
Dental Follicle Stem Cells Promote Periodontal Regeneration through Periostin-Mediated Macrophage Infiltration and Reprogramming in an Inflammatory Microenvironment
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
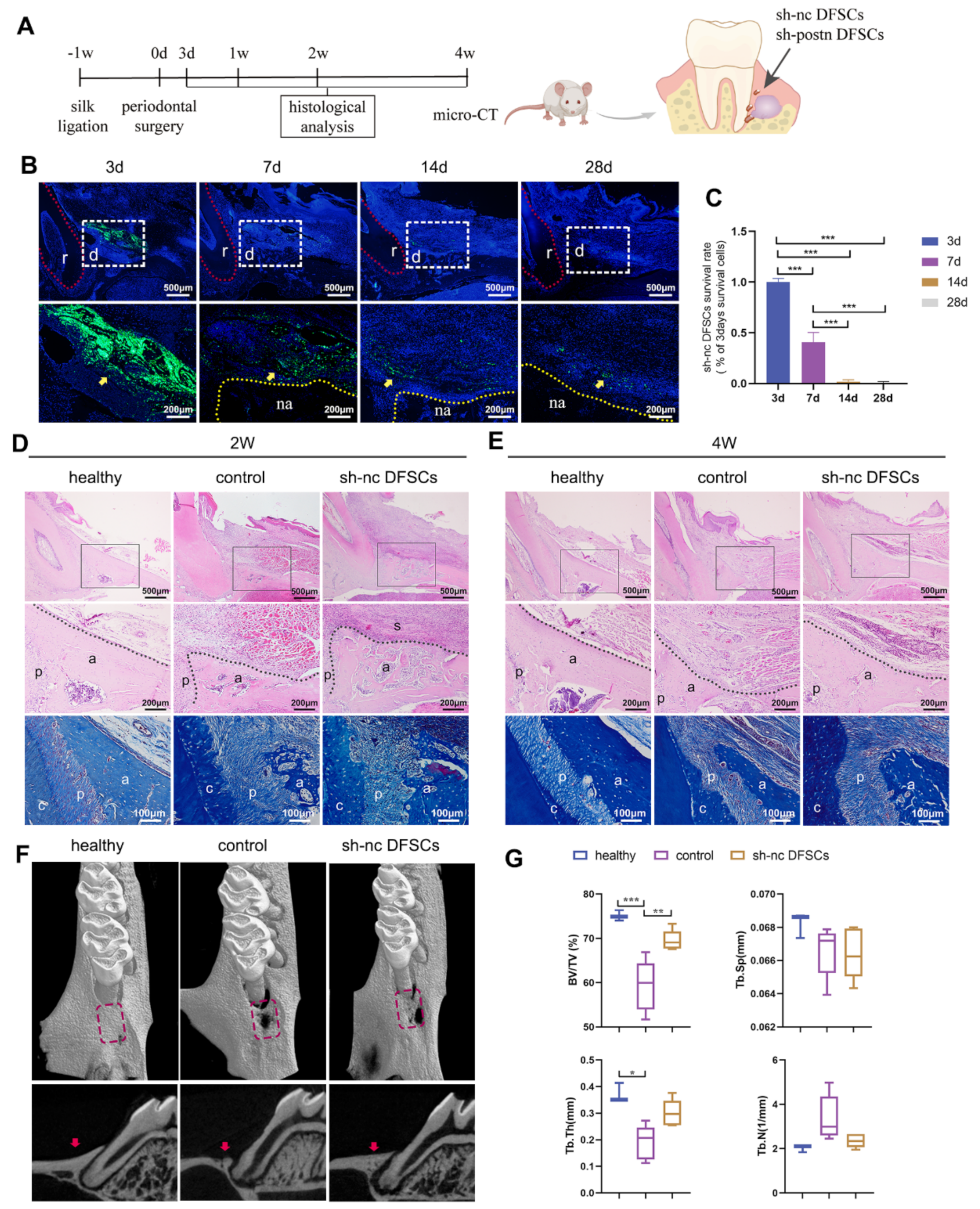
2.1. DFSCs Significantly Promoted Periodontal Regeneration while the Cell Survival Rate Was Low after Transplantation
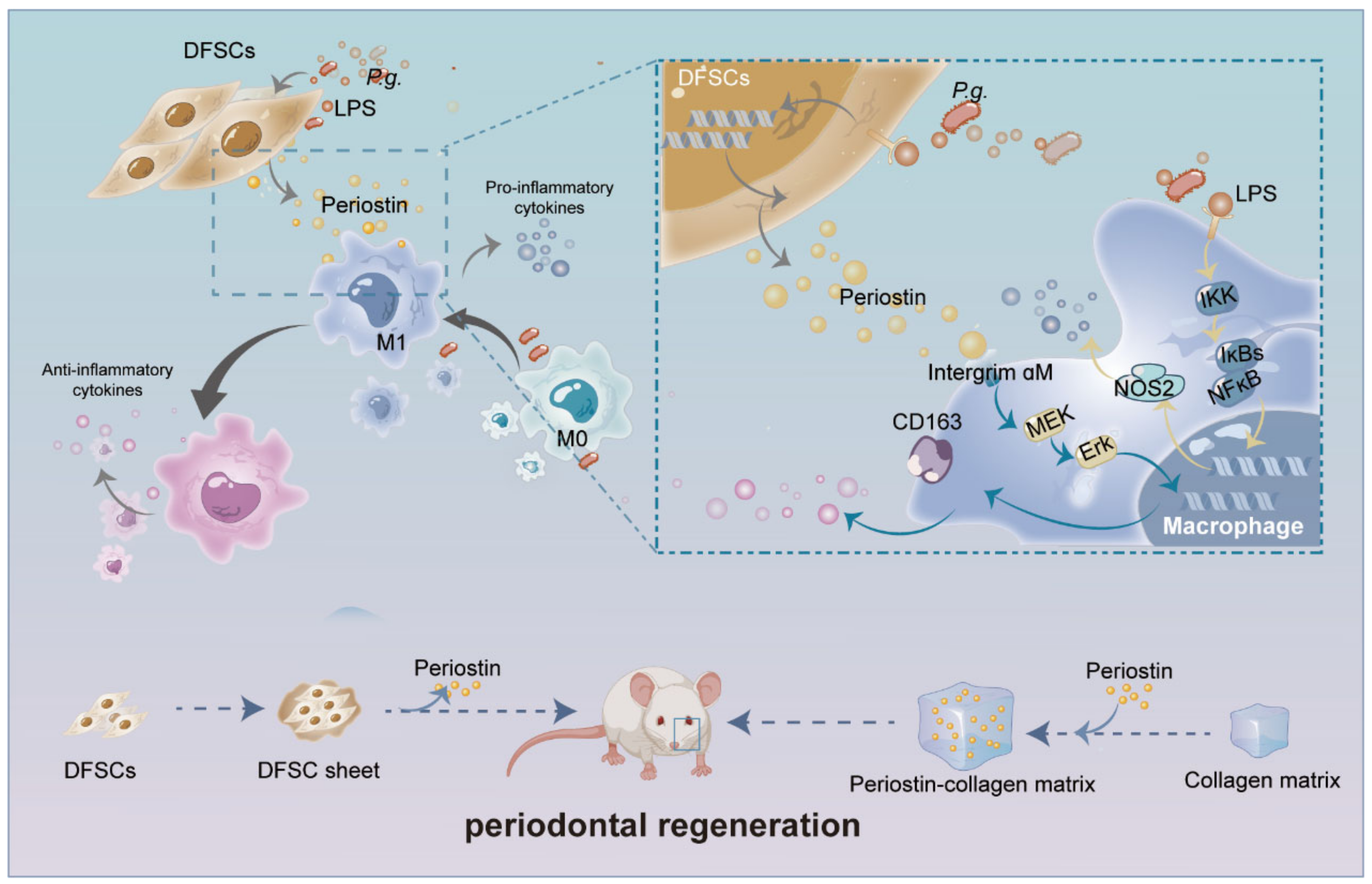
2.2. Transplanted DFSCs Suppressed Inflammatory Responses and Modulated Macrophage Infiltration and Polarization
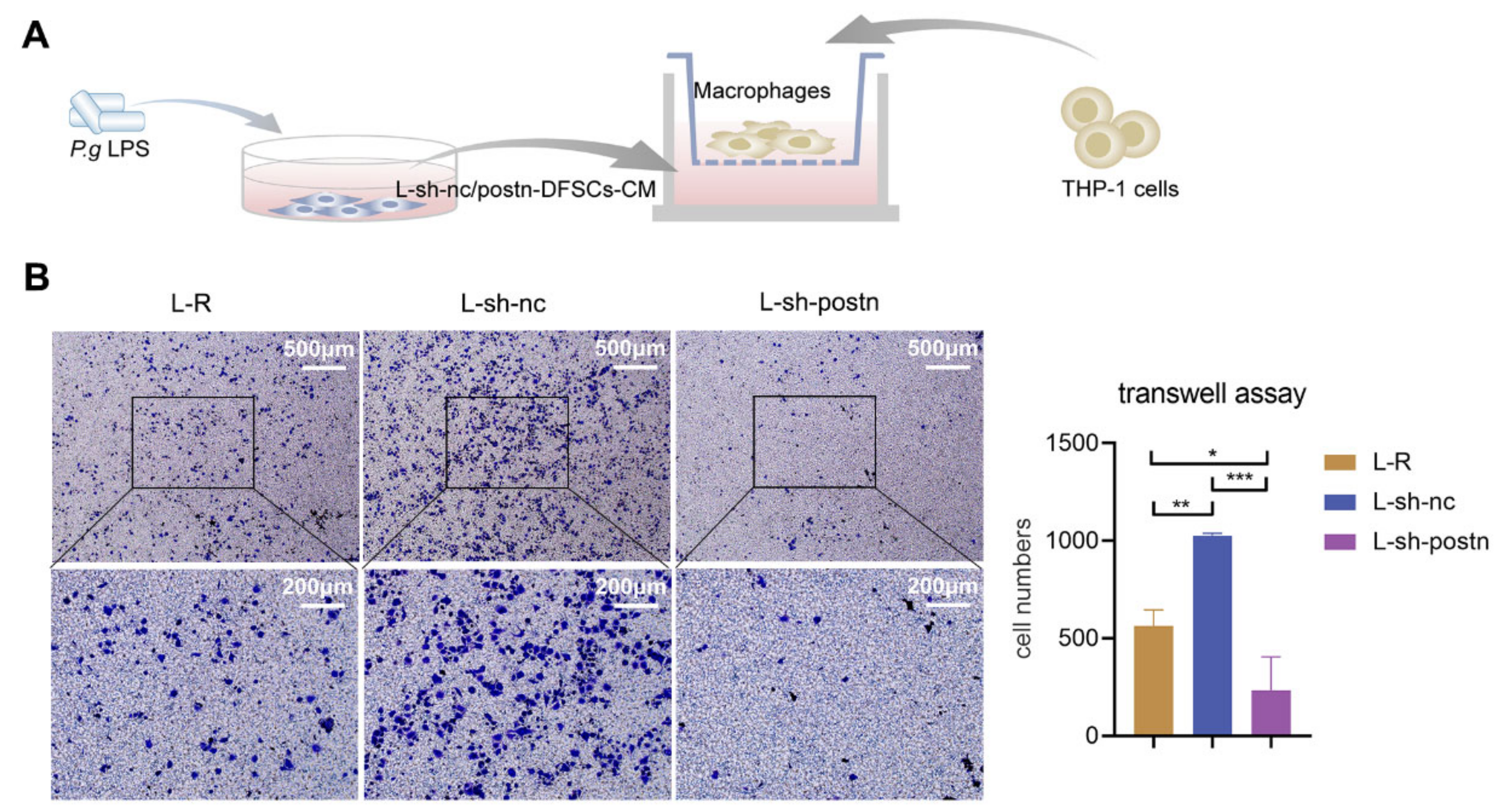
2.3. Silencing Periostin in DFSCs Prevented Macrophage Migration
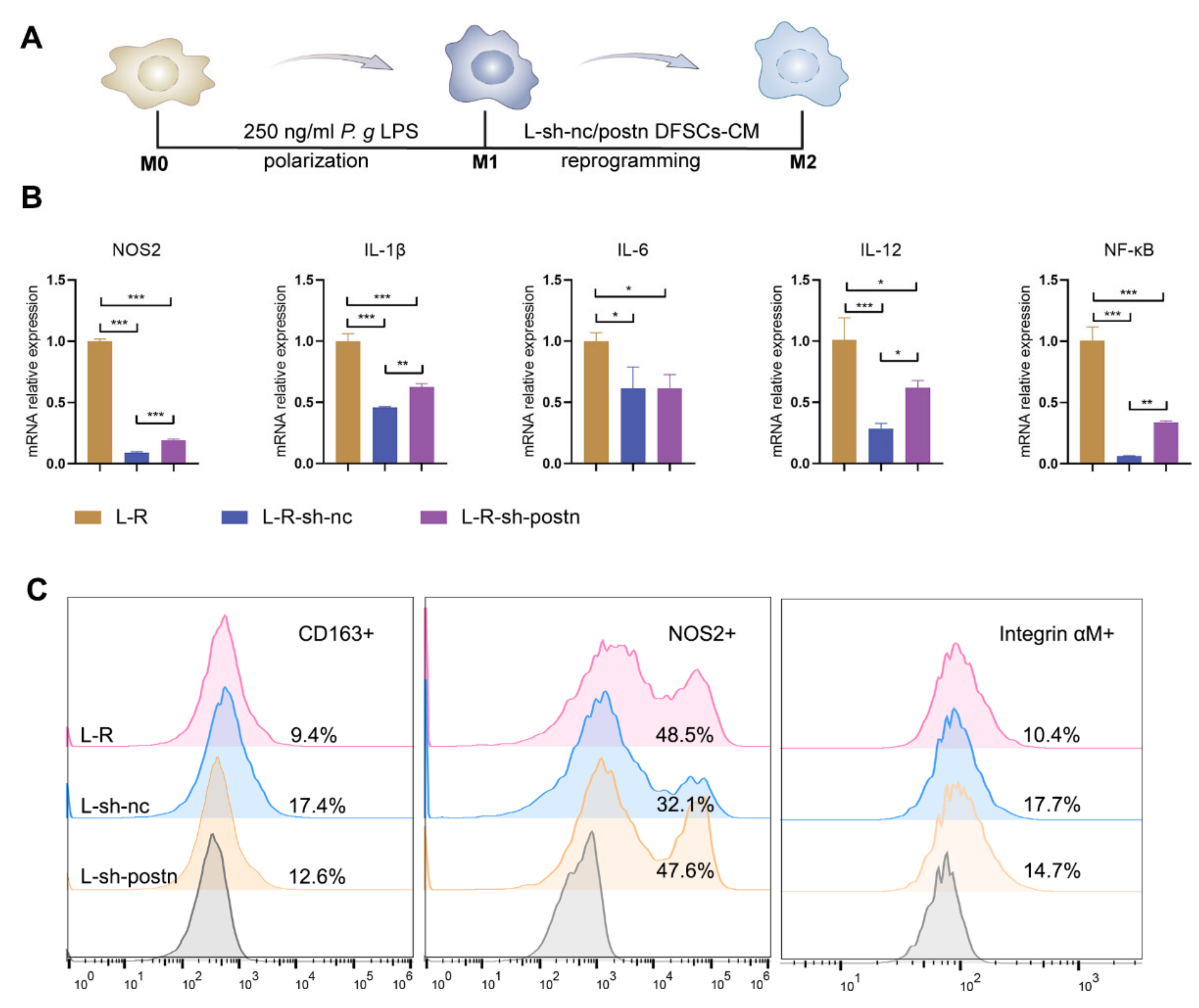
2.4. Silencing Periostin in DFSCs Inhibited the Promotion of Macrophage Reprogramming
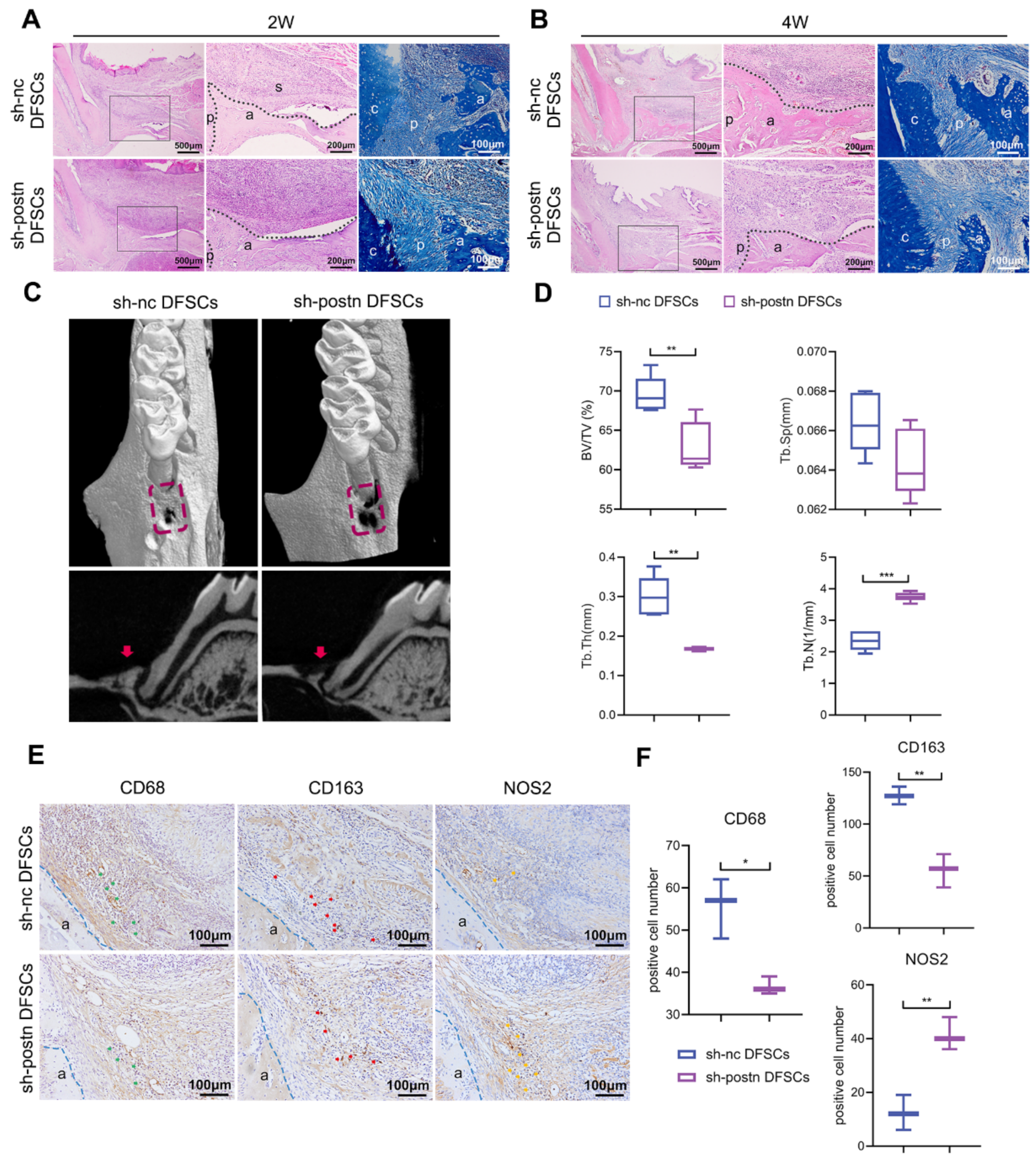
2.5. Silencing Periostin Impaired the Ability of DFSCs to Promote Periodontal Regeneration
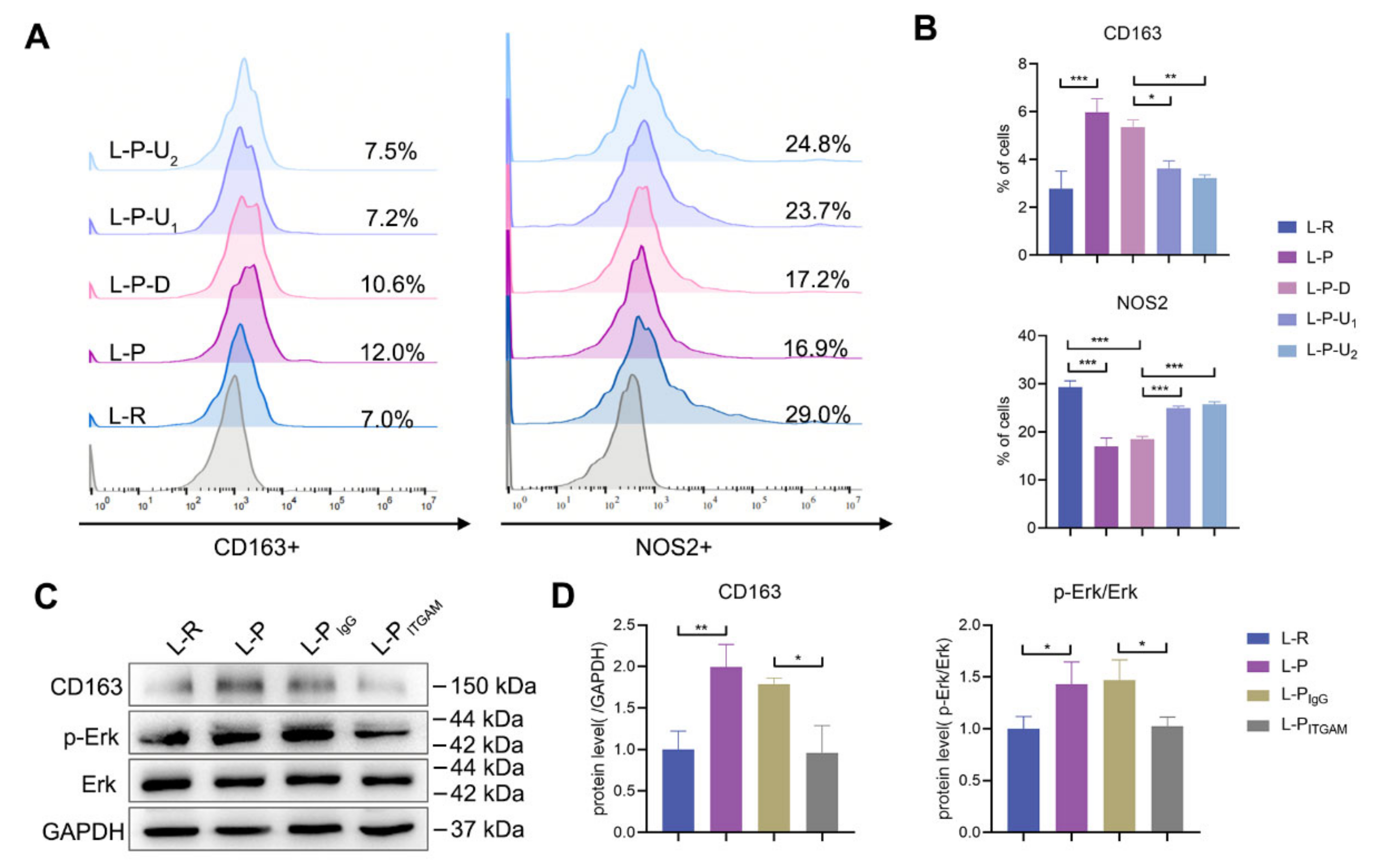
2.6. Periostin Reprogrammed M1 to M2 Macrophages in Part through Integrin αM/p-Erk/Erk Signaling
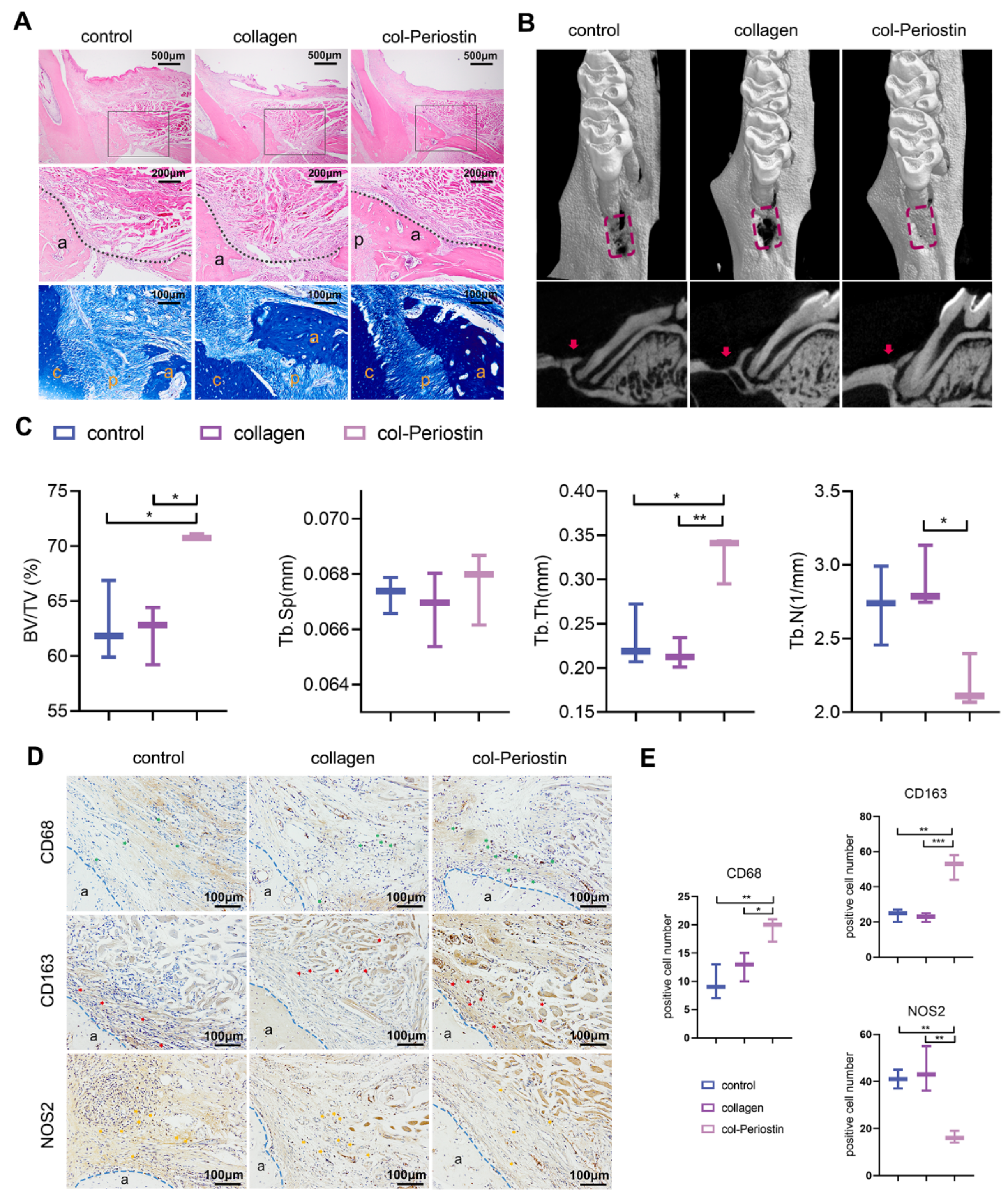
2.7. Periostin–Collagen Matrix Promoted Periodontal Regeneration
3. Discussion
4. Materials and Methods
4.1. DFSC Isolation and Culture
4.2. Lentivirus Transfection
4.3. Preparation of DFSC-Conditioned Medium
4.4. THP-1 Cell Culture
4.5. Transwell Assay
4.6. Quantitative Reverse Transcription–Polymerase Chain Reaction (qRT–PCR)
4.7. Flow Cytometry
4.8. Western Blot
4.9. Animals
4.10. Rat Periodontal Defect Model
4.11. Preparation and Use of Periostin–Collagen Matrix
4.12. Micro-Computed Tomography Evaluation
4.13. Histological, Immunohistochemical, and Immunofluorescent Analyses
4.14. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Tonetti, M.S.; Greenwell, H.; Kornman, K.S. Staging and grading of periodontitis: Framework and proposal of a new classification and case definition. J. Periodontol. 2018, 89, S159–S172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nuñez, J.; Vignoletti, F.; Caffesse, R.G.; Sanz, M. Cellular therapy in periodontal regeneration. Periodontology 2000 2019, 79, 107–116. [Google Scholar] [CrossRef] [PubMed]
- Yan, N.; Hu, B.; Xu, J.; Cai, R.; Liu, Z.; Fu, D.; Huo, B.; Liu, Z.; Zhao, Y.; Chen, C.; et al. Stem cell Janus patch for periodontal regeneration. Nano Today 2022, 42, 101336. [Google Scholar] [CrossRef]
- Li, Q.; Yang, G.; Li, J.; Ding, M.; Zhou, N.; Dong, H.; Mou, Y. Stem cell therapies for periodontal tissue regeneration: A network meta-analysis of preclinical studies. Stem Cell Res. Ther. 2020, 11, 427. [Google Scholar] [CrossRef]
- Wang, P.; Wang, W.; Geng, T.; Liu, Y.; Zhu, S.; Liu, Z.; Yuan, C. EphrinB2 regulates osteogenic differentiation of periodontal ligament stem cells and alveolar bone defect regeneration in beagles. J. Tissue Eng. 2019, 10, 2041731419894361. [Google Scholar] [CrossRef]
- Li, G.; Han, N.; Yang, H.; Zhang, X.; Cao, Y.; Cao, Y.; Shi, R.; Wang, S.; Fan, Z. SFRP2 promotes stem cells from apical papilla-mediated periodontal tissue regeneration in miniature pig. J. Oral Rehabil. 2019, 47, 12–18. [Google Scholar] [CrossRef]
- Yang, H.; Li, J.; Hu, Y.; Sun, J.; Guo, W.; Li, H.; Chen, J.; Huo, F.; Tian, W.; Li, S. Treated dentin matrix particles combined with dental follicle cell sheet stimulate periodontal regeneration. Dent. Mater. 2019, 35, 1238–1253. [Google Scholar] [CrossRef]
- Novello, S.; Debouche, A.; Philippe, M.; Naudet, F.; Jeanne, S. Clinical application of mesenchymal stem cells in periodontal regeneration: A systematic review and meta-analysis. J. Periodontal Res. 2019, 55, 1–12. [Google Scholar] [CrossRef]
- Sánchez, N.; Fierravanti, L.; Núñez, J.; Vignoletti, F.; González-Zamora, M.; Santamaría, S.; Suárez-Sancho, S.; Fernández-Santos, M.E.; Figuero, E.; Herrera, D.; et al. Periodontal regeneration using a xenogeneic bone substitute seeded with autologous periodontal ligament-derived mesenchymal stem cells: A 12-month quasi-randomized controlled pilot clinical trial. J. Clin. Periodontol. 2020, 47, 1391–1402. [Google Scholar] [CrossRef]
- Chen, F.-M.; Gao, L.-N.; Tian, B.-M.; Zhang, X.-Y.; Zhang, Y.-J.; Dong, G.-Y.; Lu, H.; Chu, Q.; Xu, J.; Yu, Y.; et al. Treatment of periodontal intrabony defects using autologous periodontal ligament stem cells: A randomized clinical trial. Stem Cell Res. Ther. 2016, 7, 33. [Google Scholar] [CrossRef] [Green Version]
- Wagoner, Z.W.; Zhao, W. Therapeutic implications of transplanted-cell death. Nat. Biomed. Eng. 2021, 5, 379–384. [Google Scholar] [CrossRef]
- He, L.; Liu, L.; Li, T.; Zhuang, D.; Dai, J.; Wang, B.; Bi, L. Exploring the Imbalance of Periodontitis Immune System from the Cellular to Molecular Level. Front. Genet. 2021, 12, 653209. [Google Scholar] [CrossRef]
- Xu, X.W.; Liu, X.; Shi, C.; Sun, H.C. Roles of Immune Cells and Mechanisms of Immune Responses in Periodontitis. Chin. J. Dent. Res. 2021, 24, 219–230. [Google Scholar]
- Liu, Y.; Liu, W.; Hu, C.; Xue, Z.; Wang, G.; Ding, B.; Luo, H.; Tang, L.; Kong, X.; Chen, X.; et al. MiR-17 Modulates Osteogenic Differentiation Through a Coherent Feed-Forward Loop in Mesenchymal Stem Cells Isolated from Periodontal Ligaments of Patients with Periodontitis. STEM CELLS 2011, 29, 1804–1816. [Google Scholar] [CrossRef]
- Liu, N.; Shi, S.; Deng, M.; Tang, L.; Zhang, G.; Liu, N.; Ding, B.; Liu, W.; Liu, Y.; Shi, H.; et al. High levels of β-catenin signaling reduce osteogenic differentiation of stem cells in inflammatory microenvironments through inhibition of the noncanonical Wnt pathway. J. Bone Miner. Res. 2011, 26, 2082–2095. [Google Scholar] [CrossRef]
- Guo, S.; Kang, J.; Ji, B.; Guo, W.; Ding, Y.; Wu, Y.; Tian, W. Periodontal-Derived Mesenchymal Cell Sheets Promote Periodontal Regeneration in Inflammatory Microenvironment. Tissue Eng. Part A 2017, 23, 585–596. [Google Scholar] [CrossRef]
- Huang, Y.; Liu, Q.; Liu, L.; Huo, F.; Guo, S.; Tian, W. Lipopolysaccharide-Preconditioned Dental Follicle Stem Cells Derived Small Extracellular Vesicles Treating Periodontitis via Reactive Oxygen Species/Mitogen-Activated Protein Kinase Signaling-Mediated Antioxidant Effect. Int. J. Nanomed. 2022, 17, 799–819. [Google Scholar] [CrossRef]
- Kii, I.; Ito, H. Periostin and its interacting proteins in the construction of extracellular architectures. Cell. Mol. Life Sci. 2017, 74, 4269–4277. [Google Scholar] [CrossRef]
- Lehmann, A.E.; Scangas, G.A.; Bergmark, R.W.; El Rassi, E.; Stankovic, K.M.; Metson, R. Periostin and Inflammatory Disease: Implications for Chronic Rhinosinusitis. Otolaryngol. Neck Surg. 2019, 160, 965–973. [Google Scholar] [CrossRef]
- Wang, Z.; Xiong, S.; Mao, Y.; Chen, M.; Ma, X.; Zhou, X.; Ma, Z.; Liu, F.; Huang, Z.; Luo, Q.; et al. Periostin promotes immunosuppressive premetastatic niche formation to facilitate breast tumour metastasis. J. Pathol. 2016, 239, 484–495. [Google Scholar] [CrossRef]
- Zhong, H.; Li, X.; Zhang, J.; Wu, X. Overexpression of periostin is positively associated with gastric cancer metastasis through promoting tumor metastasis and invasion. J. Cell. Biochem. 2019, 120, 9927–9935. [Google Scholar] [CrossRef] [PubMed]
- Du, J.; Li, M. Functions of Periostin in dental tissues and its role in periodontal tissues’ regeneration. Cell. Mol. Life Sci. 2017, 74, 4279–4286. [Google Scholar] [CrossRef] [PubMed]
- Rios, H.; Koushik, S.V.; Wang, H.Y.; Wang, J.; Zhou, H.M.; Lindsley, A.; Rogers, R.; Chen, Z.; Maeda, M.; Kruzynska-Frejtag, A.; et al. periostin null mice exhibit dwarfism, incisor enamel defects, and an early-onset periodontal disease-like phenotype. Mol. Cell. Biol. 2005, 25, 11131–11144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rios, H.; Ma, D.; Xie, Y.; Giannobile, W.; Bonewald, L.; Conway, S.; Feng, J. Periostin Is Essential for the Integrity and Function of the Periodontal Ligament During Occlusal Loading in Mice. J. Periodontol. 2008, 79, 1480–1490. [Google Scholar] [CrossRef] [PubMed]
- Matsuzawa, M.; Arai, C.; Nomura, Y.; Murata, T.; Yamakoshi, Y.; Oida, S.; Hanada, N.; Nakamura, Y. Periostin of human periodontal ligament fibroblasts promotes migration of human mesenchymal stem cell through the v3 integrin/FAK/PI3K/Akt pathway. J. Periodontal Res. 2015, 50, 855–863. [Google Scholar] [CrossRef]
- Padial-Molina, M.; Volk, S.L.; Rios, H.F. Periostin increases migration and proliferation of human periodontal ligament fibroblasts challenged by tumor necrosis factor -α and Porphyromonas gingivalis lipopolysaccharides. J. Periodontal Res. 2013, 49, 405–414. [Google Scholar] [CrossRef] [Green Version]
- Wu, Z.; Dai, W.; Wang, P.; Zhang, X.; Tang, Y.; Liu, L.; Wang, Q.; Li, M.; Tang, C. Periostin promotes migration, proliferation, and differentiation of human periodontal ligament mesenchymal stem cells. Connect. Tissue Res. 2017, 59, 108–119. [Google Scholar] [CrossRef]
- Yan, Y.; Zhang, H.; Liu, L.; Chu, Z.; Ge, Y.; Wu, J.; Liu, Y.; Tang, C. Periostin reverses high glucose-inhibited osteogenesis of periodontal ligament stem cells via AKT pathway. Life Sci. 2019, 242, 117184. [Google Scholar] [CrossRef]
- He, X.-T.; Li, X.; Xia, Y.; Yin, Y.; Wu, R.-X.; Sun, H.-H.; Chen, F.-M. Building capacity for macrophage modulation and stem cell recruitment in high-stiffness hydrogels for complex periodontal regeneration: Experimental studies in vitro and in rats. Acta Biomater. 2019, 88, 162–180. [Google Scholar] [CrossRef]
- Chew, J.R.J.; Chuah, S.J.; Teo, K.Y.W.; Zhang, S.; Lai, R.C.; Fu, J.H.; Lim, L.P.; Lim, S.K.; Toh, W.S. Mesenchymal stem cell exosomes enhance periodontal ligament cell functions and promote periodontal regeneration. Acta Biomater. 2019, 89, 252–264. [Google Scholar] [CrossRef]
- Wang, W.; Zheng, C.; Yang, J.; Li, B. Intersection between macrophages and periodontal pathogens in periodontitis. J. Leukoc. Biol. 2021, 110, 577–583. [Google Scholar] [CrossRef]
- Shi, W.; Guo, S.; Liu, L.; Liu, Q.; Huo, F.; Ding, Y.; Tian, W. Small Extracellular Vesicles from Lipopolysaccharide-Preconditioned Dental Follicle Cells Promote Periodontal Regeneration in an Inflammatory Microenvironment. ACS Biomater. Sci. Eng. 2020, 6, 5797–5810. [Google Scholar] [CrossRef]
- Yang, L.; Guo, P.; Wang, P.; Wang, W.; Liu, J. IL-6/ERK signaling pathway participates in type I IFN-programmed, unconventional M2-like macrophage polarization. Sci. Rep. 2023, 13, 1827. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Ding, H.; Liu, X.; Sheng, Y.; Liu, X.; Jiang, C. Dental Follicle Stem Cells: Tissue Engineering and Immunomodulation. Stem Cells Dev. 2019, 28, 986–994. [Google Scholar] [CrossRef]
- Bi, R.; Lyu, P.; Song, Y.; Li, P.; Song, D.; Cui, C.; Fan, Y. Function of Dental Follicle Progenitor/Stem Cells and Their Potential in Regenerative Medicine: From Mechanisms to Applications. Biomolecules 2021, 11, 997. [Google Scholar] [CrossRef]
- Tour, G.; Wendel, M.; Tcacencu, I.; Moll, G. Bone Repair Using Periodontal Ligament Progenitor Cell-seeded Constructs. J. Dent. Res. 2012, 91, 789–794. [Google Scholar] [CrossRef]
- Yang, Y.; Rossi, F.M.; Putnins, E.E. Periodontal regeneration using engineered bone marrow mesenchymal stromal cells. Biomaterials 2010, 31, 8574–8582. [Google Scholar] [CrossRef]
- Yu, X.; Ge, S.; Chen, S.; Xu, Q.; Zhang, J.; Guo, H.; Yang, P. Human Gingiva-Derived Mesenchymal Stromal Cells Contribute to Periodontal Regeneration in Beagle Dogs. Cells Tissues Organs 2013, 198, 428–437. [Google Scholar] [CrossRef]
- Iwasaki, K.; Akazawa, K.; Nagata, M.; Komaki, M.; Honda, I.; Morioka, C.; Yokoyama, N.; Ayame, H.; Yamaki, K.; Tanaka, Y.; et al. The Fate of Transplanted Periodontal Ligament Stem Cells in Surgically Created Periodontal Defects in Rats. Int. J. Mol. Sci. 2019, 20, 192. [Google Scholar] [CrossRef] [Green Version]
- Yu, N.; Bronckers, A.L.; Oortgiesen, D.A.; Yan, X.; Jansen, J.A.; Yang, F.; Walboomers, X.F. Periodontal Cell Implantation Contributes to the Regeneration of the Periodontium in an Indirect Way. Tissue Eng. Part A 2015, 21, 166–173. [Google Scholar] [CrossRef] [Green Version]
- Villarreal-Ponce, A.; Tiruneh, M.W.; Lee, J.; Guerrero-Juarez, C.F.; Kuhn, J.; David, J.A.; Dammeyer, K.; Kell, R.M.; Kwong, J.; Rabbani, P.S.; et al. Keratinocyte- Macrophage Crosstalk by the Nrf2/Ccl2/EGF Signaling Axis Orchestrates Tissue Repair. Cell Rep. 2020, 33, 108417. [Google Scholar] [CrossRef] [PubMed]
- Cattin, A.-L.; Burden, J.J.; Van Emmenis, L.; Mackenzie, F.E.; Hoving, J.J.; Calavia, N.G.; Guo, Y.; McLaughlin, M.; Rosenberg, L.H.; Quereda, V.; et al. Macrophage-Induced Blood Vessels Guide Schwann Cell-Mediated Regeneration of Peripheral Nerves. Cell 2015, 162, 1127–1139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shang, M.; Cappellesso, F.; Amorim, R.; Serneels, J.; Virga, F.; Eelen, G.; Carobbio, S.; Rincon, M.Y.; Maechler, P.; De Bock, K.; et al. Macrophage-derived glutamine boosts satellite cells and muscle regeneration. Nature 2020, 587, 626–631. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Bi, C.; Gao, L.; An, Y.; Chen, F.; Chen, F. Macrophage polarization in human gingival tissue in response to periodontal disease. Oral Dis. 2018, 25, 265–273. [Google Scholar] [CrossRef] [Green Version]
- Miao, Y.; He, L.; Qi, X.; Lin, X. Injecting Immunosuppressive M2 Macrophages Alleviates the Symptoms of Periodontitis in Mice. Front. Mol. Biosci. 2020, 7, 603817. [Google Scholar] [CrossRef]
- Yang, J.; Zhu, Y.; Duan, D.; Wang, P.; Xin, Y.; Bai, L.; Liu, Y.; Xu, Y. Enhanced activity of macrophage M1/M2 phenotypes in periodontitis. Arch. Oral Biol. 2018, 96, 234–242. [Google Scholar] [CrossRef]
- Almubarak, A.; Tanagala, K.K.K.; Papapanou, P.N.; Lalla, E.; Momen-Heravi, F. Disruption of Monocyte and Macrophage Homeostasis in Periodontitis. Front. Immunol. 2020, 11, 330. [Google Scholar] [CrossRef]
- Chen, X.C.; Yang, B.; Tian, J.; Hong, H.; Du, Y.; Li, K.; Li, X.; Wang, N.; Yu, X.Q.; Wei, X. Dental Follicle Stem Cells Ameliorate Lipopolysaccharide-Induced Inflammation by Secreting TGF-beta 3 and TSP-1 to Elicit Macrophage M2 Polarization. Cell. Physiol. Biochem. 2018, 51, 2290–2308. [Google Scholar] [CrossRef]
- Zhou, W.; Ke, S.Q.; Huang, Z.; Flavahan, W.; Fang, X.; Paul, J.; Wu, L.; Sloan, A.E.; McLendon, R.E.; Li, X.; et al. Periostin secreted by glioblastoma stem cells recruits M2 tumour-associated macrophages and promotes malignant growth. Nat. Cell Biol. 2015, 17, 170–182. [Google Scholar] [CrossRef] [Green Version]
- Guo, X.; Xue, H.; Shao, Q.; Wang, J.; Guo, X.; Chen, X.; Zhang, J.; Xu, S.; Li, T.; Zhang, P.; et al. Hypoxia promotes glioma-associated macrophage infiltration via periostin and subsequent M2 polarization by upregulating TGF-beta and M-CSFR. Oncotarget 2016, 7, 80521–80542. [Google Scholar] [CrossRef] [Green Version]
- Yokota, K.; Kobayakawa, K.; Saito, T.; Hara, M.; Kijima, K.; Ohkawa, Y.; Harada, A.; Okazaki, K.; Ishihara, K.; Yoshida, S.; et al. Periostin Promotes Scar Formation through the Interaction between Pericytes and Infiltrating Monocytes/Macrophages after Spinal Cord Injury. Am. J. Pathol. 2017, 187, 639–653. [Google Scholar] [CrossRef] [Green Version]
- Tang, M.; Liu, B.; Bu, X.; Zhao, P. Cross-talk between ovarian cancer cells and macrophages through periostin promotes macrophage recruitment. Cancer Sci. 2018, 109, 1309–1318. [Google Scholar] [CrossRef] [Green Version]
- Liao, Y.; Li, G.; Zhang, X.; Huang, W.; Xie, D.; Dai, G.; Zhu, S.; Lu, D.; Zhang, Z.; Lin, J.; et al. Cardiac Nestin+ Mesenchymal Stromal Cells Enhance Healing of Ischemic Heart through Periostin-Mediated M2 Macrophage Polarization. Mol. Ther. 2020, 28, 855–873. [Google Scholar] [CrossRef]
- Sun, X.; Gao, J.; Meng, X.; Lu, X.; Zhang, L.; Chen, R. Polarized Macrophages in Periodontitis: Characteristics, Function, and Molecular Signaling. Front. Immunol. 2021, 12, 4953. [Google Scholar] [CrossRef]
- Kudo, A.; Kii, I. Periostin function in communication with extracellular matrices. J. Cell Commun. Signal. 2018, 12, 301–308. [Google Scholar] [CrossRef] [Green Version]
- Vimalathas, P.; Farris, A.; Letner, D.; Deshpande, V.; Yajnik, V.; Shreffler, W.; Garber, J. Integrin αM activation and upregulation on esophageal eosinophils and periostin-mediated eosinophil survival in eosinophilic esophagitis. Immunol. Cell Biol. 2018, 96, 426–438. [Google Scholar] [CrossRef]
- Watanabe, T.; Yasue, A.; Fujihara, S.; Tanaka, E. PERIOSTIN regulates MMP-2 expression via the alpha v beta 3 integrin/ERK pathway in human periodontal ligament cells. Arch. Oral Biol. 2012, 57, 52–59. [Google Scholar] [CrossRef]
- Liu, J.; Wang, L.; Liu, W.; Li, Q.; Jin, Z.; Jin, Y. Dental Follicle Cells Rescue the Regenerative Capacity of Periodontal Ligament Stem Cells in an Inflammatory Microenvironment. PLoS ONE 2014, 9, e108752. [Google Scholar] [CrossRef]
- Iwasaki, K.; Peng, Y.; Kanda, R.; Umeda, M.; Ishikawa, I. Stem Cell Transplantation and Cell-Free Treatment for Periodontal Regeneration. Int. J. Mol. Sci. 2022, 23, 1011. [Google Scholar] [CrossRef]
- Li, X.; He, X.-T.; Yin, Y.; Wu, R.-X.; Tian, B.-M.; Chen, F.-M. Administration of signalling molecules dictates stem cell homing for in situ regeneration. J. Cell. Mol. Med. 2017, 21, 3162–3177. [Google Scholar] [CrossRef]
- Liu, A.-Q.; Hu, C.-H.; Jin, F.; Zhang, L.-S.; Xuan, K. Contributions of Bioactive Molecules in Stem Cell-Based Periodontal Regeneration. Int. J. Mol. Sci. 2018, 19, 1016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, X.-Y.; Li, X.; Wang, J.; He, X.-T.; Sun, H.-H.; Chen, F.-M. Concise Review: Periodontal Tissue Regeneration Using Stem Cells: Strategies and Translational Considerations. STEM CELLS Transl. Med. 2018, 8, 392–403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, Y.; Liu, L.; Wang, P.; Chen, D.; Wu, Z.; Tang, C. Periostin promotes migration and osteogenic differentiation of human periodontal ligament mesenchymal stem cells via the Jun amino-terminal kinases (JNK) pathway under inflammatory conditions. Cell Prolif. 2017, 50, e12369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, S.; Guo, W.; Ding, Y.; Gong, J.; Zou, Q.; Xie, D.; Chen, Y.; Wu, Y.; Tian, W. Comparative Study of Human Dental Follicle Cell Sheets and Periodontal Ligament Cell Sheets for Periodontal Tissue Regeneration. Cell Transplant. 2013, 22, 1061–1073. [Google Scholar] [CrossRef] [Green Version]
- Yi, G.; Zhang, S.; Ma, Y.; Yang, X.; Huo, F.; Chen, Y.; Yang, B.; Tian, W. Matrix vesicles from dental follicle cells improve alveolar bone regeneration via activation of the PLC/PKC/MAPK pathway. Stem Cell Res. Ther. 2022, 13, 1–20. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wei, X.; Guo, S.; Liu, Q.; Liu, L.; Huo, F.; Wu, Y.; Tian, W. Dental Follicle Stem Cells Promote Periodontal Regeneration through Periostin-Mediated Macrophage Infiltration and Reprogramming in an Inflammatory Microenvironment. Int. J. Mol. Sci. 2023, 24, 6353. https://doi.org/10.3390/ijms24076353
Wei X, Guo S, Liu Q, Liu L, Huo F, Wu Y, Tian W. Dental Follicle Stem Cells Promote Periodontal Regeneration through Periostin-Mediated Macrophage Infiltration and Reprogramming in an Inflammatory Microenvironment. International Journal of Molecular Sciences. 2023; 24(7):6353. https://doi.org/10.3390/ijms24076353
Chicago/Turabian StyleWei, Xiuqun, Shujuan Guo, Qian Liu, Li Liu, Fangjun Huo, Yafei Wu, and Weidong Tian. 2023. "Dental Follicle Stem Cells Promote Periodontal Regeneration through Periostin-Mediated Macrophage Infiltration and Reprogramming in an Inflammatory Microenvironment" International Journal of Molecular Sciences 24, no. 7: 6353. https://doi.org/10.3390/ijms24076353