22 ± 0.73 | ||||
| Blood | 200 individuals: | MDA was significantly higher in stroke patients than in healthy controls. | [45] | ||
| Ischemic stroke, N = 100 | 7.11 ± 1.67 | ||||
| Healthy control, N = 100 | 1.64 ± 0.82 | ||||
| 4-HNE (µmol) | Plasma | 60 men: | The plasma 4-HNE concentrations in patients with ischemic stroke were higher than those in healthy control. | [74] | |
| Ischemic stroke: N = 24, | ≈11 | ||||
| Normal men, N = 36 | ≈9 | ||||
| TBARs (µmol/L) | Serum | 180 individuals: | The concentration of TBARS was significantly higher in stroke patients than in the controls. | [61] | |
| Acute ischemic stroke, N = 100: | |||||
| Small-vessel, N = 75 | 20.7 ± 2.6 | ||||
| Large-vessel, N = 25 | 19.7 ± 1.2 | ||||
| Healthy control (age- and sex-matched), N = 80 | ≈16 | ||||
| DNA oxidation | |||||
| 8-oHdG (ng/mgCr) | Urine | 44 acute ischemic stroke patients: Lacunar, N = 9 Atherothrombotic, N = 22 Cardioembolic, N = 13 | Day 0 15.8 ± 6.9 12.8 ± 11.7 11.8 ± 5.6 Day 7 16.1 ± 5.1 16.2 ± 15.5 13.0 ± 5.0 | 8-oHdG urinary levels increase in time, following AIS. In patients with better outcomes, the level increase is significantly slower. | [86] |
| 8-oHdG (ng/L) | Blood | 241 acute ischemic stroke patients: | There was a difference between 8-oHdG levels and ischemic stroke severity in depressed versus non-depressed patients. Mild positive Spearman correlation between 8-oHdG levels and catalase activity. Urinary 8-oHdG levels could be used as reliable and valuable biomarkers to predict functional outcomes in stroke rehabilitation. | [87] | |
| Depressive post-ischemic stroke, N = 70 | 218.0 (170.6–246.7) | ||||
| Non-depressive post-ischemic stroke, N = 171 | 164.8 (121.1–208.0) | ||||
| 8-oHdG (ng/mg creatinine) | Urine | Acute ischemic stroke patients, N = 61: | There was a difference between 8-oHdG levels and ischemic stroke severity in depressed versus non-depressed patients. Mild positive Spearman correlation between 8-oHdG levels and catalase activity. Urinary 8-oHdG levels could be used as reliable and valuable biomarkers to predict functional outcomes in stroke rehabilitation. | [88] | |
| Before rehabilitation | 5.87 ± 2.77 | ||||
| After rehabilitation | 5.60 ± 2.47 | ||||
| Protein oxidation | |||||
| Protein carbonyls (nmol/mg protein) | Plasma | 163 individuals: | Protein carbonyls were not significantly different in the experimental groups compared to the controls. | [48] | |
| Healthy control (gender and age matched), N = 81 | 0.25 ± 0.04 | ||||
| Acute ischemic stroke, N = 82 | 0.28 ± 0.04 | ||||
| Homocysteine (µmol/L) | Plasma | 653 individuals: | The increase in total homocysteine concentrations was associated with a 6% to 7% increase in stroke risk | [89] | |
| Stroke patients (male and female), N = 120 | ≥18.6 | ||||
| Control subjects, N = 533 | 12.0 | ||||
| Serum | 225 individuals | Moderate hyperhomocysteinema has been proposed as an independent risk factor for stroke in middle-aged British men | [90] | ||
| Stroke patients (male), N = 107 | 13.7 | ||||
| Healthy subjects (male), N = 118 | 11.9 | ||||
| Blood | 71 individuals: | Hyperhomocysteinemia is independent risk factors for stroke | [91] | ||
| Ischemic stroke patients (47 males, 24 females), N = 71 | 22.76 × 104 ± 12.67 | ||||
| Glutathion (nmol/g of brain tissue) | Brain tissue | 36 White male rats (weighing 260–300 g; 2–3 months of age) | GSH homeostasis as an oxidative stress marker has been disturbed in global and focal ischemia. After focal and global cerebral ischemia, a significant drop was recorded in the levels of the reduced forms of GSH in blood plasma. This effect may be attributed to their oxidation. | [92] | |
| Control: | |||||
| Reduced GSH | 592 ± 28 | ||||
| Oxidized GSH | 38 ± 4 | ||||
| Bilateral occlusion of the common carotid arteries (BCAO): | |||||
| Reduced GSH | 575 ± 24 | ||||
| Oxidized GSH | 279 ± 20 | ||||
| Middle cerebral artery occlusion (MCAO): | |||||
| Reduced GSH | 593 ± 35 | ||||
| Oxidized GSH | 204 ± 22 | ||||
| Blood | 140 Individuals (33 female, 37 male): | [93] | |||
| Control group (volunteers with similar cerebrovascular risk factors), N = 70 | 2.3 ± 0.4 | ||||
| Patients with acute ischemic stroke, N = 70 | 3.9 ± 2.5 | ||||
| S-adenosylhomocysteine methylation (%) | Whole peripheral blood | 202 individuals: | In patients with ischemic stroke, the percentage of methylated reference AHCY was significantly higher than in controls. | [94] | |
| Patients with acute ischemic stroke, N = 64 | 0.13% (0.09%, 0.27%) | ||||
| Control group, N = 138 | 0.06% (0.00%, 0.17%) | ||||
| Methionine (odds ratios (oRs) | Plasma | PREDIMED Cohort, 567 women: Stroke cases, N = 59 Controls, N = 508 | OR 1.85 (95% CI 1.44–2.37) | Methionine sulfoxide was linked to an increased risk of stroke | [95] |
| Oxidative Stress Biomarkers | Detection Strategy Examples | References |
|---|---|---|
| Malondialdehyde | UV-Vis Fluorescence Electrochemistry GC-MS SERS TBARS Test | [96,97] |
| F2-isoprostanes | GC-MS GC-NICI-MS ELISA HPLC-MS/MS SPE-HPLC-MS/MS | [98,99,100,101] |
| 4-Hydroxy-2-Nonenal | 2-AP HPLC Sandwich ELISA Western Blot LC-MSMS FT-ICR MS MALDI-TOF-MS 32P-PostlabelingGC-MS DNPH Derivatization | [102,103,104] |
| Oxidized LDL | TBARS assay ELISA | [105,106] |
| 8-oxo-7,8-dihydro-2-deoxyguanosine | HPLC-ED | [107] |
| 3-Nitrotyrosine | (HPLC)-(UV-VIS) absorption Electrochemical (ECD) Diode array (DAD) LC-MS LC-MS/MS GC-MS GC-MS/MS Sandwich ELISA | [108,109] |
| Gluthatione | DTNB/GR enzyme recycling method HPLC | [110] |
| Protein carbonyls | Western blot In-gel fluorophoric tagging Levine spectrophotometric method ELISA HPLC | [111,112] |
| Homocysteine | HPLC with fluorometric detection HPLC-ED Immunonephelometric method LC-MS-MS Fluorescence polarization Immunoassay EIA | [113] |
| Methionine sulfoxide | Peptide mapping with MS detection rpHPLC HIC Weak cation-exchange chromatography | [114] |
| Myeloperoxidase | BLI CRET ADHP MPO-Gd MR imaging | [115,116] |
3. DNA Oxidation
4. Oxidative Protein Modifications
4.1. Protein Carbonyls
4.2. 3-Nitrotyrosine
4.3. Ischemia-Modified Albumin (IMA)
4.4. Advanced Glycation End Products
4.5. Advanced Lipoxigenation End Products
4.6. Methionine Sulfoxide
4.7. Homocysteine
4.8. Glutathione
5. Changes in Protein Coding Gene
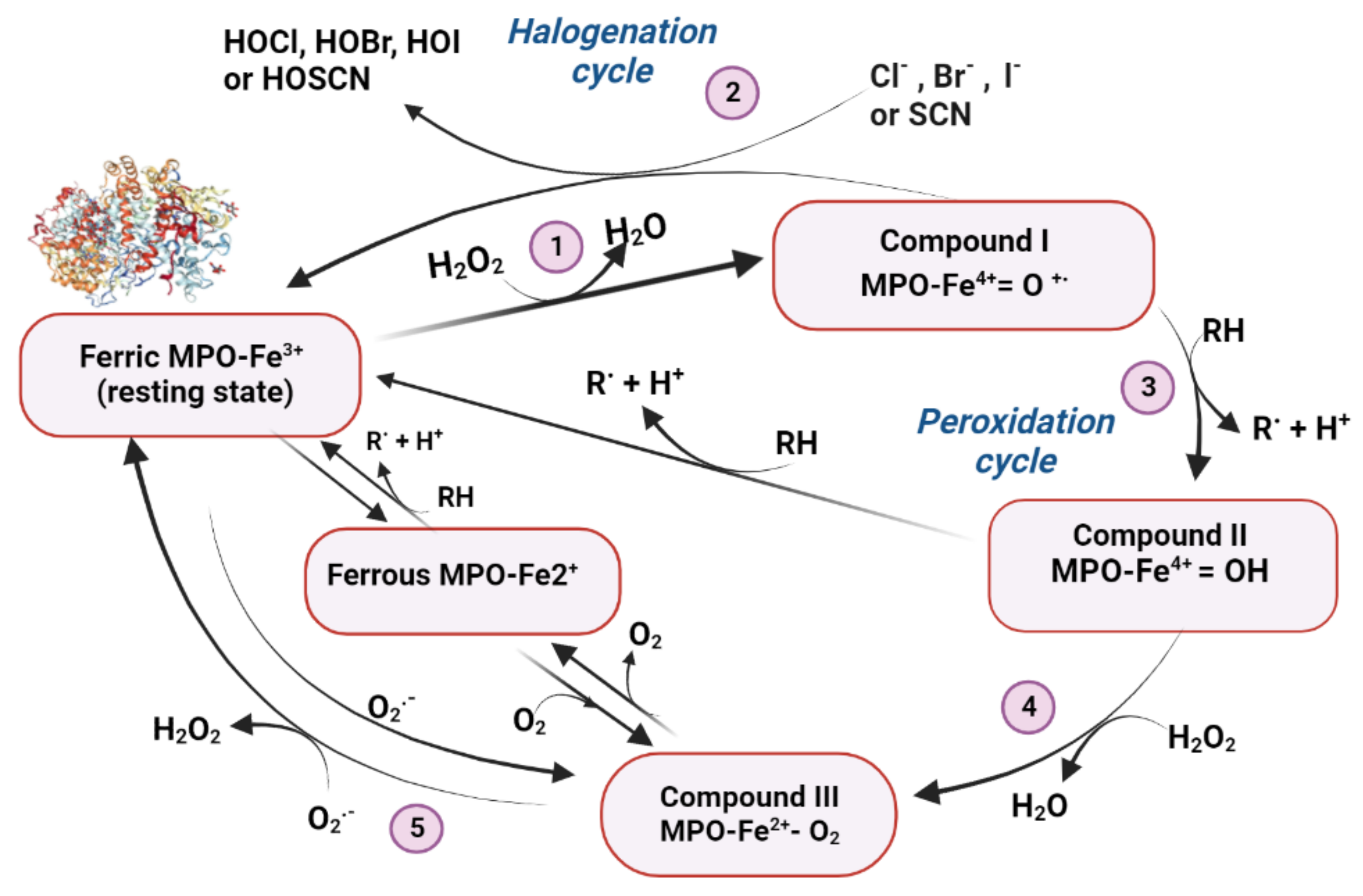
Myeloperoxidase
6. Measuring the Modification of Antioxidant Defense Status as Potential Indirect Markers of Oxidative Stress
6.1. Antioxidant Enzymes
6.2. Non-Enzymatic Antioxidants
6.3. Total Antioxidant Power
6.4. Total and Perchloric Acid Oxygen Radical Absorbance Capacity
6.5. Total Peroxyl Radical–Trapping Potential
7. Future Treatment Directions
7.1. Targeting Oxidative Stress May Be a Potential Treatment for Ischemic Stroke
7.2. Antioxidant Can Be Explored for Their Potential to Prevent Ischemic Stroke
8. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Hui, C.; Tadi, P.; Patti, L. Ischemic stroke. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2022. [Google Scholar]
- Feigin, V.L.; Brainin, M.; Norrving, B.; Martins, S.; Sacco, R.L.; Hacke, W.; Fisher, M.; Pandian, J.; Lindsay, P. World Stroke Organization (WSO): Global Stroke Fact Sheet 2022. Int. J. Stroke 2022, 17, 18–29. [Google Scholar] [CrossRef]
- Feigin, V.L.; Krishnamurthi, R.V.; Parmar, P.; Norrving, B.; Mensah, G.A.; Bennett, D.A.; Barker-Collo, S.; Moran, A.E.; Sacco, R.L.; Truelsen, T.; et al. Update on the global burden of ischemic and haemorrhagic stroke in 1990–2013: The GBD 2013 study. Neuroepidemiology 2015, 45, 161–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krafft, P.R.; Bailey, E.L.; Lekic, T.; Rolland, W.B.; Altay, O.; Tang, J.; Wardlaw, J.M.; Zhang, J.H.; Sudlow, C.L.M. Etiology of Stroke and Choice of Models. Int. J. Stroke 2012, 7, 398–406. [Google Scholar] [CrossRef]
- Cui, Q.; Naikoo, N.A. Modifiable and non-modifiable risk factors in ischemic stroke: A meta-analysis. Afr. Health Sci. 2019, 19, 2121–2129. [Google Scholar] [CrossRef]
- Boehme, A.K.; Esenwa, C.; Elkind, M.S. Stroke risk factors, genetics, and prevention. Circ. Res. 2017, 120, 472–495. [Google Scholar] [CrossRef] [PubMed]
- Donkor, E.S. Stroke in the century: A snapshot of the burden, epidemiology, and quality of life. Stroke Res. Treat. 2018, 2018, 3238165. [Google Scholar]
- Yu, H.; Chen, X.; Guo, X.; Chen, D.; Jiang, L.; Qi, Y.; Shao, J.; Tao, L.; Hang, J.; Lu, G.; et al. The clinical value of serum xanthine oxidase levels in patients with acute ischemic stroke. Redox Biol. 2023, 60, 102623. [Google Scholar] [CrossRef]
- Feigin, V.L.; Roth, A.G.; Naghavi, M.; Parmar, P.; Krishnamurthi, R.; Chugh, S.; Mensah, A.G.; Norrving, B.; Shiue, I.; Ng, M.; et al. Global burden of stroke and risk factors in 188 countries, during 1990–2013: A systematic analysis for the Global Burden of Disease Study 2013. Lancet Neurol. 2016, 15, 913–924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blanc-Labarre, C.; Delpont, B.; Hervieu-Bègue, M.; Osseby, G.-V.; Ricolfi, F.; Thouant, P.; Giroud, M.; Béjot, Y. Prise en charge aiguë des infarctus cérébraux en 2017. Rev. Méd. Interne 2018, 39, 408–413. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhang, S.; Fan, X. Role of Polyphenols as Antioxidant Supplementation in Ischemic Stroke. Oxid. Med. Cell. Longev. 2021, 2021, 5471347. [Google Scholar] [CrossRef]
- Musuka, T.D.; Wilton, S.B.; Traboulsi, M.; Hill, M.D. Diagnosis and management of acute ischemic stroke: Speed is critical. Can. Med. Assoc. J. 2015, 187, 887–893. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Sánchez, P.; Gutiérrez-Fernández, M.; Fuentes, B.; Masjuán, J.; Cases, M.A.D.L.; Novillo-López, M.E.; Díez-Tejedor, E. Biochemical and inflammatory biomarkers in ischemic stroke: Translational study between humans and two experimental rat models. J. Transl. Med. 2014, 12, 220. [Google Scholar] [CrossRef] [Green Version]
- Hill, M.D.; Jackowski, G.; Bayer, N.; Lawrence, M.; Jaeschke, R. Biochemical markers in acute ischemic stroke. Can. Med. Assoc. J. 2000, 162, 1139–1140. [Google Scholar]
- Teppo, K.; Jaakkola, J.; Biancari, F.; Halminen, O.; Putaala, J.; Mustonen, P.; Haukka, J.; Linna, M.; Kinnunen, J.; Tiili, P.; et al. Mental health conditions and risk of first-ever ischaemic stroke and death in patients with incident atrial fibrillation: A nationwide cohort study. Eur. J. Clin. Investig. 2022, 52, e13801. [Google Scholar] [CrossRef]
- Li, W.; Xiao, W.-M.; Chen, Y.-K.; Qu, J.-F.; Liu, Y.-L.; Fang, X.-W.; Weng, H.-Y.; Luo, G.-P. Anxiety in Patients With Acute Ischemic Stroke: Risk Factors and Effects on Functional Status. Front. Psychiatry 2019, 10, 257. [Google Scholar] [CrossRef]
- Hoyer, C.; Schmidt, H.L.; Kranaster, L.; Alonso, A. Impact of psychiatric comorbidity on the severity, short-term functional outcome, and psychiatric complications after acute stroke. Neuropsychiatr. Dis. Treat. 2019, 15, 1823–1831. [Google Scholar] [CrossRef]
- Chen, H.; Yoshioka, H.; Kim, G.S.; Jung, J.E.; Okami, N.; Sakata, H.; Maier, C.M.; Narasimhan, P.; Goeders, C.E.; Chan, P.H. Oxidative Stress in Ischemic Brain Damage: Mechanisms of Cell Death and Potential Molecular Targets for Neuroprotection. Antioxid. Redox Signal. 2011, 14, 1505–1517. [Google Scholar] [CrossRef] [Green Version]
- Rodrigo, R.; Fernandez-Gajardo, R.; Gutierrez, R.; Matamala, J.; Carrasco, R.; Miranda-Merchak, A.; Feuerhake, W. Oxidative Stress and Pathophysiology of Ischemic Stroke: Novel Therapeutic Opportunities. CNS Neurol. Disord.-Drug Targets 2013, 12, 698–714. [Google Scholar] [CrossRef] [PubMed]
- Lushchak, V.I.; Duszenko, M.; Gospodaryov, D.V.; Garaschuk, O. Oxidative Stress and Energy Metabolism in the Brain: Midlife as a Turning Point. Antioxidants 2021, 10, 1715. [Google Scholar] [CrossRef] [PubMed]
- Pawluk, H.; Kołodziejska, R.; Grześk, G.; Woźniak, A.; Kozakiewicz, M.; Kosinska, A.; Pawluk, M.; Grzechowiak, E.; Wojtasik, J.; Kozera, G. Increased Oxidative Stress Markers in Acute Ischemic Stroke Patients Treated with Thrombolytics. Int. J. Mol. Sci. 2022, 23, 15625. [Google Scholar] [CrossRef]
- Saver, J.L. Time Is Brain—Quantified. Stroke 2006, 37, 263–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shao, A.; Lingling, W.; Wang, L.; Tu, S.; Lenahan, C.; Zhang, J. Oxidative Stress at the Crossroads of Aging, Stroke and Depression. Aging Dis. 2020, 11, 1537–1566. [Google Scholar] [CrossRef]
- Chavda, V.; Chaurasia, B.; Garg, K.; Deora, H.; Umana, G.E.; Palmisciano, P.; Scalia, G.; Lu, B. Molecular mechanisms of oxidative stress in stroke and cancer. Brain Disord. 2021, 5, 100029. [Google Scholar] [CrossRef]
- He, J.; Liu, J.; Huang, Y.; Tang, X.; Xiao, H.; Hu, Z. Oxidative stress, inflamma-tion, and autophagy: Potential targets of mesenchymal stem cells-based therapies in is-chemic stroke. Front. Neurosci. 2021, 15, 641157. [Google Scholar] [CrossRef]
- Manzanero, S.; Santro, T.; Arumugam, T.V. Neuronal oxidative stress in acute ischemic stroke: Sources and contribution to cell injury. Neurochem. Int. 2013, 62, 712–718. [Google Scholar] [CrossRef]
- Cheng, Z.; Wang, L.; Qu, M.; Liang, H.; Li, W.; Li, Y.; Deng, L.; Zhang, Z.; Yang, G.-Y. Mesenchymal stem cells attenuate blood-brain barrier leakage after cerebral ischemia in mice. J. Neuroinflamm. 2018, 15, 135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Battelli, M.G.; Polito, L.; Bortolotti, M.; Bolognesi, A. Xanthine Oxidoreductase in Drug Metabo-lism: Beyond a Role as a Detoxifying Enzyme. Curr. Med. Chem. 2016, 23, 4027–4036. [Google Scholar] [CrossRef] [Green Version]
- Shibuya, S.; Watanabe, K.; Ozawa, Y.; Shimizu, T. Xanthine Oxidoreductase-Mediated Super-oxide Production Is Not Involved in the Age-Related Pathologies in Sod1-Deficient Mice. Int. J. Mol. Sci. 2021, 22, 3542. [Google Scholar] [CrossRef]
- Jelinek, M.; Jurajda, M.; Duris, K. Oxidative Stress in the Brain: Basic Concepts and Treatment Strategies in Stroke. Antioxidants 2021, 10, 1886. [Google Scholar] [CrossRef]
- Patt, A.; Harken, A.H.; Burton, L.K.; Rodell, T.C.; Piermattei, D.; Schorr, W.J.; Parker, N.B.; Berger, E.M.; Horesh, I.R.; Terada, L.S. Xanthine oxidase-derived hydrogen peroxide contributes to ischemia reperfusion-induced edema in gerbil brains. J. Clin. Investig. 1988, 81, 1556–1562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oka, H.; Kanemitsu, H.; Nihei, H.; Nakayama, H.; Tamura, A.; Sano, K. Change of xanthine de-hydrogenase and xanthine oxidase activities in rat brain following complete ischaemia. Neurol. Res. 1992, 14, 321–324. [Google Scholar] [CrossRef]
- Kinuta, Y.; Kimura, M.; Itokawa, Y.; Ishikawa, M.; Kikuchi, H.; Gamper, N.; Ooi, L.; Mortazavi, M.M.; Verma, K.; Harmon, O.A.; et al. Changes in xanthine oxidase in ischemic rat brain. J. Neurosurg. 1989, 71, 417–420. [Google Scholar] [CrossRef] [Green Version]
- Kahles, T.; Brandes, R.P. NADPH oxidases as therapeutic targets in ischemic stroke. Cell. Mol. Life Sci. 2012, 69, 2345–2363. [Google Scholar] [CrossRef]
- Vermot, A.; Petit-Härtlein, I.; Smith, S.M.; Fieschi, F. NADPH Oxidases (NOX): An Overview from Discovery, Molecular Mechanisms to Physiology and Pathology. Antioxidants 2021, 10, 890. [Google Scholar] [CrossRef]
- Taylor, C.J.; Weston, R.M.; Dusting, G.J.; Roulston, C.L. NADPH oxidase and angiogenesis fol-lowing endothelin-1 induced stroke in rats: Role for nox 2 in brain repair. Brain Sci. 2013, 3, 294–317. [Google Scholar] [CrossRef]
- Vallet, P.; Charnay, Y.; Steger, K.; Ogier-Denis, E.; Kovari, E.; Herrmann, F.; Michel, J.-P.; Szanto, I. Neuronal expression of the NADPH oxidase NOX4, and its regulation in mouse experimental brain ischemia. Neuroscience 2005, 132, 233–238. [Google Scholar] [CrossRef]
- Yang, S.; Li, W. Targeting oxidative stress for the treatment of ischemic stroke: Upstream and downstream therapeutic strategies. Brain Circ. 2016, 2, 153–163. [Google Scholar] [CrossRef]
- Liu, X.; Davis, C.M.; Alkayed, N.J. P450 Eicosanoids and Reactive Oxygen Species Interplay in Brain Injury and Neuroprotection. Antioxid. Redox Signal. 2018, 28, 987–1007. [Google Scholar] [CrossRef]
- Alexandrov, A.V.; Grotta, J.C. Arterial re-occlusion in stroke patients treated with intravenous tissue plasminogen activator. Neurology 2002, 59, 862–867. [Google Scholar] [CrossRef]
- Yamashita, A.; Asada, Y. Pathology and Mechanisms Underlying Thrombus Formation in Stroke. Brain Nerve 2021, 73, 965–974. [Google Scholar]
- Elsayed, W.M.; Abdel-Gawad, E.H.A.; Mesallam, D.I.; El-Serafy, T.S. The relationship between oxidative stress and acute ischemic stroke severity and func-tional outcome. Egypt. J. Neurol. Psychiatry Neurosurg. 2020, 56, 74. [Google Scholar] [CrossRef]
- Orellana-Urzúa, S.; Rojas, I.; Líbano, L.; Rodrigo, R. Pathophysiology of Ischemic Stroke: Role of Oxidative Stress. Curr. Pharm. Des. 2020, 26, 4246–4260. [Google Scholar] [CrossRef]
- Oraby, M.I.; Rabie, R.A. Blood biomarkers for stroke: The role of thioredoxin in diagnosis and prog-nosis of acute ischemic stroke. Egypt. J. Neurol. Psychiatry Neurosurg. 2020, 56, 1. [Google Scholar] [CrossRef]
- Menon, B.; Ramalingam, K.; Kumar, R. Evaluating the Role of Oxidative Stress in Acute Ischemic Stroke. J. Neurosci. Rural Pract. 2020, 11, 156–159. [Google Scholar] [CrossRef] [Green Version]
- Lorenzano, S.; Rost, N.S.; Khan, M.; Li, H.; Batista, L.M.; Chutinet, A.; Green, R.E.; Thankachan, T.K.; Thornell, B.; Muzikansky, A.; et al. Early molecular oxidative stress biomarkers of ischemic penumbra in acute stroke. Neurology 2019, 93, e1288–e1298. [Google Scholar] [CrossRef]
- Shaafi, S.; Hadisi, F.; Mahmoudinezhad, M.; Razmi, H.; Nejadghaderi, S.A.; Khalili, M. The significance of the oxidative stress markers in the one-year prognosis of patients with acute ischemic stroke: A case-control study. BMC Neurol. 2021, 21, 258. [Google Scholar] [CrossRef] [PubMed]
- Žitňanová, I.; Šiarnik, P.; Kollár, B.; Chomová, M.; Pazderová, P.; Andrezálová, L.; Ježovičová, M.; Koňariková, K.; Lauber-tová, L.; Krivošíková, Z.; et al. Oxidative Stress Markers and Their Dynamic Changes in Patients after Acute Ischemic Stroke. Oxid. Med. Cell. Longev. 2016, 2016, 9761697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Gao, S.; Lenahan, C.; Gu, Y.; Wang, X.; Fang, Y.; Xu, W.; Wu, H.; Pan, Y.; Shao, A.; et al. Melatonin as an Antioxidant Agent in Stroke: An Updated Review. Aging Dis. 2022, 13, 1823. [Google Scholar] [CrossRef]
- Wang, A.; Liu, J.; Meng, X.; Wang, H.; Wang, Y.; Su, Z.; Zhang, N.; Dai, L. Association between oxidized low-density lipoprotein and cognitive impairment in patients with ischemic stroke. Eur. J. Neurol. 2017, 25, 185–191. [Google Scholar] [CrossRef]
- Wang, A.; Yang, Y.; Su, Z.; Yue, W.; Hao, H.; Ren, L.; Wang, Y.; Cao, Y.; Wang, Y. Association of Oxidized Low-Density Lipoprotein With Prognosis of Stroke and Stroke Subtypes. Stroke 2017, 48, 91–97. [Google Scholar] [CrossRef]
- Wang, A.; Cui, Y.; Meng, X.; Su, Z.; Cao, Y.; Yang, Y.; Liu, C.; Dai, L.; Wang, Y.; Wang, Y. The relationship between oxidized low-density lipoprotein and the NIHSS score among patients with acute ischemic stroke: The SOS-Stroke Study. Atherosclerosis 2018, 270, 21–25. [Google Scholar] [CrossRef]
- Ayala, A.; Muñoz, M.F.; Argüelles, S. Lipid peroxidation: Production, metabolism, and signaling mechanisms of malondialdehyde and 4-hydroxy-2-nonenal. Oxid. Med. Cell. Longev. 2014, 2014, 360438. [Google Scholar] [CrossRef] [Green Version]
- Feng, H.; Stockwell, B.R. Unsolved mysteries: How does lipid peroxidation cause ferroptosis? PLoS Biol. 2018, 16, e2006203. [Google Scholar] [CrossRef]
- Čepelak, I.; Dodig, S.; Dodig, D. Ferroptosis: Regulated cell death. Arch. Ind. Hyg. Toxicol. 2020, 71, 99–109. [Google Scholar] [CrossRef]
- Bu, Z.Q.; Yu, H.Y.; Wang, J.; He, X.; Cui, Y.R.; Feng, J.C.; Feng, J. Emerging Role of Ferroptosis in the Pathogenesis of Ischemic Stroke: A New Therapeutic Target? ASN Neuro 2021, 13, 17590914211037505. [Google Scholar] [CrossRef]
- Fang, X.-L.; Ding, S.-Y.; Du, X.-Z.; Wang, J.-H.; Li, X.-L. Ferroptosis—A Novel Mechanism With Multifaceted Actions on Stroke. Front. Neurol. 2022, 13, 881809. [Google Scholar] [CrossRef]
- Worrall, S.; Thiele, G.M. Modification of proteins by reactive ethanol metabolites: Adduct structure, functional and pathological consequences. In Comprehensive Handbook of Alcohol Related Pathology; Preedy, V.R., Watson, R.R., Eds.; Academic Press: Cambridge, MA, USA, 2005; pp. 1209–1222. ISBN 9780125643702. [Google Scholar]
- Zahra, K.F.; Lefter, R.; Ali, A.; Abdellah, E.-C.; Trus, C.; Ciobica, A.; Timofte, D. The Involvement of the Oxidative Stress Status in Cancer Pathology: A Double View on the Role of the Antioxidants. Oxid. Med. Cell. Longev. 2021, 2021, 9965916. [Google Scholar] [CrossRef]
- Cui, X.; Gong, J.; Han, H.; He, L.; Teng, Y.; Tetley, T.; Sinharay, R.; Chung, K.F.; Islam, T.; Gilliland, F.; et al. Relationship between free and total malondialdehyde, a well-established marker of oxidative stress, in various types of human biospecimens. J. Thorac. Dis. 2018, 10, 3088–3197. [Google Scholar] [CrossRef]
- Tsai, N.-W.; Chang, Y.-T.; Huang, C.-R.; Lin, Y.-J.; Lin, W.-C.; Cheng, B.-C.; Su, C.-M.; Chiang, Y.-F.; Chen, S.-F.; Huang, C.-C.; et al. Association between Oxidative Stress and Outcome in Different Subtypes of Acute Ischemic Stroke. BioMed Res. Int. 2014, 2014, 256879. [Google Scholar] [CrossRef] [Green Version]
- Niedernhofer, L.J.; Scott Daniels, J.; Rouzer, C.A.; Greene, R.E.; Marnett, L.J. Malondialdehyde, a Product of Lipid Peroxidation, is Mutagenic in Human Cells. J. Biol. Chem. 2003, 278, 31426–31433. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Liu, Y.; Tu, X.; Shen, H.; Qiu, H.; Chen, H.; He, J. High Serum Levels of Malondialdehyde and 8-OHdG are both Associated with Early Cognitive Impairment in Patients with Acute Ischaemic Stroke. Sci. Rep. 2017, 7, 9493. [Google Scholar] [CrossRef] [PubMed]
- Szczuko, M.; Kozioł, I.; Kotlęga, D.; Brodowski, J.; Drozd, A. The Role of Thromboxane in the Course and Treatment of Ischemic Stroke: Review. Int. J. Mol. Sci. 2021, 22, 11644. [Google Scholar] [CrossRef]
- Al-Rawi, N.H.; Jaber, F.; Atiyah, K.M. Assessment of salivary and serum oxidative stress and antioxidants as plausible parameters in prediction of ischemic stroke among Iraqi samples. Internet J. Third World Med. 2009, 7, 1–9. [Google Scholar]
- Milne, G.L.; Musiek, E.S.; Morrow, J.D. F2-isoprostanes as markers of oxidative stress in vivo: An overview. Biomarkers 2005, 10, S10–S23. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Ojeda, F.J.; Olza, J.; Gil, Á.; Aguilera, C.M. Oxidative stress and inflammation in obesity and metabolic syndrome. In Obesity: Oxidative Stress and Dietary Antioxidants; Academic Press: Cambridge, MA, USA, 2018; pp. 1–15. [Google Scholar] [CrossRef]
- Putman, A.; Contreras, G.; Sordillo, L. Isoprostanes in Veterinary Medicine: Beyond a Biomarker. Antioxidants 2021, 10, 145. [Google Scholar] [CrossRef]
- Montuschi, P.; Barnes, P.J.; Roberts, L.J., 2nd. Isoprostanes: Markers and mediators of oxidative stress. FASEB J. 2004, 18, 1791–1800. [Google Scholar] [CrossRef]
- Lorenzano, S.; Rost, N.S.; Khan, M.; Li, H.; Lima, F.O.; Maas, M.B.; Green, R.E.; Thankachan, T.K.; Dipietro, A.J.; Arai, K.; et al. Oxidative stress biomarkers of brain damage: Hyperacute plasma f2-isoprostane predicts infarct growth in stroke. Stroke 2018, 49, 630–637. [Google Scholar] [CrossRef]
- Seet, R.C.; Lee, J.C.-Y.; Chan, B.P.; Sharma, V.K.; Teoh, H.-L.; Venketasubramanian, N.; Lim, E.C.; Chong, W.-L.; Looi, W.-F.; Huang, S.-H.; et al. Oxidative Damage in Ischemic Stroke Revealed Using Multiple Biomarkers. Stroke 2011, 42, 2326–2329. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Bi, R.; Sun, S.; Chen, S.; Chen, J.; Hu, B.; Jin, H. The Role of Oxidative Stress in Acute Ischemic Stroke-Related Thrombosis. Oxid. Med. Cell. Longev. 2022, 2022, 8418820. [Google Scholar] [CrossRef]
- Reilly, M.P.; Delanty, N.; Roy, L.; Rokach, J.; Callaghan, P.O.; Crean, P.; Lawson, J.A.; FitzGerald, G.A. Increased formation of the isoprostanes IPF2alpha-I and 8-epi-prostaglandin F2alpha in acute coronary angioplasty: Evidence for oxidant stress during coronary reperfusion in humans. Circulation 1997, 96, 3314–3320. [Google Scholar] [CrossRef]
- Lee, W.-C.; Wong, H.-Y.; Chai, Y.-Y.; Shi, C.-W.; Amino, N.; Kikuchi, S.; Huang, S.-H. Lipid peroxidation dysregulation in ischemic stroke: Plasma 4-HNE as a potential biomarker? Biochem. Biophys. Res. Commun. 2012, 425, 842–847. [Google Scholar] [CrossRef]
- Guo, J.-M.; Liu, A.-J.; Zang, P.; Dong, W.-Z.; Ying, L.; Wang, W.; Xu, P.; Song, X.-R.; Cai, J.; Zhang, S.-Q.; et al. ALDH2 protects against stroke by clearing 4-HNE. Cell Res. 2013, 23, 915–930. [Google Scholar] [CrossRef]
- Valente, A.J.; Irimpen, A.M.; Siebenlist, U.; Chandrasekar, B. OxLDL induces endothelial dysfunction and death via TRAF3IP2: Inhibition by HDL3 and AMPK activators. Free Radic. Biol. Med. 2014, 70, 117–128. [Google Scholar] [CrossRef] [Green Version]
- Matsuura, E.; Hughes, G.R.; Khamashta, A.M. Oxidation of LDL and its clinical implication. Autoimmun. Rev. 2008, 7, 558–566. [Google Scholar] [CrossRef]
- Parthasarathy, S.; Raghavamenon, A.; Garelnabi, M.O.; Santanam, N. Oxidized Low-Density Lipoprotein. In Free Radicals and Antioxidant Protocols: Methods in Molecular Biology; Humana Press: Totowa, NJ, USA, 2010; Volume 610, pp. 403–417. [Google Scholar] [CrossRef] [Green Version]
- Varghese, D.S.; Ali, B.R. Pathological Crosstalk Between Oxidized LDL and ER Stress in Human Diseases: A Comprehensive Review. Front. Cell Dev. Biol. 2021, 9, 674103. [Google Scholar] [CrossRef] [PubMed]
- Yan, Z.; Fu, B.; He, D.; Zhang, Y.; Liu, J.; Zhang, X. The relationship between oxidized low-density lipoprotein and related ratio and acute cerebral infarction. Medicine 2018, 97, e12642. [Google Scholar] [CrossRef]
- Gąsecka, A.; Rogula, S.; Szarpak, Ł.; Filipiak, K. LDL-Cholesterol and Platelets: Insights into Their Interactions in Atherosclerosis. Life 2021, 11, 39. [Google Scholar] [CrossRef] [PubMed]
- Berger, M.; Naseem, K.M. Oxidised Low-Density Lipoprotein-Induced Platelet Hyperactivity—Receptors and Signalling Mechanisms. Int. J. Mol. Sci. 2022, 23, 9199. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Zhou, Y.; Nabavi, S.M.; Sahebkar, A.; Little, P.J.; Xu, S.; Weng, J.; Ge, J. Mechanisms of Oxidized LDL-Mediated Endothelial Dysfunction and Its Consequences for the Development of Atherosclerosis. Front. Cardiovasc. Med. 2022, 9, 1442. [Google Scholar] [CrossRef]
- Wang, X.; Fu, Y.-F.; Liu, X.; Feng, G.; Xiong, D.; Mu, G.-F.; Chen, F.-P. ROS Promote Ox-LDL-Induced Platelet Activation by Up-Regulating Autophagy Through the Inhibition of the PI3K/AKT/mTOR Pathway. Cell. Physiol. Biochem. 2018, 50, 1779–1793. [Google Scholar] [CrossRef]
- Yang, X.; Sun, W.; Hou, D.; Wang, T.; Li, C.; Luo, Y.; Zhang, S.; Shen, L.; Liu, W.; Wu, D. The Degree of Plasma Oxidized Low-Density Lipoprotein Level Decrease Is Related to Clinical Outcomes for Patients with Acute Ischemic Stroke. Dis. Markers 2021, 2021, 4998823. [Google Scholar] [CrossRef]
- Saki, M.; Prakash, A. DNA damage related crosstalk between the nucleus and mitochondria. Free Radic. Biol. Med. 2017, 107, 216–227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakajima, H.; Unoda, K.; Ito, T.; Kitaoka, H.; Kimura, F.; Hanafusa, T. The relation of urinary 8-OHdG, a marker of oxidative stress to dna, and clinical outcomes for ischemic stroke. Open Neurol. J. 2012, 6, 51–57. [Google Scholar] [CrossRef]
- Liu, Z.; Cai, Y.; He, J. High serum levels of 8-OHdG are an independent predictor of post-stroke depression in Chinese stroke survivors. Neuropsychiatr. Dis. Treat. 2018, 14, 587–596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bots, M.L.; Launer, L.J.; Lindemans, J.; Hoes, A.W.; Hofman, A.; Witteman, J.C.; Koudstaal, P.J.; Grobbee, D.E. Homocysteine and short-term risk of myocardial infarction and stroke in the elderly: The Rotterdam Study. Arch. Intern. Med. 1999, 159, 38–44. [Google Scholar] [CrossRef] [Green Version]
- Perry, I.J.; Refsum, H.; Morris, R.W.; Ebrahim, S.B.; Ueland, P.M.; Shaper, A.G. Prospective study of serum total homocysteine concentration and risk of stroke in middle-aged British men. Lancet 1995, 346, 1395–1398. [Google Scholar] [CrossRef]
- Niazi, F.; Aslam, A.; Khattak, S.; Waheed, S. Frequency of Homocysteinemia in Young Ischemic Stroke Patients and Its Relationship with the Early Outcome of a Stroke. Cureus 2019, 11, e5625. [Google Scholar] [CrossRef] [Green Version]
- Ivanov, A.V.; Alexandrin, V.V.E.; Paltsyn, A.A.; Nikiforova, K.A.; Virus, E.D.; Luzyanin, B.P.; Kubatiev, A.A. Plasma low-molecular-weight thiol/disulphide homeostasis as an early indicator of glob-al and focal cerebral ischaemia. Redox Rep. 2017, 22, 460–466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ozkul, A.; Akyol, A.; Yenisey, C.; Arpaci, E.; Kiylioglu, N.; Tataroglu, C. Oxidative stress in acute ischemic stroke. J. Clin. Neurosci. 2007, 14, 1062–1066. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Chen, X.; Zhou, S.; Lin, Z.; Yu, X.; Huang, Y. DNA methylation of AHCY may increase the risk of ischemic stroke. Bosn. J. Basic Med. Sci. 2020, 20, 471–476. [Google Scholar] [CrossRef]
- Balasubramanian, R.; Hu, J.; Guasch-Ferre, M.; Li, J.; Sorond, F.; Zhao, Y.; Shutta, K.H.; Salas-Salvado, J.; Hu, F.; Clish, C.B.; et al. Metabolomic Profiles Associated With Incident Ischemic Stroke. Neurology 2021, 98, e483–e492. [Google Scholar] [CrossRef]
- Lu, Y.; Wan, S.; Ruan, X.; Liang, H.; Su, J.; Wang, Z.; Zhu, L. Phosphoric Acid Induced Controllable Nanoparticle Aggregation for Ultrasensitive SERS Detection of Malondialdehyde in a Microfluidic Chip. Chemosensors 2022, 10, 524. [Google Scholar] [CrossRef]
- Papastergiadis, A.; Mubiru, E.; Van Langenhove, H.; De Meulenaer, B. Malondialdehyde Measurement in Oxidized Foods: Evaluation of the Spectrophotometric Thiobarbituric Acid Reactive Substances (TBARS) Test in Various Foods. J. Agric. Food Chem. 2012, 60, 9589–9594. [Google Scholar] [CrossRef]
- Liu, W.; Morrow, J.D.; Yin, H. Quantification of F2-isoprostanes as a reliable index of oxidative stress in vivo using gas chromatography–mass spectrometry (GC-MS) method. Free Radic. Biol. Med. 2009, 47, 1101–1107. [Google Scholar] [CrossRef] [Green Version]
- Woolcock, A.D.; Leisering, A.; Deshuillers, P.; Roque-Torres, J.; Moore, G.E. Feline urinary F2-isoprostanes measured by enzyme-linked immunoassay and gas chromatography–mass spectroscopy are poorly correlated. J. Vet. Diagn. Investig. 2020, 32, 648–655. [Google Scholar] [CrossRef] [PubMed]
- Larose, J.; Julien, P.; Bilodeau, J.-F. Analysis of F2-isoprostanes in plasma of pregnant women by HPLC-MS/MS using a column packed with core-shell particles. J. Lipid Res. 2013, 54, 1505–1511. [Google Scholar] [CrossRef] [Green Version]
- Holder, C.; Adams, A.; McGahee, E.; Xia, B.; Blount, B.C.; Wang, L. High-Throughput and Sensitive Analysis of Free and Total 8-Isoprostane in Urine with Isotope-Dilution Liquid Chromatography-Tandem Mass Spectrometry. ACS Omega 2020, 5, 10919–10926. [Google Scholar] [CrossRef] [PubMed]
- Wakita, C.; Honda, K.; Shibata, T.; Akagawa, M.; Uchida, K. A method for detection of 4-hydroxy-2-nonenal adducts in proteins. Free Radic. Biol. Med. 2011, 51, 1–4. [Google Scholar] [CrossRef]
- Spickett, C.M. The lipid peroxidation product 4-hydroxy-2-nonenal: Advances in chemistry and analysis. Redox Biol. 2013, 1, 145–152. [Google Scholar] [CrossRef] [Green Version]
- Wacker, M.; Schuler, D.; Wanek, P.; Eder, E. Development of a 32P-postlabeling method for the detection of 1, N 2-propanodeoxyguanosine adducts of trans-4-hydroxy-2-nonenal in vivo. Chem. Res. Toxicol. 2000, 13, 1165–1173. [Google Scholar] [CrossRef]
- Tan, X.W.; Takenaka, F.; Takekawa, H.; Matsuura, E. Rapid and specific detection of oxidized LDL/β2GPI complexes via facile lateral flow immunoassay. Heliyon 2020, 6, e04114. [Google Scholar] [CrossRef] [PubMed]
- Itabe, H.; Ueda, M. Measurement of plasma oxidized low-density lipoprotein and its clinical implications. J. Atheroscler. Thromb. 2007, 14, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Lagadu, S.; Lechevrel, M.; Sichel, F.; Breton, J.; Pottier, D.; Couderc, R.; Moussa, F.; Prevost, V. 8-Oxo-7,8-dihydro-2’-deoxyguanosine as a biomarker of oxidative damage in oesophageal cancer patients: Lack of association with antioxidant vitamins and polymorphism of hOGG1 and GST. J. Exp. Clin. Cancer Res. 2010, 29, 157. [Google Scholar] [CrossRef] [Green Version]
- Tsikas, D.; Schwedhelm, E.; Frölich, J.C. Methodological Considerations on the Detection of 3-Nitrotyrosine in the Cardiovascular System. Circ. Res. 2002, 90, E70. [Google Scholar] [CrossRef] [Green Version]
- Teixeira, D.; Fernandes, R.; Prudêncio, C.; Vieira, M. 3-Nitrotyrosine quantification methods: Current concepts and future challenges. Biochimie 2016, 125, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tipple, T.E.; Rogers, L.K. Methods for the Determination of Plasma or Tissue Glutathione Levels. Dev. Toxicol. Methods Protoc. 2012, 889, 315–324. [Google Scholar] [CrossRef] [Green Version]
- Alomari, E.; Bruno, S.; Ronda, L.; Paredi, G.; Bettati, S.; Mozzarelli, A. Protein carbonylation detection methods: A comparison. Data Brief 2018, 19, 2215–2220. [Google Scholar] [CrossRef]
- Weber, D.; Davies, M.J.; Grune, T. Determination of protein carbonyls in plasma, cell extracts, tissue homogenates, isolated proteins: Focus on sample preparation and derivatization conditions. Redox Biol. 2015, 5, 367–380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alam, S.F.; Kumar, S.; Ganguly, P. Measurement of homocysteine: A historical perspective. J. Clin. Biochem. Nutr. 2019, 65, 171–177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jenkins, N. Posttranslation Modifications Other Than Glycosylation. J. Cell Physiol. 2011, 236, 8020–8034. [Google Scholar]
- Huang, J.; Milton, A.; Arnold, R.D.; Huang, H.; Smith, F.; Panizzi, J.R.; Panizzi, P. Methods for measuring myeloperoxidase activity toward assessing inhibitor efficacy in living systems. J. Leukoc. Biol. 2016, 99, 541–548. [Google Scholar] [CrossRef] [Green Version]
- Pulli, B.; Ali, M.; Forghani, R.; Schob, S.; Hsieh, K.L.C.; Wojtkiewicz, G.; Linnoila, J.J.; Chen, J.W. Measuring Myeloperoxidase Activity in Biological Samples. PLoS ONE 2013, 8, e67976. [Google Scholar] [CrossRef] [Green Version]
- Poetsch, A.R. The genomics of oxidative DNA damage, repair, and resulting mutagenesis. Comput. Struct. Biotechnol. J. 2020, 18, 207–219. [Google Scholar] [CrossRef] [PubMed]
- Massaad, C.A.; Klann, E. Reactive Oxygen Species in the Regulation of Synaptic Plasticity and Memory. Antioxid. Redox Signal. 2011, 14, 2013–2054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, A.-L.; Li, X.; Gu, Y.; Wright, P.M.; Chang, D.-Y. Repair of Oxidative DNA Damage: Mechanisms and Functions. Cell Biochem. Biophys. 2001, 35, 141–170. [Google Scholar] [CrossRef]
- Poulsen, H.E.; Nadal, L.L.; Broedbaek, K.; Nielsen, P.E.; Weimann, A. Detection and interpretation of 8-oxodG and 8-oxoGua in urine, plasma and cerebrospinal fluid. Biochim. Biophys. Acta (BBA)—Gen. Subj. 2014, 1840, 801–808. [Google Scholar] [CrossRef]
- Watanabe, S.; Kawasaki, Y.; Kawai, K. Salivary 8-hydroxyguanine as a lifestyle-related oxidative stress biomarker in workers. J. Clin. Biochem. Nutr. 2020, 66, 57–61. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.; Hao, W.; Pan, L.; Boldogh, I.; Ba, X. The roles of base excision repair enzyme OGG1 in gene expression. Cell. Mol. Life Sci. 2018, 75, 3741–3750. [Google Scholar] [CrossRef] [Green Version]
- Ide, T.; Tsutsui, H.; Hayashidani, S.; Kang, D.; Suematsu, N.; Nakamura, K.-I.; Utsumi, H.; Hamasaki, N.; Takeshita, A. Mitochondrial DNA Damage and Dysfunction Associated With Oxidative Stress in Failing Hearts After Myocardial Infarction. Circ. Res. 2001, 88, 529–535. [Google Scholar] [CrossRef] [Green Version]
- Hsieh, Y.-W.; Lin, K.-C.; Korivi, M.; Lee, T.-H.; Wu, C.-Y.; Wu, K.-Y. The Reliability and Predictive Ability of a Biomarker of Oxidative DNA Damage on Functional Outcomes after Stroke Rehabilitation. Int. J. Mol. Sci. 2014, 15, 6504–6516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, W.; Xiao, S.; Ahn, D.U. Protein Oxidation: Basic Principles and Implications for Meat Quality. Crit. Rev. Food Sci. Nutr. 2013, 53, 1191–1201. [Google Scholar] [CrossRef]
- Ahmad, S.; Khan, H.; Shahab, U.; Rehman, S.; Rafi, Z.; Khan, M.Y.; Ansari, A.; Siddiqui, Z.; Ashraf, J.M.; Abdullah, S.M.S.; et al. Protein oxidation an overview of metabolism of sulphur containing amino acid cysteine. Front. Biosci. 2017, 9, 71–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonos, E.S.; Kapetanou, M.; Sereikaite, J.; Bartosz, G.; Naparło, K.; Grzesik, M.; Sadowska-Bartosz, I. Origin and pathophysiology of protein carbonylation, nitration and chlorination in age-related brain diseases and aging. Aging 2018, 10, 868–901. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.R.; Kim, J.-K.; Lee, H.-S.; Kim, S.G.; Choi, E.-K. Serum levels of protein carbonyl, a marker of oxidative stress, are associated with overhydration, sarcopenia and mortality in hemodialysis patients. BMC Nephrol. 2020, 21, 281. [Google Scholar] [CrossRef]
- Ahsan, H. 3-Nitrotyrosine: A biomarker of nitrogen free radical species modified proteins in systemic autoimmunogenic conditions. Hum. Immunol. 2013, 74, 1392–1399. [Google Scholar] [CrossRef] [PubMed]
- Bandookwala, M.; Thakkar, D.; Sengupta, P. Advancements in the Analytical Quantification of Nitroxidative Stress Biomarker 3-Nitrotyrosine in Biological Matrices. Crit. Rev. Anal. Chem. 2019, 50, 265–289. [Google Scholar] [CrossRef]
- Medeiros, R.; Sousa, B.; Rossi, S.; Afonso, C.; Bonino, L.; Pitt, A.; López, E.; Spickett, C.; Borthagaray, G. Identification and relative quantification of 3-nitrotyrosine residues in fibrinogen nitrated in vitro and fibrinogen from ischemic stroke patient plasma using LC-MS/MS. Free Radic. Biol. Med. 2021, 165, 334–347. [Google Scholar] [CrossRef]
- Cichoń, N.; Bijak, M.; Miller, E.; Niwald, M.; Saluk, J. Poststroke Depression as a Factor Adversely Affecting the Level of Oxidative Damage to Plasma Proteins during a Brain Stroke. Oxid. Med. Cell. Longev. 2015, 2015, 408745. [Google Scholar] [CrossRef] [Green Version]
- Abboud, H.; Labreuche, J.; Meseguer, E.; Lavallee, P.C.; Simon, O.; Olivot, J.-M.; Mazighi, M.; Dehoux, M.; Benessiano, J.; Steg, P.G.; et al. Ischemia-Modified Albumin in Acute Stroke. Cerebrovasc. Dis. 2006, 23, 216–220. [Google Scholar] [CrossRef]
- Shevtsova, A.; Gordiienko, I.; Tkachenko, V.; Ushakova, G. Ischemia-Modified Albumin: Origins and Clinical Implications. Dis. Markers 2021, 2021, 9945424. [Google Scholar] [CrossRef]
- Menon, B.; Ramalingam, K.; Krishna, V. Study of Ischemia Modified Albumin as a Biomarker in Acute Ischaemic Stroke. Ann. Neurosci. 2018, 25, 187–190. [Google Scholar] [CrossRef]
- Jena, I.; Mohapatra, P.C.; Mohanty, N.R. Ischemia modified albumin: A biochemical marker of acute stroke. Int. J. Pharm. Biol. Sci. 2016, 7, 15–19. [Google Scholar]
- Okda, M.A.; El-Kabany, R.A.; Soliman, H.S.; El-Kholy, M.S. Ischemia-modified albumin for evaluating severity and predicting prognosis in patients with acute cerebrovascular disease. Menoufia Med. J. 2020, 33, 1072–1076. [Google Scholar]
- Goldin, A.; Beckman, J.A.; Schmidt, A.M.; Creager, M.A. Advanced glycation end products: Sparking the development of diabetic vascular injury. Circulation 2006, 114, 597–605. [Google Scholar] [CrossRef] [Green Version]
- Rabbani, N.; Thornalley, P.J. Advanced glycation end products in the pathogenesis of chronic kidney disease. Kidney Int. 2018, 93, 803–813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hegab, Z.; Gibbons, S.; Neyses, L.; Mamas, M.A. Role of advanced glycation end products in cardiovascular disease. World J. Cardiol. 2012, 4, 90–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ikeda, T.; Maruyama, K.; Ito, N.; Utagawa, A.; Nagane, M.; Shiokawa, Y. Serum Pentosidine, An Advanced Glycation End Product, Indicates Poor Outcomes After Acute Ischemic Stroke. J. Stroke Cerebrovasc. Dis. 2010, 21, 386–390. [Google Scholar] [CrossRef]
- Yokota, C. Serum level of AGE/RAGE as a potential biomarker for acute stroke. Cereb. Blood Flow Metab. (Jpn. J. Cereb. Blood Flow Metab. 2015, 26, 51–56. [Google Scholar] [CrossRef] [Green Version]
- Park, H.-Y.; Yun, K.H.; Park, D.-S. Levels of Soluble Receptor for Advanced Glycation End Products in Acute Ischemic Stroke without a Source of Cardioembolism. J. Clin. Neurol. 2009, 5, 126–132. [Google Scholar] [CrossRef] [Green Version]
- Montaner, J.; Mendioroz, M.; Ribo, M.; Delgado, P.; Quintana, M.; Penalba, A.; Chacon, P.; Molina, C.; Fernández-Cadenas, I.; Rosell, A.; et al. A panel of biomarkers including caspase-3 and D-dimer may differentiate acute stroke from stroke-mimicking conditions in the emergency department. J. Intern. Med. 2010, 270, 166–174. [Google Scholar] [CrossRef] [Green Version]
- Vistoli, G.; De Maddis, D.; Cipak, A.; Zarkovic, N.; Carini, M.; Aldini, G. Advanced glycoxidation and lipoxidation end products (AGEs and ALEs): An overview of their mechanisms of formation. Free Radic. Res. 2013, 47 (Suppl. S1), 3–27. [Google Scholar] [CrossRef] [Green Version]
- Moldogazieva, N.T.; Mokhosoev, I.M.; Mel’Nikova, T.I.; Porozov, Y.B.; Terentiev, A.A. Oxidative Stress and Advanced Lipoxidation and Glycation End Products (ALEs and AGEs) in Aging and Age-Related Diseases. Oxid. Med. Cell. Longev. 2019, 2019, 3085756. [Google Scholar] [CrossRef] [Green Version]
- Negre-Salvayre, A.; Coatrieux, C.; Ingueneau, C.; Salvayre, R. Advanced lipid peroxidation end products in oxidative damage to proteins. Potential role in diseases and therapeutic prospects for the inhibitors. Br. J. Pharmacol. 2008, 153, 6–20. [Google Scholar] [CrossRef] [Green Version]
- Pamplona, R. Advanced lipoxidation end-products. Chem. Interact. 2011, 192, 14–20. [Google Scholar] [CrossRef]
- Mashima, R.; Nakanishi-Ueda, T.; Yamamoto, Y. Simultaneous determination of methionine sulfoxide and methionine in blood plasma using gas chromatography-mass spectrometry. Anal. Biochem. 2003, 313, 28–33. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Qu, M.; Wang, N.; Wang, L.; Fan, G.; Yang, C. Betaine protects rats against ischemia/reperfusion injury-induced brain damage. J. Neurophysiol. 2022, 127, 444–451. [Google Scholar] [CrossRef] [PubMed]
- Gu, S.X.; Blokhin, I.O.; Wilson, K.M.; Dhanesha, N.; Doddapattar, P.; Grumbach, I.M.; Chauhan, A.K.; Lentz, S.R. Protein methionine oxidation augments reperfusion injury in acute ischemic stroke. J. Clin. Investig. 2016, 1, e86460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, H.; Blake, S.; Chan, K.T.; Pearson, R.B.; Kang, J. Cystathionine β-synthase in physiology and cancer. BioMed Res. Int. 2018, 1, 3205125. [Google Scholar]
- Finkelstein, J.D.; Martin, J.J. Homocysteine. Int. J. Biochem. Cell Biol. 2000, 32, 385–389. [Google Scholar] [CrossRef] [PubMed]
- Sacco, R.L.; Roberts, J.K.; Jacobs, B.S. Homocysteine as a Risk Factor for Ischemic Stroke: An Epidemiological Story in Evolution. Neuroepidemiology 1998, 17, 167–173. [Google Scholar] [CrossRef]
- Eikelboom, J.W.; Lonn, E.; Genest, J., Jr.; Hankey, G.; Yusuf, S. Homocyst (e) ine and cardiovascular disease: A critical review of the epidemiologic evidence. Ann. Intern. Med. 1999, 131, 363–375. [Google Scholar] [CrossRef] [PubMed]
- El Kossi, M.M.H.; Zakhary, M.M. Oxidative stress in the context of acute cerebrovascular stroke. Stroke 2000, 31, 1889–1892. [Google Scholar] [CrossRef]
- Brattström, L.; Lindgren, A.; Israelsson, B.; Malinow, M.R.; Norrving, B.; Upson, B.; Hamfelt, A. Hyperhomocysteinaemia in stroke: Prevalence, cause, and relationships to type of stroke and stroke risk factors. Eur. J. Clin. lnvestig. 1992, 22, 214–221. [Google Scholar] [CrossRef] [PubMed]
- Noctor, G.; Queval, G.; Mhamdi, A.; Chaouch, S.; Foyer, C.H. Glutathione. In The Arabidopsis Book; American Society of Plant Biologists: Rockville, MD, USA, 2011; Volume 9. [Google Scholar]
- Ivanov, A.V.; Maksimova, M.Y.; Nikiforova, K.A.; Ochtova, F.R.; Suanova, E.T.; Alexandrin, V.V.E.; Kruglova, M.P.; Piradov, M.A.; Kubatiev, A.A. Plasma glutathione as a risk marker for the severity and functional outcome of acute atherothrombotic and cardioem-bolic stroke. Egypt. J. Neurol. Psychiatry Neurosurg. 2022, 58, 15. [Google Scholar] [CrossRef]
- Halprin, K.M.; Ohkawara, A. The measurement of glutathione in human epi-dermis using glutathione reductase. J. Investig. Dermatol. 1967, 48, 149–152. [Google Scholar] [CrossRef] [Green Version]
- Zimmermann, C.; Winnefeld, K.; Streck, S.; Roskos, M.; Haberl, R. Antioxidant Status in Acute Stroke Patients and Patients at Stroke Risk. Eur. Neurol. 2004, 51, 157–161. [Google Scholar] [CrossRef] [Green Version]
- Aratani, Y. Myeloperoxidase: Its role for host defense, inflammation, and neutrophil function. Arch. Biochem. Biophys. 2018, 640, 47–52. [Google Scholar] [CrossRef]
- Wang, Y.C.; Lu, Y.B.; Huang, X.L.; Lao, Y.F.; Zhang, L.; Yang, J.; Shi, M.; Ma, H.L.; Pan, Y.W.; Zhang, Y.N. Myeloperoxidase: A new target for the treatment of stroke? Neural Regen. Res. 2022, 17, 1711–1716. [Google Scholar]
- Wang, Y.; Jia, Y.; Xu, Q.; Wang, R.; Sun, L.; Guo, D.; Shi, M.; Yang, P.; Wang, Y.; Liu, F.; et al. Association between myeloperoxidase and the risks of ischemic stroke, heart failure, and atrial fibrillation: A Mendelian randomization study. Nutr. Metab. Cardiovasc. Dis. 2022, 33, 210–218. [Google Scholar] [CrossRef]
- Chen, S.; Chen, H.; Du, Q.; Shen, J. Targeting Myeloperoxidase (MPO) Mediated Oxidative Stress and Inflammation for Reducing Brain Ischemia Injury: Potential Application of Natural Compounds. Front. Physiol. 2020, 11, 433. [Google Scholar] [CrossRef]
- Tziomalos, K.; Katrini, K.; Papagianni, M.; Christou, K.; Gkolfinopoulou, C.; Angelopoulou, S.-M.; Sofogianni, A.; Savopoulos, C.; Hatzitolios, A.I.; Chroni, A. Impaired antioxidative activity of high-density lipoprotein is associated with more severe acute ischemic stroke. Metabolism 2019, 98, 49–52. [Google Scholar] [CrossRef]
- Palm, F.; Pussinen, P.J.; Safer, A.; Tervahartiala, T.; Sorsa, T.; Urbanek, C.; Becher, H.; Grau, A.J. Serum matrix metalloproteinase-8, tissue inhibitor of metalloproteinase and myeloperoxidase in ischemic stroke. Atherosclerosis 2018, 271, 9–14. [Google Scholar] [CrossRef] [Green Version]
- Horváth, E.; Huțanu, A.; Chiriac, L.; Dobreanu, M.; Orădan, A.; Nagy, E.-E. Ischemic damage and early inflammatory infiltration are different in the core and penumbra lesions of rat brain after transient focal cerebral ischemia. J. Neuroimmunol. 2018, 324, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Barone, F.C.; Hillegass, L.M.; Tzimas, M.N.; Schmidt, D.B.; Foley, J.J.; White, R.F.; Price, W.J.; Feuerstein, G.Z.; Clark, R.K.; Griswold, D.E.; et al. Time-related changes in myeloperoxidase activity and leukotriene B4 receptor binding reflect leukocyte influx in cerebral focal stroke. Mol. Chem. Neuropathol. 1995, 24, 13–30. [Google Scholar] [CrossRef] [PubMed]
- Breckwoldt, M.O.; Chen, J.W.; Stangenberg, L.; Aikawa, E.; Rodriguez, E.; Qiu, S.; Moskowitz, M.A.; Weissleder, R. Tracking the inflammatory response in stroke in vivo by sensing the enzyme myeloperoxidase. Proc. Natl. Acad. Sci. USA 2008, 105, 18584–18589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forghani, R.; Kim, H.J.; Wojtkiewicz, G.; Bure, L.; Wu, Y.; Hayase, M.; Wei, Y.; Zheng, Y.; Moskowitz, A.M.; Chen, J.W. Myeloperoxidase Propagates Damage and is a Potential Therapeutic Target for Subacute Stroke. J. Cereb. Blood Flow Metab. 2015, 35, 485–493. [Google Scholar] [CrossRef]
- Kim, H.J.; Wei, Y.; Wojtkiewicz, G.R.; Lee, J.Y.; Moskowitz, A.M.; Chen, J.W. Reducing myeloperoxidase activity decreases inflammation and increases cellular protection in ischemic stroke. J. Cereb. Blood Flow Metab. 2018, 39, 1864–1877. [Google Scholar] [CrossRef]
- Podrez, E.A.; Abu-Soud, H.M.; Hazen, S.L. Myeloperoxidase-generated oxidants and atherosclerosis. Free Radic. Biol. Med. 2000, 28, 1717–1725. [Google Scholar] [CrossRef]
- Liu, D.; Liu, L.; Song, Z.; Hu, Z.; Liu, J.; Hou, D. Genetic Variations of Oxidative Stress Related Genes ALOX5, ALOX5AP and MPO Modulate Ischemic Stroke Susceptibility Through Main Effects and Epistatic Interactions in a Chinese Population. Cell. Physiol. Biochem. 2017, 43, 1588–1602. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.; Cao, F.; Xue, Y.; Tao, L.; Zhu, Y. Three Classes of Antioxidant Defense Systems and the Development of Postmenopausal Osteoporosis. Front. Physiol. 2022, 13, 199. [Google Scholar] [CrossRef]
- Gonzalez-Pinto, A.; Martinez-Cengotitabengoa, M.; Arango, C.; Baeza, I.; Otero-Cuesta, S.; Graell-Berna, M.; Soutullo, C.; Leza, J.C.; Micó, J.A. Antioxidant defense system and family environment in adolescents with family history of psychosis. BMC Psychiatry 2012, 12, 200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ciobica, A.; Balmus, I.M.; Trifan, A.; Stanciu, C. The implications of oxidative stress and antioxidant therapies in Inflammatory Bowel Disease: Clinical aspects and animal models. Saudi J. Gastroenterol. 2016, 22, 3–17. [Google Scholar] [CrossRef] [PubMed]
- Mavroudis, I.; Kazis, D.; Chowdhury, R.; Petridis, F.; Costa, V.; Balmus, I.-M.; Ciobica, A.; Luca, A.-C.; Radu, I.; Dobrin, R.P.; et al. Post-Concussion Syndrome and Chronic Traumatic Encephalopathy: Narrative Review on the Neuropathology, Neuroimaging and Fluid Biomarkers. Diagnostics 2022, 12, 740. [Google Scholar] [CrossRef]
- Krishnamurthy, P.; Wadhwani, A. Antioxidant Enzymes and Human Health. In Antioxidant Enzyme; IntechOpen: London, UK, 2012. [Google Scholar]
- Birben, E.; Sahiner, U.M.; Sackesen, C.; Erzurum, S.; Kalayci, O. Oxidative stress and antioxidant defense. World Allergy Organ. J. 2012, 5, 9–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nasiru, S.; Bulama, I.; Abdurrahman, J.H.; Abubakar, N.A.; Salisu, A.B.; Salisu, B.; Abbas, A.Y.; Yusuf, S.; Suleman, B.L. Neurobiochemical Roles of Low Molecular Weight Antioxidants on Oxidative Stress Biomarkers and Severity of Ischemic Stroke in Wistar Rats. J. Neurol. Neurol. Dis. 2018, 4, 101–111. [Google Scholar]
- Cojocaru, I.M.; Cojocaru, M.; Sapira, V.; Ionescu, A. Evaluation of oxidative stress in patients with acute ischemic stroke. Rom. J. Intern. Med. 2013, 51, 97–106. [Google Scholar] [PubMed]
- Cichon, N.; Bijak, M.; Synowiec, E.; Miller, E.; Sliwinski, T.; Saluk-Bijak, J. Modulation of antioxidant enzyme gene expression by extremely low frequency electromagnetic field in post-stroke patients. Scand. J. Clin. Lab. Investig. 2018, 78, 626–631. [Google Scholar] [CrossRef]
- Valavi, M.; Zarban, A.; Firuzabadi, M.D.; Sharifzadeh, G.; Zarezade, M. Oxidative Stress and Antioxidant Markers in Serum and Red Blood Cell of Patients with Ischemic and Hemorrhagic Stroke: A Case-Control Study. Mod. Care J. 2016, 13, 30–39. [Google Scholar] [CrossRef] [Green Version]
- Zheng, M.; Wang, X.; Yang, J.; Ma, S.; Wei, Y.; Liu, S. Changes of complement and oxidative stress parameters in patients with acute cerebral infarction or cerebral hemorrhage and the clinical significance. Exp. Ther. Med. 2019, 19, 703–709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouayed, J.; Bohn, T. Exogenous Antioxidants—Double-Edged Swords in Cellular Redox State: Health Beneficial Effects at Physiologic Doses versus Deleterious Effects at High Doses. Oxid. Med. Cell. Longev. 2010, 3, 228–237. [Google Scholar] [CrossRef] [Green Version]
- Lorente, L.; Martín, M.M.; Pérez-Cejas, A.; Abreu-González, P.; Ramos, L.; Argueso, M.; Cáceres, J.J.; Solé-Violán, J.; Jiménez, A. Association between total antioxidant capacity and mortality in ischemic stroke patients. Ann. Intensive Care 2016, 6, 39. [Google Scholar] [CrossRef] [Green Version]
- Altamura, C.; Squitti, R.; Pasqualetti, P.; Gaudino, C.; Palazzo, P.; Tibuzzi, F.; Lupoi, D.; Cortesi, M.; Rossini, P.M.; Vernieri, F. Ceruloplasmin/Transferrin System Is Related to Clinical Status in Acute Stroke. Stroke 2009, 40, 1282–1288. [Google Scholar] [CrossRef] [Green Version]
- Guldiken, B.; Demir, M.; Guldiken, S.; Turgut, N.; Turgut, B.; Tugrul, A. Oxidative Stress and Total Antioxidant Capacity in Diabetic and Nondiabetic Acute Ischemic Stroke Patients. Clin. Appl. Thromb. 2008, 15, 695–700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Del Rio, D.; Agnoli, C.; Pellegrini, N.; Krogh, V.; Brighenti, F.; Mazzeo, T.; Masala, G.; Bendinelli, B.; Berrino, F.; Sieri, S.; et al. Total Antioxidant Capacity of the Diet Is Associated with Lower Risk of Ischemic Stroke in a Large Italian Cohort. J. Nutr. 2011, 141, 118–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, G.; Shukitt-Hale, B.; Bickford, P.C.; Joseph, J.A.; McEwen, J.; Prior, R.L. Hyperoxia-induced changes in antioxidant capacity and the effect of dietary antioxidants. J. Appl. Physiol. 1999, 86, 1817–1822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prior, R.L. Oxygen radical absorbance capacity (ORAC): New horizons in relating dietary antioxidants/bioactives and health benefits. J. Funct. Foods 2015, 18, 797–810. [Google Scholar] [CrossRef]
- Niculescu, L.; Stancu, C.; Sima, A.; Toporan, D.; Simionescu, M. The total peroxyl radical trapping potential in serum—An assay to define the stage of atherosclerosis. J. Cell. Mol. Med. 2002, 5, 285–294. [Google Scholar] [CrossRef]
- Valkonen, M.; Kuusi, T. Spectrophotometric assay for total peroxyl radical-trapping antioxidant potential in human serum. J. Lipid Res. 1997, 38, 823–833. [Google Scholar] [CrossRef]
- Leinonen, J.S.; Ahonen, J.-P.; Loönnrot, K.; Jehkonen, M.; Dastidar, P.; Molnaár, G.; Alho, H. Low Plasma Antioxidant Activity Is Associated With High Lesion Volume and Neurological Impairment in Stroke. Stroke 2000, 31, 33–39. [Google Scholar] [CrossRef] [Green Version]
- Rajkovic, O.; Gourmel, C.; D’Arcy, R.; Wong, R.; Rajkovic, I.; Tirelli, N.; Pinteaux, E. Reactive Oxygen Species-Responsive Nanoparticles for the Treatment of Ischemic Stroke. Adv. Ther. 2019, 2, 1900038. [Google Scholar] [CrossRef] [Green Version]
- Baker, K.; Marcus, C.B.; Huffman, K.; Kruk, H.; Malfroy, B.; Doctrow, S.R. Synthetic combined superoxide dismutase/catalase mimetics are protective as a delayed treatment in a rat stroke model: A key role for reactive oxygen species in ischemic brain injury. J. Pharmacol. Exp. Ther. 1998, 284, 215–221. [Google Scholar]
- Ducruet, A.F.; Mack, W.J.; Mocco, J.; Hoh, D.J.; Coon, A.L.; D’Ambrosio, A.L.; Winfree, C.J.; Pinsky, D.J.; Connolly, E.S. Preclinical evaluation of post-ischemic dehydroascorbic Acid administration in a large-animal stroke model. Transl. Stroke Res. 2011, 2, 399–403. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Xiong, X.; Wu, X.; Ye, Y.; Jian, Z.; Zhi, Z.; Gu, L. Targeting Oxidative Stress and Inflammation to Prevent Ischemia-Reperfusion Injury. Front. Mol. Neurosci. 2020, 13, 28. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Zhang, X.; Xiong, X.; Zhu, H.; Chen, R.; Zhang, S.; Chen, G.; Jian, Z. Nrf2 Regulates Oxidative Stress and Its Role in Cerebral Ischemic Stroke. Antioxidants 2022, 11, 2377. [Google Scholar] [CrossRef] [PubMed]
- Puzio, M.; Moreton, N.; O’Connor, J.J. Neuroprotective strategies for acute ischemic stroke: Targeting oxidative stress and prolyl hydroxylase domain inhibition in synaptic signalling. Brain Disord. 2022, 5, 100030. [Google Scholar] [CrossRef]
- Yuan, J.; Li, L.; Yang, Q.; Ran, H.; Wang, J.; Hu, K.; Pu, W.; Huang, J.; Wen, L.; Zhou, L.; et al. Targeted Treatment of Ischemic Stroke by Bioactive Nanoparticle-Derived Reactive Oxygen Species Responsive and Inflammation-Resolving Nanotherapies. ACS Nano 2021, 15, 16076–16094. [Google Scholar] [CrossRef]
- Ayoka, T.O.; Ezema, B.O.; Eze, C.N.; Nnadi, C.O. Antioxidants for the Prevention and Treatment of Non-communicable Diseases. J. Explor. Res. Pharmacol. 2022, 7, 178–188. [Google Scholar] [CrossRef]
- Lobo, V.; Patil, A.; Phatak, A.; Chandra, N. Free radicals, antioxidants and functional foods: Impact on human health. Pharmacogn. Rev. 2010, 4, 118–126. [Google Scholar] [CrossRef] [Green Version]
- Uzombah, T.A. The Implications of Replacing Synthetic Antioxidants with Natural Ones in the Food Systems. In Natural Food Additives; Prieto, M.A., Otero, P., Eds.; IntechOpen: London, UK, 2022; Available online: https://www.intechopen.com/chapters/81679 (accessed on 14 February 2023).
- Gaire, B.P. Herbal Medicine in Ischemic Stroke: Challenges and Prospective. Chin. J. Integr. Med. 2018, 24, 243–246. [Google Scholar] [CrossRef]
- Raeeszadeh, M.; Mortazavi, P.; Atashin-Sadafi, R. The Antioxidant, Anti-Inflammatory, Pathological, and Behavioural Effects of Medicago sativa L. (Alfalfa) Extract on Brain Injury Caused by Nicotine in Male Rats. Evid.-Based Complement. Altern. Med. 2021, 2021, 6694629. [Google Scholar] [CrossRef]
- Horvat, D.; Viljevac Vuletić, M.; Andrić, L.; Baličević, R.; Kovačević Babić, M.; Tucak, M. Characterization of Forage Quality, Phenolic Profiles, and Antioxidant Activity in Alfalfa (Medicago sativa L.). Plants 2022, 11, 2735. [Google Scholar] [CrossRef]
- Quiñones-Muñoz, T.A.; Villanueva-Rodríguez, S.J.; Torruco-Uco, J.G. Nutraceutical Properties of Medicago sativa L.; Agave spp.; Zea mays L. and Avena sativa L.: A Review of Metabolites and Mechanisms. Metabolites 2022, 12, 806. [Google Scholar] [CrossRef] [PubMed]
- Raeeszadeh, M.; Rezaee, M.; Akbari, A.; Khademi, N. The comparison of the effect of Origanum vulgare L. extract and vitamin C on the gentamycin-induced nephrotoxicity in rats. Drug Chem. Toxicol. 2021, 45, 2031–2038. [Google Scholar] [CrossRef] [PubMed]
- Raeeszadeh, M.; Shokrollahi, B.; Khademi, N.; Akbari, A. Superior effect of broccoli methanolic extract on control of oxidative damage of sperm cryopreservation and reproductive performance in rats: A comparison with vitamin C and E antioxidant. Theriogenology 2022, 181, 50–58. [Google Scholar] [CrossRef]
- E Silva, M.L.A.; Lucarini, R.; dos Santos, F.F.; Martins, C.H.G.; Pauletti, P.M.; Januario, A.H.; Santos, M.F.C.; Cunha, W.R. Hypoglycemic effect of rosmarinic acid-rich infusion (RosCE) from Origanum vulgare in alloxan-induced diabetic rats. Nat. Prod. Res. 2021, 36, 4519–4525. [Google Scholar] [CrossRef]
- Hwang, J.-H.; Lim, S.-B. Antioxidant and Anti-inflammatory Activities of Broccoli Florets in LPS-stimulated RAW 264.7 Cells. Prev. Nutr. Food Sci. 2014, 19, 89–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, N.; Singhvi, R. Effects of Chemical Fertilizers and Pesticides on Human Health and Environment: A Review. Int. J. Agric. Environ. Biotechnol. 2017, 10, 675. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kamal, F.Z.; Lefter, R.; Jaber, H.; Balmus, I.-M.; Ciobica, A.; Iordache, A.-C. The Role of Potential Oxidative Biomarkers in the Prognosis of Acute Ischemic Stroke and the Exploration of Antioxidants as Possible Preventive and Treatment Options. Int. J. Mol. Sci. 2023, 24, 6389. https://doi.org/10.3390/ijms24076389
Kamal FZ, Lefter R, Jaber H, Balmus I-M, Ciobica A, Iordache A-C. The Role of Potential Oxidative Biomarkers in the Prognosis of Acute Ischemic Stroke and the Exploration of Antioxidants as Possible Preventive and Treatment Options. International Journal of Molecular Sciences. 2023; 24(7):6389. https://doi.org/10.3390/ijms24076389
Chicago/Turabian StyleKamal, Fatima Zahra, Radu Lefter, Hassna Jaber, Ioana-Miruna Balmus, Alin Ciobica, and Alin-Constantin Iordache. 2023. "The Role of Potential Oxidative Biomarkers in the Prognosis of Acute Ischemic Stroke and the Exploration of Antioxidants as Possible Preventive and Treatment Options" International Journal of Molecular Sciences 24, no. 7: 6389. https://doi.org/10.3390/ijms24076389