
Voluntary Exercise to Reduce Anxiety Behaviour in Traumatic Brain Injury Shown to Alleviate Inflammatory Brain Response in Mice
 ,
,
Abstract
:1. Introduction
2. Results
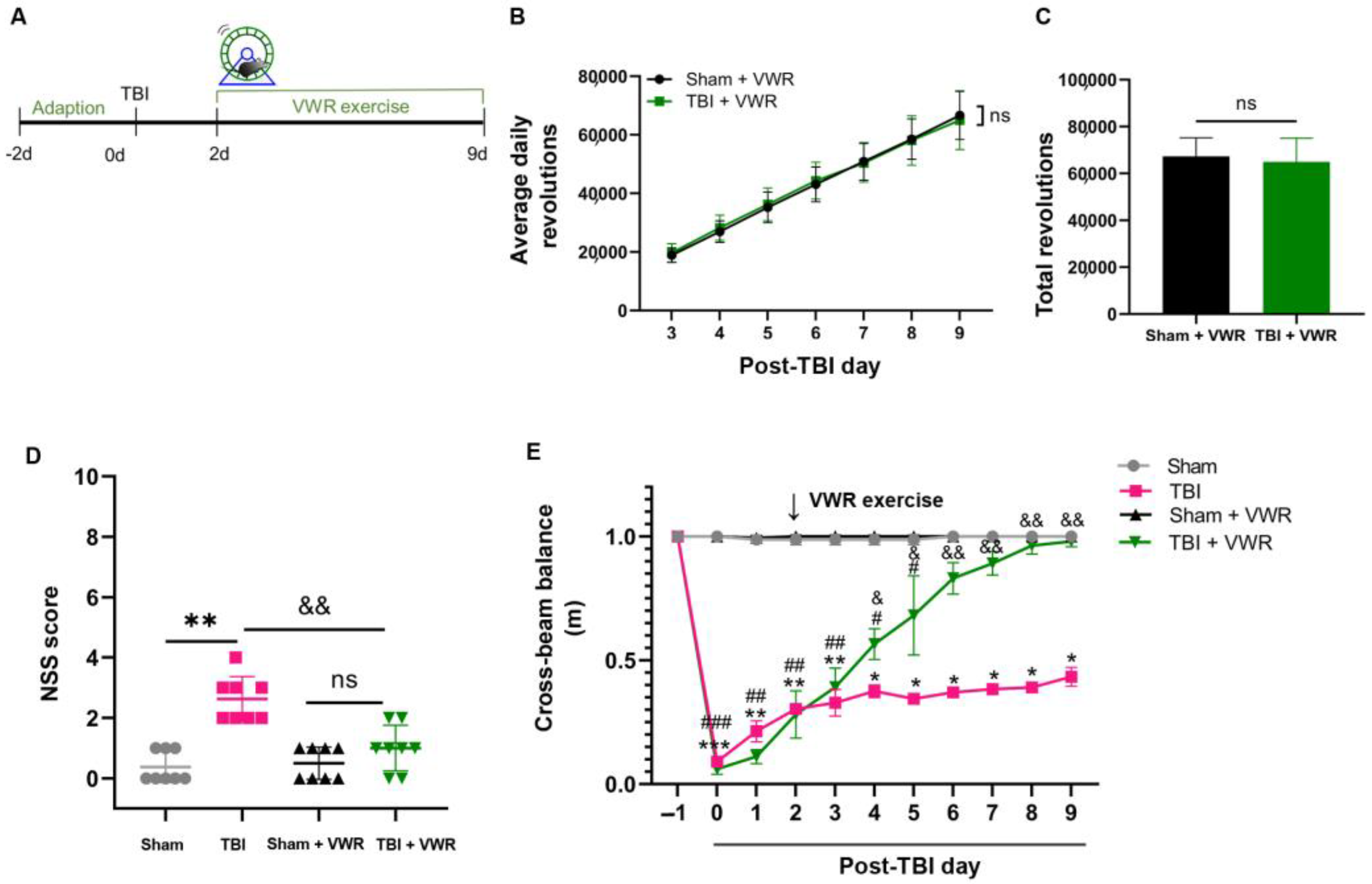
2.1. VWR Improved the Neurological Recovery of TBI Mice
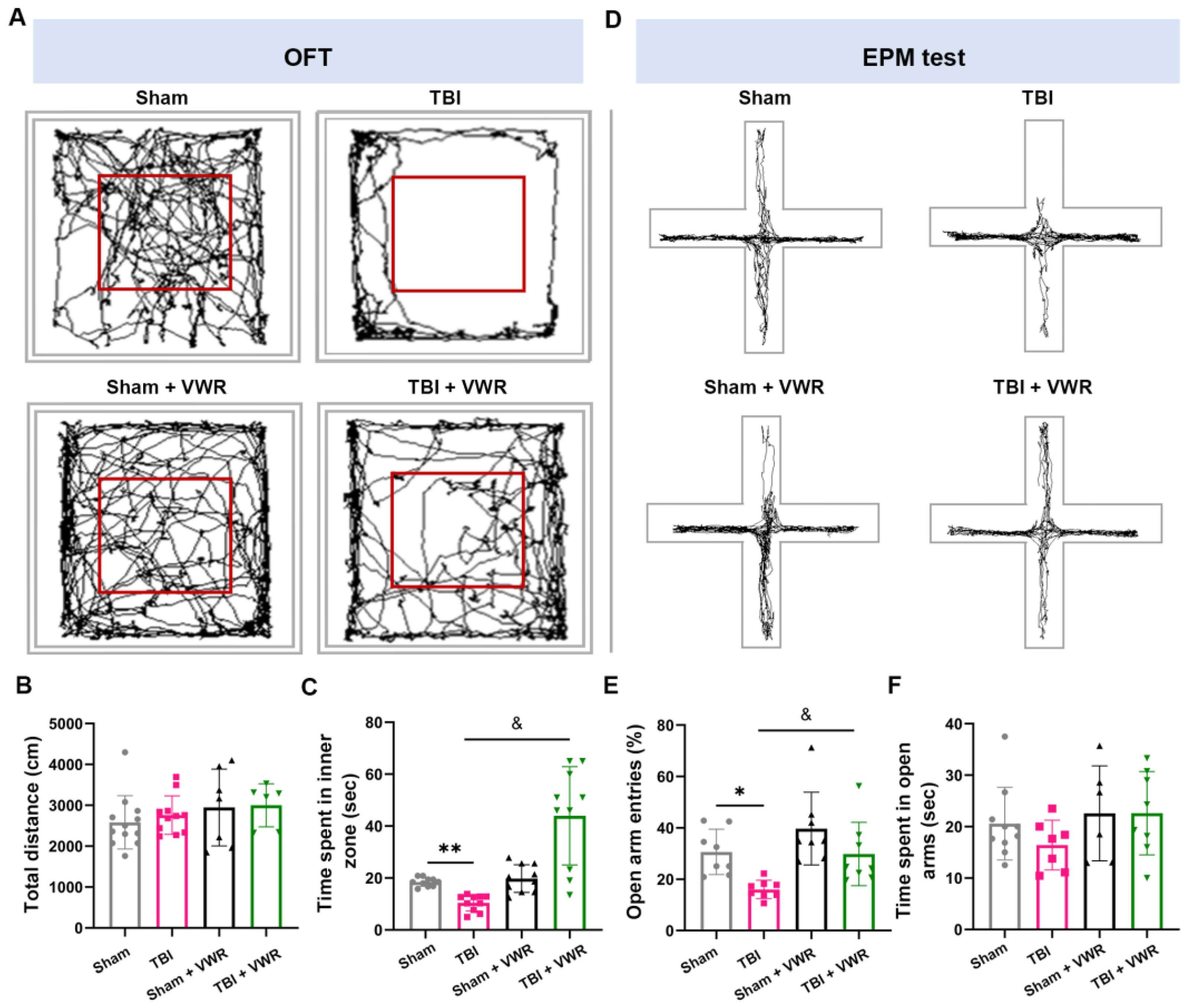
2.2. VWR Eased Anxiety in TBI Mice
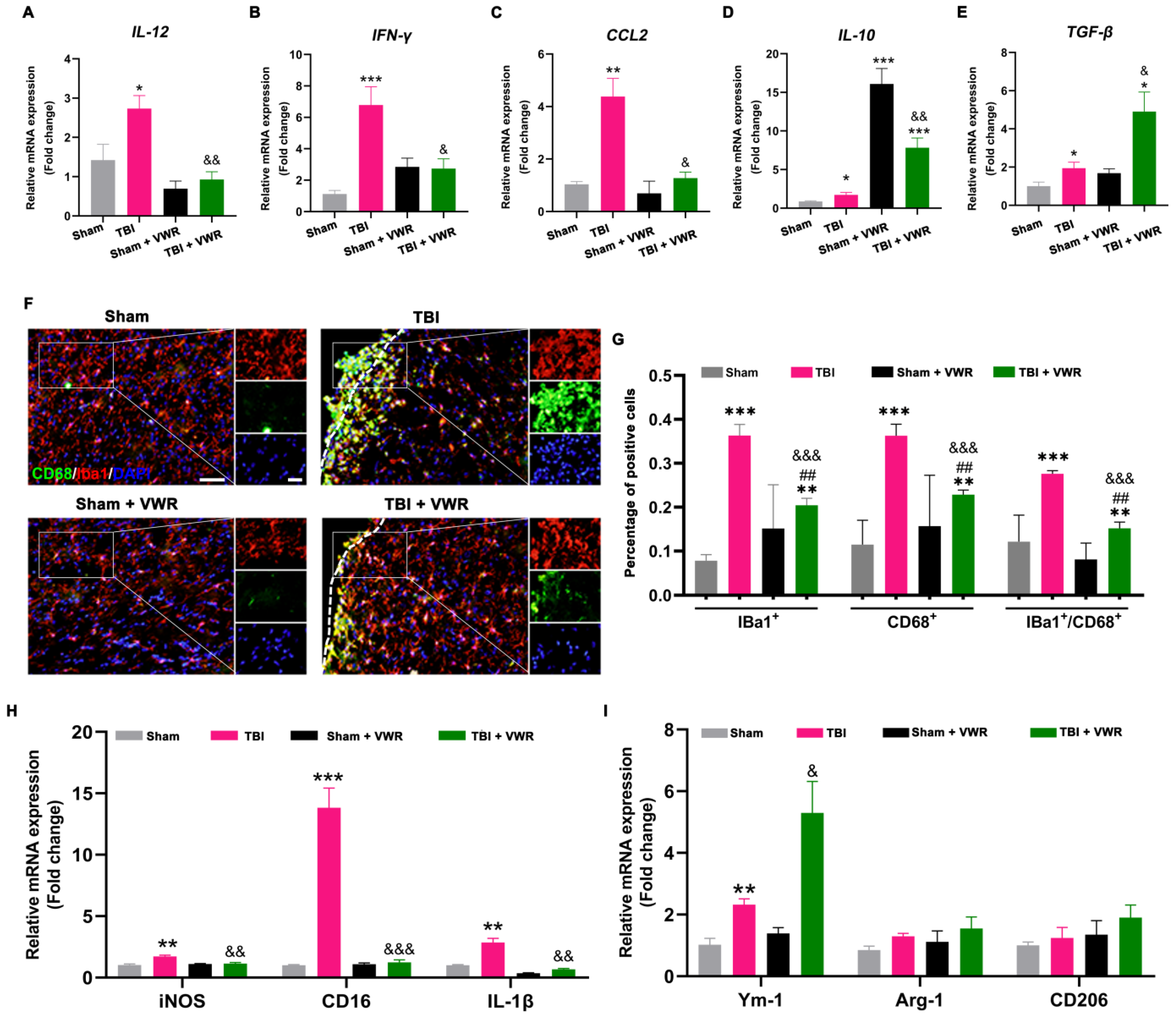
2.3. VWR Inhibited the Neuroinflammation Response after TBI
2.4. VWR Attenuated the Activation of Microglia and Promoted M2 Phenotypic Polarization after TBI
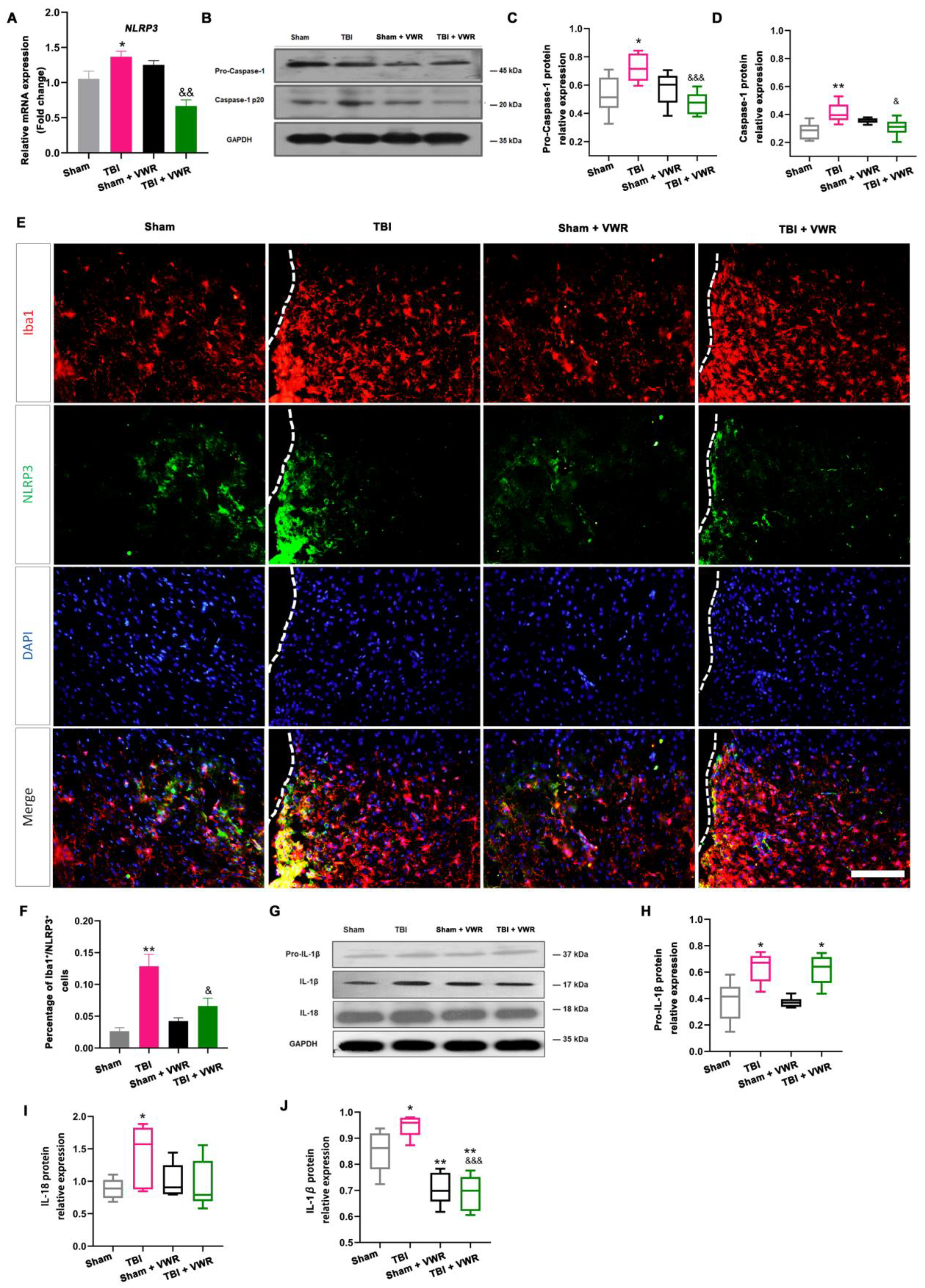
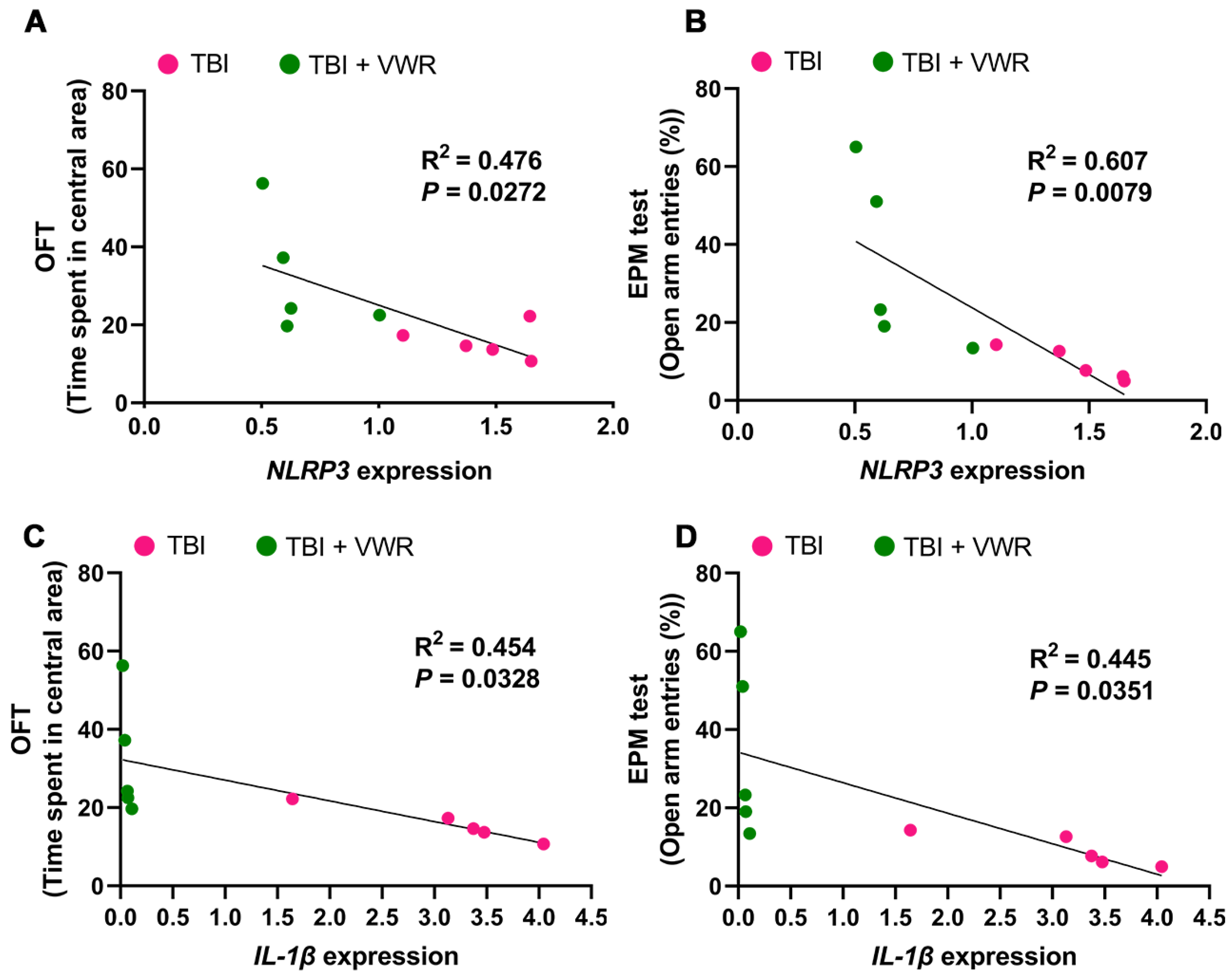
2.5. VWR Eased Anxiety Behavior in Mice via the Inhibition of NLRP3 Activation
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Construction of a Mouse Model of TBI
4.3. Voluntary Exercise Strategy
4.4. NSS Test
4.5. Motor Coordination and Balance Test
4.6. Behavioral Tests for Anxiety
4.6.1. OFT
4.6.2. EPM Test
4.7. Histology and Immunofluorescence Staining
4.8. Western Blotting Assay
4.9. qRT-PCR Analysis
4.10. Data Collection and Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liu, B. Current status and development of traumatic brain injury treatments in China. Chin. J. Traumatol. 2015, 18, 135–136. [Google Scholar] [CrossRef]
- Gardner, A.J.; Zafonte, R. Neuroepidemiology of traumatic brain injury. Handb. Clin. Neurol. 2016, 138, 207–223. [Google Scholar] [CrossRef] [PubMed]
- GBD 2016 Neurology Collaborators. Global, regional, and national burden of traumatic brain injury and spinal cord in-jury, 1990–2016: A systematic analysis for the Global Burden of Disease Study 2016. Lancet Neurol. 2019, 18, 56–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, J.Y.; Gao, G.Y.; Feng, J.F.; Mao, Q.; Chen, L.G.; Yang, X.F.; Liu, J.F.; Wang, Y.H.; Qiu, B.H.; Huang, X.J. Traumatic brain injury in China. Lancet Neurol. 2019, 18, 286–295. [Google Scholar] [CrossRef]
- Ahmed, S.; Venigalla, H.; Mekala, H.M.; Dar, S.; Hassan, M.; Ayub, S. Traumatic brain injury and neuropsychiatric complications. Indian J. Psychol. Med. 2017, 39, 114–121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braga, M.F.M.; Juranek, J.; Eiden, L.E.; Li, Z.; Figueiredo, T.H.; de Araujo Furtado, M.; Marini, A.M. GABAergic circuits of the basolateral amygdala and generation of anxiety after traumatic brain injury. Amino Acids 2022, 54, 1229–1249. [Google Scholar] [CrossRef]
- Osborn, A.J.; Mathias, J.L.; Fairweather-Schmidt, A.K. Prevalence of anxiety following adult traumatic brain injury: A meta-analysis comparing measures, samples and postinjury intervals. Neuropsychology 2016, 30, 247–261. [Google Scholar] [CrossRef]
- Albrecht, J.S.; Peters, M.E.; Smith, G.S.; Rao, V. Anxiety and posttraumatic stress disorder among medicare beneficiaries after traumatic brain injury. J. Head Trauma. Rehabil. 2017, 32, 178–184. [Google Scholar] [CrossRef]
- Popovitz, J.; Mysore, S.P.; Adwanikar, H. Long-term effects of traumatic brain injury on anxiety-like behaviors in mice: Behavioral and neural correlates. Front. Behav. Neurosci. 2019, 13, 6. [Google Scholar] [CrossRef] [Green Version]
- Tucker, L.B.; Burke, J.F.; Fu, A.H.; McCabe, J.T. Neuropsychiatric symptom modeling in male and female c57bl/6j mice after experimental traumatic brain injury. J. Neurotrauma 2017, 34, 890–905. [Google Scholar] [CrossRef] [Green Version]
- Scholten, A.C.; Haagsma, J.A.; Cnossen, M.C.; Olff, M.; van Beeck, E.F.; Polinder, S. Prevalence of and risk factors for anxiety and depressive disorders after traumatic brain injury: A systematic review. J. Neurotrauma 2016, 33, 1969–1994. [Google Scholar] [CrossRef] [PubMed]
- Diaz, A.P.; Schwarzbold, M.L.; Thais, M.E.; Hohl, A.; Bertotti, M.M.; Schmoeller, R.; Nunes, J.C.; Prediger, R.; Linhares, M.N.; Guarnieri, R.; et al. Psychiatric disorders and health-related quality of life after severe traumatic brain injury: A prospective study. J. Neurotrauma 2012, 29, 1029–1037. [Google Scholar] [CrossRef]
- Zheng, R.Z.; Lee, K.Y.; Qi, Z.X.; Wang, Z.; Xu, Z.Y.; Wu, X.H.; Mao, Y. Neuroinflammation following traumatic brain injury: Take it seriously or not. Front. Immunol. 2022, 13, 855701. [Google Scholar] [CrossRef]
- Jassam, Y.N.; Izzy, S.; Whalen, M.; McGavern, D.B.; El Khoury, J. Neuroimmunology of traumatic brain injury: Time for a paradigm shift. Neuron 2017, 95, 1246–1265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mira, R.G.; Lira, M.; Cerpa, W. Traumatic brain injury: Mechanisms of glial response. Front. Physiol. 2021, 12, 740939. [Google Scholar] [CrossRef]
- Karve, I.P.; Taylor, J.M.; Crack, P.J. The contribution of astrocytes and microglia to traumatic brain injury. Br. J. Pharmacol. 2016, 173, 692–702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.D.; Li, W.; Chen, Z.R.; Hu, Y.C.; Zhang, D.D.; Shen, W.; Zhou, M.L.; Zhu, L.; Hang, C.H. Expression of the NLRP3 inflammasome in cerebral cortex after traumatic brain injury in a rat model. Neurochem. Res. 2013, 38, 2072–2083. [Google Scholar] [CrossRef]
- Irrera, N.; Russo, M.; Pallio, G.; Bitto, A.; Mannino, F.; Minutoli, L.; Altavilla, D.; Squadrito, F. The role of NLRP3 inflammasome in the pathogenesis of traumatic brain injury. Int. J. Mol. Sci. 2020, 21, 6024. [Google Scholar] [CrossRef]
- Wei, X.; Hu, C.C.; Zhang, Y.L.; Yao, S.L.; Mao, W.K. Telmisartan reduced cerebral edema by inhibiting NLRP3 inflammasome in mice with cold brain injury. J. Huazhong Univ. Sci. Technol. Med. Sci. 2016, 36, 576–583. [Google Scholar] [CrossRef]
- Ma, J.; Xiao, W.; Wang, J.; Wu, J.; Ren, J.; Hou, J.; Gu, J.; Fan, K.; Yu, B. Propofol inhibits NLRP3 inflammasome and attenuates blast-induced traumatic brain injury in rats. Inflammation 2016, 39, 2094–2103. [Google Scholar] [CrossRef]
- O’Brien, W.T.; Pham, L.; Symons, G.F.; Monif, M.; Shultz, S.R.; McDonald, S.J. The NLRP3 inflammasome in traumatic brain injury: Potential as a biomarker and therapeutic target. J. Neuroinflamm. 2020, 17, 104. [Google Scholar] [CrossRef]
- Rashno, M.; Ghaderi, S.; Nesari, A.; Khorsandi, L.; Farbood, Y.; Sarkaki, A. Chrysin attenuates traumatic brain injury-induced recognition memory decline, and anxiety/depression-like behaviors in rats: Insights into underlying mechanisms. Psychopharmacology 2020, 237, 1607–1619. [Google Scholar] [CrossRef]
- Zhao, J.; Qu, D.; Xi, Z.; Huan, Y.; Zhang, K.; Yu, C.; Yang, D.; Kang, J.; Lin, W.; Wu, S.; et al. Mitochondria transplantation protects traumatic brain injury via promoting neuronal survival and astrocytic BDNF. Transl. Res. 2021, 235, 102–114. [Google Scholar] [CrossRef]
- Mele, C.; Pingue, V.; Caputo, M.; Zavattaro, M.; Pagano, L.; Prodam, F.; Nardone, A.; Aimaretti, G.; Marzullo, P. Neuroinflammation and hypothalamo-pituitary dysfunction: Focus of traumatic brain injury. Int. J. Mol. Sci. 2021, 22, 2686. [Google Scholar] [CrossRef]
- Rodgers, K.M.; Bercum, F.M.; McCallum, D.L.; Rudy, J.W.; Frey, L.C.; Johnson, K.W.; Watkins, L.R.; Barth, D.S. Acute neuroimmune modulation attenuates the development of anxiety-like freezing behavior in an animal model of traumatic brain injury. J. Neurotrauma 2012, 29, 1886–1897. [Google Scholar] [CrossRef] [Green Version]
- Rodgers, K.M.; Deming, Y.K.; Bercum, F.M.; Chumachenko, S.Y.; Wieseler, J.L.; Johnson, K.W.; Watkins, L.R.; Barth, D.S. Reversal of established traumatic brain injury-induced, anxiety-like behavior in rats after delayed, post-injury neuroimmune suppression. J. Neurotrauma 2014, 31, 487–497. [Google Scholar] [CrossRef] [Green Version]
- Yang, F.; Wang, Z.; Wei, X.; Han, H.; Meng, X.; Zhang, Y.; Shi, W.; Li, F.; Xin, T.; Pang, Q.; et al. NLRP3 deficiency ameliorates neurovascular damage in experimental ischemic stroke. J. Cereb. Blood Flow. Metab. 2014, 34, 660–667. [Google Scholar] [CrossRef] [Green Version]
- Irrera, N.; Pizzino, G.; Calo, M.; Pallio, G.; Mannino, F.; Fama, F.; Arcoraci, V.; Fodale, V.; David, A.; Francesca, C.; et al. Lack of the Nlrp3 inflammasome improves mice recovery following traumatic brain injury. Front. Pharmacol. 2017, 8, 459. [Google Scholar] [CrossRef] [Green Version]
- Zheng, B.; Zhang, S.; Ying, Y.; Guo, X.; Li, H.; Xu, L.; Ruan, X. Administration of dexmedetomidine inhibited NLRP3 inflammasome and microglial cell activities in hippocampus of traumatic brain injury rats. Biosci. Rep. 2018, 38, BSR20180892. [Google Scholar] [CrossRef] [Green Version]
- Kuwar, R.; Rolfe, A.; Di, L.; Xu, H.; He, L.; Jiang, Y.; Zhang, S.; Sun, D. A novel small molecular NLRP3 inflammasome inhibitor alleviates neuroinflammatory response following traumatic brain injury. J. Neuroinflamm. 2019, 16, 81. [Google Scholar] [CrossRef] [Green Version]
- Ismael, S.; Nasoohi, S.; Ishrat, T. MCC950, the selective inhibitor of nucleotide oligomerization domain-like receptor protein-3 inflammasome, protects mice against traumatic brain injury. J. Neurotrauma 2018, 35, 1294–1303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.; Nakagawa, S.; An, Y.; Ito, K.; Kitaichi, Y.; Kusumi, I. The exercise-glucocorticoid paradox: How exercise is beneficial to cognition, mood, and the brain while increasing glucocorticoid levels. Front. Neuroendocrinol. 2017, 44, 83–102. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Y.; Mahmood, A.; Chopp, M. Emerging treatments for traumatic brain injury. Expert Opin. Emerg. Drugs 2009, 14, 67–84. [Google Scholar] [CrossRef] [PubMed]
- Stephan, J.S.; Sleiman, S.F. Exercise factors as potential mediators of cognitive rehabilitation following traumatic brain injury. Curr. Opin. Neurol. 2019, 32, 808–814. [Google Scholar] [CrossRef] [PubMed]
- Kodali, M.; Mishra, V.; Hattiangady, B.; Attaluri, S.; Gonzalez, J.J.; Shuai, B.; Shetty, A.K. Moderate, intermittent voluntary exercise in a model of Gulf War Illness improves cognitive and mood function with alleviation of activated microglia and astrocytes, and enhanced neurogenesis in the hippocampus. Brain Behav. Immun. 2021, 97, 135–149. [Google Scholar] [CrossRef]
- Griesbach, G.S.; Tio, D.L.; Nair, S.; Hovda, D.A. Recovery of stress response coincides with responsiveness to voluntary exercise after traumatic brain injury. J. Neurotrauma 2014, 31, 674–682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amorós-Aguilar, L.; Portell-Cortés, I.; Costa-Miserachs, D.; Torras-Garcia, M.; Riubugent-Camps, È.; Almolda, B.; Coll-Andreu, M. The benefits of voluntary physical exercise after traumatic brain injury on rat’s object recognition memory: A comparison of different temporal schedules. Exp. Neurol. 2020, 326, 113178. [Google Scholar] [CrossRef]
- Griesbach, G.S.; Tio, D.L.; Vincelli, J.; McArthur, D.L.; Taylor, A.N. Differential effects of voluntary and forced exercise on stress responses after traumatic brain injury. J. Neurotrauma 2012, 29, 1426–1433. [Google Scholar] [CrossRef] [Green Version]
- Scrimgeour, A.G.; Condlin, M.L.; Loban, A.; DeMar, J.C. Omega-3 fatty acids and vitamin d decrease plasma t-tau, gfap, and uch-l1 in experimental traumatic brain injury. Front. Nutr. 2021, 8, 685220. [Google Scholar] [CrossRef]
- Zhao, Z.; Sabirzhanov, B.; Wu, J.; Faden, A.I.; Stoica, B.A. Voluntary exercise preconditioning activates multiple antiapoptotic mechanisms and improves neurological recovery after experimental traumatic brain injury. J. Neurotrauma 2015, 32, 1347–1360. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Drudis, L.; Amorós-Aguilar, L.; Torras-Garcia, M.; Serra-Elias, B.; Costa-Miserachs, D.; Portell-Cortés, I.; Coll-Andreu, M. Delayed voluntary physical exercise restores “when” and “where” object recognition memory after traumatic brain injury. Behav. Brain Res. 2021, 400, 113048. [Google Scholar] [CrossRef] [PubMed]
- Risbrough, V.B.; Vaughn, M.N.; Friend, S.F. Role of inflammation in traumatic brain injury-associated risk for neuropsychiatric disorders: State of the evidence and where do we go from here. Biol. Psychiatry 2022, 91, 438–448. [Google Scholar] [CrossRef] [PubMed]
- Osborn, A.J.; Mathias, J.L.; Fairweather-Schmidt, A.K.; Anstey, K.J. Anxiety and comorbid depression following traumatic brain injury in a community-based sample of young, middle-aged and older adults. J. Affect. Disord. 2017, 213, 214–221. [Google Scholar] [CrossRef] [PubMed]
- Yurgil, K.A.; Barkauskas, D.A.; Vasterling, J.J.; Nievergelt, C.M.; Larson, G.E.; Schork, N.J.; Litz, B.T.; Nash, W.P.; Baker, D.G. Association between traumatic brain injury and risk of posttraumatic stress disorder in active-duty Marines. JAMA Psychiatry 2014, 71, 149–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stein, M.B.; Kessler, R.C.; Heeringa, S.G.; Jain, S.; Campbell-Sills, L.; Colpe, L.J.; Fullerton, C.S.; Nock, M.K.; Sampson, N.A.; Schoenbaum, M.; et al. Prospective longitudinal evaluation of the effect of deployment-acquired traumatic brain injury on posttraumatic stress and related disorders: Results from the Army Study to Assess Risk and Resilience in Servicemembers (Army STARRS). Am. J. Psychiatry 2015, 172, 1101–1111. [Google Scholar] [CrossRef] [Green Version]
- Rodney, T.; Taylor, P.; Dunbar, K.; Perrin, N.; Lai, C.; Roy, M.; Gill, J. High IL-6 in military personnel relates to multiple traumatic brain injuries and post-traumatic stress disorder. Behav. Brain Res. 2020, 392, 112715. [Google Scholar] [CrossRef]
- Stein, M.B.; Jain, S.; Giacino, J.T.; Levin, H.; Dikmen, S.; Nelson, L.D.; Vassar, M.J.; Okonkwo, D.O.; Diaz-Arrastia, R.; Robertson, C.S.; et al. Risk of posttraumatic stress disorder and major depression in civilian patients after mild traumatic brain injury: A track-tbi study. JAMA Psychiatry 2019, 76, 249–258. [Google Scholar] [CrossRef]
- Devoto, C.; Arcurio, L.; Fetta, J.; Ley, M.; Rodney, T.; Kanefsky, R.; Gill, J. Inflammation relates to chronic behavioral and neurological symptoms in military personnel with traumatic brain injuries. Cell Transplant. 2017, 26, 1169–1177. [Google Scholar] [CrossRef]
- Chiu, C.C.; Liao, Y.E.; Yang, L.Y.; Wang, J.Y.; Tweedie, D.; Karnati, H.K.; Greig, N.H.; Wang, J.Y. Neuroinflammation in animal models of traumatic brain injury. J. Neurosci. Methods 2016, 272, 38–49. [Google Scholar] [CrossRef] [Green Version]
- Simon, D.W.; McGeachy, M.J.; Bayır, H.; Clark, R.S.; Loane, D.J.; Kochanek, P.M. The far-reaching scope of neuroinflammation after traumatic brain injury. Nat. Rev. Neurol. 2017, 13, 171–191. [Google Scholar] [CrossRef] [Green Version]
- Semple, B.D.; Bye, N.; Rancan, M.; Ziebell, J.M.; Morganti-Kossmann, M.C. Role of CCL2 (MCP-1) in traumatic brain injury (TBI): Evidence from severe TBI patients and CCL2−/− mice. J. Cereb. Blood Flow. Metab. 2010, 30, 769–782. [Google Scholar] [CrossRef]
- Flierl, M.A.; Stahel, P.F.; Beauchamp, K.M.; Morgan, S.J.; Smith, W.R.; Shohami, E. Mouse closed head injury model induced by a weight-drop device. Nat. Protoc. 2009, 4, 1328–1337. [Google Scholar] [CrossRef]
- Ge, Q.; Hu, X.; Ma, N.; Sun, M.; Zhang, L.; Cai, Z.; Tan, R.; Lu, H. Maternal high-salt diet during pregnancy impairs synaptic plasticity and memory in offspring. Faseb J. 2021, 35, e21244. [Google Scholar] [CrossRef]
- Chen, W.J.; Niu, J.Q.; Chen, Y.T.; Deng, W.J.; Xu, Y.Y.; Liu, J.; Luo, W.F.; Liu, T. Unilateral facial injection of botulinum neurotoxin a attenuates bilateral trigeminal neuropathic pain and anxiety-like behaviors through inhibition of TLR2-mediated neuroinflammation in mice. J. Headache Pain 2021, 22, 38. [Google Scholar] [CrossRef]
- Verdonk, F.; Roux, P.; Flamant, P.; Fiette, L.; Bozza, F.A.; Simard, S.; Lemaire, M.; Plaud, B.; Shorte, S.L.; Sharshar, T.; et al. Phenotypic clustering: A novel method for microglial morphology analysis. J. Neuroinflamm. 2016, 13, 153. [Google Scholar] [CrossRef] [Green Version]
- Zhuang, H.; Yang, J.; Huang, Z.; Liu, H.; Li, X.; Zhang, H.; Wang, J.; Yu, S.; Liu, K.; Liu, R.; et al. Accelerated age-related decline in hippocampal neurogenesis in mice with noise-induced hearing loss is associated with hippocampal microglial degeneration. Aging 2020, 12, 19493–19519. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Task | Points |
|---|---|
| Exit circle | 1 |
| Monoparesis/hemiparesis | 1 |
| Straight walk | 1 |
| Startle reflex | 1 |
| Seeking behavior | 1 |
| Beam balancing | 1 |
| Round stick balancing | 1 |
| Beam walk: 3 cm | 1 |
| Beam walk: 2 cm | 1 |
| Beam walk: 1 cm | 1 |
| Maximal score | 10 |
| Gene | Forward Primer | Reverse Primer |
|---|---|---|
| IL-12 | GTCCTCAGAAGCTAACCATCTCC | CCAGAGCCTATGACTCCATGTC |
| INF-γ | GAACTGGCAAAAGGATGGTGA | TGTGGGTTGTTGACCTCAAAC |
| CCL2 | TTAAAAACCTGGATCGGAACCAA | GCATTAGCTTCAGATTTACGGGT |
| IL-10 | GGCAGAGAAGCATGGCCCAGAA | AATCGATGACAGCGCCTCAGCC |
| TGF-β | CTCCCGTGGCTTCTAGTGC | GCCTTAGTTTGGACAGGATCTG |
| iNOS | CACCAAGCTGAACTTGAGCG | CGTGGCTTTGGGCTCCTC |
| CD16 | TTTGGACACCCAGATGTTTCAG | GTCTTCCTTGAGCACCTGGATC |
| IL-1β | TGGGAAACAACAGTGGTCAGG | CCATCAGAGGCAAGGAGGAA |
| Ym-1 | CAAGTTGAAGGCTCAGTGGCTC | CAAATCATTGTGTAAAGCTCCTCTC |
| Arg-1 | CCAGAAGAATGGAAGAGTCAGTGT | GCAGATATGCAGGGAGTCACC |
| CD206 | TCTTTGCCTTTCCCAGTCTCC | TGACACCCAGCGGAATTTC |
| NLRP3 | ATTACCCGCCCGAGAAAGG | CATGAGTGTGGCTAGATCCAAG |
| GAPDH | GCCAAGGCTGTGGGCAAGGT | TCTCCAGGCGGCACGTCAGA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, X.; Ou, Y.; Li, J.; Sun, M.; Ge, Q.; Pan, Y.; Cai, Z.; Tan, R.; Wang, W.; An, J.; et al. Voluntary Exercise to Reduce Anxiety Behaviour in Traumatic Brain Injury Shown to Alleviate Inflammatory Brain Response in Mice. Int. J. Mol. Sci. 2023, 24, 6365. https://doi.org/10.3390/ijms24076365
Hu X, Ou Y, Li J, Sun M, Ge Q, Pan Y, Cai Z, Tan R, Wang W, An J, et al. Voluntary Exercise to Reduce Anxiety Behaviour in Traumatic Brain Injury Shown to Alleviate Inflammatory Brain Response in Mice. International Journal of Molecular Sciences. 2023; 24(7):6365. https://doi.org/10.3390/ijms24076365
Chicago/Turabian StyleHu, Xiaoxuan, Yuhang Ou, Jiashuo Li, Meiqi Sun, Qian Ge, Yongqi Pan, Zhenlu Cai, Ruolan Tan, Wenyu Wang, Jing An, and et al. 2023. "Voluntary Exercise to Reduce Anxiety Behaviour in Traumatic Brain Injury Shown to Alleviate Inflammatory Brain Response in Mice" International Journal of Molecular Sciences 24, no. 7: 6365. https://doi.org/10.3390/ijms24076365