
Effects of Exogenous Ergothioneine on Brassica rapa Clubroot Development Revealed by Transcriptomic Analysis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Effects of Different EGT Concentrations on the Activity of the Resting Spores of P. brassicae
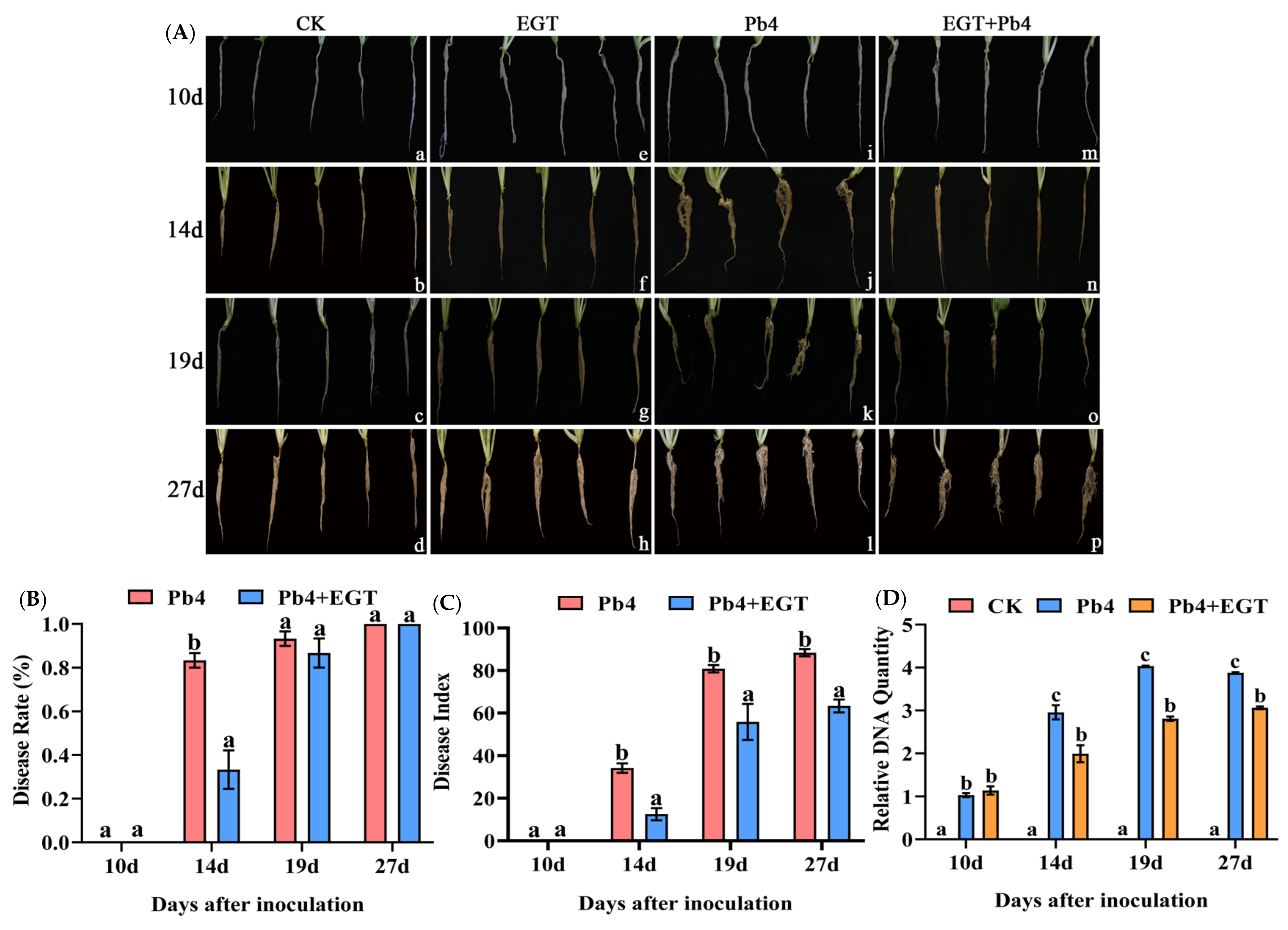
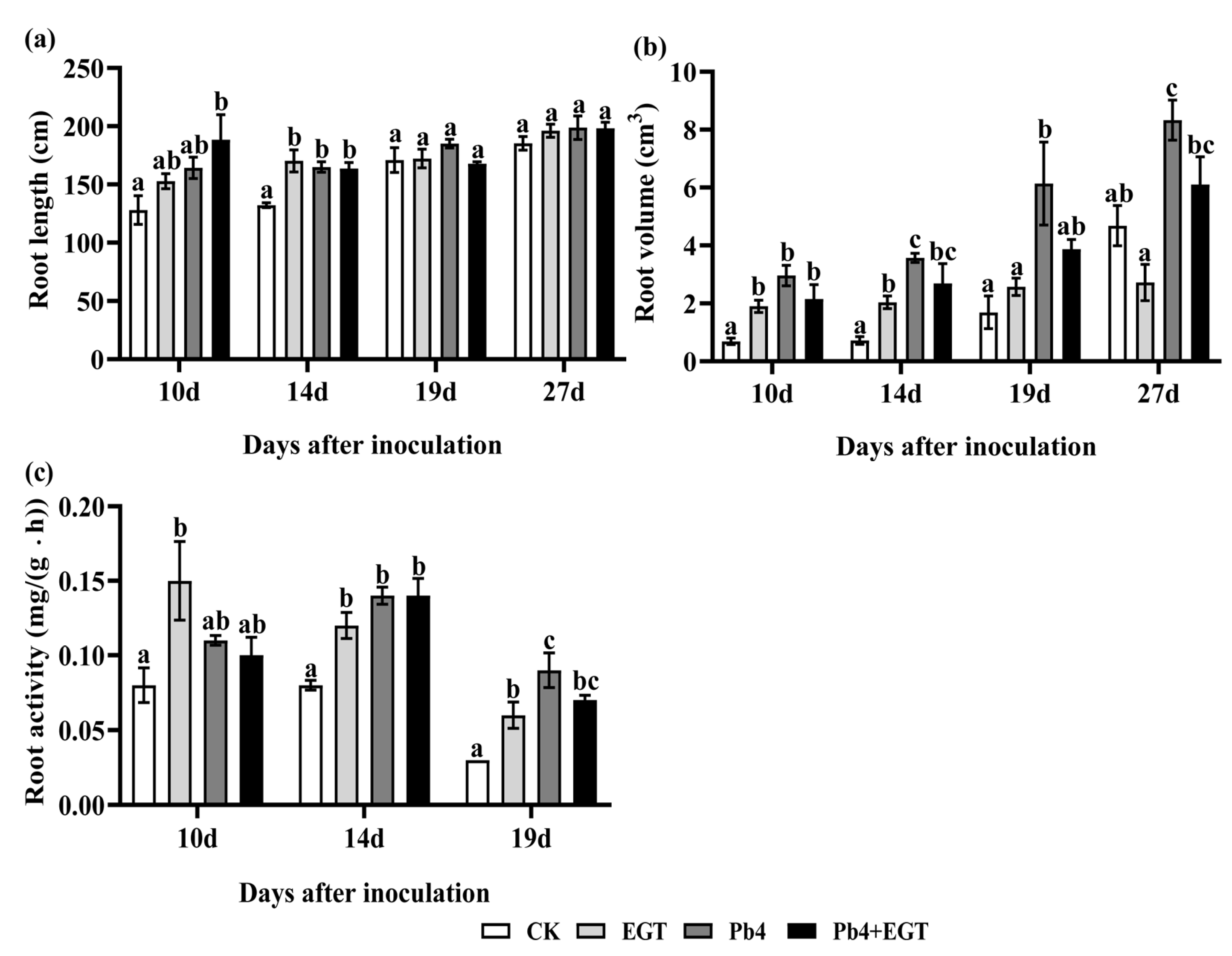
2.2. Effect of EGT on Chinese Cabbage Infected with P. brassicae
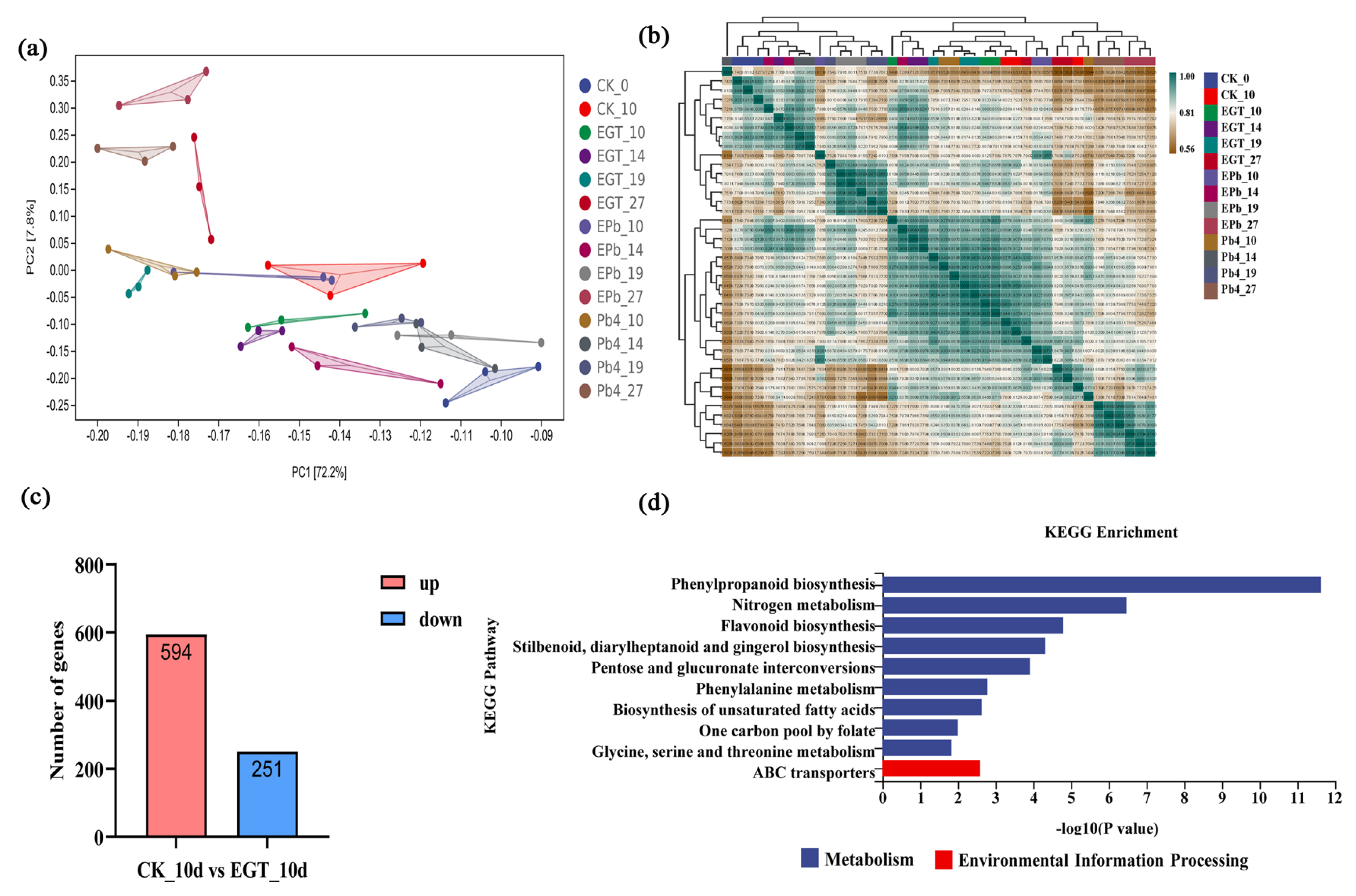
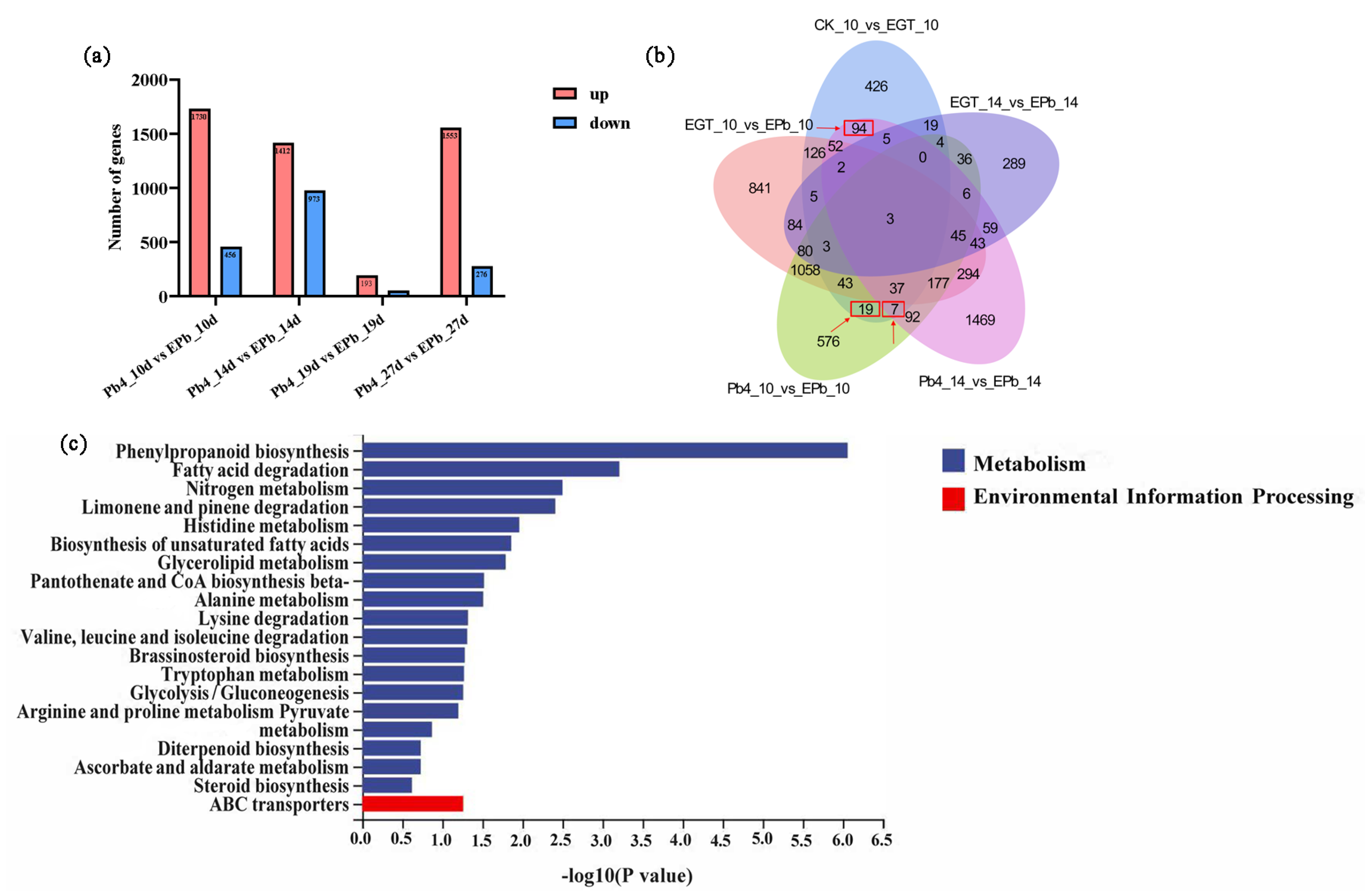
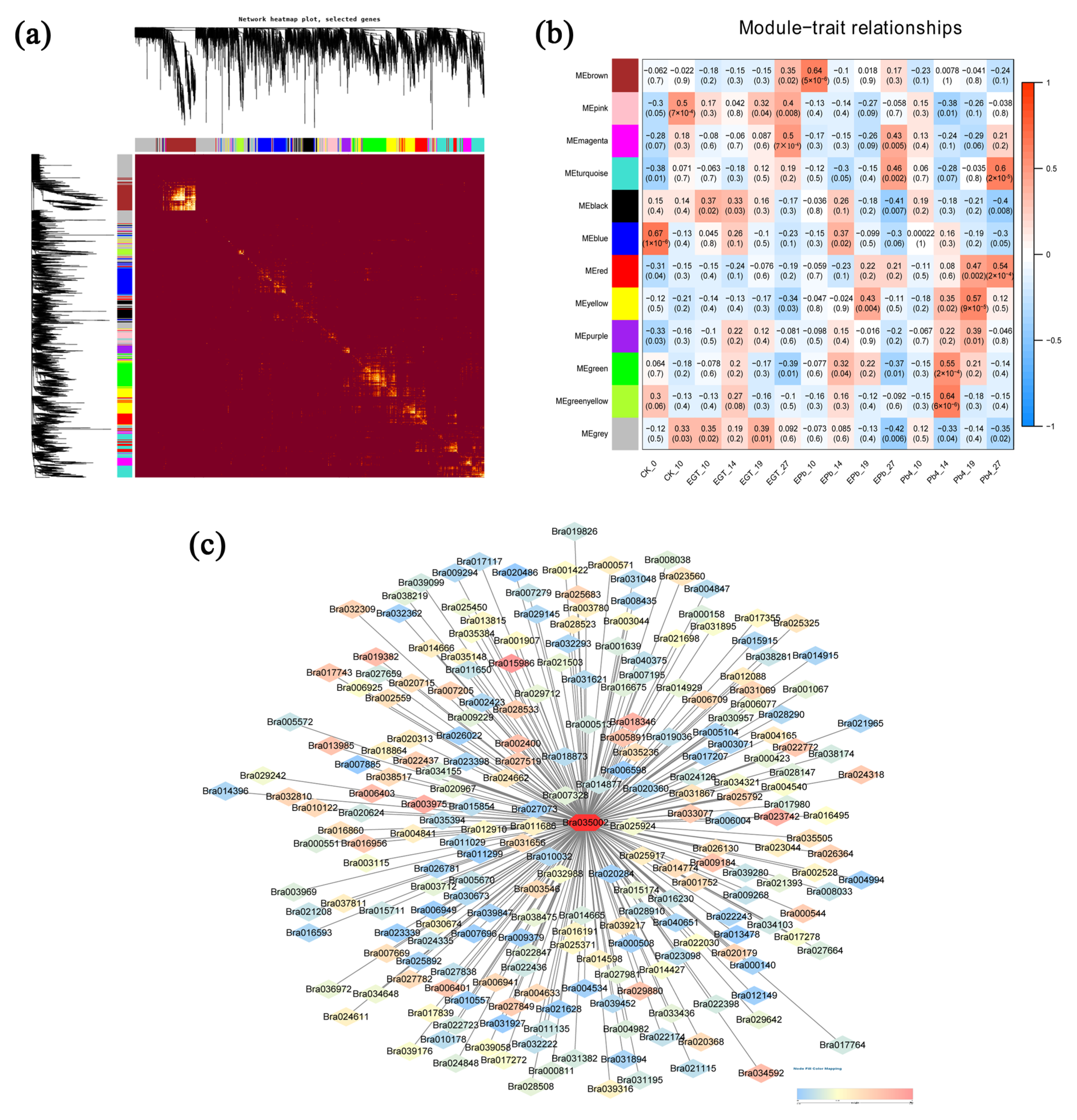
2.3. DEG Analysis of Exogenous EGT Treatment on Chinese Cabbage Root Development
2.4. DEG Analysis of Chinese Cabbage in Response to P. brassicae Infection
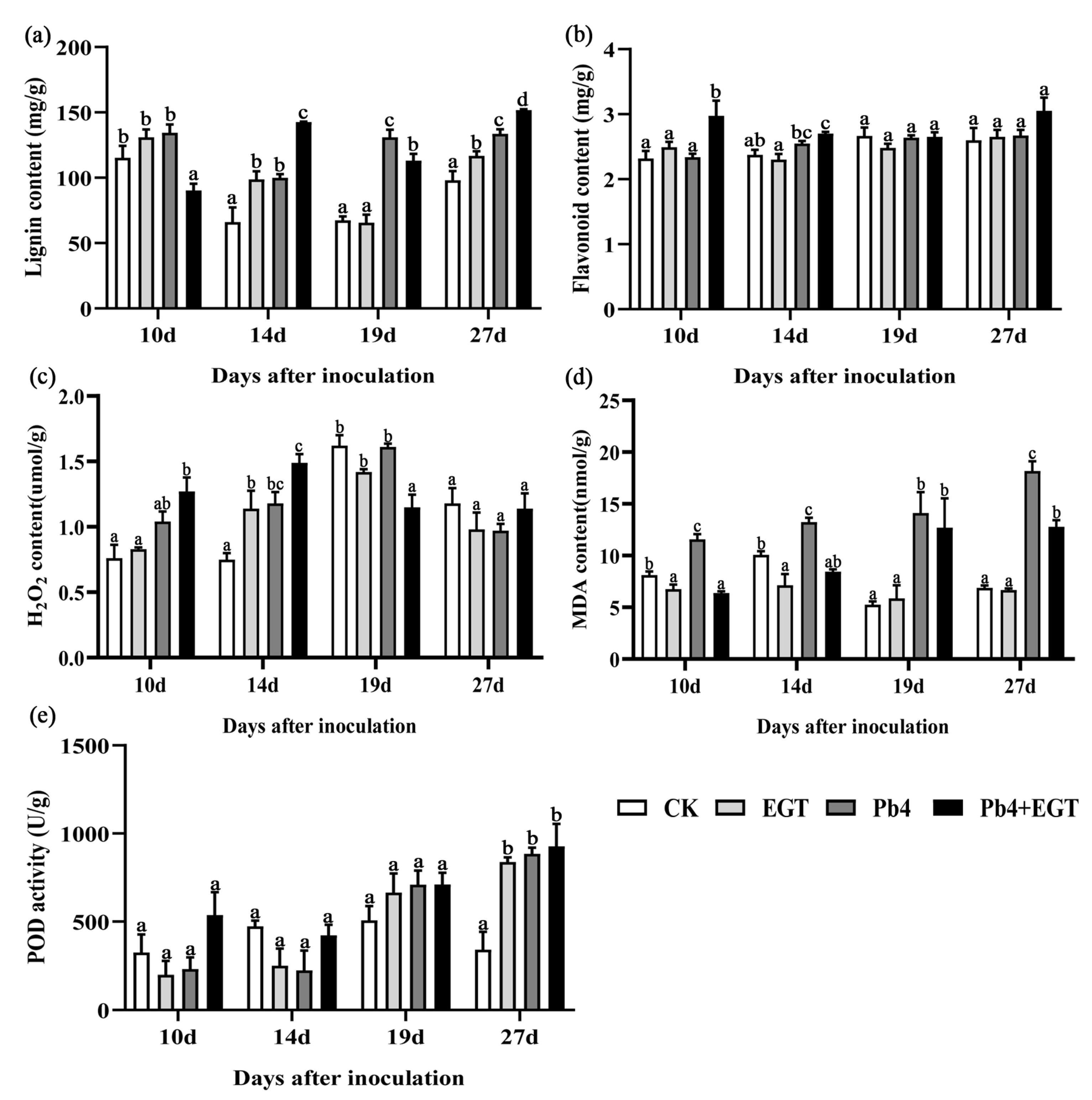
2.5. Effect of EGT on Lignin and Flavonoid Content
2.6. Effect of EGT on the H2O2, MDA, and POD Content in the Roots
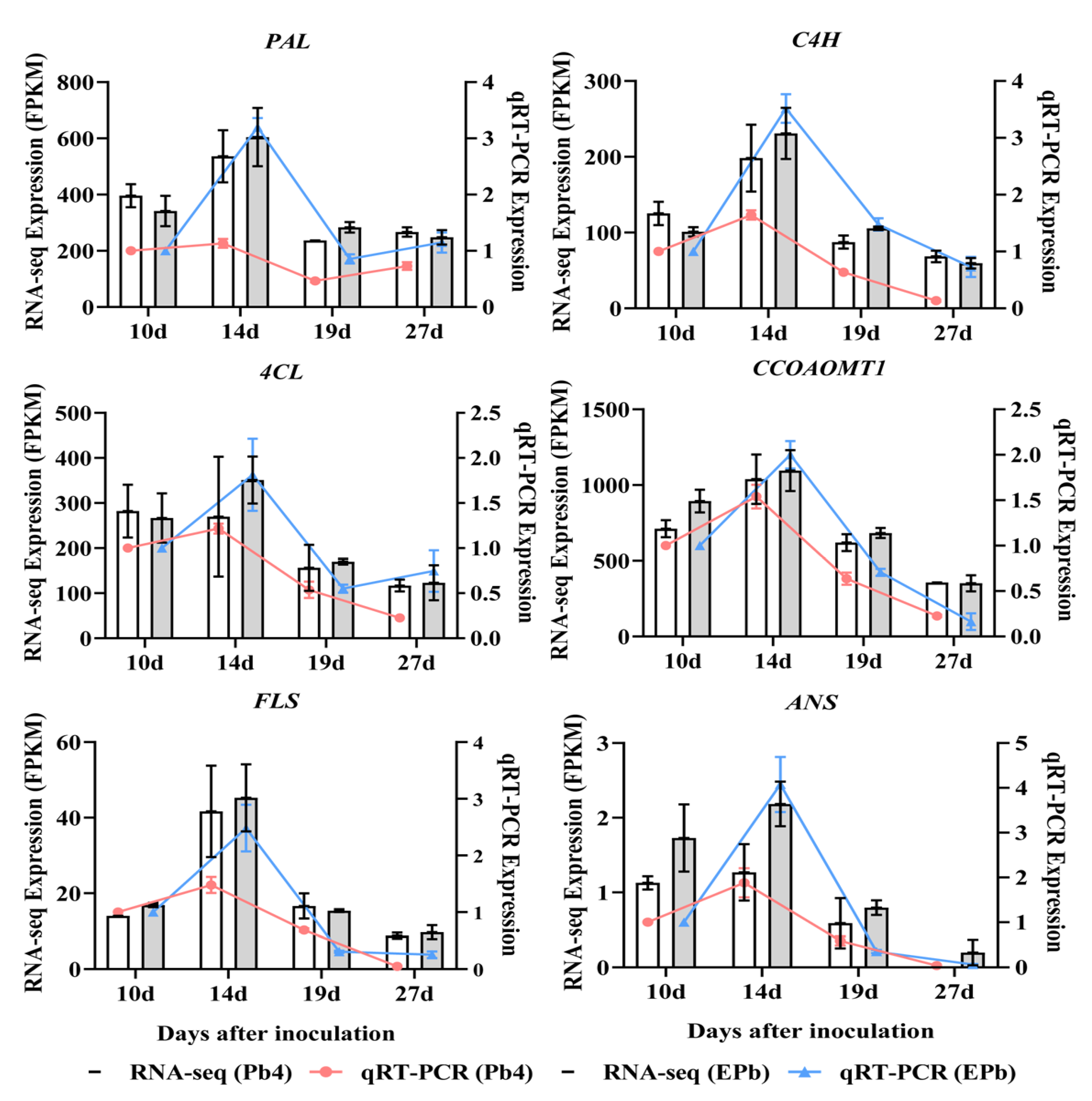
2.7. Quantitative RT-PCR Verification of the Phenylpropanoid Pathway Genes
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Pathogen Inoculation
4.2. Effects of Different EGT Concentrations on the Resting Spores of P. brassicae
4.3. Exogenous EGT Treatment
4.4. Disease Assessment
4.5. Detection of Root Physiological Indices and Activity
4.6. RNA Extraction and Transcriptome Sequencing
4.7. Identification of Differentially Expressed Genes and Validation of RNA Sequencing by qRT-PCR
4.8. Construction of Gene Co-Expression Networks and Prediction of Key Genes
4.9. Determination of Lignin, Total Flavonoid, H2O2, MDA, and POD Content
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dixon, G.R. The occurrence and economic impact of Plasmodiophora brassicae and clubroot disease. J. Plant Growth Regul. 2009, 28, 194–202. [Google Scholar] [CrossRef]
- Voorrips, R.E.; Jongerius, M.C.; Kanne, H.J. Quantitative trait loci for clubroot resistance in Brassica oleracea. Brassica Legum. Genome Struct. Breed. 2003, 52, 87–91. [Google Scholar]
- Howard, R.J.; Strelkov, S.E.; Harding, M.W. Clubroot of cruciferous crops—New perspectives on an old disease. Can. J. Plant Pathol. 2010, 32, 43–57. [Google Scholar] [CrossRef]
- Underwood, W. The plant cell wall: A dynamic barrier against pathogen invasion. Front. Plant Sci. 2012, 3, 85. [Google Scholar] [CrossRef] [Green Version]
- Piao, Z.; Ramchiary, N.; Lim, Y.P. Genetics of Clubroot Resistance in Brassica Species. J. Plant Growth Regul. 2009, 28, 252–264. [Google Scholar] [CrossRef]
- Badstoeber, J.; Ciaghi, S.; Neuhauser, S. Dynamic cell wall modifications in brassicas during clubroot disease. bioRxiv 2020. [Google Scholar] [CrossRef] [Green Version]
- Buendgen, M.R.; Coors, J.G.; Grombacher, A.W.; Russell, W.A. European Corn Borer Resistance and Cell Wall Composition of Three Maize Populations. Crop Sci. 1990, 30, 505–510. [Google Scholar] [CrossRef]
- Bonello, P.; Blodgett, J.T. Pinus nigra–Sphaeropsis sapinea as a model pathosystem to investigate local and systemic effects of fungal infection of pines. Physiol. Mol. Plant Pathol. 2003, 63, 249–261. [Google Scholar] [CrossRef]
- Smith, A.H.; Gill, W.M.; Pinkard, E.A.; Mohammed, C.L. Anatomical and histochemical defence responses induced in juvenile leaves of Eucalyptus globulus and Eucalyptus nitens by Mycosphaerella infection. For. Pathol. 2007, 37, 361–373. [Google Scholar] [CrossRef]
- Miedes, E.; Vanholme, R.; Boerjan, W.; Molina, A. The role of the secondary cell wall in plant resistance to pathogens. Front. Plant Sci. 2014, 5, 358. [Google Scholar] [CrossRef] [Green Version]
- Irani, S.; Todd, C.D.; Wei, Y.; Bonham-Smith, P.C. Changes in phenylpropanoid pathway gene expression in roots and leaves of susceptible and resistant Brassica napus lines in response to Plasmodiophora brassicae inoculation. Physiol. Mol. Plant Pathol. 2019, 106, 196–203. [Google Scholar] [CrossRef]
- Zhao, Y.; Bi, K.; Gao, Z.; Chen, T.; Liu, H.; Xie, J.; Cheng, J.; Fu, Y.; Jiang, D. Transcriptome Analysis of Arabidopsis thaliana in Response to Plasmodiophora brassicae during Early Infection. Front. Microbiol. 2017, 8, 673. [Google Scholar] [CrossRef] [PubMed]
- He, Z.; Gao, Z.; Bi, K.; Zhao, Y. Metabolic changes of Arabidopsis thaliana infected with Plasmodiophora brassicae based on LC-MS analysis. J. Huazhong Agric. Univ. 2017, 36, 30–38. [Google Scholar]
- Päsold, S.; Siegel, I.; Seidel, C.; Ludwig-Müller, J. Flavonoid accumulation in Arabidopsis thaliana root galls caused by the obligate biotrophic pathogen Plasmodiophora brassicae. Mol. Plant Pathol. 2010, 11, 545–562. [Google Scholar] [CrossRef] [PubMed]
- Tanret, C. Sur une base nouvelle retiree du seigle ergote, l’ergothioneine. Rend. Acad. Sci. 1909, 149, 222–224. [Google Scholar]
- Akanmu, D.; Cecchini, R.; Aruoma, O.I.; Halliwell, B. The antioxidant action of ergothioneine. Arch. Biochem. Biophys. 1991, 288, 10–16. [Google Scholar] [CrossRef]
- Aruoma, O.I.; Whiteman, M.; England, T.G.; Halliwell, B. Antioxidant Action of Ergothioneine: Assessment of Its Ability to Scavenge Peroxynitrite. Biochem. Biophys. Res. Commun. 1997, 231, 389–391. [Google Scholar] [CrossRef]
- Franzoni, F.; Colognato, R.; Galetta, F.; Laurenza, I.; Barsotti, M.C.; Di Stefano, R.; Bocchetti, R.; Regoli, F.; Carpi, A.; Balbarini, A.; et al. An in vitro study on the free radical scavenging capacity of ergothioneine: Comparison with reduced glutathione, uric acid and trolox. Biomed. Pharmacother. 2006, 60, 453–457. [Google Scholar] [CrossRef]
- Cheah, I.K.; Halliwell, B. Ergothioneine; antioxidant potential, physiological function and role in disease. Biochim. Biophys. Acta 2012, 1822, 784–793. [Google Scholar] [CrossRef] [Green Version]
- Asahi, T.; Wu, X.; Shimoda, H.; Hisaka, S.; Harada, E.; Kanno, T.; Nakamura, Y.; Kato, Y.; Osawa, T. A mushroom-derived amino acid, ergothioneine, is a potential inhibitor of inflammation-related DNA halogenation. Biosci. Biotechnol. Biochem. 2016, 80, 313–317. [Google Scholar] [CrossRef]
- Kawano, H.; Murata, H.; Iriguchi, S.; Mayumi, T.; Hama, T. Studies on ergothioneine. XI. Inhibitory effect on lipid peroxide formation in mouse liver. Chem. Pharm. Bull. 1983, 31, 1682–1687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaudière, J.; Ferrari-Iliou, R. Intracellular antioxidants: From chemical to biochemical mechanisms. Food Chem. Toxicol. 1999, 37, 949–962. [Google Scholar] [CrossRef]
- Colognato, R.; Laurenza, I.; Fontana, I.; Coppede, F.; Siciliano, G.; Coecke, S.; Aruoma, O.; Benzi, L.; Migliore, L. Modulation of hydrogen peroxide-induced DNA damage, MAPKs activation and cell death in PC12 by ergothioneine. Clin. Nutr. 2006, 25, 135–145. [Google Scholar] [CrossRef] [PubMed]
- Paul, B.D.; Snyder, S.H. The unusual amino acid L-ergothioneine is a physiologic cytoprotectant. Cell Death Differ. 2009, 17, 1134–1140. [Google Scholar] [CrossRef] [Green Version]
- Spicer, S.S.; Wooley, J.G.; Kessler, V. Ergothioneine Depletion in Rabbit Erythrocytes and its Effect on Methemoglobin Formation and Reversion. Exp. Biol. Med. 1951, 77, 418–420. [Google Scholar] [CrossRef] [PubMed]
- Arduini, A.; Mancinelli, G.; Radatti, G.L.; Hochstein, P.; Cadenas, E. Possible mechanism of inhibition of nitrite-induced oxidation of oxyhemoglobin by ergothioneine and uric acid. Arch. Biochem. Biophys. 1992, 294, 398–402. [Google Scholar] [CrossRef]
- Hatano, T.; Saiki, S.; Okuzumi, A.; Mohney, R.P.; Hattori, N. Identification of novel biomarkers for Parkinson’s disease by metabolomic technologies. J. Neurol. Neurosurg. Psychiatry 2016, 87, 295–301. [Google Scholar] [CrossRef]
- Cheah, I.K.; Feng, L.; Tang, R.M.Y.; Lim, K.H.C.; Halliwell, B. Ergothioneine levels in an elderly population decrease with age and incidence of cognitive decline; a risk factor for neurodegeneration? Biochem. Biophys. Res. Commun. 2016, 6, 162–167. [Google Scholar] [CrossRef]
- Lai, Y.; Xue, J.; Liu, C.W.; Gao, B.; Chi, L.; Tu, P.; Lu, K.; Ru, H. Serum Metabolomics Identifies Altered Bioenergetics, Signaling Cascades in Parallel with Exposome Markers in Crohn’s Disease. Molecules 2019, 24, 449. [Google Scholar] [CrossRef] [Green Version]
- Kameda, M.; Teruya, T.; Yanagida, M.; Kondoh, H. Frailty markers comprise blood metabolites involved in antioxidation, cognition, and mobility. Proc. Natl. Acad. Sci. USA 2020, 117, 9483–9489. [Google Scholar] [CrossRef] [Green Version]
- Smith, E.; Ottosson, F.; Hellstrand, S.; Ericson, U.; Orho-Melander, M.; Fernandez, C.; Melander, O. Ergothioneine is associated with reduced mortality and decreased risk of cardiovascular disease. Heart 2020, 106, 691–697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winkels, R.M.; van Brakel, L.; van Baar, H.; Beelman, R.B.; van Duijnhoven, F.J.B.; Geijsen, A.; van Halteren, H.K.; Hansson, B.M.E.; Richie, J.P.; Sun, D.; et al. Are Ergothioneine Levels in Blood Associated with Chronic Peripheral Neuropathy in Colorectal Cancer Patients Who Underwent Chemotherapy? Nutr. Cancer 2020, 72, 451–459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Macfarlane, I. Germination of resting spores of Plasmodiophora brassicae. Trans. Br. Mycol. Soc. 1970, 55, 97–112. [Google Scholar] [CrossRef]
- Yang, D.; Hu, C.; Cheng, Q.; Jia, W.; Zhao, X. Plasmodiophora brassicae and Plant Roots: The Interaction and Its Influencing Factors. Chin. Agric. Sci. Bull. 2019, 35, 77–81. [Google Scholar]
- Alscher, R.G.; Donahue, J.L.; Cramer, C.L. Reactive oxygen species and antioxidants: Relationships in green cells. Physiol. Plant. 1997, 100, 224–233. [Google Scholar] [CrossRef]
- Mallick, N.; Mohn, F.H. Reactive oxygen species: Response of algal cells. J. Plant Physiol. 2000, 157, 183–193. [Google Scholar] [CrossRef]
- Bowler, C.; Fluhr, R. The role of calcium and activated oxygens as signals for controlling cross-tolerance. Trends Plant Sci. 2000, 5, 241–246. [Google Scholar] [CrossRef]
- Hou, L. Effects of H2O2 on Some Physiological Indexes of Rape Seedlings under Chilling Stress. Hubei Agric. Sci. 2013, 52, 5144–5146. [Google Scholar]
- Thumma, B.R.; Nolan, M.F.; Evans, R.; Moran, G.F. Polymorphisms in Cinnamoyl CoA Reductase (CCR) are Associated with Variation in Microfibril Angle in Eucalyptus spp. Genetics 2005, 171, 1257–1265. [Google Scholar] [CrossRef] [Green Version]
- Piquemal, J.; Lapierre, C.; Myton, K.; O’Connell, A.; Schuch, W.; Grima-Pettenati, J.; Boudet, A.-M. Down-regulation of cinnamoyl-CoA reductase induces significant changes of lignin profiles in transgenic tobacco plants. Plant J. 1998, 13, 71–83. [Google Scholar] [CrossRef]
- Cao, T.; Srivastava, S.; Rahman, M.H.; Kav, N.N.; Hotte, N.; Deyholos, M.K.; Strelkov, S.E. Proteome-level changes in the roots of Brassica napus as a result of Plasmodiophora brassicae infection. Plant Sci. 2008, 174, 97–115. [Google Scholar] [CrossRef]
- Agarwal, A.; Kaul, V.; Faggian, R.; Rookes, J.E.; Ludwigmüller, J.; Cahill, D.M. Analysis of global host gene expression during the primary phase of the Arabidopsis thaliana–Plasmodiophora brassicae interaction. Funct. Plant Biol. 2011, 38, 462–478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Liu, Y.; Fang, Z.; Li, Z.; Yang, L.; Zhuang, M.; Zhang, Y.; Lv, H. Comparative Transcriptome Analysis between Broccoli (Brassica oleracea var. italica) and Wild Cabbage (Brassica macrocarpa Guss.) in Response to Plasmodiophora brassicae during Different Infection Stages. Front. Plant Sci. 2016, 7, 1929. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pang, W.; Liang, Y.; Zhan, Z.; Li, X.; Piao, Z. Development of a Sinitic Clubroot Differential Set for the Pathotype Classification of Plasmodiophora brassicae. Front. Plant Sci. 2020, 11, 568771. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Ma, S.; Huang, Y.; Wang, Z. Modality and biological characteristic of Plasmodiophora brassicae in rape. Sichuan Nongye Daxue Xuebao 2006, 24, 161. [Google Scholar]
- Naiki, T.; Dixon, G.; Ikegami, H. Quantitative estimation of spore germination of Plasmodiophora brassicae. Trans. Br. Mycol. Soc. 1987, 89, 569–572. [Google Scholar] [CrossRef]
- Chaube, H.S.; Singh, U.S. Plant Disease Management: Principles and Practices; CRC Press: Boca Raton, FL, USA, 1991. [Google Scholar]
- Siemens, J.; Nagel, M.; Ludwig-Muller, J.; Sacristán, M.D. The Interaction of Plasmodiophora brassicae and Arabidopsis thaliana Parameters for Disease Quantification and Screening of Mutant Lines. J. Phytopathol. 2002, 150, 592–605. [Google Scholar] [CrossRef]
- Mugai, E.N.; Agong, S.G.; Matsumoto, H. Aluminium tolerance mechanisms in Phaseolus vulgaris L.: Citrate synthase activity and TTC reduction are well correlated with citrate secretion. Soil Sci. Plant Nutr. 2000, 46, 939–950. [Google Scholar] [CrossRef] [Green Version]
- Clemensson-Lindell, A. Triphenyltetrazolium chloride as an indicator of fine-root vitality and environmental stress in coniferous forest stands: Applications and limitations. Plant Soil 1994, 159, 297–300. [Google Scholar] [CrossRef]
- Anders, S.; Huber, W. Differential expression analysis for sequence count data. Nat. Preced. 2010, 5, 1. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using RealTime Quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Wang, L.; Liu, B. Determination of Trace lignin in Flax Fiber by Turbidimetry. Chem. Adhes. 2004, 2004, 368–369. [Google Scholar]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Cao, G.; Li, X.; Piao, Z. Effects of Exogenous Ergothioneine on Brassica rapa Clubroot Development Revealed by Transcriptomic Analysis. Int. J. Mol. Sci. 2023, 24, 6380. https://doi.org/10.3390/ijms24076380
Zhang Y, Cao G, Li X, Piao Z. Effects of Exogenous Ergothioneine on Brassica rapa Clubroot Development Revealed by Transcriptomic Analysis. International Journal of Molecular Sciences. 2023; 24(7):6380. https://doi.org/10.3390/ijms24076380
Chicago/Turabian StyleZhang, Yuting, Guizhu Cao, Xiaonan Li, and Zhongyun Piao. 2023. "Effects of Exogenous Ergothioneine on Brassica rapa Clubroot Development Revealed by Transcriptomic Analysis" International Journal of Molecular Sciences 24, no. 7: 6380. https://doi.org/10.3390/ijms24076380