

Karyotype Evolution in Triatominae (Hemiptera, Reduviidae): The Role of Chromosomal Rearrangements in the Diversification of Chagas Disease Vectors
 ,
,  ,
,  and
and
Abstract
:1. Introduction
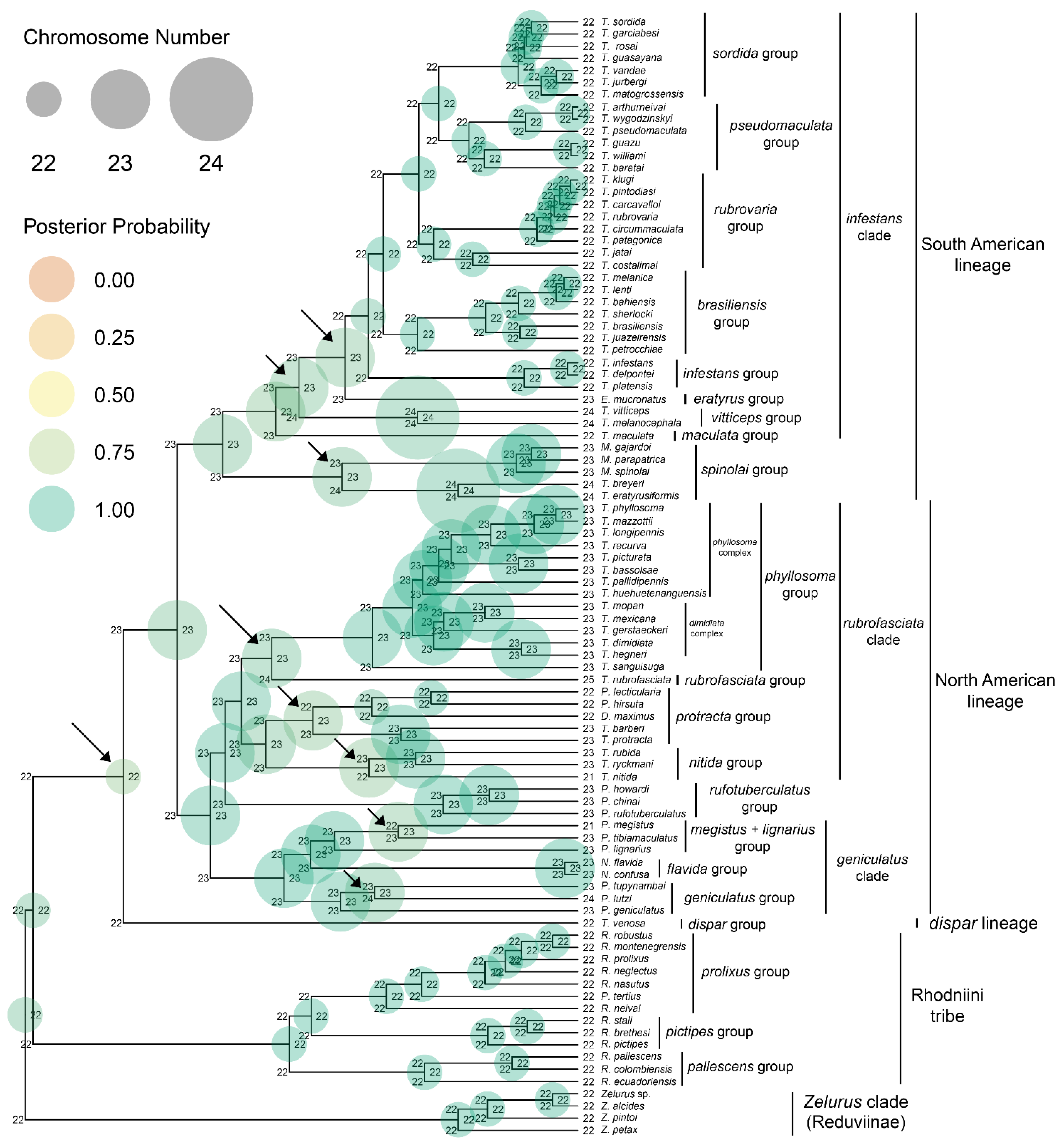
2. Results and Discussion
2.1. Dispar Lineage
2.2. North American Lineage
2.3. South American Lineage
2.4. Chromosomal Changes as a Reproductive Barrier
3. Materials and Methods
3.1. Chromosomal Evolution Modeling
3.2. Experimental Crosses
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rassi, A.; Rassi, A.; Marin-Neto, J.A. Chagas disease. Lancet 2010, 375, 1388–1402. [Google Scholar] [CrossRef]
- World Health Organization. Chagas Disease (American trypanosomiasis). Available online: http://www.who.int/news-room/fact-sheets/detail/chagas-disease-(american-trypanosomiasis) (accessed on 6 October 2022).
- Galvão, C. Vetores da Doença de Chagas no Brasil; Sociedade Brasileira de Zoologia: Curitiba, Brasil, 2014; p. 289. [Google Scholar]
- Dias, J.C.P.; Neto, V.A.; Luna, E.J.A. Alternative transmission mechanisms of Trypanosoma Cruzi in Brazil and proposals for their prevention. Ver. Soc. Bras. Med. Trop. 2011, 44, 375–379. [Google Scholar] [CrossRef] [Green Version]
- Alevi, K.C.C.; Oliveira, J.; Rocha, D.S.; Galvão, C. Trends in taxonomy of Chagas disease vectors (Hemiptera, Reduviidae, Triatominae): From Linnaean to integrative taxonomy. Pathogens 2021, 10, 1627. [Google Scholar] [CrossRef]
- Correia, J.P.S.O.; Gil-Santana, H.R.; Dale, C.; Galvão, C. Triatoma guazu Lent and Wygodzinsky is a junior synonym of Triatoma williami Galvão, Souza and Lima. Insects 2022, 13, 591. [Google Scholar] [CrossRef]
- Gil-Santana, H.R.; Chavez, T.; Pita, S.; Panzera, F.; Galvão, C. Panstrongylus noireaui, a remarkable new species of Triatominae (Hemiptera, Reduviidae) from Bolivia. ZooKeys 2022, 1104, 203–225. [Google Scholar] [CrossRef]
- Ravazi, A.; Oliveira, J.; Campos, F.F.; Madeira, F.F.; Reis, Y.V.; Oliveira, A.B.B.; Azeredo-Oliveira, M.T.V.; Rosa, J.A.; Galvão, C.; Alevi, K.C.C. Trends in evolution of the Rhodniini tribe (Hemiptera, Triatominae): Experimental crosses between Psammolestes tertius Lent & Jurberg, 1965 and P. coreodes Bergroth, 1911 and analysis of the reproductive isolating mechanisms. Parasites Vectors 2021, 14, 350. [Google Scholar] [CrossRef]
- Martínez-Ibarra, J.A.; Ventura-Rodríguez, L.V.; Meillon-Isais, K.; Barajas-Martínez, H.; Alejandre-Aguilar, R.; Lupercio-Coronel, P. Biological and genetic aspects of experimental hybrids from species of the phyllosoma complex (Hemiptera: Reduviidae: Triatominae). Mem. Inst. Oswaldo Cruz 2008, 103, 236–243. [Google Scholar] [CrossRef]
- Martínez-Ibarra, J.A.; Grant-Guillén, Y.; Ventura-Rodríguez, L.V.; Osorio-Pelayo, P.D.; Macías-Amezcua, M.D.; Meillón-Isáis, K. Biological and genetic aspects of crosses between species of the genus Meccus (Hemiptera: Reduviidae Triatominae). Mem. Inst. Oswaldo Cruz 2011, 106, 293–300. [Google Scholar] [CrossRef]
- Mendonça, V.J.; Alevi, K.C.C.; Medeiros, L.M.O.; Nascimento, J.D.; Azeredo-Oliveira, M.T.V.; Rosa, J.A. Cytogenetic and morphologic approaches of hybrids from experimental crosses between Triatoma lenti Sherlock & Serafim, 1967 and T. sherlocki Papa et al., 2002 (Hemiptera: Reduviidae). Infect. Genet. Evol. 2014, 26, 123–131. [Google Scholar] [CrossRef] [Green Version]
- Alevi, K.C.C.; Pinotti, H.; Araújo, R.F.; Azeredo-Oliveira, M.T.V.; Rosa, J.A.; Mendonça, V.J. Hybrid colapse confirm the specific status of Triatoma bahiensis Sherlock and Serafim, 1967 (Hemiptera, Triatominae). Am. J. Trop. Med. Hyg. 2018, 98, 475–477. [Google Scholar] [CrossRef] [Green Version]
- Costa, J.; Almeida, C.E.; Dujardin, J.P.; Beard, C.B. Crossing experiments detect genetic incompatibility among populations of Triatoma brasiliensis Neiva, 1911 (Heteroptera, Reduviidae, Triatominae). Mem. Inst. Oswaldo Cruz 2003, 98, 637–639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mendonca, V.J.; Alevi, K.C.C.; Pinotti, H.; Gurgel-Goncalves, R.; Pita, S.; Guerra, A.L.; Panzera, F.; Araujo, R.F.; Azeredo-Oliveira, M.T.V.; Rosa, J.A. Revalidation of Triatoma bahiensis Sherlock & serafim, 1967 (Hemiptera: Reduviidae) and phylogeny of the T. brasiliensis species complex. Zootaxa 2016, 4107, 239–254. [Google Scholar] [CrossRef] [PubMed]
- Campos-Soto, R.; Panzera, F.; Pita, S.; Lages, C.; Solari, A.; Botto-Mahan, C. Experimental crosses between Mepraia gajardoi and M. spinolai and hybrid chromosome analyses reveal the occurrence of several isolation mechanisms. Infect. Genet. Evol. 2016, 45, 205–212. [Google Scholar] [CrossRef] [PubMed]
- Nascimento, J.D.; Rosa, J.A.; Salgado-Roa, F.C.; Hernández, C.; Pardo-Diaz, C.; Alevi, K.C.C.; Ravazi, A.; Oliveira, J.; Azeredo-Oliveira, M.T.V.; Salazar, C.; et al. Taxonomical over splitting in the Rhodnius prolixus (Insecta: Hemiptera: Reduviidae) clade: Are R. taquarussuensis (Da Rosa et al., 2017) and R. neglectus (Lent, 1954) the same species? PLoS ONE 2019, 14, e0213043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cesaretto, N.R.; Oliveira, J.; Ravazi, A.; Madeira, F.F.; Reis, Y.V.; Oliveira, A.B.B.; Vicente, R.D.; Cristal, D.C.; Galvão, C.; Azeredo-Oliveira, M.T.V.; et al. Trends in taxonomy of Triatomini (Hemiptera, Reduviidae, Triatominae): Reproductive compatibility reinforces the synonymization of Meccus Stål, 1859 with Triatoma Laporte, 1832. Parasites Vectors 2021, 14, 340. [Google Scholar] [CrossRef] [PubMed]
- Neves, J.M.S.; Sousa, P.S.; Oliveira, J.; Ravazi, A.; Madeira, F.F.; Reis, Y.V.; Oliveira, A.B.B.; Pinotti, H.; Azeredo-Oliveira, M.T.V.; Rosa, J.A.; et al. Prezygotic isolation confirms the exclusion of Triatoma melanocephala, T. vitticeps and T. tibiamaculata of the T. brasiliensis subcomplex (Hemiptera, Triatominae). Infect. Genet. Evol. 2020, 79, 104149. [Google Scholar] [CrossRef]
- Alevi, K.C.C.; Oliveira, J.; Garcia, A.C.C.; Cristal, D.C.; Delgado, L.M.G.; Bittinelli, I.D.F.; Reis, Y.V.; Ravazi, A.; Oliveira, A.B.B.; Galvão, C.; et al. Triatoma rosai sp. nov. (Hemiptera, Triatominae): A new species of Argentinian Chagas disease vector described based on integrative taxonomy. Insects 2020, 11, 830. [Google Scholar] [CrossRef]
- Pinotti, H.; Oliveira, J.; Ravazi, A.; Madeira, F.F.; Reis, Y.V.; Oliveira, A.B.B.; Azeredo-Oliveira, M.T.V.; Rosa, J.A.; Alevi, K.C.C. Revisiting the hybridization processes in the Triatoma brasiliensis complex (Hemiptera, Triatominae): Interspecific genomic compatibility point to a possible recent diversification of the species grouped in this monophyletic complex. PLoS ONE 2021, 16, e0257992. [Google Scholar] [CrossRef]
- Madeira, F.F.; Delgado, L.M.G.; Bittinelli, I.F.; Oliveira, J.; Ravazi, A.; Reis, Y.V.; Oliveira, A.B.B.; Cristal, D.C.; Galvao, C.; Azeredo-Oliveira, M.T.V.; et al. Triatoma sordida (Stål, 1859) from La Paz, Bolivia (Hemiptera, Triatominae): An incipient species or an intraspecific chromosomal polymorphism? Parasites Vectors 2021, 14, 553. [Google Scholar] [CrossRef]
- Delgado, L.M.G.; Oliveira, J.; Ravazi, A.; Madeira, F.F.; Reis, Y.V.; Pinotti, H.; Oliveira, A.B.B.; Masarin, I.S.; Lilioso, M.; Folly-Ramos, E.; et al. Revisiting the hybridization processes in the Triatoma brasiliensis complex (Hemiptera, Triatominae): Reproductive isolation between Triatoma petrocchiae and T. b. brasiliensis and T. lenti. Insects 2021, 12, 1015. [Google Scholar] [CrossRef]
- Ravazi, A.; Oliveira, J.; Madeira, F.F.; Reis, Y.V.; Oliveira, A.B.B.; Galvão, C.; Azeredo-Oliveira, M.T.V.; Rosa, J.A.; Alevi, K.C.C. Trends in taxonomy of the Rhodniini tribe (Hemiptera, Triatominae): Reproductive incompatibility between Rhodnius neglectus Lent, 1954 and Psammolestes spp. confirms the generic status of Psammolestes Bergroth, 1911. Diversity 2022, 14, 761. [Google Scholar] [CrossRef]
- Martínez-Ibarra, J.A.; Nogueda-Torres, B.; Salazar-Schettino, P.M.; Cabrera-Bravo, M.; Vences-Blanco, M.O.; Rocha-Chávez, G. Transmission capacity of Trypanosoma cruzi (Trypanosomatida: Trypanosomatidae) by three subspecies of Meccus phyllosomus (Heteroptera: Reduviidae) and their hybrids. J. Med. Entomol. 2016, 53, 928–934. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Ibarra, J.A.; Nogueda-Torres, B.; Salazar-Montaño, L.F.; García-Lino, J.C.; Arroyo-Reyes, D.; Hernández-Navarro, J.Á. Comparison of biological fitness in crosses between subspecies of Meccus phyllosomus (Hemiptera: Reduviidae: Triatominae) in southern Mexico. Insect Sci. 2017, 24, 114–121. [Google Scholar] [CrossRef] [PubMed]
- Meraz-Medina, T.; Nogueda-Torres, B.; Ceballos-Rodríguez, R.S.; Godínez-Aceves, K.C.; Martínez-Ibarra, J.A. Enhancing fitness in offspring of crosses between two triatomine species. J. Vector Ecol. 2019, 44, 173–178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez-Ibarra, J.A.; Cárdenas-Sosa, M.A.; Montañez-Valdez, O.D.; Michel-Parra, J.G.; Nogueda-Torres, B. Biological parameters and estimation of the vectorial capacity of two subspecies of Triatoma protracta (Uhler) and their laboratory hybrids in Mexico. J. Vector Ecol. 2021, 46, 148–154. [Google Scholar] [CrossRef] [PubMed]
- Díaz, S.; Panzera, F.; Jaramillo-O, N.; Pérez, R.; Fernández, R.; Vallejo, G.; Saldaña, A.; Calzada, J.E.; Triana, O.; Gómez-Palacio, A. Genetic, cytogenetic and morphological trends in the evolution of the Rhodnius (Triatominae: Rhodniini) trans-andean group. PLoS ONE 2014, 9, e87493. [Google Scholar] [CrossRef] [Green Version]
- Perlowagora-Szumlewics, A.; Correia, M.V. Induction of male sterility manipulation of genetic mechanisms present in vector species of Chagas disease (remarks on integrating sterile-male release with insecticidal control measures against vectors of Chagas disease). Rev. Inst. Med. Trop. São Paulo 1972, 14, 360–371. [Google Scholar]
- Pérez, R.; Hernández, M.; Quintero, O.; Scovortzoff, E.; Canale, D.; Méndez, L.; Cohanoff, C.; Martino, M.; Panzera, F. Cytogenetic analysis of experimental hybrids in species of Triatominae (Hemiptera-Reduviidae). Genetica 2005, 125, 261–270. [Google Scholar] [CrossRef]
- Perlowagora-Szumlewicz, A.; Correia, M.V.; Trinchet, A.M.R. Induction of male sterility through manipulation of genetic mechanisms present in vector species of triatominae. II. Partial restoration of male fertility. Ver. Soc. Bras. Med. Trop. 1976, 10, 367–383. [Google Scholar] [CrossRef] [Green Version]
- Reis, Y.V.; Oliveira, J.; Madeira, F.F.; Ravazi, A.; Oliveira, A.B.B.; Mello, D.V.; Campos, F.F.; Azeredo-Oliveira, M.T.V.; Rosa, J.A.; Galvão, C.; et al. Trends in evolution of the Triatomini tribe (Hemiptera, Triatominae): Reproductive incompatibility between four species of geniculatus clade. Parasites Vectors 2022, 15, 403. [Google Scholar] [CrossRef]
- Vos, J.M.; Augustijnen, H.; Bätscher, L.; Lucek, K. Speciation through chromosomal fusion and fission in Lepidoptera. Philos. Trans. R. Soc. 2020, 375, 20190539. [Google Scholar] [CrossRef]
- Ueshima, N. Cytotaxonomy of the Triatominae (Reduviidae: Hemiptera). Chromosoma 1966, 18, 97–122. [Google Scholar] [CrossRef]
- Panzera, F.; Pérez, R.; Panzera, Y.; Ferrandis, I.; Ferreiro, M.J.; Calleros, L. Cytogenetics and genome evolution in the subfamily Triatominae (Hemiptera, Reduviidae). Cytogenet. Genome Res. 2010, 128, 77–87. [Google Scholar] [CrossRef]
- Panzera, F.; Pita, S.; Lorite, P. Chromosome structure and evolution of triatominae: A review. In Triatominae—The Biology of Chagas Disease Vectors, Entomology in Focus 5; Guarneri, A., Lorenzo, M., Eds.; Springer: Cham, Switzerland, 2021; pp. 65–99. [Google Scholar]
- Pita, S.; Lorite, P.; Cuadrado, A.; Panzera, Y.; Oliveira, J.; Alevi, K.C.C.; Rosa, J.A.; Freitas, S.P.C.; Gómez-Palacio, A.; Solari, A.; et al. High chromosomal mobility of ribosomal clusters in holocentric chromosomes of Triatominae, vectors of Chagas disease (Hemiptera-Reduviidae). Med. Vet. Entomol. 2022, 36, 66–80. [Google Scholar] [CrossRef]
- Alevi, K.C.C.; Oliveira, J.; Rosa, J.A.; Azeredo-Oliveira, M.T.V. Karyotype evolution of Chagas disease vectors (Hemiptera, Triatominae). Am. J. Trop Med. Hyg. 2018, 99, 87–89. [Google Scholar] [CrossRef]
- Reis, Y.V.; Alevi, K.C.C. Revisão cariotípica dos vetores da doença de Chagas. In Atualidades em Medicina Tropical na América do Sul: Vetores, 1st ed.; Oliveira, J., Alevi, K.C.C., Camargo, L.M.A., Meneguetti, D.U.O., Eds.; Stricto Sensu: Rio Branco, Acre, Brasil, 2021; pp. 70–79. [Google Scholar]
- Mayrose, I.; Barker, M.S.; Otto, S.P. Probabilistic models of chromosome number evolution and the inference of polyploidy. Syst. Biol. 2010, 59, 132–144. [Google Scholar] [CrossRef] [Green Version]
- Glick, L.; Mayrose, I. ChromEvol: Assessing the pattern of chromosome number evolution and the inference of polyploidy along a phylogeny. Mol. Biol. Evol. 2014, 31, 1914–1922. [Google Scholar] [CrossRef]
- Freyman, W.A.; Höhna, S. Cladogenetic and anagenetic models of chromosome number evolution: A bayesian model averaging approach. Syst. Biol. 2018, 67, 195–215. [Google Scholar] [CrossRef] [Green Version]
- Justi, S.A.; Russo, C.A.M.; Mallet, J.R.D.S.; Obara, M.T.; Galvão, C. Molecular phylogeny of Triatomini (Hemiptera: Reduviidae: Triatominae). Parasites Vectors 2014, 7, 149. [Google Scholar] [CrossRef] [Green Version]
- Justi, S.A.; Galvão, C.; Schrago, C.G. Geological changes of the Americas and their influence on the diversification of the Neotropical kissing bugs (Hemiptera: Reduviidae: Triatominae). PLoS Negl. Trop. Dis. 2016, 10, e0004527. [Google Scholar] [CrossRef] [Green Version]
- Belintani, T.; Oliveira, J.; Pinotti, H.; Silva, L.A.; Alevi, K.C.C.; Galvão, C.; Rosa, J.A. Phylogenetic and phenotypic relationships of the Triatoma sordida subcomplex (Hemiptera: Reduviidae: Triatominae). Acta Trop. 2020, 212, 105679. [Google Scholar] [CrossRef]
- Bittinelli, I.F.; Oliveira, J.; Reis, Y.V.; Ravazi, A.; Madeira, F.F.; Oliveira, A.B.B.; Montanari, G.; Gomes, A.J.C.; Cesaretto, L.P.; Massarin, I.D.S.; et al. Do not judge a book by its cover: Would Triatoma tibiamaculata (Pinto, 1926) belong to Triatoma Laporte, 1832, or to Panstrongylus Berg, 1879, with misleading homoplasies? Parasites Vectors 2022, 15, 184. [Google Scholar] [CrossRef]
- Pita, S.; Panzera, F.; Sánchez, A.; Panzera, Y.; Palomeque, T.; Lorite, P. Distribution and evolution of repeated sequences in genomes of Triatominae (Hemiptera-Reduviidae) inferred from Genomic In Situ Hybridization. PLoS ONE 2014, 9, e114298. [Google Scholar] [CrossRef]
- Kieran, T.J.; Gordon, E.R.L.; Zaldívar-Riverón, A.; Ibarra-Cerdeña, C.N.; Glenn, T.C.; Weirauch, C. Ultraconserved elements reconstruct the evolution of Chagas disease-vectoring kissing bugs (Reduviidae: Triatominae). Syst. Entomol. 2021, 46, 725–740. [Google Scholar] [CrossRef]
- Coyne, J.A.; Orr, H.A. Patterns of speciation in Drosophila. Evolution 1989, 43, 362–381. [Google Scholar] [CrossRef]
- Marcilla, A.; Bargues, M.D.; Ramsey, J.M.; Magallón-Gastelum, E.; Salazar-Schettino, P.M.; Abad-Franch, F.; Dujardin, J.P.; Schofield, C.J.; Mas-Coma, S. The ITS-2 of the nuclear rDNA as a molecular marker for populations, species, and phylogenetic relationships in Triatominae (Hemiptera: Reduviidae), vectors of Chagas disease. Mol. Phylog. Evol. 2001, 18, 136–142. [Google Scholar] [CrossRef]
- Monteiro, F.A.; Weirauch, C.; Felix, M.; Lazoski, C.; Abad-Franch, F. Evolution, systematics, and biogeography of the Triatominae, vectors of Chagas disease. Adv. Parasitol. 2018, 99, 265–344. [Google Scholar] [CrossRef]
- Pita, S.; Gómez-Palacio, A.; Lorite, P.; Dujardin, J.P.; Chavez, T.; Villacís, A.G.; Galvão, C.; Panzera, Y.; Calleros, L.; Pereyra-Mello, S.; et al. Multidisciplinary approach detects speciation within the kissing bug Panstrongylus rufotuberculatus populations (Hemiptera, Heteroptera, Reduviidae). Mem. Inst. Oswaldo Cruz 2021, 116, e210259. [Google Scholar] [CrossRef]
- Carcavallo, R.U.; Girón, I.G.; Jurberg, J.; Galvao, C.; Noireau, F.; Canale, D. Mutations, Hybrids and Teratologies. In Atlas of Chagas Disease Vectors in the Americas; Carcavallo, R.U., Girón, I.G., Jurberg, J., Lent, H., Eds.; Editora FIOCRUZ: Rio de Janeiro, Brazil, 1998; pp. 515–536. [Google Scholar]
- Paiva, V.F.; Oliveira, J.; Galvão, C.; Justi, S.A.; Landa, J.M.A.; Rosa, J.A. Formal assignation of the kissing bug Triatoma lecticularia (Hemiptera: Reduviidae: Triatominae) to the genus Paratriatoma. Insects 2021, 12, 538. [Google Scholar] [CrossRef]
- Galvão, C.; Carcavallo, R.; Rocha, D.D.S.; Jurberg, J. A checklist of the current valid species of the subfamily triatominae Jeannel, 1919 (Hemiptera, Reduviidae) and their geographical distribution, with nomenclatural and taxonomic notes. Zootaxa 2003, 202, 1–36. [Google Scholar] [CrossRef] [Green Version]
- Abbott, R.; Albach, D.; Ansell, S.; Arntzen, J.W.; Baird, S.J.E.; Bierne, N.; Boughman, J.; Brelsford, A.; Buerkle, C.A.; Buggs, R.; et al. Hybridization and speciation. J. Evol. Biol. 2013, 26, 229–246. [Google Scholar] [CrossRef] [Green Version]
- Filée, J.; Merle, M.; Bastide, H.; Mougel, F.; Bérenger, J.M.; Folly-Ramos, E.; Almeida, C.E.; Harry, M. phylogenomics for Chagas disease vectors of the Rhodnius genus (Hemiptera, Triatominae): What we learn from mito-nuclear conflicts and recommendations. Front. Ecol. Evol. 2022, 9, 750317. [Google Scholar] [CrossRef]
- Schreiber, G.; Pellegrino, J. Eteropicnosi di autosomi come possible meccanismo di speciazione. Sci. Genet. 1950, 3, 215–226. [Google Scholar]
- White, M.J.D. Chromosomal rearrangements and speciation in animals. Annu. Ver. Genet. 1969, 3, 75–98. [Google Scholar] [CrossRef]
- White, M.J.D. Animal Citology and Evolution, 3rd ed.; Cambridge University Press: Cambridge, UK, 1973; p. 961. [Google Scholar]
- White, M.J.D. Modes of Speciation; W. H. Freeman: San Francisco, CA, USA, 1978; p. 455. [Google Scholar]
- Noor, M.A.F.; Gratos, K.L.; Bertucci, L.A.; Reiland, J. Chromosomal inversions and the reproductive isolation of species. Proc. Natl. Acad. Sci. USA 2001, 98, 12084–12088. [Google Scholar] [CrossRef] [Green Version]
- Rieseberg, L.H. Chromosomal rearrangements and speciation. Trends Ecol. Evol. 2001, 16, 351–358. [Google Scholar] [CrossRef]
- Navarro, A.; Barton, N.H. Accumulating postzygotic isolation in parapatry: A new twist on chromosomal speciation. Evolution 2003, 57, 447–459. [Google Scholar] [CrossRef]
- Faria, R.; Navarro, A. Chromosomal speciation revisited: Rearranging theory with pieces of evidence. Trends Ecol. Evol. 2010, 25, 660–669. [Google Scholar] [CrossRef]
- Coyne, J.A.; Orr, H.A. Speciation; Sinauer Associates: Sunderland, MA, USA, 2004; p. 545. [Google Scholar]
- Lucek, K.; Augustijnen, H.; Escudero, M. A holocentric twist to chromosomal speciation? Trends Ecol. Evol. 2022, 37, 655–662. [Google Scholar] [CrossRef]
- Slijepcevic, P. Telomeres and mechanisms of Robertsonian fusion. Chromosoma 1998, 107, 136–140. [Google Scholar] [CrossRef]
- Escudero, M.; Hahn, M.; Brown, B.H.; Lueders, K.; Hipp, A.L. Chromosomal rearrangements in holocentric organisms lead to reproductive isolation by hybrid dysfunction: The correlation between karyotype rearrangements and germination rates in sedges. Am. J. Bot. 2016, 103, 1529–1536. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular evolutionary genetics analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [Green Version]
- Gouy, M.; Guindon, S.; Gascuel, O. SeaView version 4: A multiplatform graphical user interface for sequence alignment and phylogenetic tree building. Mol. Biol. Evol. 2010, 27, 221–224. [Google Scholar] [CrossRef] [Green Version]
- Drummond, A.J.; Suchard, M.A.; Xie, D.; Rambaut, A. Bayesian phylogenetics with BEAUti and the BEAST 1.7. Mol. Biol. Evol. 2012, 29, 1969–1973. [Google Scholar] [CrossRef] [Green Version]
- Yule, G.U., II. A mathematical theory of evolution, based on the conclusions of Dr. J. C. Willis, F.R.S. Phil. Trans. R. Soc. Lond. B. 1925, 213, 21–87. [Google Scholar] [CrossRef] [Green Version]
- Gernhard, T. The conditioned reconstructed process. J. Theor. Biol. 2008, 253, 769–778. [Google Scholar] [CrossRef] [Green Version]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior summarisation in Bayesian phylogenetics using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef] [Green Version]
- Hohna, S.; Landis, M.J.; Heath, T.A.; Boussau, B.; Lartillot, N.; Moore, B.R.; Huelsenbeck, J.P.; Ronquist, F. RevBayes: Bayesian phylogenetic inference using graphical models and an interactive model-specification language. Syst. Biol. 2016, 65, 726–736. [Google Scholar] [CrossRef] [Green Version]
- Poggio, M.G.; Provecho, Y.M.; Papeschi, A.G.; Bressa, M.J. Possible origin of polymorphism for chromosome number in the assassin bug Zelurus femoralis longispinis (Reduviidae: Reduviinae). Biol. J. Linn. Soc. 2013, 110, 757–764. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, J.; Alevi, K.C.C.; Gil-Santana, H.R.; Galvão, C. Biological, ecological, morphological and cytogenetic analyses, with taxonomic notes of Zelurus ochripennis (Stål, 1854) (Hemiptera: Heteroptera: Reduviidae: Reduviinae). Zootaxa 2021, 4958, 345–358. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. Available online: http://www.r-project.org. (accessed on 3 October 2022).
- Yu, G. Using Ggtree to visualize data on tree-like structures. Curr. Protoc. Bioinform. 2020, 69, e96. [Google Scholar] [CrossRef]
- Yu, G.; Lam, T.T.Y.; Zhu, H.; Guan, Y. Two methods for mapping and visualizing associated data on phylogeny using GGTree. Mol. Biol. Evol. 2018, 35, e96. [Google Scholar] [CrossRef]
- Yu, G. Data Integration, Manipulation and Visualization of Phylogenetic Trees, 1st ed.; Chapman and Hall/CRC: New York, NY, USA, 2022; p. 276. [Google Scholar] [CrossRef]
- Yu, G.; Smith, D.K.; Zhu, H.; Guan, Y.; Lam, T.T.Y. Ggtree: An R package for visualization and annotation of phylogenetic trees with their covariates and other associated data. Methods Ecol. Evol. 2017, 8, 28–36. [Google Scholar] [CrossRef]
- Xu, S.; Li, L.; Luo, X.; Chen, M.; Tang, W.; Zhan, L.; Dai, Z.; Lam, T.T.; Guan, Y.; Yu, G. Ggtree: A serialized data object for visualization of a phylogenetic tree and annotation data. iMeta 2022, 1, e56. [Google Scholar] [CrossRef]
- Tribble, C.M.; Freyman, W.A.; Landis, M.J.; Lim, J.Y.; Barido-Sottani, J.; Kopperud, B.T.; Hӧhna, S.; May, M.R. RevGadgets: An R package for visualizing bayesian phylogenetic analyses from RevBayes. Methods Ecol. Evol. 2022, 13, 314–323. [Google Scholar] [CrossRef]
- Olaia, N.; Alevi, K.C.C.; Oliveira, J.; Cacini, G.L.; Souza, E.D.S.; Pinotti, H.; Silva, L.A.; Rosa, J.A. Biology of Chagas disease vectors: Biological cycle and emergence rates of Rhodnius marabaensis Souza et al., 2016 (Hemiptera, Reduviidae, Triatominae) under laboratory conditions. Parasitol. Res. 2021, 120, 2939–2945. [Google Scholar] [CrossRef]


{kind=link}
| Experimental Crosses | Number of Eggs | Hatching Rate (%) | Reference | ||
|---|---|---|---|---|---|
| Female (CN) | x | Male (CN) | |||
| Interspecific cross | |||||
| T. longipennis (23) | x | T. vitticeps (24) | 26 | 0 | This paper |
| T. vitticeps (24) | x | T. longipennis (23) | 72 | 0 | This paper |
| T. longipennis (23) | x | T. infestans (22) | 38 | 0 | This paper |
| T. infestans (22) | x | T. longipennis (23) | 45 | 0 | This paper |
| T. infestans (22) | x | T. protracta (23) | 146 | 0 | This paper |
| T. protracta (23) | x | T. infestans (22) | 93 | 0 | This paper |
| T. protracta (23) | x | P. lecticularia (22) | 160 | 0 | This paper |
| P. lecticularia (22) | x | T. protracta (23) | 82 | 0 | This paper |
| P. tibiamaculatus (23) | x | T. brasiliensis (22) | 60 | 0 | This paper |
| T. brasiliensis (22) | x | P. tibiamaculatus (23) | 193 | 0 | This paper |
| T. pseudomaculata (22) | x | P. tibiamaculatus (23) | 150 | 0 | This paper |
| P. tibiamaculatus (23) | x | T. pseudomaculata (22) | 102 | 0 | This paper |
| T. melanocephala (24) | x | P. tibiamaculatus (23) | 102 | 0 | This paper |
| P. tibiamaculatus (23) | x | T. melanocephala (24) | 237 | 0 | This paper |
| T. rubrovaria (22) | x | P. tibiamaculatus (23) | 21 | 0 | This paper |
| P. tibiamaculatus (23) | x | T. rubrovaria (22) | 53 | 0 | This paper |
| T. infestans (22) | x | P. tibiamaculatus (23) | 174 | 0 | This paper |
| P. tibiamaculatus (23) | x | T. infestans (22) | 93 | 0 | This paper |
| T. brasiliensis (22) | x | T. vitticeps (24) | 90 | 0 | [18] |
| T. vitticeps (24) | x | T. brasiliensis (22) | 147 | 0 | [18] |
| T. melanocephala (24) | x | T. brasiliensis (22) | 78 | 0 | [18] |
| T. brasiliensis (22) | x | T. melanocephala (24) | 63 | 0 | [18] |
| P. megistus (21) | x | P. tibiamaculatus (23) | 107 | 0 | [32] |
| P. tibiamaculatus (23) | x | P. megistus (21) | 265 | 0 | [32] |
| P. megistus (21) | x | P. lignarius (23) | 157 | 0 | [32] |
| P. lignarius (23) | x | P. megistus (21) | 523 | 0 | [32] |
| Intraspecific cross | |||||
| T. infestans | x | T. infestans | 439 | 62 | This paper |
| T. protracta | x | T. protracta | 278 | 86 | This paper |
| P. lignarius | x | P. lignarius | 700 | 51 | [32] |
| P. megistus | x | P. megistus | 372 | 68 | [32] |
| P. tibiamaculatus | x | P. tibiamaculatus | 190 | 65 | [18] |
| T. brasiliensis | x | T. brasiliensis | 271 | 59 | [18] |
| T. melanocephala | x | T. melanocephala | 302 | 63 | [18] |
| T. vitticeps | x | T. vitticeps | 353 | 70 | [18] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reis, Y.V.d.; de Oliveira, J.; Madeira, F.F.; Ravazi, A.; Oliveira, A.B.B.d.; Bittinelli, I.d.S.; Delgado, L.M.G.; de Azeredo-Oliveira, M.T.V.; Rosa, J.A.d.; Galvão, C.; et al. Karyotype Evolution in Triatominae (Hemiptera, Reduviidae): The Role of Chromosomal Rearrangements in the Diversification of Chagas Disease Vectors. Int. J. Mol. Sci. 2023, 24, 6350. https://doi.org/10.3390/ijms24076350
Reis YVd, de Oliveira J, Madeira FF, Ravazi A, Oliveira ABBd, Bittinelli IdS, Delgado LMG, de Azeredo-Oliveira MTV, Rosa JAd, Galvão C, et al. Karyotype Evolution in Triatominae (Hemiptera, Reduviidae): The Role of Chromosomal Rearrangements in the Diversification of Chagas Disease Vectors. International Journal of Molecular Sciences. 2023; 24(7):6350. https://doi.org/10.3390/ijms24076350
Chicago/Turabian StyleReis, Yago Visinho dos, Jader de Oliveira, Fernanda Fernandez Madeira, Amanda Ravazi, Ana Beatriz Bortolozo de Oliveira, Isadora da Silva Bittinelli, Luiza Maria Grzyb Delgado, Maria Tercília Vilela de Azeredo-Oliveira, João Aristeu da Rosa, Cleber Galvão, and et al. 2023. "Karyotype Evolution in Triatominae (Hemiptera, Reduviidae): The Role of Chromosomal Rearrangements in the Diversification of Chagas Disease Vectors" International Journal of Molecular Sciences 24, no. 7: 6350. https://doi.org/10.3390/ijms24076350