
Sigma-2 Receptor Ligand Binding Modulates Association between TSPO and TMEM97
 , , , , , , , ,
, , , , , , , ,  , , , , , , , , and
, , , , , , , , and
Abstract
:1. Introduction
2. Results
2.1. TSPO and TMEM97 Are Bound by the S2R Ligand SW120 in MP Cells
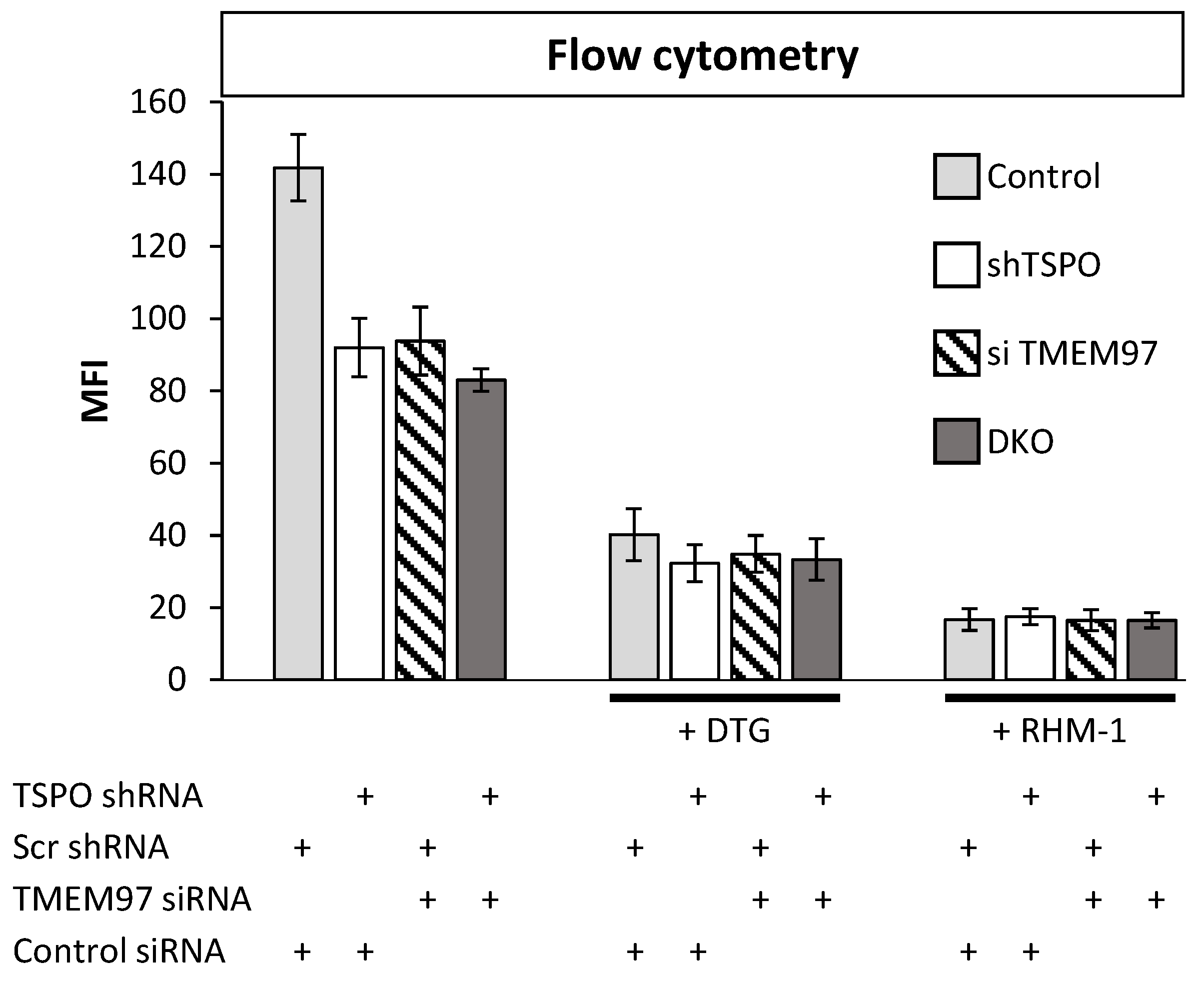
2.2. TSPO and TMEM97 Affect the Same SW120-Binding Sites in MP Cells
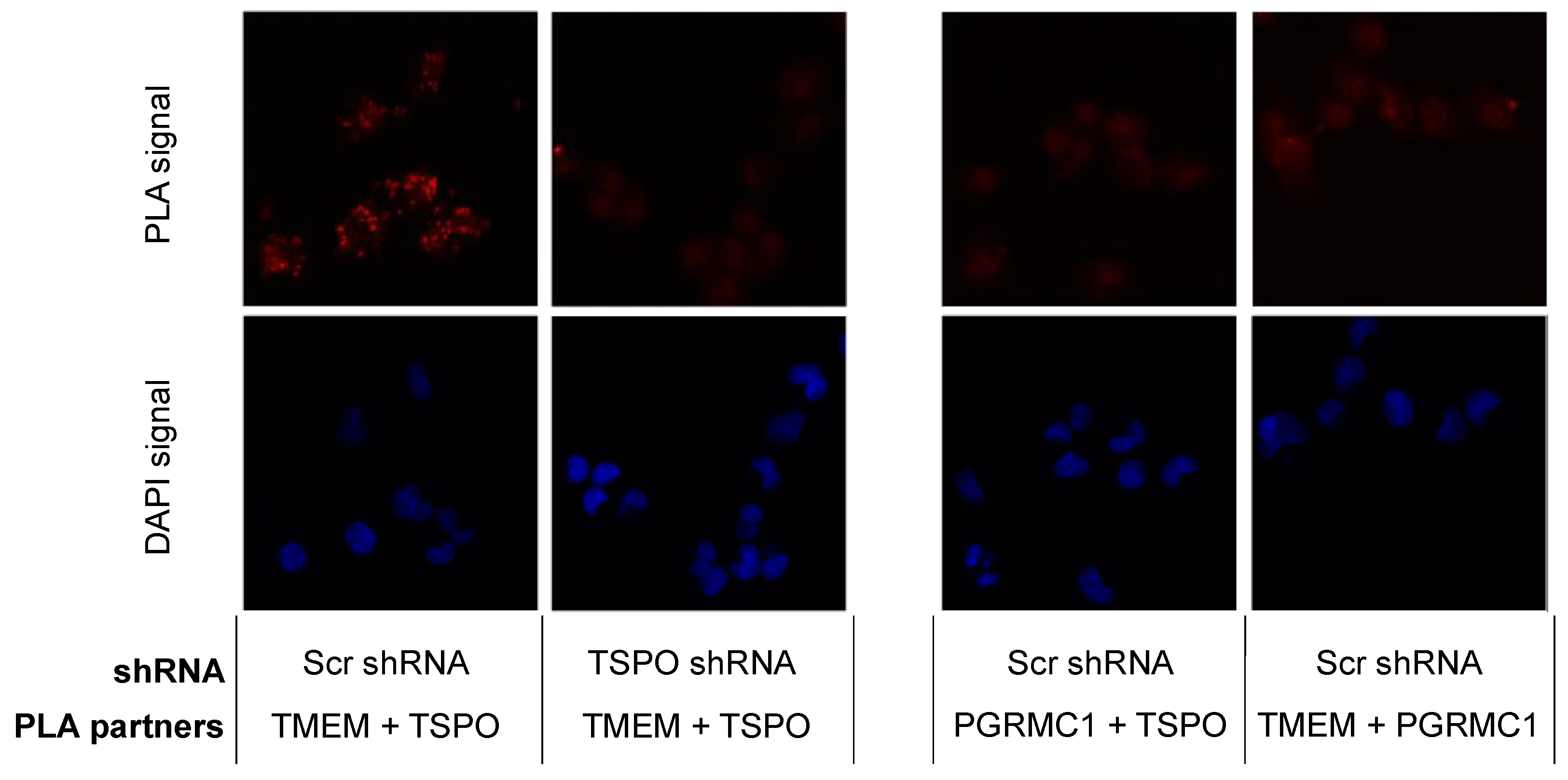
2.3. TSPO Colocalizes with TMEM97 in MP Cells
2.4. Saturation Analyses by Radioligand Binding to Detect TMEM97 and TSPO in Functionally Different Cell Lines
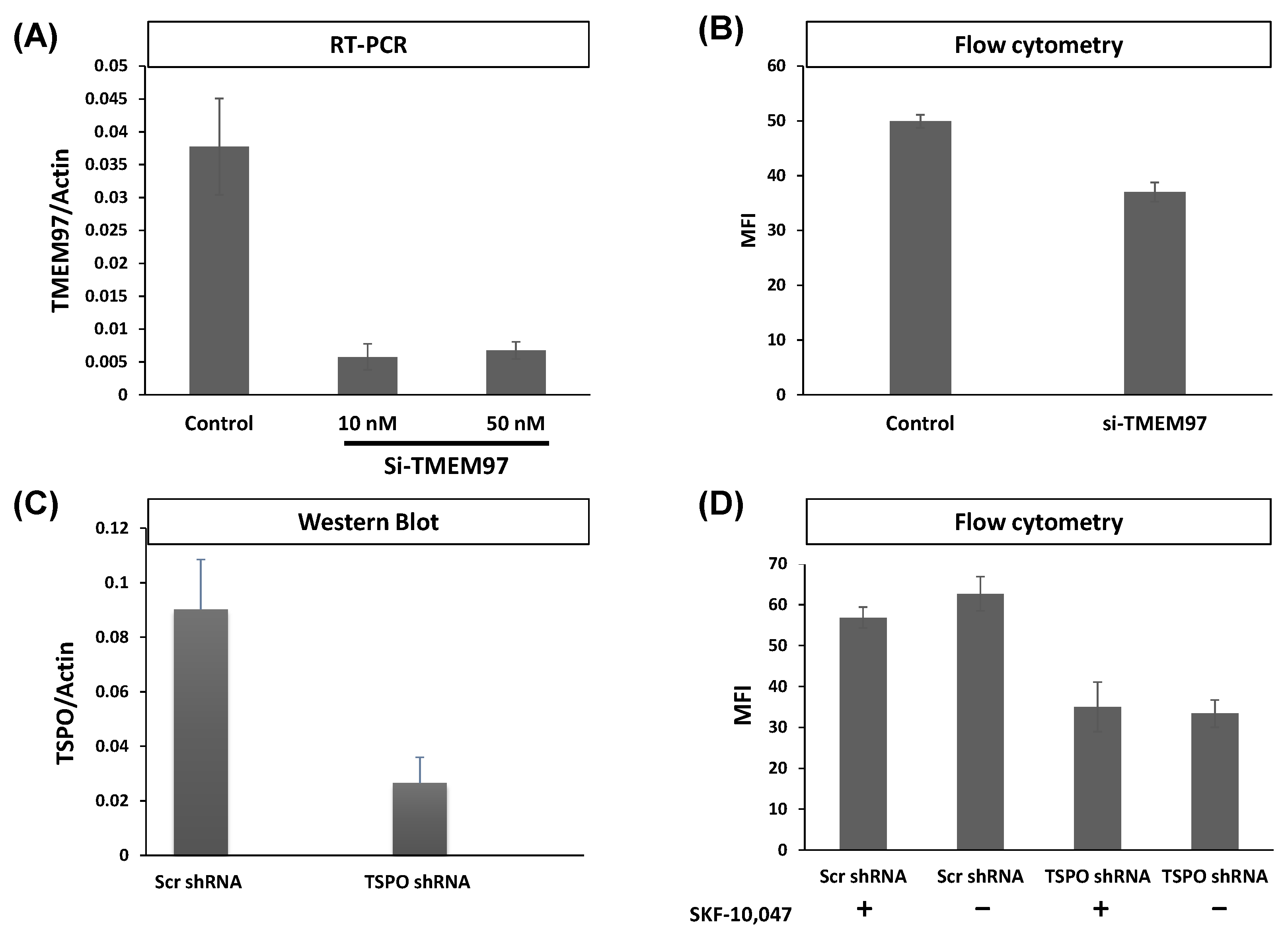
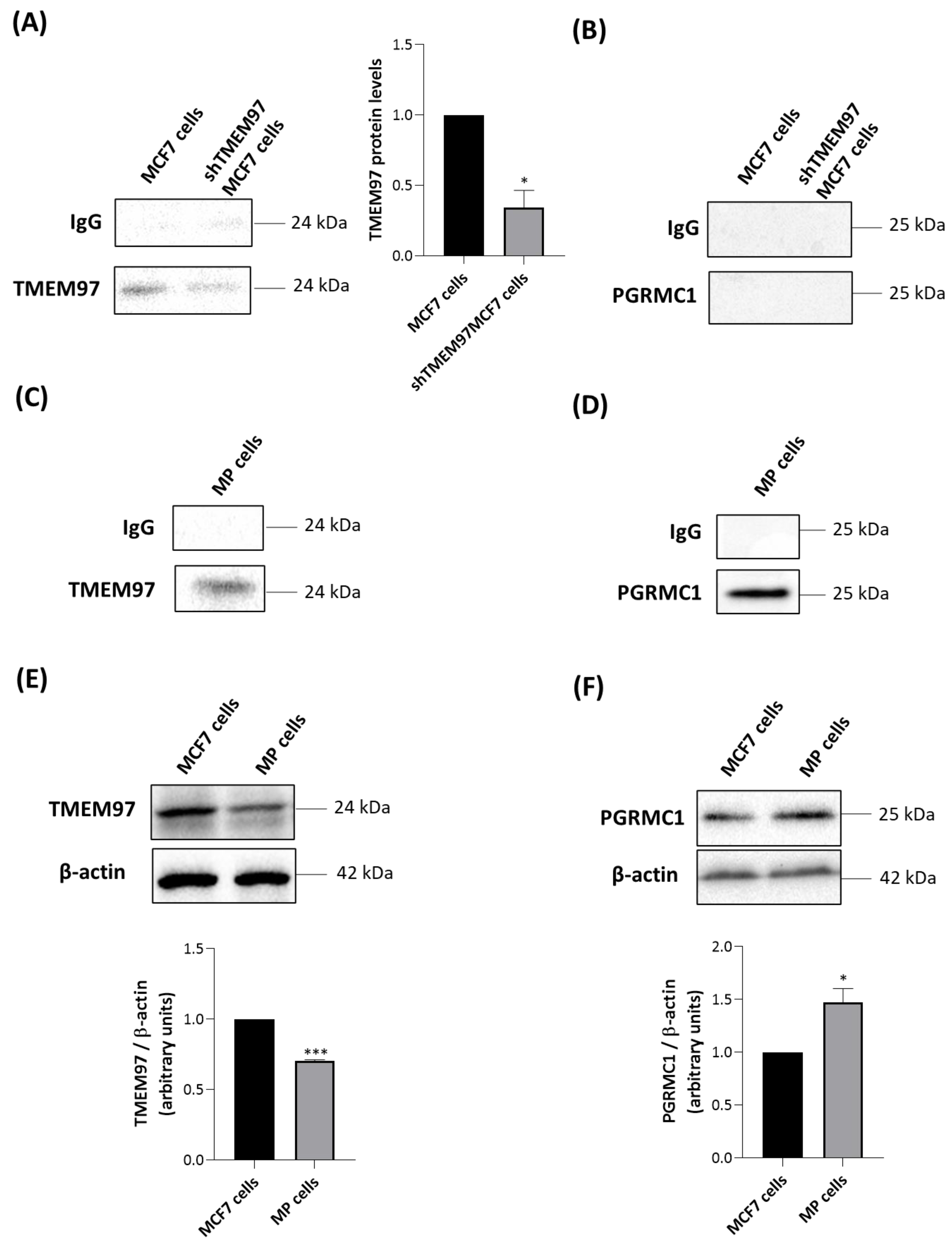
2.5. Generation of MCF7 and MP Cells with Attenuated Levels of TMEM97/S2R
2.6. TMEM97 Co-Immunoprecipitates with TSPO
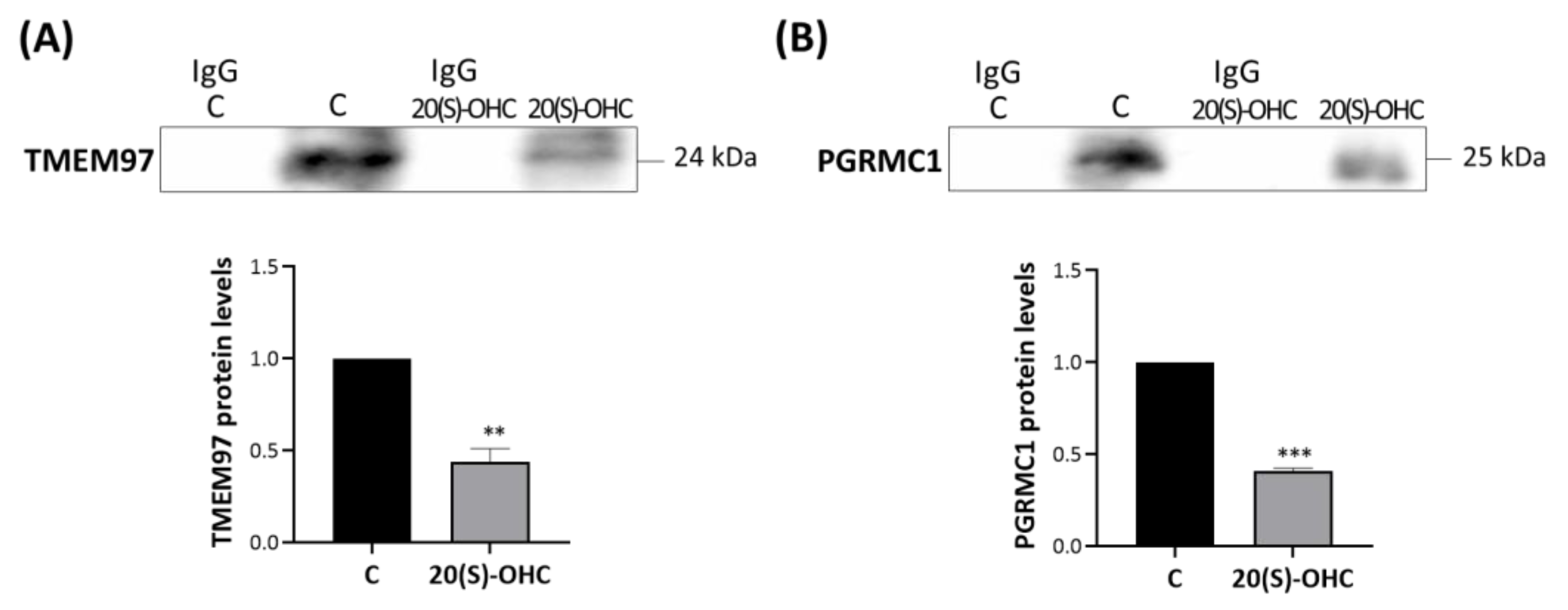
2.7. 20S-OHC Reduced the Associations TSPO-TMEM97 and TSPO-PGRMC1 in MP Cells
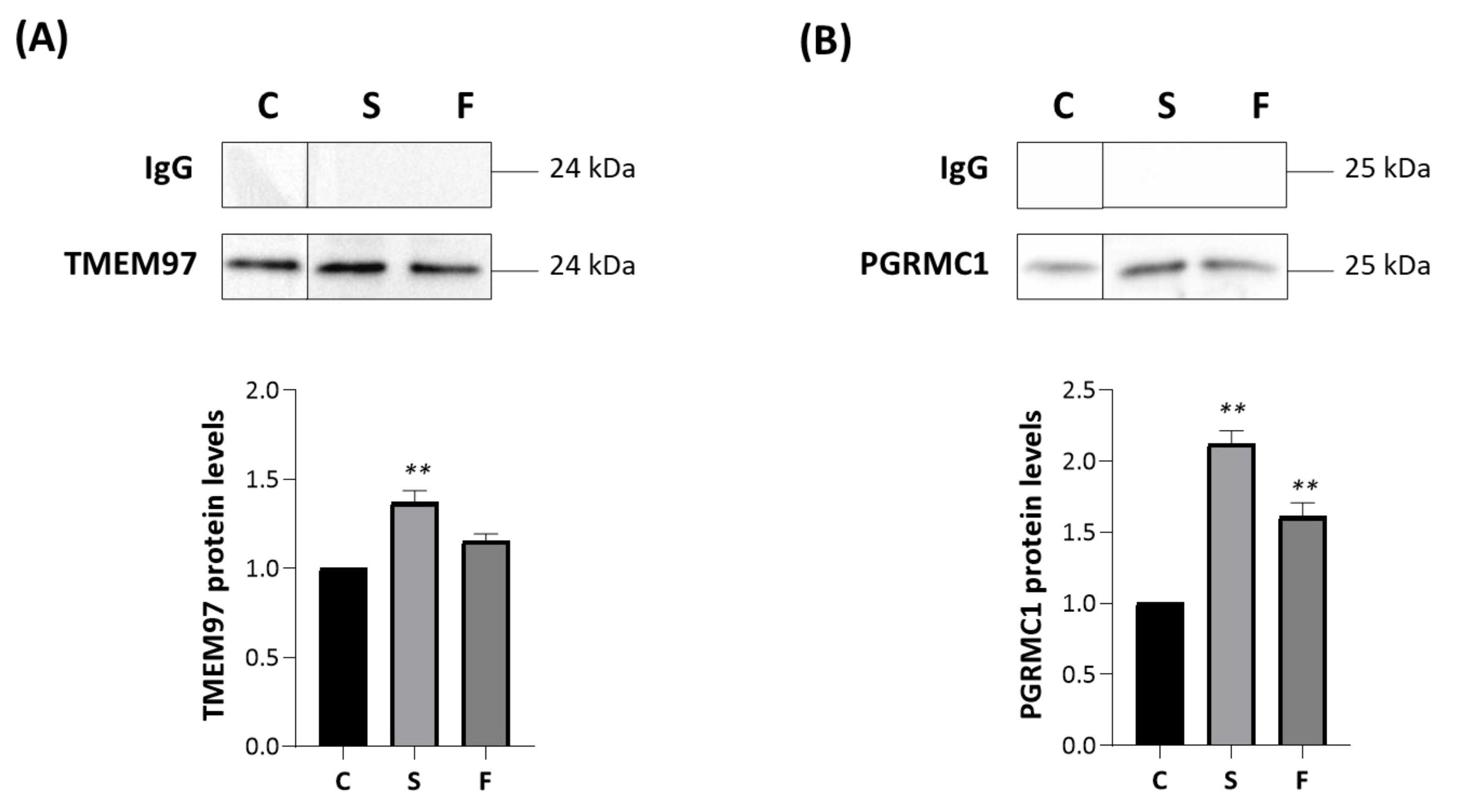
2.8. Effect of Siramesine Oxalate Salt and FA10 Hydrochloric Salt on TSPO Interactions-With TMEM97 and PGRMC1 in MP Cells
3. Discussion
4. Methods
4.1. Cell Culture
4.2. Saturation Binding Assay
4.2.1. Membrane Preparation
4.2.2. Saturation Binding Assay by Radioligand at TSPO
4.2.3. Saturation Binding Assay by Radioligand at TMEM97/Sigma-2 Receptor
4.2.4. Materials
4.3. Co-Immunoprecipitation
4.4. Western Blotting
4.5. Statistical Analysis
4.6. shRNA Knockdown and Cell Lines Generation
4.7. siRNA Knockdown of TMEM97 in MP Cells
4.8. MCF7 Transfection with sh_RNA Targeting TMEM97
4.9. RT-PCR
4.10. Flow Cytometry
4.11. Binding Assay for SW120 and SKF 10,047
4.12. Proximity Ligation Assay
4.13. Confocal Microscopy Studies
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hornick, J.R.; Spitzer, D.; Goedegebuure, P.; Mach, R.H.; Hawkins, W.G. Therapeutic targeting of pancreatic cancer utilizing sigma-2 ligands. Surgery 2012, 152, S152–S156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mach, R.H.; Smith, C.R.; Al-Nabulsi, I.; Whirrett, B.R.; Childers, S.R.; Wheeler, K.T. Sigma 2 receptors as potential biomarkers of proliferation in breast cancer. Cancer Res. 1997, 57, 156–161. [Google Scholar] [PubMed]
- Mach, R.H.; Zeng, C.; Hawkins, W.G. The sigma2 receptor: A novel protein for the imaging and treatment of cancer. J. Med. Chem. 2013, 56, 7137–7160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yi, B.; Sahn, J.J.; Ardestani, P.M.; Evans, A.K.; Scott, L.L.; Chan, J.Z.; Iyer, S.; Crisp, A.; Zuniga, G.; Pierce, J.T.; et al. Small molecule modulator of sigma 2 receptor is neuroprotective and reduces cognitive deficits and neuroinflammation in experimental models of Alzheimer’s disease. J. Neurochem. 2017, 140, 561–575. [Google Scholar] [CrossRef] [Green Version]
- Blass, B.E.; Rogers, J.P. The sigma-2 (sigma-2) receptor: A review of recent patent applications: 2013–2018. Expert. Opin. Ther. Pat. 2018, 28, 655–663. [Google Scholar] [CrossRef]
- Zeng, C.; Vangveravong, S.; Jones, L.A.; Hyrc, K.; Chang, K.C.; Xu, J.; Rothfuss, J.M.; Goldberg, M.P.; Hotchkiss, R.S.; Mach, R.H. Characterization and evaluation of two novel fluorescent sigma-2 receptor ligands as proliferation probes. Mol. Imaging 2011, 10, 420–433. [Google Scholar] [CrossRef] [Green Version]
- Zeng, C.; Weng, C.C.; Schneider, M.E., Jr.; Puentes, L.; Riad, A.; Xu, K.; Makvandi, M.; Jin, L.; Hawkins, W.G.; Mach, R.H. TMEM97 and PGRMC1 do not mediate sigma-2 ligand-induced cell death. Cell Death Discov. 2019, 5, 58. [Google Scholar] [CrossRef] [Green Version]
- Hanner, M.; Moebius, F.F.; Flandorfer, A.; Knaus, H.G.; Striessnig, J.; Kempner, E.; Glossmann, H. Purification, molecular cloning, and expression of the mammalian sigma1-binding site. Proc. Natl. Acad. Sci. USA 1996, 93, 8072–8077. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, H.R.; Betz, R.M.; Dror, R.O.; Kruse, A.C. Structural basis for σ(1) receptor ligand recognition. Nat. Struct. Mol. Biol. 2018, 25, 981–987. [Google Scholar] [CrossRef]
- Xu, J.; Zeng, C.; Chu, W.; Pan, F.; Rothfuss, J.M.; Zhang, F.; Tu, Z.; Zhou, D.; Zeng, D.; Vangveravong, S.; et al. Identification of the PGRMC1 protein complex as the putative sigma-2 receptor binding site. Nature Commun. 2011, 2, 380. [Google Scholar] [CrossRef] [Green Version]
- Cahill, M.A.; Jazayeri, J.A.; Kovacevic, Z.; Richardson, D.R. PGRMC1 regulation by phosphorylation: Potential new insights in controlling biological activity. Oncotarget 2016, 7, 50822–50827. [Google Scholar] [CrossRef] [Green Version]
- Cahill, M.A.; Medlock, A.E. Thoughts on interactions between PGRMC1 and diverse attested and potential hydrophobic ligands. J. Steroid Biochem. Mol. Biol. 2017, 171, 11–33. [Google Scholar] [CrossRef]
- Abate, C.; Niso, M.; Infantino, V.; Menga, A.; Berardi, F. Elements in support of the ’non-identity’ of the PGRMC1 protein with the sigma2 receptor. Eur. J. Pharmacol. 2015, 758, 16–23. [Google Scholar] [CrossRef]
- Pati, M.L.; Groza, D.; Riganti, C.; Kopecka, J.; Niso, M.; Berardi, F.; Hager, S.; Heffeter, P.; Hirai, M.; Tsugawa, H.; et al. Sigma-2 receptor and progesterone receptor membrane component 1 (PGRMC1) are two different proteins: Proofs by fluorescent labeling and binding of sigma-2 receptor ligands to PGRMC1. Pharmacol. Res. 2017, 117, 67–74. [Google Scholar] [CrossRef]
- Hiranita, T. Identification of the Sigma-2 Receptor: Distinct from the Progesterone Receptor Membrane Component 1 (PGRMC1). J. Alcohol. Drug. Depend. 2016, 4, e130. [Google Scholar] [CrossRef] [Green Version]
- Chu, U.B.; Mavlyutov, T.A.; Chu, M.L.; Yang, H.; Schulman, A.; Mesangeau, C.; McCurdy, C.R.; Guo, L.W.; Ruoho, A.E. The Sigma-2 Receptor and Progesterone Receptor Membrane Component 1 are Different Binding Sites Derived From Independent Genes. EBioMedicine 2015, 2, 1806–1813. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, I.S.; Chamberlain, C.; Craven, R.J. S2R(Pgrmc1): The cytochrome-related sigma-2 receptor that regulates lipid and drug metabolism and hormone signaling. Expert. Opin. Drug. Metab. Toxicol. 2012, 8, 361–370. [Google Scholar] [CrossRef]
- Kabe, Y.; Nakane, T.; Koike, I.; Yamamoto, T.; Sugiura, Y.; Harada, E.; Sugase, K.; Shimamura, T.; Ohmura, M.; Muraoka, K.; et al. Haem-dependent dimerization of PGRMC1/Sigma-2 receptor facilitates cancer proliferation and chemoresistance. Nature Commun. 2016, 7, 11030. [Google Scholar] [CrossRef] [Green Version]
- Izzo, N.J.; Xu, J.; Zeng, C.; Kirk, M.J.; Mozzoni, K.; Silky, C.; Rehak, C.; Yurko, R.; Look, G.; Rishton, G.; et al. Alzheimer’s therapeutics targeting amyloid beta 1-42 oligomers II: Sigma-2/PGRMC1 receptors mediate Abeta 42 oligomer binding and synaptotoxicity. PLoS ONE 2014, 9, e111899. [Google Scholar] [CrossRef] [PubMed]
- Alon, A.; Schmidt, H.R.; Wood, M.D.; Sahn, J.J.; Martin, S.F.; Kruse, A.C. Identification of the gene that codes for the sigma2 receptor. Proc. Natl. Acad. Sci. USA 2017, 114, 7160–7165. [Google Scholar] [CrossRef] [Green Version]
- Riad, A.; Zeng, C.; Weng, C.C.; Winters, H.; Xu, K.; Makvandi, M.; Metz, T.; Carlin, S.; Mach, R.H. Sigma-2 Receptor/TMEM97 and PGRMC-1 Increase the Rate of Internalization of LDL by LDL Receptor through the Formation of a Ternary Complex. Sci. Rep. 2018, 8, 16845. [Google Scholar] [CrossRef] [PubMed]
- Alon, A.; Lyu, J.; Braz, J.M.; Tummino, T.A.; Craik, V.; O’Meara, M.J.; Webb, C.M.; Radchenko, D.S.; Moroz, Y.S.; Huang, X.P.; et al. Structures of the σ(2) receptor enable docking for bioactive ligand discovery. Nature 2021, 600, 759–764. [Google Scholar] [CrossRef] [PubMed]
- Papadopoulos, V.; Baraldi, M.; Guilarte, T.R.; Knudsen, T.B.; Lacapere, J.J.; Lindemann, P.; Norenberg, M.D.; Nutt, D.; Weizman, A.; Zhang, M.R.; et al. Translocator protein (18kDa): New nomenclature for the peripheral-type benzodiazepine receptor based on its structure and molecular function. Trends Pharmacol. Sci. 2006, 27, 402–409. [Google Scholar] [CrossRef] [PubMed]
- Tu, L.N.; Zhao, A.H.; Hussein, M.; Stocco, D.M.; Selvaraj, V. Translocator Protein (TSPO) Affects Mitochondrial Fatty Acid Oxidation in Steroidogenic Cells. Endocrinology 2016, 157, 1110–1121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Selvaraj, V.; Tu, L.N. Current status and future perspectives: TSPO in steroid neuroendocrinology. J. Endocrinol. 2016, 231, R1–R30. [Google Scholar] [CrossRef] [Green Version]
- Cantonero, C.; Camello, P.J.; Abate, C.; Berardi, F.; Salido, G.M.; Rosado, J.A.; Redondo, P.C. NO1, a New Sigma 2 Receptor/TMEM97 Fluorescent Ligand, Downregulates SOCE and Promotes Apoptosis in the Triple Negative Breast Cancer Cell Lines. Cancers 2020, 12, 257. [Google Scholar] [CrossRef] [Green Version]
- Anobile, D.P.; Niso, M.; Puerta, A.; Fraga Rodrigues, S.M.; Abatematteo, F.S.; Avan, A.; Abate, C.; Riganti, C.; Giovannetti, E. New Pharmacological Strategies against Pancreatic Adenocarcinoma: The Multifunctional Thiosemicarbazone FA4. Molecules 2022, 27, 1682. [Google Scholar] [CrossRef]
- Cassano, G.; Gasparre, G.; Niso, M.; Contino, M.; Scalera, V.; Colabufo, N.A. F281, synthetic agonist of the sigma-2 receptor, induces Ca2+ efflux from the endoplasmic reticulum and mitochondria in SK-N-SH cells. Cell Calcium 2009, 45, 340–345. [Google Scholar] [CrossRef]
- Bagchi, S.; Fredriksson, R.; Wallén-Mackenzie, Å. In Situ Proximity Ligation Assay (PLA). Methods Mol. Biol. 2015, 1318, 149–159. [Google Scholar]
- Cheng, Y.S.; Zhang, T.; Ma, X.; Pratuangtham, S.; Zhang, G.C.; Ondrus, A.A.; Mafi, A.; Lomenick, B.; Jones, J.J.; Ondrus, A.E. A proteome-wide map of 20(S)-hydroxycholesterol interactors in cell membranes. Nat. Chem. Biol. 2021, 17, 1271–1280. [Google Scholar] [CrossRef]
- Zou, L.B.; Yamada, K.; Nabeshima, T. Sigma receptor ligands (+)-SKF10,047 and SA4503 improve dizocilpine-induced spatial memory deficits in rats. Eur. J. Pharmacol. 1998, 355, 1–10. [Google Scholar] [CrossRef]
- Zeng, C.; Garg, N.; Mach, R.H. The PGRMC1 Protein Level Correlates with the Binding Activity of a Sigma-2 Fluorescent Probe (SW120) in Rat Brain Cells. Mol. Imaging Biol. 2016, 18, 172–179. [Google Scholar] [CrossRef]
- Xu, J.; Tu, Z.; Jones, L.A.; Vangveravong, S.; Wheeler, K.T.; Mach, R.H. [3H]N-[4-(3,4-dihydro-6,7-dimethoxyisoquinolin-2(1H)-yl)butyl]-2-methoxy-5-methylbenzamide: A novel sigma-2 receptor probe. Eur. J. Pharmacol. 2005, 525, 8–17. [Google Scholar] [CrossRef]
- Weng, C.C.; Riad, A.; Lieberman, B.P.; Xu, K.; Peng, X.; Mikitsh, J.L.; Mach, R.H. Characterization of Sigma-2 Receptor-Specific Binding Sites Using [(3)H]DTG and [(125)I]RHM-4. Pharmaceuticals 2022, 15, 1564. [Google Scholar] [CrossRef]
- Moon, B.S.; Jung, J.H.; Park, H.S.; Contino, M.; Denora, N.; Lee, B.C.; Kim, S.E. Preclinical comparison study between [(18)F]fluoromethyl-PBR28 and its deuterated analog in a rat model of neuroinflammation. Bioorg. Med. Chem. Lett. 2018, 28, 2925–2929. [Google Scholar] [CrossRef]
- Perrone, M.; Moon, B.S.; Park, H.S.; Laquintana, V.; Jung, J.H.; Cutrignelli, A.; Lopedota, A.; Franco, M.; Kim, S.E.; Lee, B.C.; et al. A Novel PET Imaging Probe for the Detection and Monitoring of Translocator Protein 18 kDa Expression in Pathological Disorders. Sci. Rep. 2016, 6, 20422. [Google Scholar] [CrossRef]
- Denora, N.; Laquintana, V.; Lopalco, A.; Iacobazzi, R.M.; Lopedota, A.; Cutrignelli, A.; Iacobellis, G.; Annese, C.; Cascione, M.; Leporatti, S.; et al. In vitro targeting and imaging the translocator protein TSPO 18-kDa through G(4)-PAMAM-FITC labeled dendrimer. J. Control. Release 2013, 172, 1111–1125. [Google Scholar] [CrossRef]
- Colabufo, N.A.; Berardi, F.; Contino, M.; Niso, M.; Abate, C.; Perrone, R.; Tortorella, V. Antiproliferative and cytotoxic effects of some sigma2 agonists and sigma1 antagonists in tumour cell lines. Naunyn Schmiedebergs Arch. Pharmacol. 2004, 370, 106–113. [Google Scholar] [CrossRef]
- Niso, M.; Kopecka, J.; Abatematteo, F.S.; Berardi, F.; Riganti, C.; Abate, C. Multifunctional thiosemicarbazones targeting sigma receptors: In vitro and in vivo antitumor activities in pancreatic cancer models. Cell. Oncol. 2021, 44, 1307–1323. [Google Scholar] [CrossRef]
- Pati, M.L.; Niso, M.; Spitzer, D.; Berardi, F.; Contino, M.; Riganti, C.; Hawkins, W.G.; Abate, C. Multifunctional thiosemicarbazones and deconstructed analogues as a strategy to study the involvement of metal chelation, Sigma-2 (σ(2)) receptor and P-gp protein in the cytotoxic action: In vitro and in vivo activity in pancreatic tumors. Eur. J. Med. Chem. 2018, 144, 359–371. [Google Scholar] [CrossRef]
- Abate, C.; Niso, M.; Berardi, F. Sigma-2 receptor: Past, present and perspectives on multiple therapeutic exploitations. Future Med. Chem. 2018, 10, 1997–2018. [Google Scholar] [CrossRef] [PubMed]
- Pati, M.L.; Niso, M.; Ferorelli, S.; Abate, C.; Berardi, F. Novel metal chelators thiosemicarbazones with activity at the σ2 receptors and P-glycoprotein: An innovative strategy for resistant tumor treatment. RSC Adv. 2015, 5, 103131–103146. [Google Scholar] [CrossRef]
- Abatematteo, F.S.; Majellaro, M.; Montsch, B.; Prieto-Díaz, R.; Niso, M.; Contino, M.; Stefanachi, A.; Riganti, C.; Mangiatordi, G.F.; Delre, P.; et al. Development of Fluorescent 4-[4-(3H-Spiro[isobenzofuran-1,4′-piperidin]-1′-yl)butyl]indolyl Derivatives as High-Affinity Probes to Enable the Study of σ Receptors via Fluorescence-Based Techniques. J. Med. Chem. 2023; in press. [Google Scholar] [CrossRef] [PubMed]
- Ostenfeld, M.S.; Høyer-Hansen, M.; Bastholm, L.; Fehrenbacher, N.; Olsen, O.D.; Groth-Pedersen, L.; Puustinen, P.; Kirkegaard-Sørensen, T.; Nylandsted, J.; Farkas, T.; et al. Anti-cancer agent siramesine is a lysosomotropic detergent that induces cytoprotective autophagosome accumulation. Autophagy 2008, 4, 487–499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niso, M.; Pati, M.L.; Berardi, F.; Abate, C. Rigid versus flexible anilines or anilides confirm the bicyclic ring as the hydrophobic portion for optimal σ2 receptor binding and provide novel tools for the development of future σ2 receptor PET radiotracers. RSC Adv. 2016, 6, 88508–88518. [Google Scholar] [CrossRef]
- Schmit, K.; Michiels, C. TMEM Proteins in Cancer: A Review. Front. Pharmacol. 2018, 9, 1345. [Google Scholar] [CrossRef] [Green Version]
- Bartz, F.; Kern, L.; Erz, D.; Zhu, M.; Gilbert, D.; Meinhof, T.; Wirkner, U.; Erfle, H.; Muckenthaler, M.; Pepperkok, R.; et al. Identification of cholesterol-regulating genes by targeted RNAi screening. Cell Metab. 2009, 10, 63–75. [Google Scholar] [CrossRef] [Green Version]
- Riad, A.; Lengyel-Zhand, Z.; Zeng, C.; Weng, C.C.; Lee, V.M.; Trojanowski, J.Q.; Mach, R.H. The Sigma-2 Receptor/TMEM97, PGRMC1, and LDL Receptor Complex Are Responsible for the Cellular Uptake of Abeta42 and Its Protein Aggregates. Mol. Neurobiol. 2020, 57, 3803–3813. [Google Scholar] [CrossRef]
- Barresi, E.; Robello, M.; Costa, B.; Da Pozzo, E.; Baglini, E.; Salerno, S.; Da Settimo, F.; Martini, C.; Taliani, S. An update into the medicinal chemistry of translocator protein (TSPO) ligands. Eur. J. Med. Chem. 2021, 209, 112924. [Google Scholar] [CrossRef]
- Rosa, M.; Noel, T.; Harris, M.; Ladds, G. Emerging roles of adhesion G protein-coupled receptors. Biochem. Soc. Trans. 2021, 49, 1695–1709. [Google Scholar] [CrossRef]
- Denora, N.; Natile, G. An Updated View of Translocator Protein (TSPO). Int. J. Mol. Sci. 2017, 18, 2640. [Google Scholar] [CrossRef] [Green Version]
- Manku, G.; Wang, Y.; Thuillier, R.; Rhodes, C.; Culty, M. Developmental expression of the translocator protein 18 kDa (TSPO) in testicular germ cells. Curr. Mol. Med. 2012, 12, 467–475. [Google Scholar]
- Gonzales, P.A.; Pisitkun, T.; Hoffert, J.D.; Tchapyjnikov, D.; Star, R.A.; Kleta, R.; Wang, N.S.; Knepper, M.A. Large-scale proteomics and phosphoproteomics of urinary exosomes. J. Am. Soc. Nephrol. 2009, 20, 363–379. [Google Scholar] [CrossRef] [Green Version]
- Cahill, M.A.; Jazayeri, J.A.; Catalano, S.M.; Toyokuni, S.; Kovacevic, Z.; Richardson, D.R. The emerging role of progesterone receptor membrane component 1 (PGRMC1) in cancer biology. Biochim. Biophys. Acta 2016, 1866, 339–349. [Google Scholar] [CrossRef]
- Piel, R.B., 3rd; Shiferaw, M.T.; Vashisht, A.A.; Marcero, J.R.; Praissman, J.L.; Phillips, J.D.; Wohlschlegel, J.A.; Medlock, A.E. A Novel Role for Progesterone Receptor Membrane Component 1 (PGRMC1): A Partner and Regulator of Ferrochelatase. Biochemistry 2016, 55, 5204–5217. [Google Scholar] [CrossRef] [Green Version]
- Zeng, C.; Riad, A.; Mach, R.H. The Biological Function of Sigma-2 Receptor/TMEM97 and Its Utility in PET Imaging Studies in Cancer. Cancers 2020, 12, 1877. [Google Scholar] [CrossRef]
- Cahill, M.A. Unde venisti PGRMC? Grand-Scale Biology from Early Eukaryotes and Eumetazoan Animal Origins. Front. Biosci. 2022, 27, 317. [Google Scholar] [CrossRef]
- Tamarit, D.; Teakel, S.; Marama, M.; Aragao, D.; Gerdes, S.Y.; Forwood, J.K.; Ettema, T.J.G.; Cahill, M.A. MAPR origins reveal a new class of prokaryotic cytochrome b5 proteins and possible role in eukaryogenesis. bioRxiv 2021. [Google Scholar] [CrossRef]
- Zhang, X.; Krause, K.H.; Xenarios, I.; Soldati, T.; Boeckmann, B. Evolution of the ferric reductase domain (FRD) superfamily: Modularity, functional diversification, and signature motifs. PLoS ONE 2013, 8, e58126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santarsiero, A.; Convertini, P.; Todisco, S.; Pierri, C.L.; De Grassi, A.; Williams, N.C.; Iacobazzi, D.; De Stefano, G.; O’Neill, L.A.J.; Infantino, V. ACLY Nuclear Translocation in Human Macrophages Drives Proinflammatory Gene Expression by NF-κB Acetylation. Cells 2021, 10, 2962. [Google Scholar] [CrossRef]
- Thejer, B.M.; Adhikary, P.P.; Kaur, A.; Teakel, S.L.; Van Oosterum, A.; Seth, I.; Pajic, M.; Hannan, K.M.; Pavy, M.; Poh, P.; et al. PGRMC1 phosphorylation affects cell shape, motility, glycolysis, mitochondrial form and function, and tumor growth. BMC Mol. Cell. Biol. 2020, 21, 24. [Google Scholar] [CrossRef] [Green Version]
- Gurusinghe, S.; Young, P.; Michelsen, J.; Strappe, P. Suppression of dedifferentiation and hypertrophy in canine chondrocytes through lentiviral vector expression of Sox9 and induced pluripotency stem cell factors. Biotechnol. Lett. 2015, 37, 1495–1504. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Lines | Bmax, pmol/mg of Protein | |
|---|---|---|
| TSPO | TMEM97/S2R | |
| C6 rat glioma | 3.60 | 2.15 |
| MCF7 | 1.02 | 1.795 |
| shMCF7TMEM97 | 2.06 | 0.891 |
| LoVo | 3.43 | 0.54 |
| Mia PaCa-2 (MP) | 7.30 | 1.14 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thejer, B.M.; Infantino, V.; Santarsiero, A.; Pappalardo, I.; Abatematteo, F.S.; Teakel, S.; Van Oosterum, A.; Mach, R.H.; Denora, N.; Lee, B.C.; et al. Sigma-2 Receptor Ligand Binding Modulates Association between TSPO and TMEM97. Int. J. Mol. Sci. 2023, 24, 6381. https://doi.org/10.3390/ijms24076381
Thejer BM, Infantino V, Santarsiero A, Pappalardo I, Abatematteo FS, Teakel S, Van Oosterum A, Mach RH, Denora N, Lee BC, et al. Sigma-2 Receptor Ligand Binding Modulates Association between TSPO and TMEM97. International Journal of Molecular Sciences. 2023; 24(7):6381. https://doi.org/10.3390/ijms24076381
Chicago/Turabian StyleThejer, Bashar M., Vittoria Infantino, Anna Santarsiero, Ilaria Pappalardo, Francesca S. Abatematteo, Sarah Teakel, Ashleigh Van Oosterum, Robert H. Mach, Nunzio Denora, Byung Chul Lee, and et al. 2023. "Sigma-2 Receptor Ligand Binding Modulates Association between TSPO and TMEM97" International Journal of Molecular Sciences 24, no. 7: 6381. https://doi.org/10.3390/ijms24076381