
Proanthocyanidin Structure-Activity Relationship Analysis by Path Analysis Model



Abstract
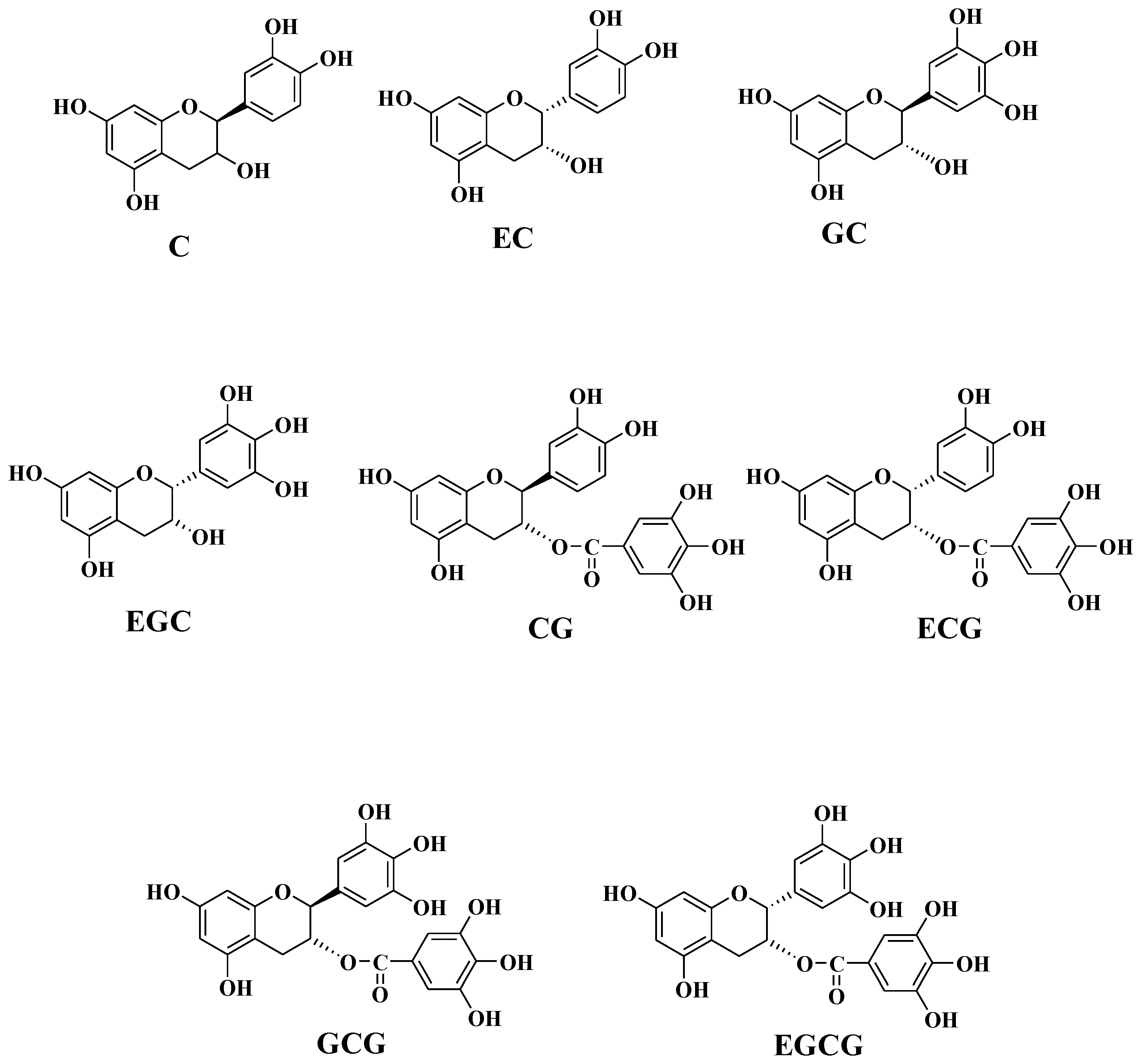
:1. Introduction
2. Results
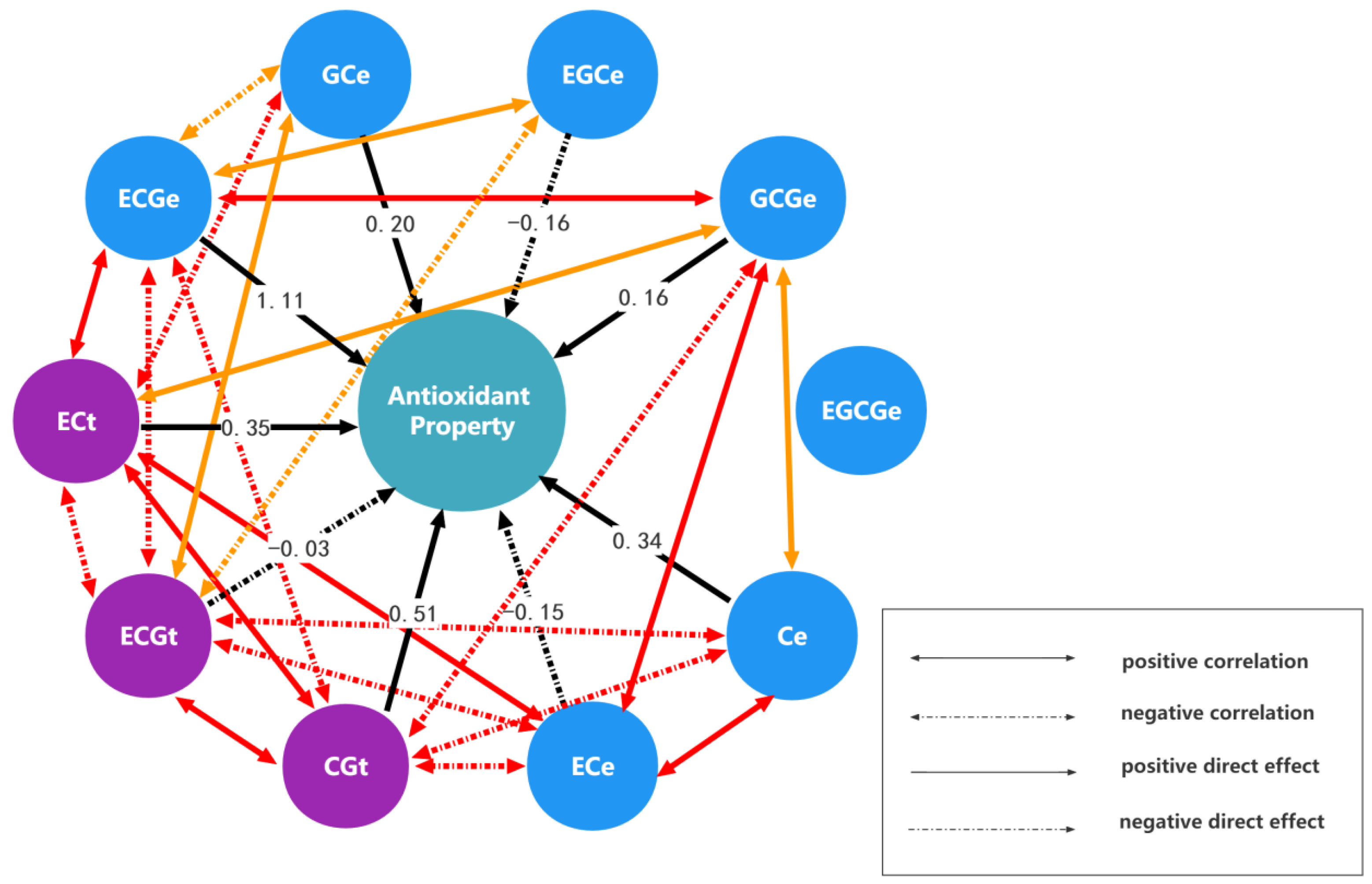
2.1. The Results of Monomer Units and Antioxidant Activities Analysis
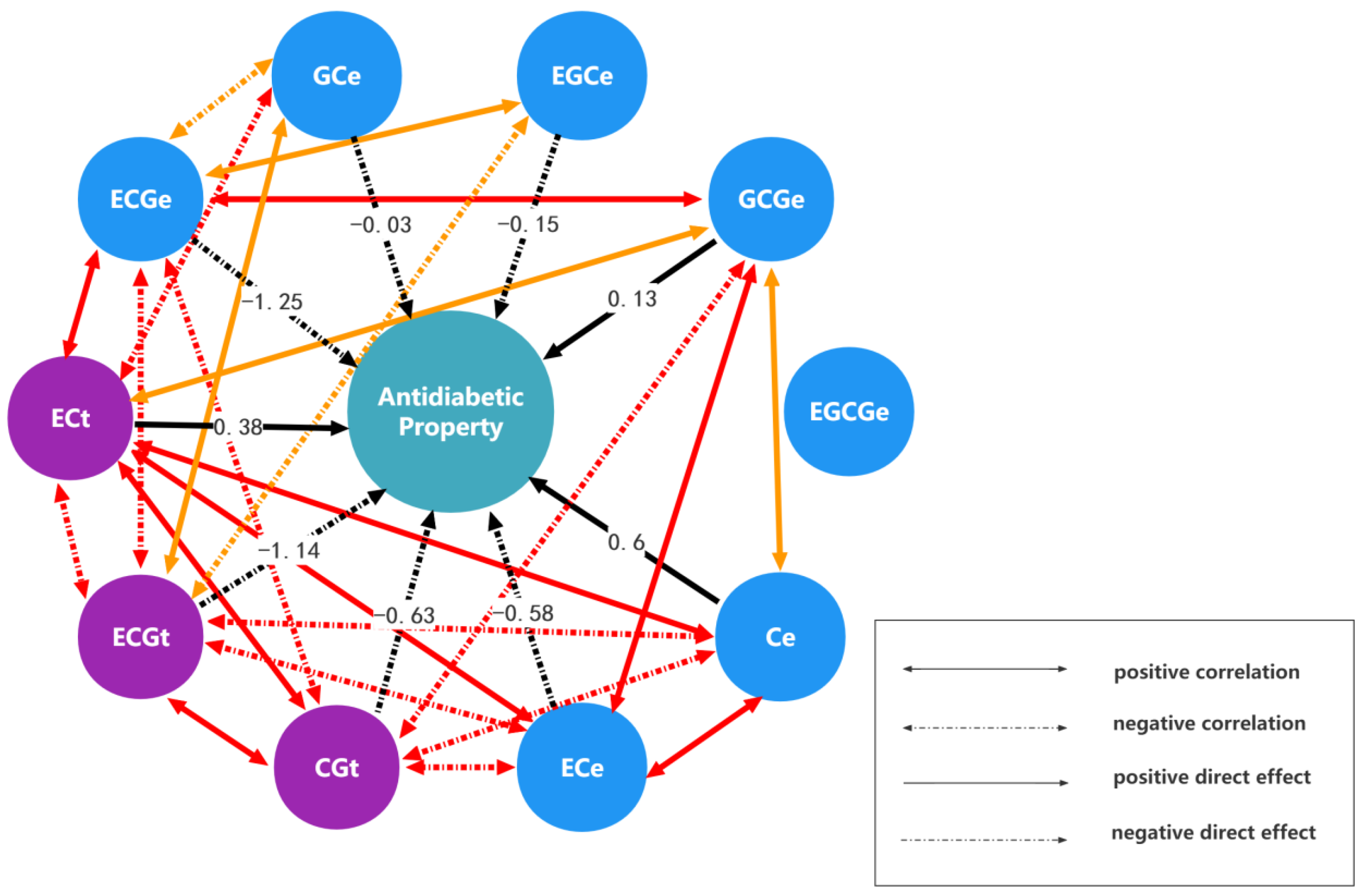
2.2. The Results of Monomer Units and Anti-Diabetes Bioactivities Analysis
2.3. The Comparison of Monomer Units on Antioxidant and Anti-Diabetes Activities Analysis Result
3. Discussion
4. Methods and Materials
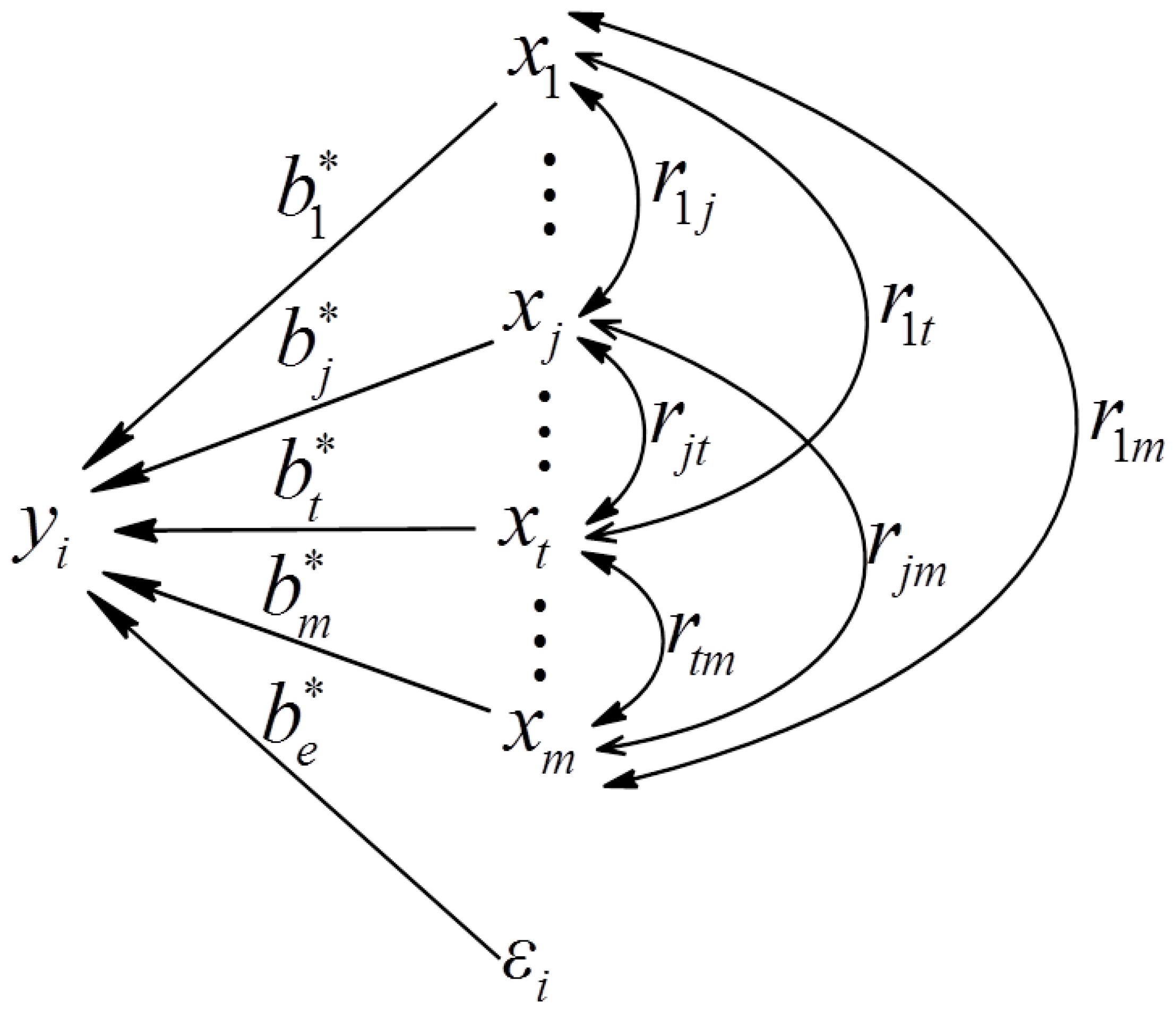
4.1. The Path Analysis Model for Structure-Activity Relation of PAs
4.2. Structural Analysis and Activity Determination
4.3. Materials
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviation
| C | Catechin |
| EC | Epicatechin |
| EGC | Epigallocatechin |
| ECG | Epicatechin gallate |
| EGCG | Epigallocatechin Gallate |
References
- Appeldoorn, M.M.; Sanders, M.; Vincken, J.P.; Cheynier, V.; le Guernevé, C.; Hollman, P.C.H.; Gruppen, H. Efficient isolation of major procyanidin A-type dimers from peanut skins and B-type dimers from grape seeds. Food Chem. 2009, 117, 713–720. [Google Scholar] [CrossRef]
- Hümmer, W.; Schreier, P. Analysis of proanthocyanidins. Mol. Nutr. Food Res. 2008, 52, 1381–1398. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Deng, J.; Joshi, S.; Liu, P.; Zhang, C.; Yu, Y.; Zhang, R.; Fan, D.; Yang, H.; D’Souza, D.H. Monomeric catechin and dimeric procyanidin B2 against human norovirus surrogates and their physicochemical interactions. Food Microbiol. 2018, 76, 346–353. [Google Scholar] [CrossRef]
- Li, M.N.; Wang, H.Y.; Wang, R.; Li, C.R.; Shen, B.Q.; Gao, W.; Li, P.; Yang, H. A modified data filtering strategy for targeted characterization of polymers in complex matrixes using drift tube ion mobility-mass spectrometry: Application to analysis of procyanidins in the grape seed extracts. Food Chem. 2020, 321, 126693. [Google Scholar] [CrossRef]
- Kiselova-Kaneva, Y.; Galunska, B.; Nikolova, M.; Dincheva, I.; Badjakov, I. High resolution LC-MS/MS characterization of polyphenolic composition and evaluation of antioxidant activity of Sambucus ebulus fruit tea traditionally used in Bulgaria as a functional food. Food Chem. 2022, 367, 130759. [Google Scholar] [CrossRef] [PubMed]
- Park, M.; Cho, H.; Jung, H.; Lee, H.; Hwang, K.T. Antioxidant and Anti-Inflammatory Activities of Tannin Fraction of the Extract from Black Raspberry Seeds Compared to Grape Seeds. J. Food Biochem. 2014, 38, 259–270. [Google Scholar] [CrossRef]
- Suo, H.; Tian, R.; Xu, W.; Li, L.; Cui, Y.; Zhang, S.; Sun, B. Novel Catechin-Tiopronin Conjugates Derived from Grape Seed Proanthocyanidin Degradation: Process Optimization, High-Speed Counter-Current Chromatography Preparation, as Well as Antibacterial Activity. J. Agric. Food Chem. 2019, 67, 11508–11517. [Google Scholar] [CrossRef]
- di Meo, F.; Aversano, R.; Diretto, G.; Demurtas, O.C.; Villano, C.; Cozzolino, S.; Filosa, S.; Carputo, D.; Crispi, S. Anti-cancer activity of grape seed semi-polar extracts in human mesothelioma cell lines. J. Funct. Foods 2019, 61, 103515. [Google Scholar] [CrossRef]
- Shahat, A.A.; Cos, P.; de Bruyne, T.; Apers, S.; Hammouda, F.M.; Ismail, S.I.; Azzam, S.; Claeys, M.; Goovaerts, E.; Pieters, L.; et al. Antiviral and antioxidant activity of flavonoids and proanthocyanidins from Crataegus sinaica. Planta Med. 2002, 68, 539–541. [Google Scholar] [CrossRef]
- Jiao, J.; Wei, Y.; Chen, J.; Chen, X.; Zhang, Y. Anti-aging and redox state regulation effects of A-type proanthocyanidins-rich cranberry concentrate and its comparison with grape seed extract in mice. J. Funct. Foods 2017, 30, 63–73. [Google Scholar] [CrossRef]
- Cedó, L.; Castell-Auví, A.; Pallarès, V.; Blay, M.; Ardévol, A.; Arola, L.; Pinent, M. Grape seed procyanidin extract modulates proliferation and apoptosis of pancreatic beta-cells. Food Chem. 2013, 138, 524–530. [Google Scholar] [CrossRef] [PubMed]
- Ge, Z.Z.; Dong, X.Q.; Zhu, W.; Zhang, Y.; Li, C.M. Metabolites and Changes in Antioxidant Activity of A-Type and B-Type Proanthocyanidin Dimers after Incubation with Rat Intestinal Microbiota. J. Agric. Food Chem. 2015, 63, 8991–8998. [Google Scholar] [CrossRef] [PubMed]
- Joshi, S.S.; Howell, A.B.; D’Souza, D.H. Reduction of Enteric Viruses by Blueberry Juice and Blueberry Proanthocyanidins. Food Environ. Virol. 2016, 8, 235–243. [Google Scholar] [CrossRef]
- Zhou, P.; Zhang, L.; Li, W.; Zhang, S.; Luo, L.; Wang, J.; Sun, B. In vitro evaluation of the anti-digestion and antioxidant effects of grape seed procyanidins according to their degrees of polymerization. J. Funct. Foods 2018, 49, 85–95. [Google Scholar] [CrossRef]
- Alejo-Armijo, A.; Glibota, N.; Frías, M.P.; Altarejos, J.; Gálvez, A.; Salido, S.; Ortega-Morente, E. Synthesis and Evaluation of Antimicrobial and Antibiofilm Properties of A-Type Procyanidin Analogues against Resistant Bacteria in Food. J. Agric. Food Chem. 2018, 66, 2151–2158. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, D.; Qiao, J.; Ni, Y.; Liu, P.; Huang, D.; Huo, J. Structure, degree of polymerization, and starch hydrolase inhibition activities of bird cherry (Prunus padus) proanthocyanidins. Food Chem. 2022, 385, 132588. [Google Scholar] [CrossRef]
- Xu, Y.Q.; Gao, Y.; Granato, D. Effects of epigallocatechin gallate, epigallocatechin and epicatechin gallate on the chemical and cell-based antioxidant activity, sensory properties, and cytotoxicity of a catechin-free model beverage. Food Chem. 2021, 339, 128060. [Google Scholar] [CrossRef]
- Ou-Yang, C.; Chai, W.; Xu, X.; Song, S.; Wei, Q.; Huang, Q.; Zou, Z. Inhibitory potential of proanthocyanidins from the fruit pulp of Clausena lansium (Lour.) Skeels against α-glucosidase and non-enzymatic glycation: Activity and mechanism. Process Biochem. 2020, 91, 364–373. [Google Scholar] [CrossRef]
- Ahmadi, S.; Mehrabi, M.; Rezaei, S.; Mardafkan, N. Structure-activity relationship of the radical scavenging activities of some natural antioxidants based on the graph of atomic orbitals. J. Mol. Struct. 2019, 1191, 165–174. [Google Scholar] [CrossRef]
- Alov, P.; Tsakovska, I.; Pajeva, I. Send Orders for Reprints to reprints@benthamscience.ae Computational Studies of Free Radical-Scavenging Properties of Phenolic Compounds. Curr. Top. Med. Chem. 2015, 15, 85–104. [Google Scholar] [CrossRef] [Green Version]
- Bouarab-Chibane, L.; Forquet, V.; Lantéri, P.; Clément, Y.; Léonard-Akkari, L.; Oulahal, N.; Degraeve, P.; Bordes, C. Antibacterial properties of polyphenols: Characterization and QSAR (Quantitative structure-activity relationship) models. Front. Microbiol. 2019, 10, 829. [Google Scholar] [CrossRef] [Green Version]
- Halder, A.K.; Moura, A.S.; Cordeiro MN, D.S. Moving Average-Based Multitasking In Silico Classification Modeling: Where Do We Stand and What Is Next? Int. J. Mol. Sci. 2022, 23, 4937. [Google Scholar] [CrossRef] [PubMed]
- Helguera, A.M.; Cordeiro, M.N.D.; Pérez, M.Á.C.; Combes, R.D.; González, M.P. Quantitative structure carcinogenicity relationship for detecting structural alerts in nitroso-compounds. Species: Rat; Sex: Male; Route of administration: Water. Toxicol. Appl. Pharmacol. 2008, 231, 197–207. [Google Scholar] [CrossRef] [PubMed]
- Sonam, K.S.; Guleria, S. Synergistic Antioxidant Activity of Natural Products. Ann. Pharmacol. Pharm. 2017, 2, 1086. [Google Scholar]
- Slavova-Kazakova, A.; Janiak, M.A.; Sulewska, K.; Kancheva, V.D.; Karamać, M. Synergistic, additive, and antagonistic antioxidant effects in the mixtures of curcumin with (−)-epicatechin and with a green tea fraction containing (−)-epicatechin. Food Chem. 2021, 360, 129994. [Google Scholar] [CrossRef] [PubMed]
- Weremfo, A.; Adulley, F.; Dabie, K.; Abassah-Oppong, S.; Peprah-Yamoah, E. Optimization of ultrasound-assisted extraction of phenolic antioxidants from turkey berry (Solanum torvum Sw) fruits using response surface methodology. J. Appl. Res. Med. Aromat. Plants 2022, 30, 100387. [Google Scholar] [CrossRef]
- Du, J.; Yuan, Z.; Ma, Z.; Song, J.; Xie, X.; Chen, Y. KEGG-PATH: Kyoto encyclopedia of genes and genomes-based pathway analysis using a path analysis model. Mol. BioSystems 2014, 10, 2441–2447. [Google Scholar] [CrossRef]
- Chiang, H.M.; Chen, H.C.; Wu, C.S.; Wu, P.Y.; Wen, K.C. Rhodiola plants: Chemistry and biological activity. J. Food Drug Anal. 2015, 23, 359–369. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.W.; Kim, Y.S.; Dong, X.; Park, J.S.; Shin, W.; bin Kim, S.J.; Go, E.A.; Park, P.J. Anti-inflammatory effect of Rhodiola crenulata extracts through the down-regulation of MyD88 dependent pathway and induction of autophagy. J. Funct. Foods 2020, 64, 103703. [Google Scholar] [CrossRef]
- Ma, D.; Wang, L.; Jin, Y.; Gu, L.; Yin, G.; Wang, J.; Yu, X.A.; Huang, H.; Zhang, Z.; Wang, B.; et al. Chemical characteristics of Rhodiola Crenulata and its mechanism in acute mountain sickness using UHPLC-Q-TOF-MS/MS combined with network pharmacology analysis. J. Ethnopharmacol. 2022, 294, 115345. [Google Scholar] [CrossRef]
- Salah, N.; Miller, N.J.; Paganga, G.; Tijburg, L.; Paul Bolwell, G.; Riceevans, C. Polyphenolic Flavanols as Scavengers of Aqueous Phase Radicals and as Chain-Breaking Antioxidants. Arch. Biochem. Biophys. 1995, 322, 339–346. [Google Scholar] [CrossRef] [PubMed]
- Almajano, M.P.; Delgado, M.E.; Gordon, M.H. Albumin causes a synergistic increase in the antioxidant activity of green tea catechins in oil-in-water emulsions. Food Chem. 2007, 102, 1375–1382. [Google Scholar] [CrossRef]
- He, J.; Xu, L.; Yang, L.; Wang, X. Epigallocatechin gallate is the most effective catechin against antioxidant stress via hydrogen peroxide and radical scavenging activity. Med. Sci. Monit. 2018, 24, 8198–8206. [Google Scholar] [CrossRef] [PubMed]
- Johnston, K.; Sharp, P.; Clifford, M.; Morgan, L. Dietary polyphenols decrease glucose uptake by human intestinal Caco-2 cells. FEBS Lett. 2005, 579, 1653–1657. [Google Scholar] [CrossRef] [Green Version]
- Loureiro, G.; Martel, F. The effect of dietary polyphenols on intestinal absorption of glucose and fructose: Relation with obesity and type 2 diabetes. Food Rev. Int. 2019, 35, 390–406. [Google Scholar] [CrossRef]
- Kobayashi, Y.; Suzuki, M.; Satsu, H.; Arai, S.; Hara, Y.; Suzuki, K.; Miyamoto, Y.; Shimizu, M. Green tea polyphenols inhibit the sodium-dependent glucose transporter of intestinal epithelial cells by a competitive mechanism. J. Agric. Food Chem. 2000, 48, 5618–5623. [Google Scholar] [CrossRef]
- Li, X.; Liu, J.; Chang, Q.; Zhou, Z.; Han, R.; Liang, Z. Antioxidant and antidiabetic activity of proanthocyanidins from fagopyrum dibotrys. Molecules 2021, 26, 2417. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Monomeric Units | ECt | CGt | ECGt | GCe | EGCe | GCGe | EGCGe | Ce | ECe | ECGe |
|---|---|---|---|---|---|---|---|---|---|---|
| The direct effect and indirect effect | 0.3521 | −0.3137 | −0.3253 | −0.3121 | 0.1765 | 0.273 | −0.0702 | 0.3245 | 0.3251 | 0.31 |
| −0.4532 | 0.5087 | 0.4964 | 0.3476 | −0.4228 | −0.4744 | 0.233 | −0.4594 | −0.4497 | −0.4982 | |
| 0.0254 | −0.0268 | −0.0274 | −0.0221 | 0.0203 | 0.0239 | −0.0139 | 0.0237 | 0.0233 | 0.0273 | |
| −0.1795 | 0.1384 | 0.1629 | 0.2026 | −0.0483 | −0.0978 | 0.0522 | −0.1376 | −0.1391 | −0.1524 | |
| −0.0821 | 0.136 | 0.1211 | 0.039 | −0.1637 | −0.1392 | 0.0919 | −0.0995 | −0.0998 | −0.13 | |
| 0.1237 | −0.1488 | −0.1392 | −0.077 | 0.1358 | 0.1596 | −0.0576 | 0.1307 | 0.1386 | 0.1405 | |
| 0.0006 | −0.0014 | −0.0016 | −0.0008 | 0.0017 | 0.0011 | −0.0031 | 0.0006 | 0.0001 | 0.0018 | |
| 0.3142 | −0.3079 | −0.2941 | −0.2316 | 0.2072 | 0.2792 | −0.0633 | 0.3409 | 0.3079 | 0.2857 | |
| −0.1416 | 0.1356 | 0.1304 | 0.1053 | −0.0936 | −0.1332 | 0.0039 | −0.1386 | −0.1534 | −0.1247 | |
| 0.9787 | −1.0886 | −1.1053 | −0.8361 | 0.8832 | 0.9789 | −0.6388 | 0.9315 | 0.9036 | 1.1117 | |
| Sum of indirect effects | 0.5861 | −1.4772 | −0.9548 | −0.9876 | 0.8601 | 0.7115 | −0.4629 | 0.5759 | 1.01 | −0.1401 |
| Total effect | 0.9382 | −0.9685 | −0.9822 | −0.785 | 0.6964 | 0.8711 | −0.466 | 0.9168 | 0.8566 | 0.9716 |
| Monomeric Units | ECt | CGt | ECGt | GCe | EGCe | GCGe | EGCGe | Ce | ECe | ECGe |
|---|---|---|---|---|---|---|---|---|---|---|
| The direct effect and indirect effect | 0.3837 | −0.3418 | −0.3545 | −0.34 | 0.1923 | 0.2975 | −0.0765 | 0.3536 | 0.3542 | 0.3378 |
| 0.5586 | −0.627 | −0.6118 | −0.4284 | 0.5211 | 0.5847 | −0.2872 | 0.5662 | 0.5542 | 0.614 | |
| 1.05 | −1.109 | −1.1364 | −0.914 | 0.8405 | 0.9913 | −0.5774 | 0.9805 | 0.9661 | 1.1299 | |
| 0.0257 | −0.0198 | −0.0233 | −0.029 | 0.0069 | 0.014 | −0.0075 | 0.0197 | 0.0199 | 0.0218 | |
| −0.0759 | 0.1258 | 0.112 | 0.0361 | −0.1514 | −0.1288 | 0.085 | −0.092 | −0.0923 | −0.1203 | |
| 0.1012 | −0.1217 | −0.1139 | −0.063 | 0.111 | 0.1305 | −0.0471 | 0.1069 | 0.1134 | 0.1149 | |
| −0.0014 | 0.0032 | 0.0035 | 0.0018 | −0.0039 | −0.0025 | 0.007 | −0.0013 | −0.0002 | −0.004 | |
| 0.5587 | −0.5475 | −0.5231 | −0.4118 | 0.3685 | 0.4966 | −0.1125 | 0.6063 | 0.5477 | 0.508 | |
| −0.5354 | 0.5126 | 0.493 | 0.3982 | −0.3536 | −0.5036 | 0.0146 | −0.5238 | −0.5799 | −0.4713 | |
| −1.1012 | 1.2249 | 1.2436 | 0.9407 | −0.9937 | −1.1014 | 0.7188 | −1.0481 | −1.0167 | −1.2508 | |
| Sum of indirect effects | 0.5803 | −0.2734 | 0.2255 | −0.7804 | 0.6891 | 0.6477 | −0.2898 | 0.3617 | 1.4463 | 2.1308 |
| Total effect | 0.964 | −0.9004 | −0.9109 | −0.8094 | 0.5377 | 0.7782 | −0.2828 | 0.968 | 0.8664 | 0.88 |

| Monomeric Units | |||||||
|---|---|---|---|---|---|---|---|
| The direct effect and the indirect effect | |||||||
| Sum of indirect effects | |||||||
| Total effect |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Z.; Liu, J.; You, J.; Li, X.; Liang, Z.; Du, J. Proanthocyanidin Structure-Activity Relationship Analysis by Path Analysis Model. Int. J. Mol. Sci. 2023, 24, 6379. https://doi.org/10.3390/ijms24076379
Li Z, Liu J, You J, Li X, Liang Z, Du J. Proanthocyanidin Structure-Activity Relationship Analysis by Path Analysis Model. International Journal of Molecular Sciences. 2023; 24(7):6379. https://doi.org/10.3390/ijms24076379
Chicago/Turabian StyleLi, Zhaoxuan, Jingling Liu, Jie You, Xin Li, Zongsuo Liang, and Junli Du. 2023. "Proanthocyanidin Structure-Activity Relationship Analysis by Path Analysis Model" International Journal of Molecular Sciences 24, no. 7: 6379. https://doi.org/10.3390/ijms24076379