
Aging-Related Mechanisms Contribute to Corticosteroid Insensitivity in Elderly Asthma


Abstract
:1. Introduction
2. Glucocorticoid Receptor Signaling and Hormones in the Aging
3. Asthma in the Elderly Pathogenesis
3.1. Airway Inflammation
3.2. Airway Structure and Function
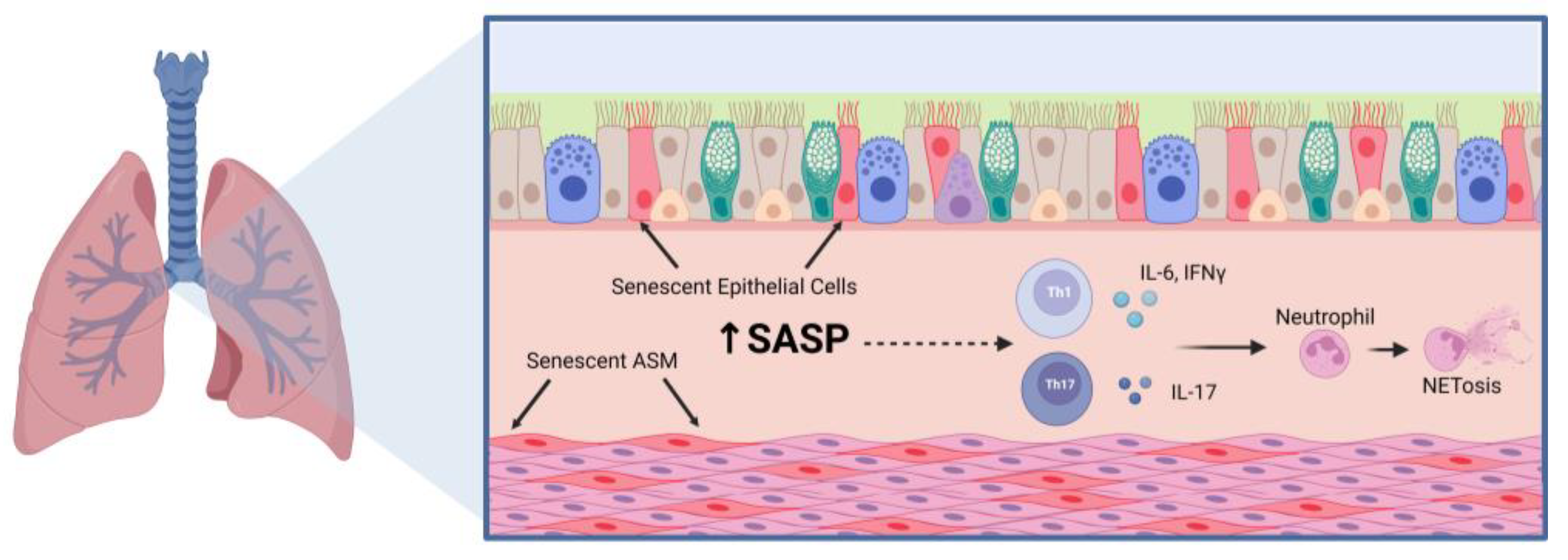
4. Aging-Related Mechanisms in Asthma
4.1. Cellular Senescence
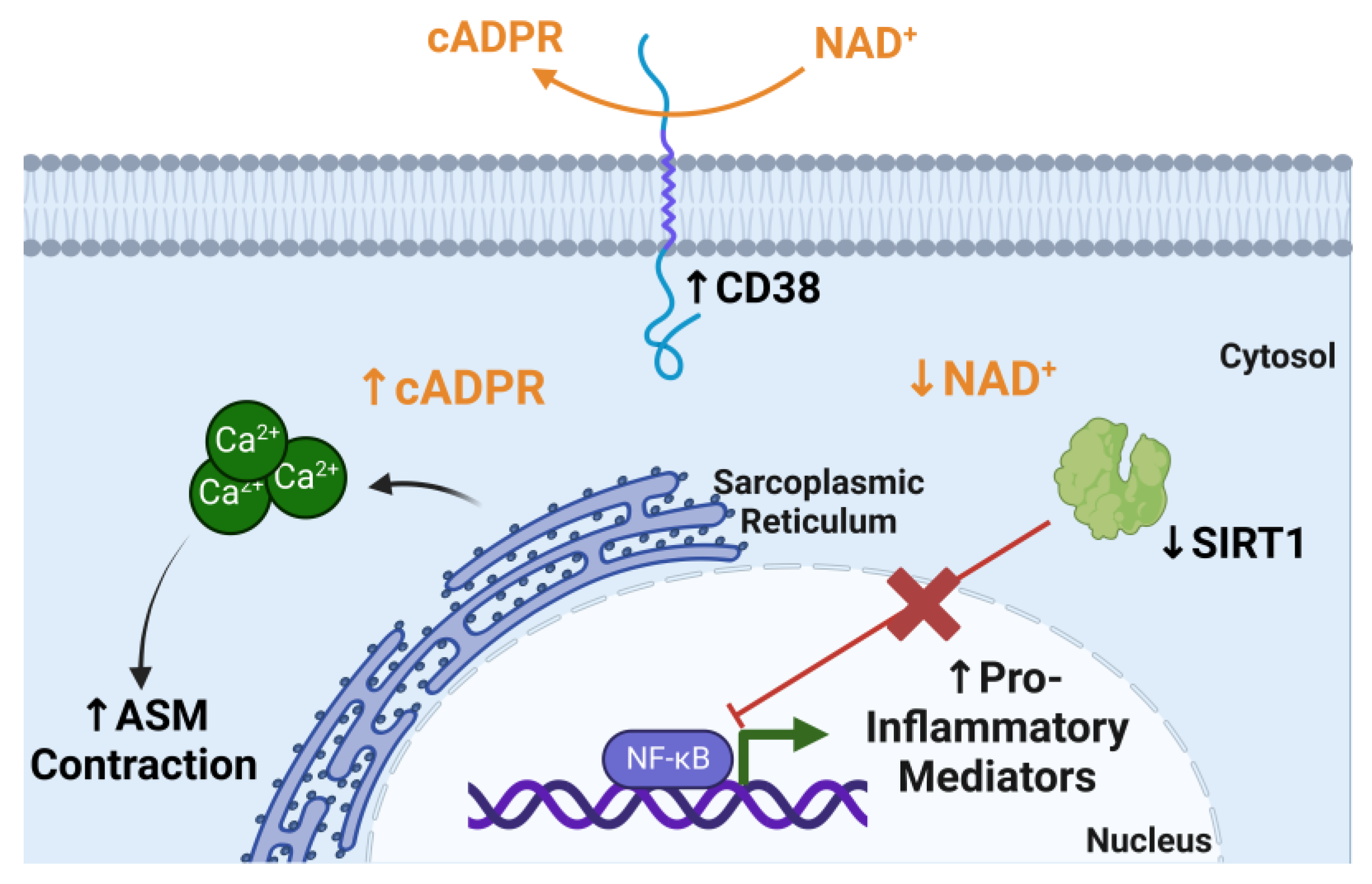
4.2. CD38 and NAD+ Metabolism
5. Factors and Comorbidities That May Influence Asthma in the Elderly
6. Conclusions

{kind=link}
{kind=link}
| Mechanisms | Characteristics | References |
|---|---|---|
| Hormone Production |
| [25,26,27,28,29] |
| Immune Cell Infiltration |
| [38,45,48] |
| Lung Function |
| [82,108,109] |
| Cellular Senescence |
| [39,52,59,63,64] |
| NAD+ Metabolism |
| [68,69,79,80] |
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Thannickal, V.J.; Murthy, M.; Balch, W.E.; Chandel, N.S.; Meiners, S.; Eickelberg, O.; Selman, M.; Pardo, A.; White, E.S.; Levy, B.D.; et al. Blue journal conference. Aging and susceptibility to lung disease. Am. J. Respir. Crit. Care Med. 2015, 191, 261–269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, E.T.; Guppy, M.; Straus, S.E.; Bell, K.J.L.; Glasziou, P. Rate of normal lung function decline in ageing adults: A systematic review of prospective cohort studies. BMJ Open 2019, 9, e028150. [Google Scholar] [CrossRef] [PubMed]
- Dunn, R.M.; Busse, P.J.; Wechsler, M.E. Asthma in the elderly and late-onset adult asthma. Allergy 2018, 73, 284–294. [Google Scholar] [CrossRef] [PubMed]
- Boulet, L.P. Irreversible airway obstruction in asthma. Curr. Allergy Asthma Rep. 2009, 9, 168–173. [Google Scholar] [CrossRef]
- Skloot, G.S.; Busse, P.J.; Braman, S.S.; Kovacs, E.J.; Dixon, A.E.; Vaz Fragoso, C.A.; Scichilone, N.; Prakash, Y.S.; Pabelick, C.M.; Mathur, S.K.; et al. An Official American Thoracic Society Workshop Report: Evaluation and Management of Asthma in the Elderly. Ann. Am. Thorac. Soc. 2016, 13, 2064–2077. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez-Garcia, M.; Caballero, A.; Jaramillo, C.; Maldonado, D.; Torres-Duque, C.A. Prevalence, risk factors and underdiagnosis of asthma and wheezing in adults 40 years and older: A population-based study. J. Asthma 2015, 52, 823–830. [Google Scholar] [CrossRef]
- Tsai, C.L.; Delclos, G.L.; Huang, J.S.; Hanania, N.A.; Camargo, C.A., Jr. Age-related differences in asthma outcomes in the United States, 1988–2006. Ann. Allergy Asthma Immunol. 2013, 110, 240–246.e1. [Google Scholar] [CrossRef]
- Ray, A.; Camiolo, M.; Fitzpatrick, A.; Gauthier, M.; Wenzel, S.E. Are We Meeting the Promise of Endotypes and Precision Medicine in Asthma? Physiol. Rev. 2020, 100, 983–1017. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, X.; Zhang, L.; Liu, Y.; Wang, G.; Zhang, H.P.; Wang, L.; Kang, Y.; Oliver, B.G.; Wan, H.J.; et al. Age-Related Clinical Characteristics, Inflammatory Features, Phenotypes, and Treatment Response in Asthma. J. Allergy Clin. Immunol. Pract. 2023, 11, 210–219.e3. [Google Scholar] [CrossRef]
- Choy, D.F.; Hart, K.M.; Borthwick, L.A.; Shikotra, A.; Nagarkar, D.R.; Siddiqui, S.; Jia, G.; Ohri, C.M.; Doran, E.; Vannella, K.M.; et al. TH2 and TH17 inflammatory pathways are reciprocally regulated in asthma. Sci. Transl. Med. 2015, 7, 301ra129. [Google Scholar] [CrossRef] [Green Version]
- Raundhal, M.; Morse, C.; Khare, A.; Oriss, T.B.; Milosevic, J.; Trudeau, J.; Huff, R.; Pilewski, J.; Holguin, F.; Kolls, J.; et al. High IFN-gamma and low SLPI mark severe asthma in mice and humans. J. Clin. Investig. 2015, 125, 3037–3050. [Google Scholar] [CrossRef] [Green Version]
- Isoyama, S.; Ishikawa, N.; Hamai, K.; Matsumura, M.; Kobayashi, H.; Nomura, A.; Ueno, S.; Tanimoto, T.; Maeda, H.; Iwamoto, H.; et al. Efficacy of mepolizumab in elderly patients with severe asthma and overlapping COPD in real-world settings: A retrospective observational study. Respir. Investig. 2021, 59, 478–486. [Google Scholar] [CrossRef]
- Patruno, C.; Napolitano, M.; Argenziano, G.; Peris, K.; Ortoncelli, M.; Girolomoni, G.; Offidani, A.; Ferrucci, S.M.; Amoruso, G.F.; Rossi, M.; et al. Dupilumab therapy of atopic dermatitis of the elderly: A multicentre, real-life study. J. Eur. Acad. Dermatol. Venereol. 2021, 35, 958–964. [Google Scholar] [CrossRef] [PubMed]
- Hinks, T.S.C.; Levine, S.J.; Brusselle, G.G. Treatment options in type-2 low asthma. Eur. Respir. J. 2021, 57, 2000528. [Google Scholar] [CrossRef] [PubMed]
- Desmet, S.J.; De Bosscher, K. Glucocorticoid receptors: Finding the middle ground. J. Clin. Investig. 2017, 127, 1136–1145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouazza, B.; Krytska, K.; Debba-Pavard, M.; Amrani, Y.; Honkanen, R.E.; Tran, J.; Tliba, O. Cytokines alter glucocorticoid receptor phosphorylation in airway cells: Role of phosphatases. Am. J. Respir. Cell Mol. Biol. 2012, 47, 464–473. [Google Scholar] [CrossRef] [Green Version]
- Chang, P.J.; Michaeloudes, C.; Zhu, J.; Shaikh, N.; Baker, J.; Chung, K.F.; Bhavsar, P.K. Impaired nuclear translocation of the glucocorticoid receptor in corticosteroid-insensitive airway smooth muscle in severe asthma. Am. J. Respir. Crit. Care Med. 2015, 191, 54–62. [Google Scholar] [CrossRef]
- Hamid, Q.A.; Wenzel, S.E.; Hauk, P.J.; Tsicopoulos, A.; Wallaert, B.; Lafitte, J.J.; Chrousos, G.P.; Szefler, S.J.; Leung, D.Y. Increased glucocorticoid receptor beta in airway cells of glucocorticoid-insensitive asthma. Am. J. Respir. Crit. Care Med. 1999, 159, 1600–1604. [Google Scholar] [CrossRef]
- Matthews, J.G.; Ito, K.; Barnes, P.J.; Adcock, I.M. Defective glucocorticoid receptor nuclear translocation and altered histone acetylation patterns in glucocorticoid-resistant patients. J. Allergy Clin. Immunol. 2004, 113, 1100–1108. [Google Scholar] [CrossRef]
- Veldhuis, J.D.; Sharma, A.; Roelfsema, F. Age-dependent and gender-dependent regulation of hypothalamic-adrenocorticotropic-adrenal axis. Endocrinol. Metab. Clin. N. Am. 2013, 42, 201–225. [Google Scholar] [CrossRef] [Green Version]
- Cho, Y.J.; Lee, K.E. Decreased glucocorticoid binding affinity to glucocorticoid receptor is important in the poor response to steroid therapy of older-aged patients with severe bronchial asthma. Allergy Asthma Proc. 2003, 24, 353–358. [Google Scholar] [PubMed]
- Nimmagadda, S.R.; Szefler, S.J.; Spahn, J.D.; Surs, W.; Leung, D.Y. Allergen exposure decreases glucocorticoid receptor binding affinity and steroid responsiveness in atopic asthmatics. Am. J. Respir. Crit. Care Med. 1997, 155, 87–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lightman, S.L.; Birnie, M.T.; Conway-Campbell, B.L. Dynamics of ACTH and Cortisol Secretion and Implications for Disease. Endocr. Rev. 2020, 41, bnaa002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dickmeis, T.; Weger, B.D.; Weger, M. The circadian clock and glucocorticoids--interactions across many time scales. Mol. Cell Endocrinol. 2013, 380, 2–15. [Google Scholar] [CrossRef] [PubMed]
- Paragliola, R.M.; Papi, G.; Pontecorvi, A.; Corsello, S.M. Treatment with Synthetic Glucocorticoids and the Hypothalamus-Pituitary-Adrenal Axis. Int. J. Mol. Sci. 2017, 18, 2201. [Google Scholar] [CrossRef] [Green Version]
- Giri, A.; Wang, Q.; Rahman, I.; Sundar, I.K. Circadian molecular clock disruption in chronic pulmonary diseases. Trends Mol. Med. 2022, 28, 513–527. [Google Scholar] [CrossRef]
- Landstra, A.M.; Postma, D.S.; Boezen, H.M.; van Aalderen, W.M. Role of serum cortisol levels in children with asthma. Am. J. Respir. Crit. Care Med. 2002, 165, 708–712. [Google Scholar] [CrossRef]
- Kachroo, P.; Stewart, I.D.; Kelly, R.S.; Stav, M.; Mendez, K.; Dahlin, A.; Soeteman, D.I.; Chu, S.H.; Huang, M.; Cote, M.; et al. Metabolomic profiling reveals extensive adrenal suppression due to inhaled corticosteroid therapy in asthma. Nat. Med. 2022, 28, 814–822. [Google Scholar] [CrossRef]
- Seale, J.P. Is the pharmacology of corticosteroids in the lung modified by age? Med. J. Aust. 2005, 183, S47–S48. [Google Scholar] [CrossRef]
- Vermeulen, A. Dehydroepiandrosterone sulfate and aging. Ann. N. Y. Acad. Sci. 1995, 774, 121–127. [Google Scholar] [CrossRef]
- Yu, C.K.; Yang, B.C.; Lei, H.Y.; Chen, Y.C.; Liu, Y.H.; Chen, C.C.; Liu, C.W. Attenuation of house dust mite Dermatophagoides farinae-induced airway allergic responses in mice by dehydroepiandrosterone is correlated with down-regulation of TH2 response. Clin. Exp. Allergy 1999, 29, 414–422. [Google Scholar] [CrossRef]
- Dashtaki, R.; Whorton, A.R.; Murphy, T.M.; Chitano, P.; Reed, W.; Kennedy, T.P. Dehydroepiandrosterone and analogs inhibit DNA binding of AP-1 and airway smooth muscle proliferation. J. Pharmacol. Exp. Ther. 1998, 285, 876–883. [Google Scholar] [PubMed]
- Gandhi, V.D.; Cephus, J.Y.; Norlander, A.E.; Chowdhury, N.U.; Zhang, J.; Ceneviva, Z.J.; Tannous, E.; Polosukhin, V.V.; Putz, N.D.; Wickersham, N.; et al. Androgen receptor signaling promotes Treg suppressive function during allergic airway inflammation. J. Clin. Investig. 2022, 132, e153397. [Google Scholar] [CrossRef] [PubMed]
- Kalidhindi, R.S.R.; Ambhore, N.S.; Balraj, P.; Schmidt, T.; Khan, M.N.; Sathish, V. Androgen receptor activation alleviates airway hyperresponsiveness, inflammation, and remodeling in a murine model of asthma. Am. J. Physiol. Lung Cell. Mol. Physiol. 2021, 320, L803–L818. [Google Scholar] [CrossRef] [PubMed]
- Zein, J.G.; McManus, J.M.; Sharifi, N.; Erzurum, S.C.; Marozkina, N.; Lahm, T.; Giddings, O.; Davis, M.D.; DeBoer, M.D.; Comhair, S.A.; et al. Benefits of Airway Androgen Receptor Expression in Human Asthma. Am. J. Respir. Crit. Care Med. 2021, 204, 285–293. [Google Scholar] [CrossRef] [PubMed]
- Zein, J.; Gaston, B.; Bazeley, P.; DeBoer, M.D.; Igo, R.P., Jr.; Bleecker, E.R.; Meyers, D.; Comhair, S.; Marozkina, N.V.; Cotton, C.; et al. HSD3B1 genotype identifies glucocorticoid responsiveness in severe asthma. Proc. Natl. Acad. Sci. USA 2020, 117, 2187–2193. [Google Scholar] [CrossRef] [Green Version]
- Busse, P.J.; Birmingham, J.M.; Calatroni, A.; Manzi, J.; Goryachokovsky, A.; Fontela, G.; Federman, A.D.; Wisnivesky, J.P. Effect of aging on sputum inflammation and asthma control. J. Allergy Clin. Immunol. 2017, 139, 1808–1818.e6. [Google Scholar] [CrossRef] [Green Version]
- Ducharme, M.E.; Prince, P.; Hassan, N.; Nair, P.; Boulet, L.P. Expiratory flows and airway inflammation in elderly asthmatic patients. Respir. Med. 2011, 105, 1284–1289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaiswal, A.K.; Yadav, J.; Makhija, S.; Sandey, M.; Suryawanshi, A.; Mitra, A.K.; Mishra, A. Short palate, lung, and nasal epithelial clone 1 (SPLUNC1) level determines steroid-resistant airway inflammation in aging. Am. J. Physiol. Lung Cell. Mol. Physiol. 2022, 322, L102–L115. [Google Scholar] [CrossRef]
- Birmingham, J.M.; Gillespie, V.L.; Srivastava, K.; Li, X.M.; Busse, P.J. Influenza A infection enhances antigen-induced airway inflammation and hyperresponsiveness in young but not aged mice. Clin. Exp. Allergy 2014, 44, 1188–1199. [Google Scholar] [CrossRef] [Green Version]
- Brandenberger, C.; Li, N.; Jackson-Humbles, D.N.; Rockwell, C.E.; Wagner, J.G.; Harkema, J.R. Enhanced allergic airway disease in old mice is associated with a Th17 response. Clin. Exp. Allergy 2014, 44, 1282–1292. [Google Scholar] [CrossRef]
- Busse, P.J.; Zhang, T.F.; Srivastava, K.; Schofield, B.; Li, X.M. Effect of ageing on pulmonary inflammation, airway hyperresponsiveness and T and B cell responses in antigen-sensitized and -challenged mice. Clin. Exp. Allergy 2007, 37, 1392–1403. [Google Scholar] [CrossRef] [PubMed]
- Alcorn, J.F.; Crowe, C.R.; Kolls, J.K. TH17 cells in asthma and COPD. Annu. Rev. Physiol. 2010, 72, 495–516. [Google Scholar] [CrossRef] [PubMed]
- Cai, C.W.; Eickhoff, C.S.; Meza, K.A.; Blase, J.R.; Audette, R.E.; Chan, D.H.; Bockerstett, K.A.; DiPaolo, R.J.; Hoft, D.F. Th17 Cells Provide Mucosal Protection against Gastric Trypanosoma cruzi Infection. Infect. Immun. 2021, 89, e0073820. [Google Scholar] [CrossRef]
- Zhang, D.; Chen, G.; Manwani, D.; Mortha, A.; Xu, C.; Faith, J.J.; Burk, R.D.; Kunisaki, Y.; Jang, J.E.; Scheiermann, C.; et al. Neutrophil ageing is regulated by the microbiome. Nature 2015, 525, 528–532. [Google Scholar] [CrossRef] [Green Version]
- Green, R.H.; Brightling, C.E.; Woltmann, G.; Parker, D.; Wardlaw, A.J.; Pavord, I.D. Analysis of induced sputum in adults with asthma: Identification of subgroup with isolated sputum neutrophilia and poor response to inhaled corticosteroids. Thorax 2002, 57, 875–879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nyenhuis, S.M.; Schwantes, E.A.; Evans, M.D.; Mathur, S.K. Airway neutrophil inflammatory phenotype in older subjects with asthma. J. Allergy Clin. Immunol. 2010, 125, 1163–1165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mathur, S.K.; Schwantes, E.A.; Jarjour, N.N.; Busse, W.W. Age-related changes in eosinophil function in human subjects. Chest 2008, 133, 412–419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Childs, B.G.; Durik, M.; Baker, D.J.; van Deursen, J.M. Cellular senescence in aging and age-related disease: From mechanisms to therapy. Nat. Med. 2015, 21, 1424–1435. [Google Scholar] [CrossRef] [Green Version]
- Munoz-Espin, D.; Canamero, M.; Maraver, A.; Gomez-Lopez, G.; Contreras, J.; Murillo-Cuesta, S.; Rodriguez-Baeza, A.; Varela-Nieto, I.; Ruberte, J.; Collado, M.; et al. Programmed cell senescence during mammalian embryonic development. Cell 2013, 155, 1104–1118. [Google Scholar] [CrossRef] [Green Version]
- Storer, M.; Mas, A.; Robert-Moreno, A.; Pecoraro, M.; Ortells, M.C.; Di Giacomo, V.; Yosef, R.; Pilpel, N.; Krizhanovsky, V.; Sharpe, J.; et al. Senescence is a developmental mechanism that contributes to embryonic growth and patterning. Cell 2013, 155, 1119–1130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Acosta, J.C.; Banito, A.; Wuestefeld, T.; Georgilis, A.; Janich, P.; Morton, J.P.; Athineos, D.; Kang, T.W.; Lasitschka, F.; Andrulis, M.; et al. A complex secretory program orchestrated by the inflammasome controls paracrine senescence. Nat. Cell Biol. 2013, 15, 978–990. [Google Scholar] [CrossRef]
- Coppe, J.P.; Patil, C.K.; Rodier, F.; Sun, Y.; Munoz, D.P.; Goldstein, J.; Nelson, P.S.; Desprez, P.Y.; Campisi, J. Senescence-associated secretory phenotypes reveal cell-nonautonomous functions of oncogenic RAS and the p53 tumor suppressor. PLoS Biol. 2008, 6, e301. [Google Scholar] [CrossRef]
- Schafer, M.J.; White, T.A.; Iijima, K.; Haak, A.J.; Ligresti, G.; Atkinson, E.J.; Oberg, A.L.; Birch, J.; Salmonowicz, H.; Zhu, Y.; et al. Cellular senescence mediates fibrotic pulmonary disease. Nat. Commun. 2017, 8, 14532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yao, C.; Guan, X.; Carraro, G.; Parimon, T.; Liu, X.; Huang, G.; Mulay, A.; Soukiasian, H.J.; David, G.; Weigt, S.S.; et al. Senescence of Alveolar Type 2 Cells Drives Progressive Pulmonary Fibrosis. Am. J. Respir. Crit. Care Med. 2021, 203, 707–717. [Google Scholar] [CrossRef]
- Prakash, Y.S. Emerging concepts in smooth muscle contributions to airway structure and function: Implications for health and disease. Am. J. Physiol. Lung Cell. Mol. Physiol. 2016, 311, L1113–L1140. [Google Scholar] [CrossRef] [Green Version]
- Parikh, P.; Britt, R.D., Jr.; Manlove, L.J.; Wicher, S.A.; Roesler, A.; Ravix, J.; Teske, J.; Thompson, M.A.; Sieck, G.C.; Kirkland, J.L.; et al. Hyperoxia-induced Cellular Senescence in Fetal Airway Smooth Muscle Cells. Am. J. Respir. Cell Mol. Biol. 2019, 61, 51–60. [Google Scholar] [CrossRef]
- Wang, Z.N.; Su, R.N.; Yang, B.Y.; Yang, K.X.; Yang, L.F.; Yan, Y.; Chen, Z.G. Potential Role of Cellular Senescence in Asthma. Front. Cell Dev. Biol. 2020, 8, 59. [Google Scholar] [CrossRef] [PubMed]
- Aghali, A.; Khalfaoui, L.; Lagnado, A.B.; Drake, L.Y.; Teske, J.J.; Pabelick, C.M.; Passos, J.F.; Prakash, Y.S. Cellular senescence is increased in airway smooth muscle cells of elderly persons with asthma. Am. J. Physiol. Lung Cell. Mol. Physiol. 2022, 323, L558–L568. [Google Scholar] [CrossRef] [PubMed]
- Wicher, S.A.; Roos, B.B.; Teske, J.J.; Fang, Y.H.; Pabelick, C.; Prakash, Y.S. Aging increases senescence, calcium signaling, and extracellular matrix deposition in human airway smooth muscle. PLoS ONE 2021, 16, e0254710. [Google Scholar] [CrossRef]
- Lambrecht, B.N.; Hammad, H. The airway epithelium in asthma. Nat. Med. 2012, 18, 684–692. [Google Scholar] [CrossRef] [PubMed]
- de Vries, M.; Nwozor, K.O.; Muizer, K.; Wisman, M.; Timens, W.; van den Berge, M.; Faiz, A.; Hackett, T.L.; Heijink, I.H.; Brandsma, C.A. The relation between age and airway epithelial barrier function. Respir. Res. 2022, 23, 43. [Google Scholar] [CrossRef] [PubMed]
- Yuan, L.; Du, X.; Tang, S.; Wu, S.; Wang, L.; Xiang, Y.; Qu, X.; Liu, H.; Qin, X.; Liu, C. ITGB4 deficiency induces senescence of airway epithelial cells through p53 activation. FEBS J. 2019, 286, 1191–1203. [Google Scholar] [CrossRef] [PubMed]
- Yuan, L.; Liu, H.; Du, X.; Yao, Y.; Qin, L.; Xia, Z.; Zhou, K.; Wu, X.; Yuan, Y.; Qing, B.; et al. Airway epithelial ITGB4 deficiency induces airway remodeling in a mouse model. J. Allergy Clin. Immunol. 2023, 151, 431–446.e16. [Google Scholar] [CrossRef]
- Hachim, M.Y.; Elemam, N.M.; Ramakrishnan, R.K.; Bajbouj, K.; Olivenstein, R.; Hachim, I.Y.; Al Heialy, S.; Hamid, Q.; Busch, H.; Hamoudi, R. Wnt Signaling Is Deranged in Asthmatic Bronchial Epithelium and Fibroblasts. Front. Cell Dev. Biol. 2021, 9, 641404. [Google Scholar] [CrossRef]
- Hasegawa, A.; Miki, T.; Hosokawa, H.; Hossain, M.B.; Shimizu, C.; Hashimoto, K.; Kimura, M.Y.; Yamashita, M.; Nakayama, T. Impaired GATA3-dependent chromatin remodeling and Th2 cell differentiation leading to attenuated allergic airway inflammation in aging mice. J. Immunol. 2006, 176, 2546–2554. [Google Scholar] [CrossRef] [Green Version]
- Piedra-Quintero, Z.L.; Wilson, Z.; Nava, P.; Guerau-de-Arellano, M. CD38: An Immunomodulatory Molecule in Inflammation and Autoimmunity. Front. Immunol. 2020, 11, 597959. [Google Scholar] [CrossRef]
- Camacho-Pereira, J.; Tarrago, M.G.; Chini, C.C.S.; Nin, V.; Escande, C.; Warner, G.M.; Puranik, A.S.; Schoon, R.A.; Reid, J.M.; Galina, A.; et al. CD38 Dictates Age-Related NAD Decline and Mitochondrial Dysfunction through an SIRT3-Dependent Mechanism. Cell Metab. 2016, 23, 1127–1139. [Google Scholar] [CrossRef] [Green Version]
- Chini, C.C.S.; Tarrago, M.G.; Chini, E.N. NAD and the aging process: Role in life, death and everything in between. Mol. Cell. Endocrinol. 2017, 455, 62–74. [Google Scholar] [CrossRef]
- Deshpande, D.A.; Guedes, A.G.P.; Lund, F.E.; Subramanian, S.; Walseth, T.F.; Kannan, M.S. CD38 in the pathogenesis of allergic airway disease: Potential therapeutic targets. Pharmacol. Ther. 2017, 172, 116–126. [Google Scholar] [CrossRef] [Green Version]
- Tliba, O.; Cidlowski, J.A.; Amrani, Y. CD38 expression is insensitive to steroid action in cells treated with tumor necrosis factor-alpha and interferon-gamma by a mechanism involving the up-regulation of the glucocorticoid receptor beta isoform. Mol. Pharmacol. 2006, 69, 588–596. [Google Scholar] [CrossRef] [PubMed]
- Jackson, D.; Walum, J.; Banerjee, P.; Lewis, B.W.; Prakash, Y.S.; Sathish, V.; Xu, Z.; Britt, R.D., Jr. Th1 cytokines synergize to change gene expression and promote corticosteroid insensitivity in pediatric airway smooth muscle. Respir. Res. 2022, 23, 126. [Google Scholar] [CrossRef] [PubMed]
- Guedes, A.G.; Jude, J.A.; Paulin, J.; Kita, H.; Lund, F.E.; Kannan, M.S. Role of CD38 in TNF-alpha-induced airway hyperresponsiveness. Am. J. Physiol. Lung Cell. Mol. Physiol. 2008, 294, L290–L299. [Google Scholar] [CrossRef]
- Guedes, A.G.; Paulin, J.; Rivero-Nava, L.; Kita, H.; Lund, F.E.; Kannan, M.S. CD38-deficient mice have reduced airway hyperresponsiveness following IL-13 challenge. Am. J. Physiol. Lung Cell Mol. Physiol. 2006, 291, L1286–L1293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guedes, A.G.; Jude, J.A.; Paulin, J.; Rivero-Nava, L.; Kita, H.; Lund, F.E.; Kannan, M.S. Airway responsiveness in CD38-deficient mice in allergic airway disease: Studies with bone marrow chimeras. Am. J. Physiol. Lung Cell. Mol. Physiol. 2015, 308, L485–L493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gally, F.; Hartney, J.M.; Janssen, W.J.; Perraud, A.L. CD38 plays a dual role in allergen-induced airway hyperresponsiveness. Am. J. Respir. Cell Mol. Biol. 2009, 40, 433–442. [Google Scholar] [CrossRef] [Green Version]
- Cui, H.; Xie, N.; Banerjee, S.; Dey, T.; Liu, R.M.; Antony, V.B.; Sanders, Y.Y.; Adams, T.S.; Gomez, J.L.; Thannickal, V.J.; et al. CD38 Mediates Lung Fibrosis by Promoting Alveolar Epithelial Cell Aging. Am. J. Respir. Crit. Care Med. 2022, 206, 459–475. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Shi, G. Roles of sirtuins in asthma. Respir. Res. 2022, 23, 251. [Google Scholar] [CrossRef]
- Rahman, I.; Kinnula, V.L.; Gorbunova, V.; Yao, H. SIRT1 as a therapeutic target in inflammaging of the pulmonary disease. Prev. Med. 2012, 54, S20–S28. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Sun, Y.; Rong, W.; Fan, L.; Cai, Y.; Qu, Q.; Gao, Y.; Zhao, H. miR-221 participates in the airway epithelial cells injury in asthma via targeting SIRT1. Exp. Lung Res. 2018, 44, 272–279. [Google Scholar] [CrossRef]
- Pal, S.; Tyler, J.K. Epigenetics and aging. Sci. Adv. 2016, 2, e1600584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ciprandi, G.; Schiavetti, I.; Ricciardolo, F.L.M. The impact of aging on outpatients with asthma in a real-world setting. Respir. Med. 2018, 136, 58–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanania, N.A.; King, M.J.; Braman, S.S.; Saltoun, C.; Wise, R.A.; Enright, P.; Falsey, A.R.; Mathur, S.K.; Ramsdell, J.W.; Rogers, L.; et al. Asthma in the elderly: Current understanding and future research needs—A report of a National Institute on Aging (NIA) workshop. J. Allergy Clin. Immunol. 2011, 128, S4-24. [Google Scholar] [CrossRef] [PubMed]
- Hekking, P.P.; Amelink, M.; Wener, R.R.; Bouvy, M.L.; Bel, E.H. Comorbidities in Difficult-to-Control Asthma. J. Allergy Clin. Immunol. Pract. 2018, 6, 108–113. [Google Scholar] [CrossRef] [PubMed]
- Wee, J.H.; Park, M.W.; Min, C.; Byun, S.H.; Park, B.; Choi, H.G. Association between asthma and cardiovascular disease. Eur. J. Clin. Investig. 2021, 51, e13396. [Google Scholar] [CrossRef]
- Tattersall, M.C.; Guo, M.; Korcarz, C.E.; Gepner, A.D.; Kaufman, J.D.; Liu, K.J.; Barr, R.G.; Donohue, K.M.; McClelland, R.L.; Delaney, J.A.; et al. Asthma predicts cardiovascular disease events: The multi-ethnic study of atherosclerosis. Arter. Thromb. Vasc. Biol. 2015, 35, 1520–1525. [Google Scholar] [CrossRef] [Green Version]
- Schatz, M.; Zeiger, R.S.; Yang, S.J.; Chen, W.; Sajjan, S.; Allen-Ramey, F.; Camargo, C.A., Jr. Prospective Study on the Relationship of Obesity to Asthma Impairment and Risk. J. Allergy Clin. Immunol. Pract. 2015, 3, 560–565.e1. [Google Scholar] [CrossRef]
- Wu, W.; Bang, S.; Bleecker, E.R.; Castro, M.; Denlinger, L.; Erzurum, S.C.; Fahy, J.V.; Fitzpatrick, A.M.; Gaston, B.M.; Hastie, A.T.; et al. Multiview Cluster Analysis Identifies Variable Corticosteroid Response Phenotypes in Severe Asthma. Am. J. Respir. Crit. Care Med. 2019, 199, 1358–1367. [Google Scholar] [CrossRef]
- Peters, M.C.; McGrath, K.W.; Hawkins, G.A.; Hastie, A.T.; Levy, B.D.; Israel, E.; Phillips, B.R.; Mauger, D.T.; Comhair, S.A.; Erzurum, S.C.; et al. Plasma interleukin-6 concentrations, metabolic dysfunction, and asthma severity: A cross-sectional analysis of two cohorts. Lancet Respir. Med. 2016, 4, 574–584. [Google Scholar] [CrossRef] [Green Version]
- Telenga, E.D.; Tideman, S.W.; Kerstjens, H.A.; Hacken, N.H.; Timens, W.; Postma, D.S.; van den Berge, M. Obesity in asthma: More neutrophilic inflammation as a possible explanation for a reduced treatment response. Allergy 2012, 67, 1060–1068. [Google Scholar] [CrossRef]
- Strunk, R.C.; Colvin, R.; Bacharier, L.B.; Fuhlbrigge, A.; Forno, E.; Arbelaez, A.M.; Tantisira, K.G.; Childhood Asthma Management Program Research, G. Airway Obstruction Worsens in Young Adults with Asthma Who Become Obese. J. Allergy Clin. Immunol. Pract. 2015, 3, 765–771.e2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burgess, J.A.; Matheson, M.C.; Diao, F.; Johns, D.P.; Erbas, B.; Lowe, A.J.; Gurrin, L.C.; Lodge, C.J.; Thomas, P.S.; Morrison, S.; et al. Bronchial hyperresponsiveness and obesity in middle age: Insights from an Australian cohort. Eur. Respir. J. 2017, 50, 1602181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orfanos, S.; Jude, J.; Deeney, B.T.; Cao, G.; Rastogi, D.; van Zee, M.; Pushkarsky, I.; Munoz, H.E.; Damoiseaux, R.; Di Carlo, D.; et al. Obesity increases airway smooth muscle responses to contractile agonists. Am. J. Physiol. Lung Cell. Mol. Physiol. 2018, 315, L673–L681. [Google Scholar] [CrossRef] [PubMed]
- Peters, M.C.; Schiebler, M.L.; Cardet, J.C.; Johansson, M.W.; Sorkness, R.; DeBoer, M.D.; Bleecker, E.R.; Meyers, D.A.; Castro, M.; Sumino, K.; et al. The Impact of Insulin Resistance on Loss of Lung Function and Response to Treatment in Asthma. Am. J. Respir. Crit. Care Med. 2022, 206, 1096–1106. [Google Scholar] [CrossRef]
- Kim, H.Y.; Lee, H.J.; Chang, Y.J.; Pichavant, M.; Shore, S.A.; Fitzgerald, K.A.; Iwakura, Y.; Israel, E.; Bolger, K.; Faul, J.; et al. Interleukin-17-producing innate lymphoid cells and the NLRP3 inflammasome facilitate obesity-associated airway hyperreactivity. Nat. Med. 2014, 20, 54–61. [Google Scholar] [CrossRef] [Green Version]
- Woldhuis, R.R.; de Vries, M.; Timens, W.; van den Berge, M.; Demaria, M.; Oliver, B.G.G.; Heijink, I.H.; Brandsma, C.A. Link between increased cellular senescence and extracellular matrix changes in COPD. Am. J. Physiol. Lung Cell. Mol. Physiol. 2020, 319, L48–L60. [Google Scholar] [CrossRef]
- Yanagisawa, H.; Hashimoto, M.; Minagawa, S.; Takasaka, N.; Ma, R.; Moermans, C.; Ito, S.; Araya, J.; Budelsky, A.; Goodsell, A.; et al. Role of IL-17A in murine models of COPD airway disease. Am. J. Physiol. Lung Cell. Mol. Physiol. 2017, 312, L122–L130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, Z.; Zhu, L. Update on molecular mechanisms of corticosteroid resistance in chronic obstructive pulmonary disease. Pulm Pharmacol. Ther. 2016, 37, 1–8. [Google Scholar] [CrossRef]
- Tommola, M.; Ilmarinen, P.; Tuomisto, L.E.; Lehtimaki, L.; Haanpaa, J.; Niemela, O.; Kankaanranta, H. Differences between asthma-COPD overlap syndrome and adult-onset asthma. Eur. Respir. J. 2017, 49, 1602383. [Google Scholar] [CrossRef] [Green Version]
- Tu, X.; Kim, R.Y.; Brown, A.C.; de Jong, E.; Jones-Freeman, B.; Ali, M.K.; Gomez, H.M.; Budden, K.F.; Starkey, M.R.; Cameron, G.J.M.; et al. Airway and parenchymal transcriptomics in a novel model of asthma and COPD overlap. J. Allergy Clin. Immunol. 2022, 150, 817–829.e6. [Google Scholar] [CrossRef]
- Shah, R.; Newcomb, D.C. Sex Bias in Asthma Prevalence and Pathogenesis. Front. Immunol. 2018, 9, 2997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sathish, V.; Martin, Y.N.; Prakash, Y.S. Sex steroid signaling: Implications for lung diseases. Pharmacol. Ther. 2015, 150, 94–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, D.J.; Walters, E.H.; Perret, J.L.; Lodge, C.J.; Lowe, A.J.; Matheson, M.C.; Dharmage, S.C. Age-of-asthma onset as a determinant of different asthma phenotypes in adults: A systematic review and meta-analysis of the literature. Expert Rev. Respir. Med. 2015, 9, 109–123. [Google Scholar] [CrossRef] [PubMed]
- Zein, J.G.; Denson, J.L.; Wechsler, M.E. Asthma over the Adult Life Course: Gender and Hormonal Influences. Clin. Chest Med. 2019, 40, 149–161. [Google Scholar] [CrossRef] [PubMed]
- Pasha, M.A.; Sundquist, B.; Townley, R. Asthma pathogenesis, diagnosis, and management in the elderly. Allergy Asthma Proc. 2017, 38, 184–191. [Google Scholar] [CrossRef]
- Khosa, J.K.; Louie, S.; Lobo Moreno, P.; Abramov, D.; Rogstad, D.K.; Alismail, A.; Matus, M.J.; Tan, L.D. Asthma Care in the Elderly: Practical Guidance and Challenges for Clinical Management—A Framework of 5 “Ps”. J. Asthma Allergy 2023, 16, 33–43. [Google Scholar] [CrossRef] [PubMed]
- Park, H.W.; Cho, S.H. Management of Elderly Asthma: Key Questions and Tentative Answers. Allergy Asthma Immunol. Res. 2023, 15, 8–18. [Google Scholar] [CrossRef]
- Inoue, H.; Niimi, A.; Takeda, T.; Matsumoto, H.; Ito, I.; Matsuoka, H.; Jinnai, M.; Otsuka, K.; Oguma, T.; Nakaji, H.; et al. Pathophysiological characteristics of asthma in the elderly: A comprehensive study. Ann. Allergy Asthma Immunol. 2014, 113, 527–533. [Google Scholar] [CrossRef] [Green Version]
- Kanazawa, H.; Tochino, Y.; Kyoh, S.; Ichimaru, Y.; Asai, K.; Hirata, K. Potential roles of pentosidine in age-related and disease-related impairment of pulmonary functions in patients with asthma. J. Allergy Clin. Immunol. 2011, 127, 899–904. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ford, M.L.; Ruwanpathirana, A.; Lewis, B.W.; Britt, R.D., Jr. Aging-Related Mechanisms Contribute to Corticosteroid Insensitivity in Elderly Asthma. Int. J. Mol. Sci. 2023, 24, 6347. https://doi.org/10.3390/ijms24076347
Ford ML, Ruwanpathirana A, Lewis BW, Britt RD Jr. Aging-Related Mechanisms Contribute to Corticosteroid Insensitivity in Elderly Asthma. International Journal of Molecular Sciences. 2023; 24(7):6347. https://doi.org/10.3390/ijms24076347
Chicago/Turabian StyleFord, Maria L., Anushka Ruwanpathirana, Brandon W. Lewis, and Rodney D. Britt, Jr. 2023. "Aging-Related Mechanisms Contribute to Corticosteroid Insensitivity in Elderly Asthma" International Journal of Molecular Sciences 24, no. 7: 6347. https://doi.org/10.3390/ijms24076347