
Evaluation of Changes in Some Functional Properties of Human Mesenchymal Stromal Cells Induced by Low Doses of Ionizing Radiation
 , , and
, , and
Abstract
:1. Introduction
2. Results
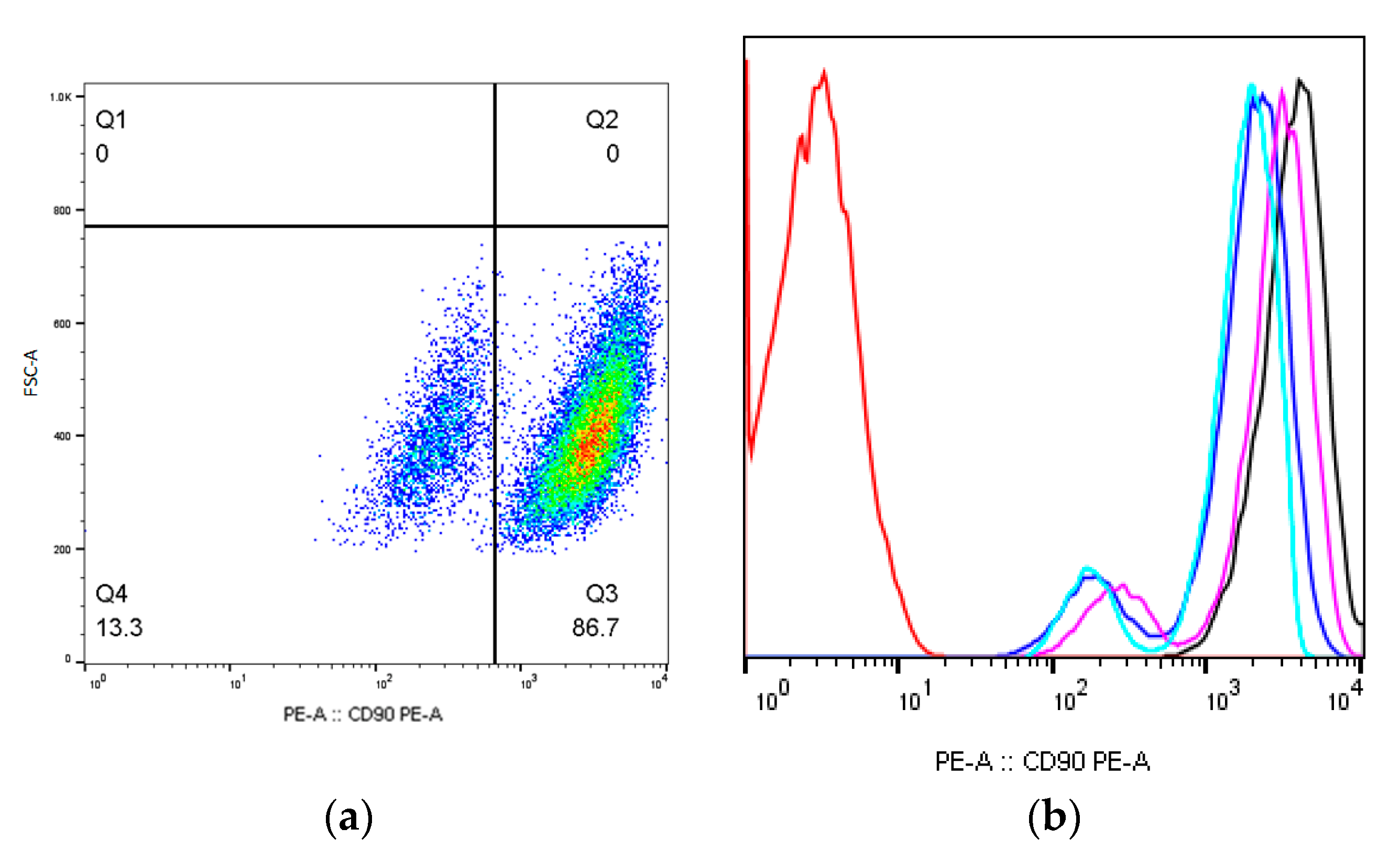
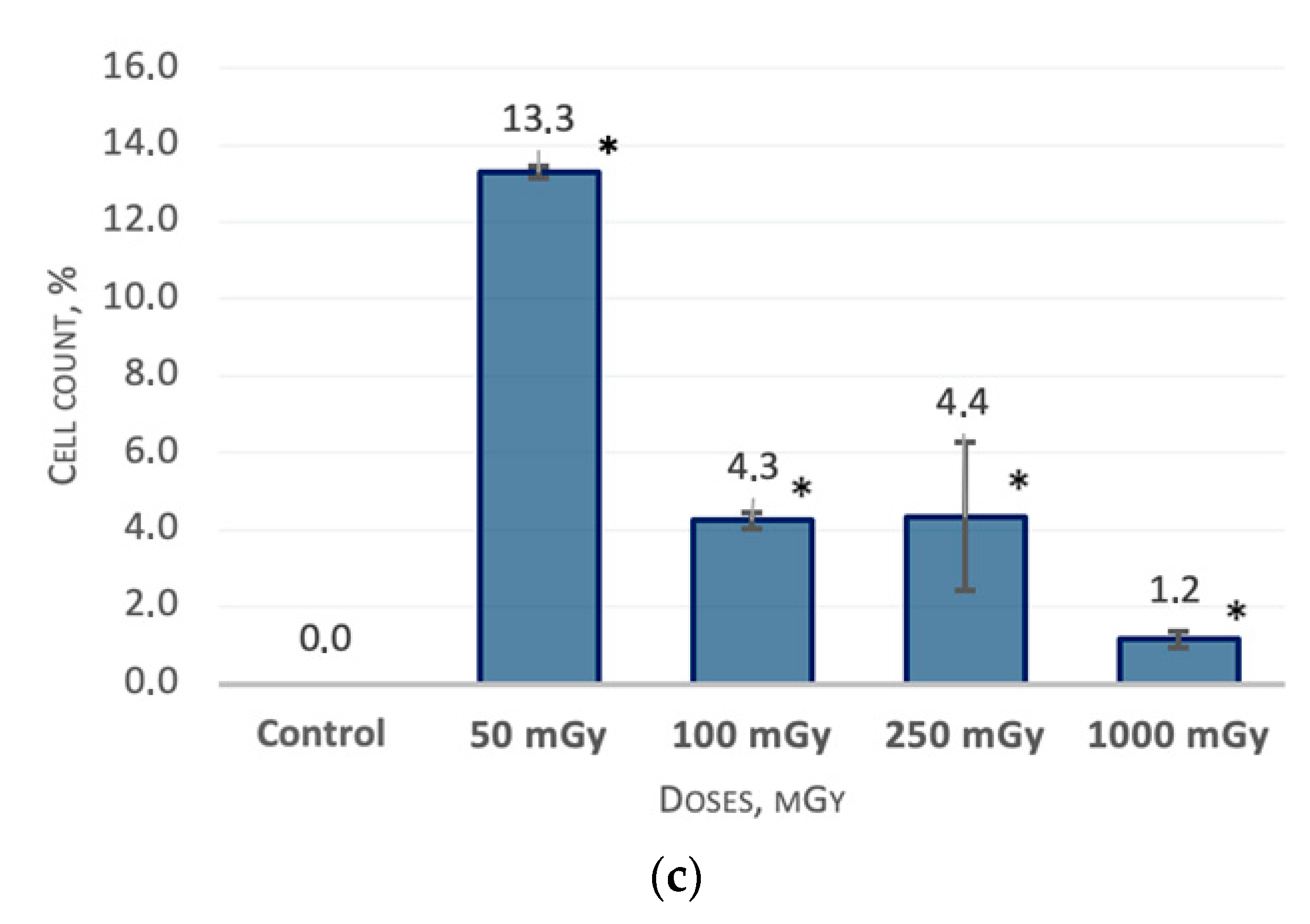
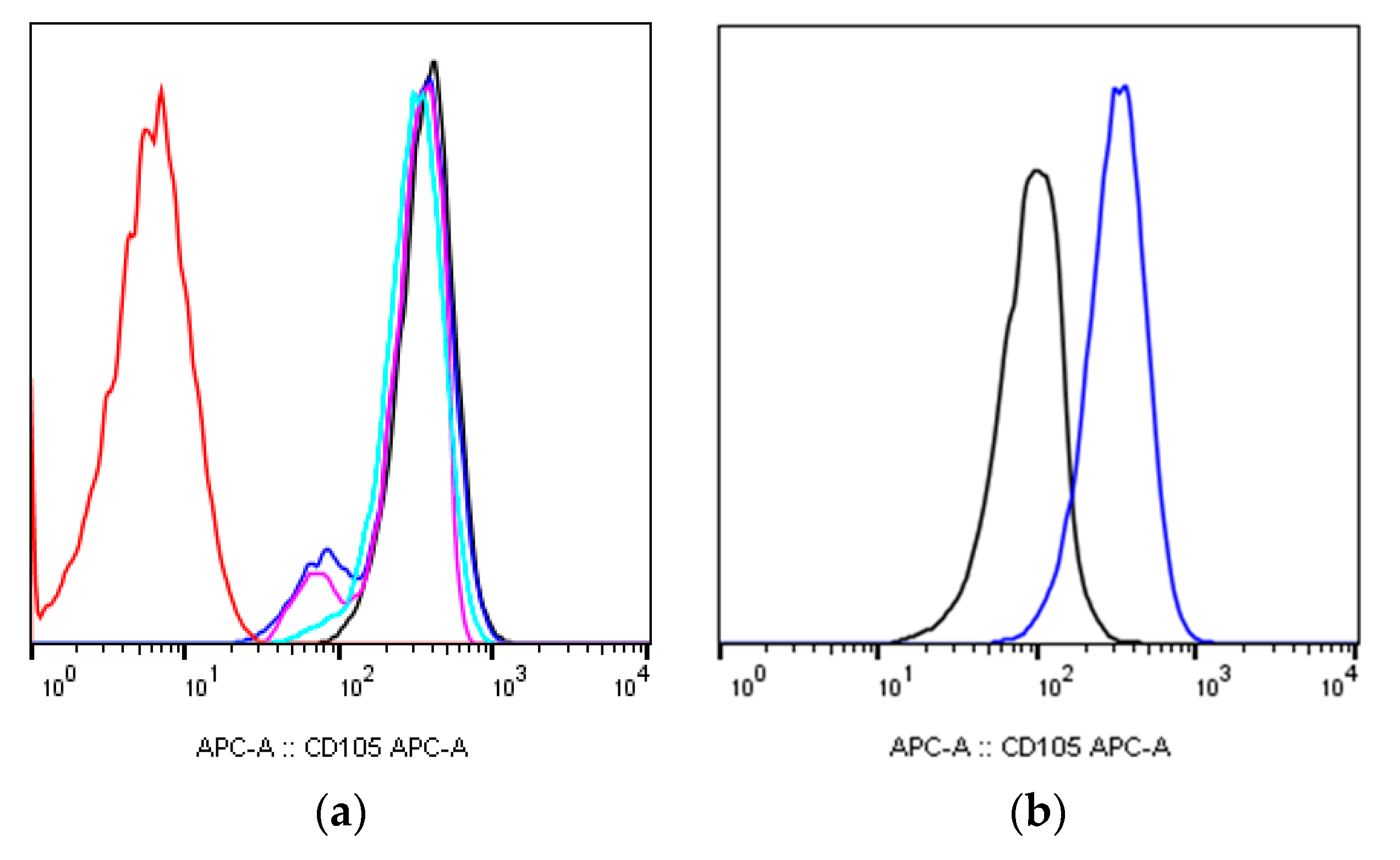
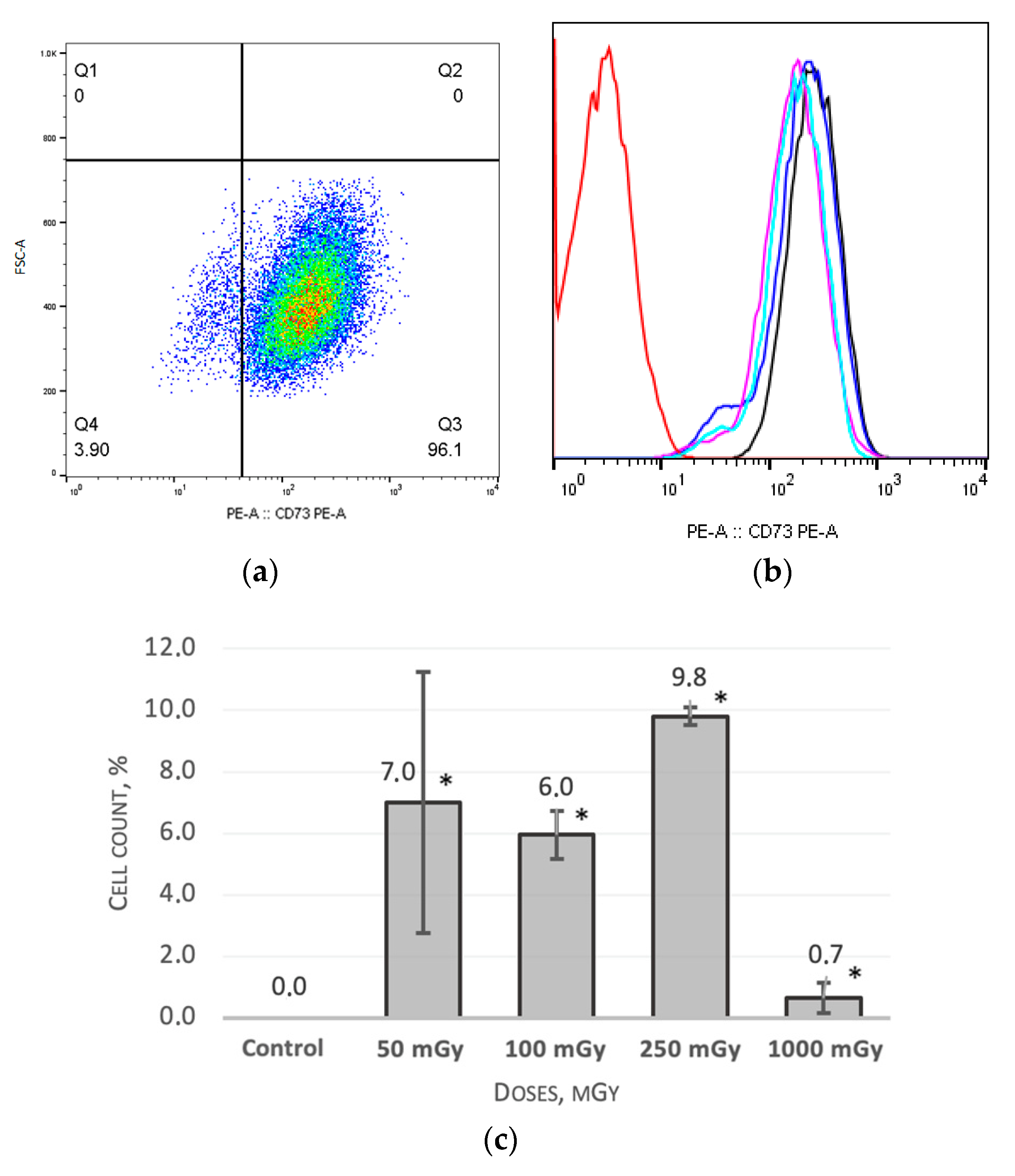
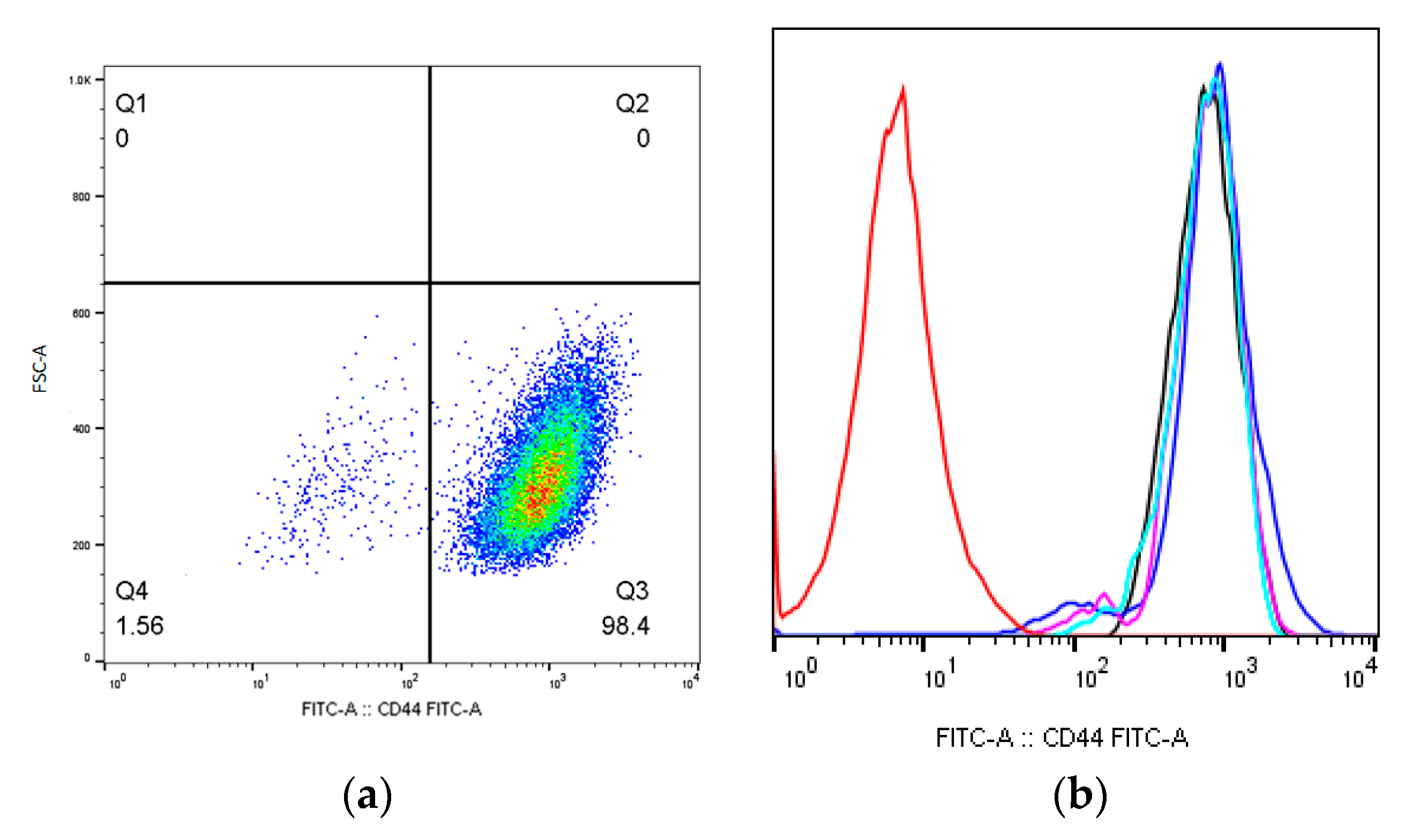
2.1. MSCs Immunophenotype after X-ray Irradiation
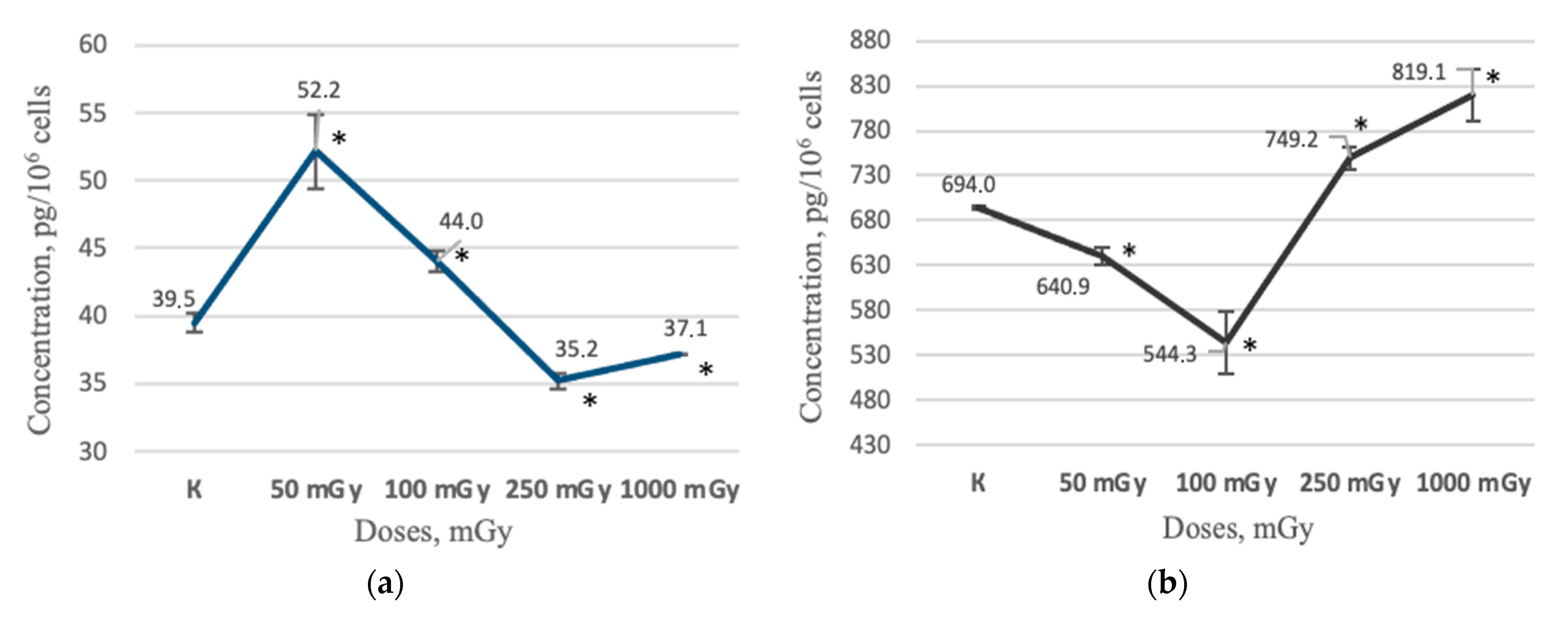
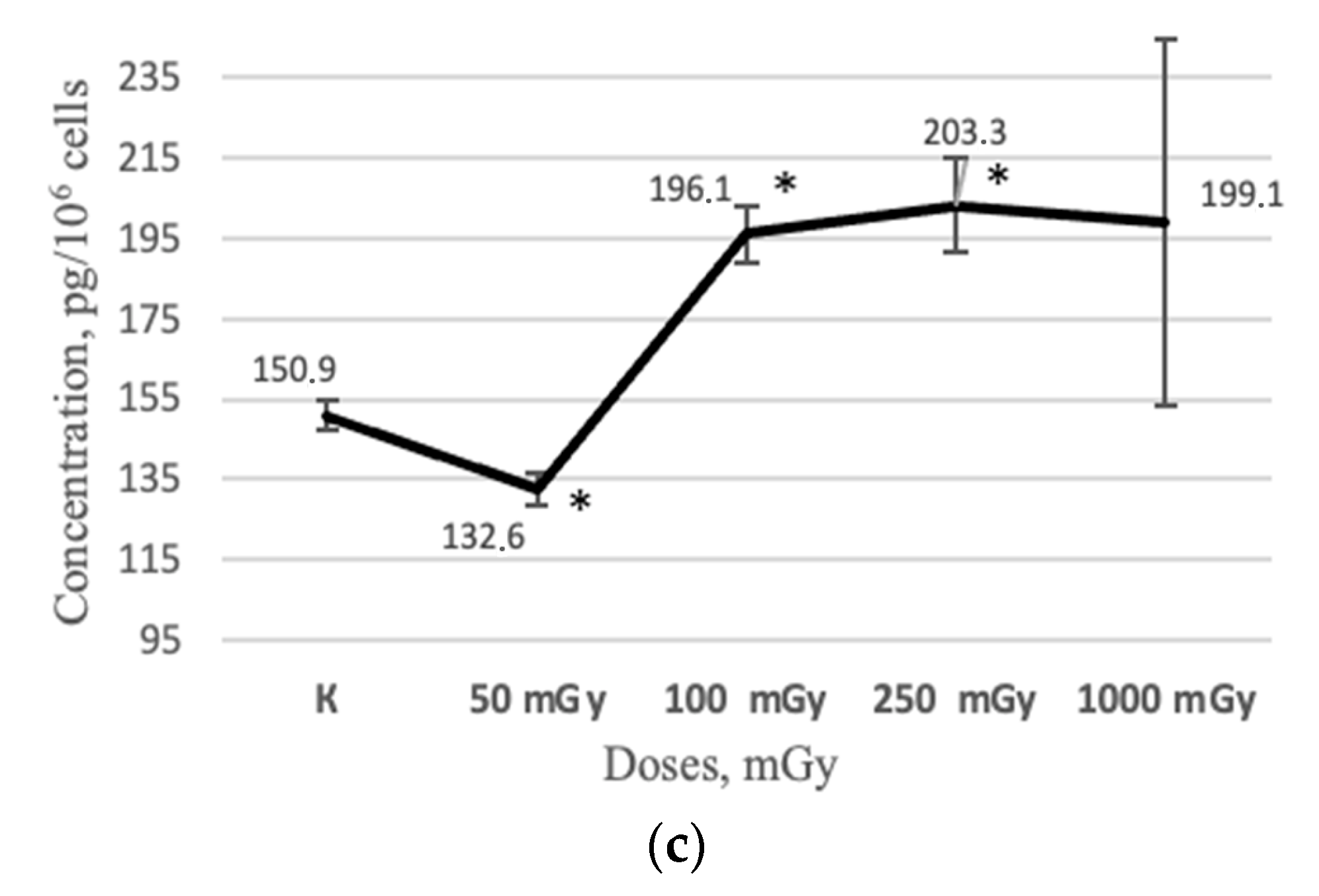
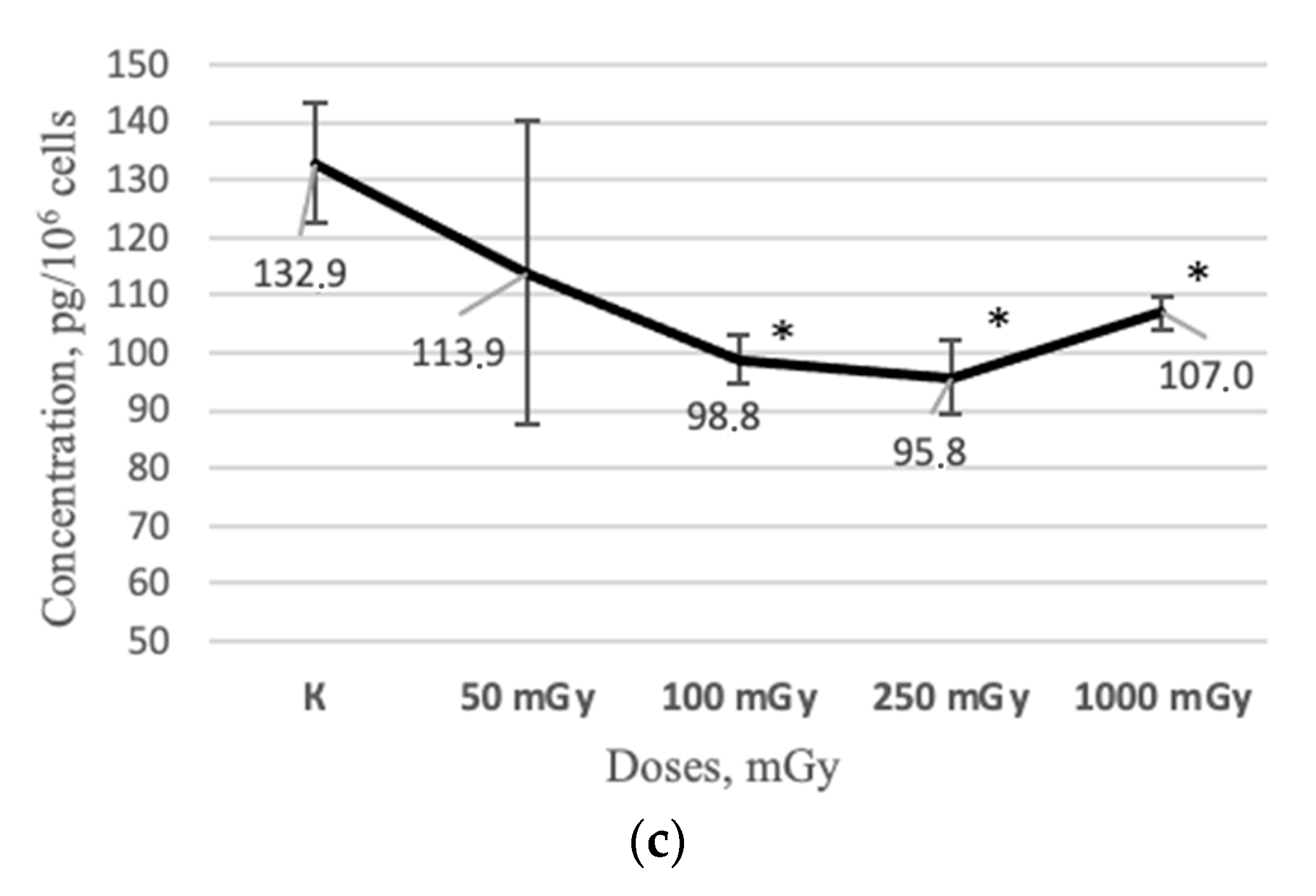
2.2. MSCs Secretory Profile after X-ray Irradiation
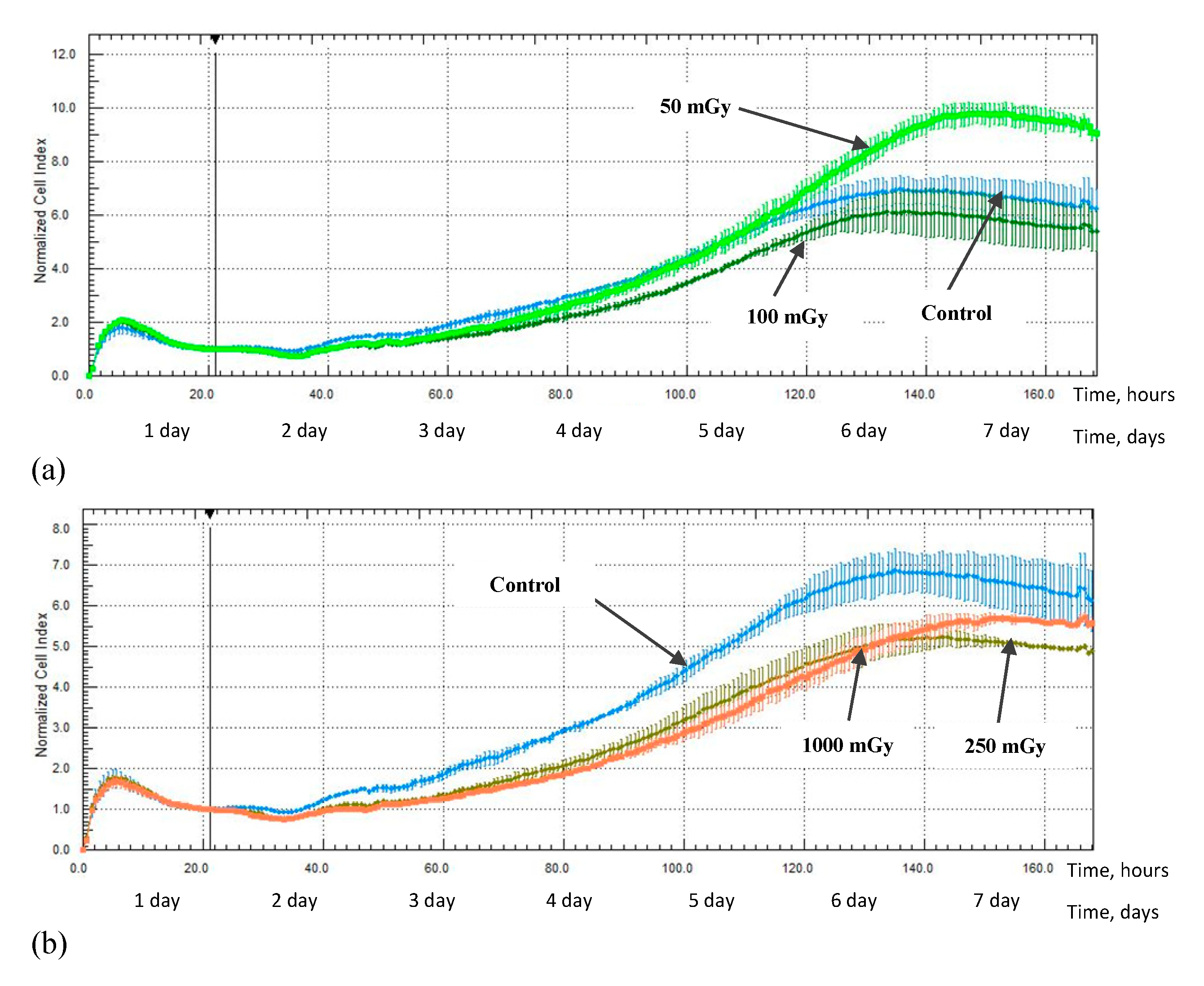
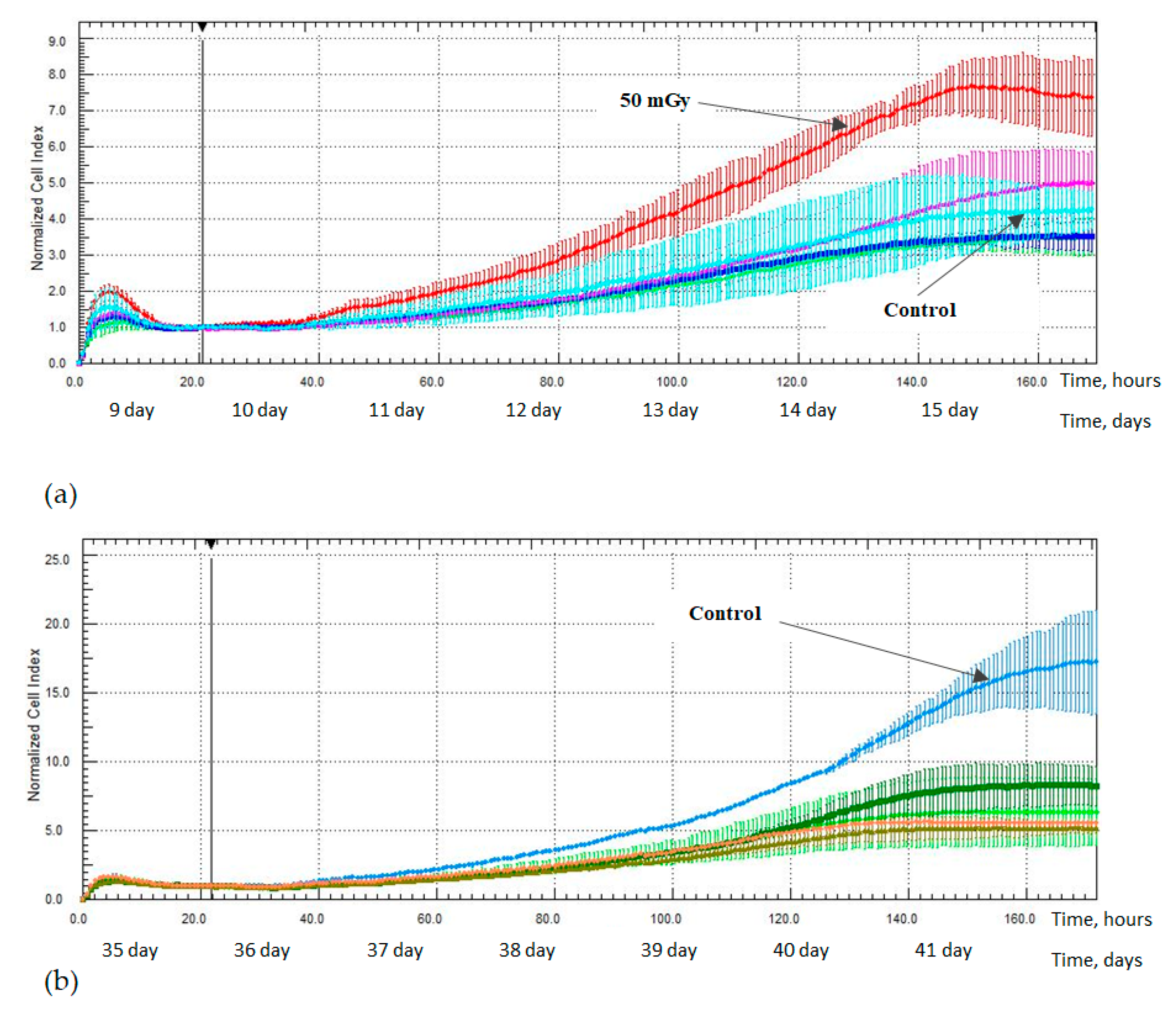
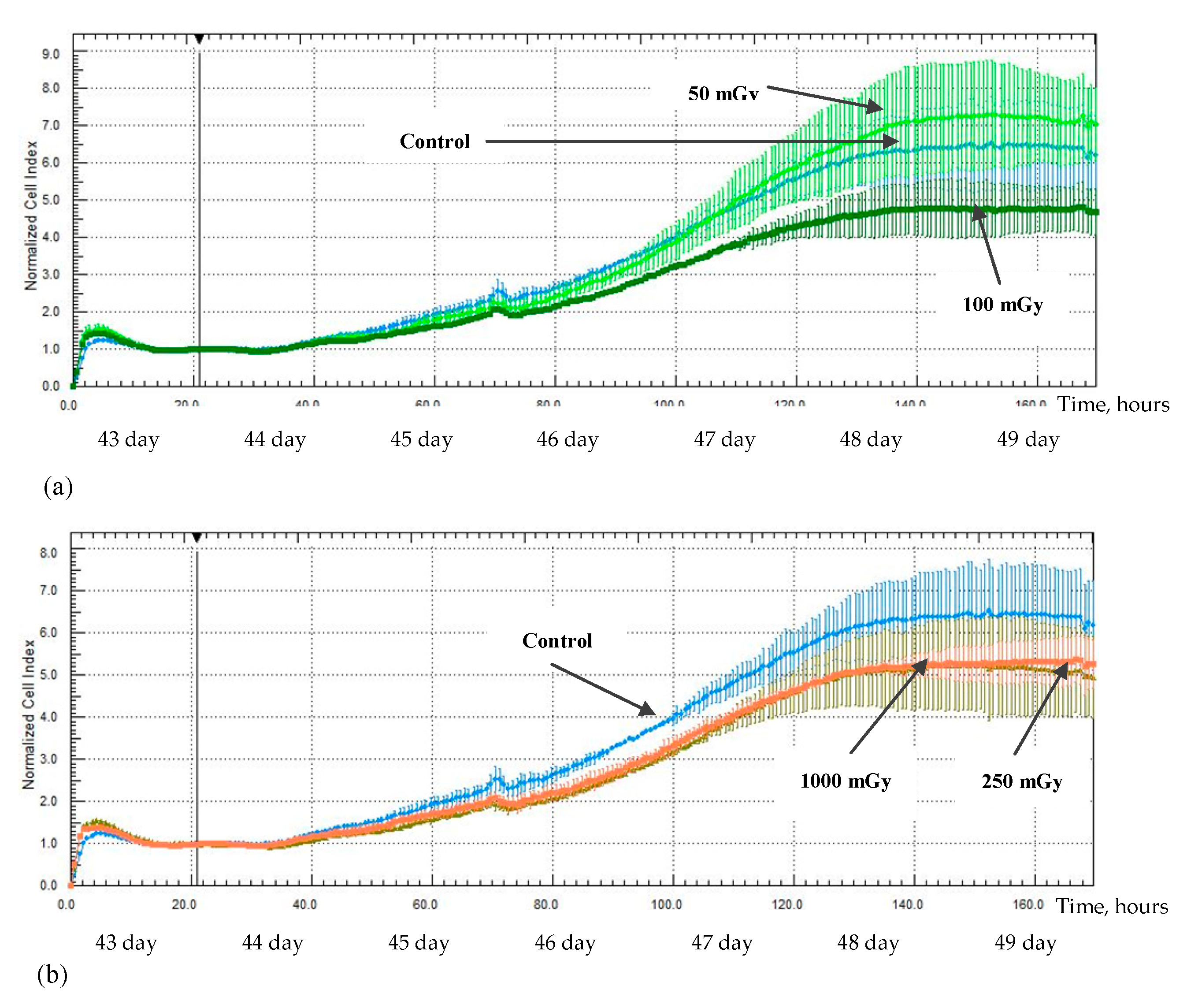
2.3. MSCs Proliferation Activity after X-ray Irradiation
3. Discussion
4. Materials and Methods
4.1. Isolation MSCs from the Samples of Human Connective Gingiva Tissue (hG-MSCs)
4.2. Cell Line Cultivation
4.3. Cryopreservation and Storage of Conditioned Media
4.4. X-ray Irradiation
4.5. Immunophenotyping of Cells
4.6. The Enzyme-Linked Immunosorbent Assay (ELISA)
- Human IL-8/CXCL8 Quantikine Elisa Kit (#D8000C, R&D systems a bio-techne brand, Minneapolis, MN, USA);
- Human IL-6 Elisa Kit (#BMS213-2, Invitrogen, Waltham, MA, USA);
- Human IL10 Elisa kit (#BMS215-2, Invitrogen, Waltham, MA, USA);
- Human VEGF-A Elisa kit (#BMS277-2, Invitrogen, Waltham, MA, USA);
- Human IDO Elisa kit (#EH246RB, Invitrogen, Waltham, MA, USA).
4.7. Evaluation of Cell Proliferative Activity
4.8. Statistics
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fazel, R.; Krumholz, H.; Wang, Y. Exposure to Low-Dose Ionizing Radiation from Medical Imaging Procedures. J. Vasc. Surg. 2009, 50, 1526–1527. [Google Scholar] [CrossRef] [Green Version]
- ICRP. 2007 Recommendations of the International Commission on Radiological Protection. Ann. ICRP 2007, 37, 9–34. [Google Scholar] [CrossRef]
- Squillaro, T.; Galano, G.; De Rosa, R.; Peluso, G.; Galderisi, U. Concise Review: The Effect of Low-Dose Ionizing Radiation on Stem Cell Biology: A Contribution to Radiation Risk. Stem Cells 2018, 36, 1146–1153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osipov, A.N.; Pustovalova, M.; Grekhova, A.; Eremin, P.; Vorobyova, N.; Pulin, A.; Zhavoronkov, A.; Roumiantsev, S.; Klokov, D.Y.; Eremin, I. Low doses of X-rays induce prolonged and ATM-independent persistence of γH2AX foci in human gingival mesenchymal stem cells. Oncotarget 2015, 6, 27275–27287. [Google Scholar] [CrossRef] [Green Version]
- Ji, K.; Wang, Y.; Du, L.; Xu, C.; Liu, Y.; He, N.; Wang, J.; Liu, Q. Research Progress on the Biological Effects of Low-Dose Radiation in China. Dose-Response 2019, 17, 1559325819833488. [Google Scholar] [CrossRef] [Green Version]
- Doss, M. Are We Approaching the End of the Linear No-Threshold Era? J. Nucl. Med. 2018, 59, 1786–1793. [Google Scholar] [CrossRef] [Green Version]
- Sokolov, M.V.; Neumann, R.D. Human Embryonic Stem Cell Responses to Ionizing Radiation Exposures: Current State of Knowledge and Future Challenges. Stem Cells Int. 2012, 2012, 579104. [Google Scholar] [CrossRef] [Green Version]
- Prise, K.M.; Saran, A. Concise Review: Stem Cell Effects in Radiation Risk. Stem Cells 2011, 29, 1315–1321. [Google Scholar] [CrossRef]
- Islam, M.S.; Stemig, M.E.; Takahashi, Y.; Hui, S.K. Radiation response of mesenchymal stem cells derived from bone marrow and human pluripotent stem cells. J. Radiat. Res. 2014, 56, 269–277. [Google Scholar] [CrossRef] [Green Version]
- Pustovalova, M.V.; Grekhova, A.K.; Osipov, A.N. Mesenchymal stem cells: Effects of exposure to ionizing radiation in small doses // Radiation biology. Radioecology 2018, 58, 352–362. [Google Scholar] [CrossRef]
- Wu, P.-K.; Wang, J.-Y.; Chen, C.-F.; Chao, K.-Y.; Chang, M.-C.; Chen, W.-M.; Hung, S.-C. Early Passage Mesenchymal Stem Cells Display Decreased Radiosensitivity and Increased DNA Repair Activity. Stem Cells Transl. Med. 2017, 6, 1504–1514. [Google Scholar] [CrossRef]
- Langen, B.; Vorontsov, E.; Spetz, J.; Swanpalmer, J.; Sihlbom, C.; Helou, K.; Forssell-Aronsson, E. Age and sex effects across the blood proteome after ionizing radiation exposure can bias biomarker screening and risk assessment. Sci. Rep. 2022, 12, 7000. [Google Scholar] [CrossRef]
- Kovalchuk, O.; Ponton, A.; Filkowski, J.; Kovalchuk, I. Dissimilar genome response to acute and chronic low-dose radiation in male and female mice. Mutat. Res. Mol. Mech. Mutagen. 2004, 550, 59–72. [Google Scholar] [CrossRef]
- Besplug, J.; Burke, P.; Ponton, A.; Filkowski, J.; Titov, V.; Kovalchuk, I.; Kovalchuk, O. Sex and tissue-specific differences in low-dose radiation-induced oncogenic signaling. Int. J. Radiat. Biol. 2005, 81, 157–168. [Google Scholar] [CrossRef]
- Silasi, G.; Diaz-Heijtz, R.; Besplug, J.; Rodriguez-Juarez, R.; Titov, V.; Kolb, B.; Kovalchuk, O. Selective brain responses to acute and chronic low-dose X-ray irradiation in males and females. Biochem. Biophys. Res. Commun. 2004, 325, 1223–1235. [Google Scholar] [CrossRef]
- Parihar, V.K.; Angulo, M.C.; Allen, B.D.; Syage, A.; Usmani, M.T.; De La Chapelle, E.P.; Amin, A.N.; Flores, L.; Lin, X.; Giedzinski, E.; et al. Sex-Specific Cognitive Deficits Following Space Radiation Exposure. Front. Behav. Neurosci. 2020, 14, 535885. [Google Scholar] [CrossRef]
- Lawler, P.R.; Afilalo, J.; Eisenberg, M.J.; Pilote, L. Comparison of Cancer Risk Associated With Low-Dose Ionizing Radiation from Cardiac Imaging and Therapeutic Procedures After Acute Myocardial Infarction in Women Versus Men. Am. J. Cardiol. 2013, 112, 1545–1550. [Google Scholar] [CrossRef]
- Narendran, N.; Luzhna, L.; Kovalchuk, O. Sex Difference of Radiation Response in Occupational and Accidental Exposure. Front. Genet. 2019, 10, 260. [Google Scholar] [CrossRef]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.C.; Krause, D.S.; Deans, R.J.; Keating, A.; Prockop, D.J.; Horwitz, E.M. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef]
- Park, C.W.; Kim, K.-S.; Bae, S.; Son, H.K.; Myung, P.-K.; Hong, H.J.; Kim, H. Cytokine Secretion Profiling of Human Mesenchymal Stem Cells by Antibody Array. Int. J. Stem Cells 2009, 2, 59–68. [Google Scholar] [CrossRef] [Green Version]
- Elman, J.S.; Li, M.; Wang, F.; Gimble, J.M.; Parekkadan, B. A comparison of adipose and bone marrow-derived mesenchymal stromal cell secreted factors in the treatment of systemic inflammation. J. Inflamm. 2014, 11, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferreira, J.R.; Teixeira, G.Q.; Santos, S.G.; Barbosa, M.A.; Almeida-Porada, G.; Gonçalves, R.M. Mesenchymal Stromal Cell Secretome: Influencing Therapeutic Potential by Cellular Pre-conditioning. Front. Immunol. 2018, 9, 2837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Netsch, P.; Elvers-Hornung, S.; Uhlig, S.; Klüter, H.; Huck, V.; Kirschhöfer, F.; Brenner-Weiß, G.; Janetzko, K.; Solz, H.; Wuchter, P.; et al. Human mesenchymal stromal cells inhibit platelet activation and aggregation involving CD73-converted adenosine. Stem Cell Res. Ther. 2018, 9, 184. [Google Scholar] [CrossRef] [PubMed]
- Canepa, D.D.; Casanova, E.A.; Arvaniti, E.; Tosevski, V.; Märsmann, S.; Eggerschwiler, B.; Halvachizadeh, S.; Buschmann, J.; Barth, A.A.; Plock, J.A.; et al. Identification of ALP+/CD73+ defining markers for enhanced osteogenic potential in human adipose-derived mesenchymal stromal cells by mass cytometry. Stem Cell Res. Ther. 2021, 12, 7. [Google Scholar] [CrossRef]
- Moraes, D.A.; Sibov, T.T.; Pavon, L.F.; Alvim, P.Q.; Bonadio, R.S.; Da Silva, J.R.; Pic-Taylor, A.; Toledo, O.A.; Marti, L.C.; Azevedo, R.B.; et al. A reduction in CD90 (THY-1) expression results in increased differentiation of mesenchymal stromal cells. Stem Cell Res. Ther. 2016, 7, 97. [Google Scholar] [CrossRef] [Green Version]
- James, A.W. Review of Signaling Pathways Governing MSC Osteogenic and Adipogenic Differentiation. Scientifica 2013, 2013, 684736. [Google Scholar] [CrossRef]
- Saalbach, A.; Anderegg, U. Thy-1: More than a marker for mesenchymal stromal cells. FASEB J. 2019, 33, 6689–6696. [Google Scholar] [CrossRef]
- Anderson, P.; Carrillo-Gálvez, A.B.; García-Pérez, A.; Cobo, M.; Martín, F. CD105 (Endoglin)-Negative Murine Mesenchymal Stromal Cells Define a New Multipotent Subpopulation with Distinct Differentiation and Immunomodulatory Capacities. PLoS ONE 2013, 8, e76979. [Google Scholar] [CrossRef]
- Rada, T.; Reis, R.L.; Gomes, M.E. Distinct stem cells subpopulations isolated from human adipose tissue exhibit different chondrogenic and osteogenic differentiation potential. Stem Cell Rev. Rep. 2010, 7, 64–76. [Google Scholar] [CrossRef] [Green Version]
- Jiang, T.; Liu, W.; Lv, X.; Sun, H.; Zhang, L.; Liu, Y.; Zhang, W.J.; Cao, Y.; Zhou, G. Potent in vitro chondrogenesis of CD105 enriched human adipose-derived stem cells. Biomaterials 2010, 31, 3564–3571. [Google Scholar] [CrossRef]
- Lesley, J.; Hascall, V.C.; Tammi, M.; Hyman, R. Hyaluronan binding by cell surface CD44. J. Biol. Chem. 2000, 35, 26967–26975. [Google Scholar] [CrossRef]
- Zhu, H.; Mitsuhashi, N.; Klein, A.; Barsky, L.W.; Weinberg, K.; Barr, M.L.; Demetriou, A.; Wu, G.D. The role of the hyaluronan receptor CD44 in mesenchymal stem cell migration in the extracellular matrix. Stem Cells 2006, 24, 928–935. [Google Scholar] [CrossRef]
- Sapaeth, E.L.; Labaff, A.M.; Toole, B.P.; Klopp, A.; Andreeff, M.; Marini, F.C. Mesenchymal CD44 expression contributes to the acquisition of an activated fibroblast phenotype via TWIST activation in the tumor microenvironment. Cancer Res. 2013, 73, 5347–5359. [Google Scholar] [CrossRef] [Green Version]
- Barker, T.H.; Grenett, H.E.; MacEwen, M.W.; Tilden, S.G.; Fuller, G.M.; Settleman, J.; Woods, A.; Murphy-Ullrich, J.; Hagood, J.S. Thy-1 regulates fibroblast focal adhesions, cytoskeletal organization and migration through modulation of p190 RhoGAP and Rho GTPase activity. Exp. Cell Res. 2004, 295, 488–496. [Google Scholar] [CrossRef]
- Tabata, C.; Kubo, H.; Tabata, R.; Wada, M.; Sakuma, K.; Ichikawa, M.; Fujita, S.; Mio, T.; Mishima, M. All-trans retinoic acid modulates radiation-induced proliferation of lung fibroblasts via IL-6/IL-6R system. Am. J. Physiol. Cell. Mol. Physiol. 2006, 290, L597–L606. [Google Scholar] [CrossRef] [Green Version]
- Brach, M.A.; Gruss, H.J.; Kaisho, T.; Asano, Y.; Hirano, T.; Herrmann, F. Ionizing radiation induces expression of interleukin 6 by human fibro- blasts involving activation of nuclear factor-kappa B. J. Biol. Chem. 1993, 268, 8466–8472. [Google Scholar] [CrossRef]
- Martin, M.; Vozenin, M.C.; Gault, N.; Crechet, F.; Pfarr, C.M.; Lefaix, J.L. Co-activation of AP-1 activity and TGF-b1 gene expression in the stress response of normal skin cells to ionizing radiation. Oncogene 1997, 15, 981–989. [Google Scholar] [CrossRef] [Green Version]
- Müller, K.; Meineke, V. Radiation-induced alterations in cytokine production by skin cells. Exp. Hematol. 2007, 35, 96–104. [Google Scholar] [CrossRef]
- Beetz, A.; Peter, R.U.; Ried, C.; Ruzicka, T.; Michel, G. Uniform induction of TNF-a and IL-8 in human keratinocytes by ionizing radiation is accompanied by non-uniform regulation of corresponding receptors. J. Eur. Acad. Dermatol. Venereol. 1996, 7, 188–190. [Google Scholar] [CrossRef]
- Narayanan, P.K.; LaRue, K.E.A.; Goodwin, E.H.; Lehnert, B.E. Alpha Particles Induce the Production of Interleukin-8 by Human Cells. Radiat. Res. 1999, 152, 57–63. [Google Scholar] [CrossRef]
- Belmans, N.; Gilles, L.; Welkenhuysen, J.; Vermeesen, R.; Baselet, B.; Salmon, B.; Baatout, S.; Jacobs, R.; Lucas, S.; Lambrichts, I.; et al. In vitro Assessment of the DNA Damage Response in Dental Mesenchymal Stromal Cells Following Low Dose X-ray Exposure. Front. Public Health 2021, 9, 584484. [Google Scholar] [CrossRef] [PubMed]
- Bernal, A.J.; Dolinoy, D.C.; Huang, D.; Skaar, D.A.; Weinhouse, C.; Jirtle, R.L. Adaptive radiation-induced epigenetic alterations mitigated by antioxidants. FASEB J. 2012, 27, 665–671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grdina, D.J.; Murley, J.S.; Miller, R.C.; Mauceri, H.J.; Sutton, H.G.; Thirman, M.J.; Li, J.J.; Woloschak, G.E.; Weichselbaum, R.R. A Manganese Superoxide Dismutase (SOD2)-Mediated Adaptive Response. Radiat. Res. 2013, 179, 115–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fabbri, E.; Brognara, E.; Montagner, G.; Ghimenton, C.; Eccher, A.; Cantù, C.; Khalil, S.; Bezzerri, V.; Provezza, L.; Bianchi, N.; et al. Regulation of IL-8 gene expression in gliomas by microRNA miR-93. BMC Cancer 2015, 15, 661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, X.; So, Y.H.; Cui, J.; Ma, K.; Xu, X.; Zhao, Y.; Cai, L.; Li, W. The low-dose ionizing radiation stimulates cell proliferation via activation of the mapk/erk pathway in rat cultured mesenchymal stem cells. J. Radiat. Res. 2011, 52, 380–386. [Google Scholar] [CrossRef] [Green Version]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | K | 50 mGy | 100 mGy | 250 mGy | 1000 mGy |
|---|---|---|---|---|---|
| % of Positive Cells | |||||
| CD45 | 0.0 ± 0.00 | 0.0 ± 0.00 | 0.0 ± 0.00 | 0.0 ± 0.00 | 0.0 ± 0.00 |
| CD90 | 99.9 ± 0.00 | 99.9 ± 0.20 | 99.9 ± 0.12 | 99.9 ± 0.00 | 99.9 ± 0.08 |
| CD105 | 99.9 ± 0.15 | 99.9 ± 0.24 | 99.9 ± 0.21 | 99.9 ± 0.10 | 99.9 ± 0.00 |
| CD34 | 0.0 ± 0.00 | 0.0 ± 0.00 | 0.0 ± 0.00 | 0.0 ± 0.00 | 0.0 ± 0.00 |
| CD73 | 99.9 ± 0.00 | 99.9 ± 0.00 | 99.9 ± 0.23 | 99.9 ± 0.33 | 99.9 ± 0.19 |
| HLA-DR | 0.0 ± 0.00 | 0.0 ± 0.00 | 0.0 ± 0.00 | 0.0 ± 0.00 | 0.0 ± 0.00 |
| CD44 | 99.9 ± 0.00 | 98.6 ± 0.20 | 99.9 ± 0.36 | 99.9 ± 0.16 | 99.9 ± 0.31 |
| Tube 1 | Tube 2 | Tube 3 | Tube 4 | Tube 5 |
|---|---|---|---|---|
| Unstained control | 7AAD | CD45 FITC | CD34 FITC | CD44 FITC |
| CD90 PE | CD73 PE | - | ||
| CD105 APC | HLA-DR APC | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Usupzhanova, D.Y.; Astrelina, T.A.; Kobzeva, I.V.; Suchkova, Y.B.; Brunchukov, V.A.; Rastorgueva, A.A.; Nikitina, V.A.; Samoilov, A.S. Evaluation of Changes in Some Functional Properties of Human Mesenchymal Stromal Cells Induced by Low Doses of Ionizing Radiation. Int. J. Mol. Sci. 2023, 24, 6346. https://doi.org/10.3390/ijms24076346
Usupzhanova DY, Astrelina TA, Kobzeva IV, Suchkova YB, Brunchukov VA, Rastorgueva AA, Nikitina VA, Samoilov AS. Evaluation of Changes in Some Functional Properties of Human Mesenchymal Stromal Cells Induced by Low Doses of Ionizing Radiation. International Journal of Molecular Sciences. 2023; 24(7):6346. https://doi.org/10.3390/ijms24076346
Chicago/Turabian StyleUsupzhanova, Daria Yu., Tatiana A. Astrelina, Irina V. Kobzeva, Yulia B. Suchkova, Vitaliy A. Brunchukov, Anna A. Rastorgueva, Victoria A. Nikitina, and Alexander S. Samoilov. 2023. "Evaluation of Changes in Some Functional Properties of Human Mesenchymal Stromal Cells Induced by Low Doses of Ionizing Radiation" International Journal of Molecular Sciences 24, no. 7: 6346. https://doi.org/10.3390/ijms24076346