
Transcriptomic Analysis from Normal Glucose Tolerance to T2D of Obese Individuals Using Bioinformatic Tools

Abstract
:1. Introduction
2. Results
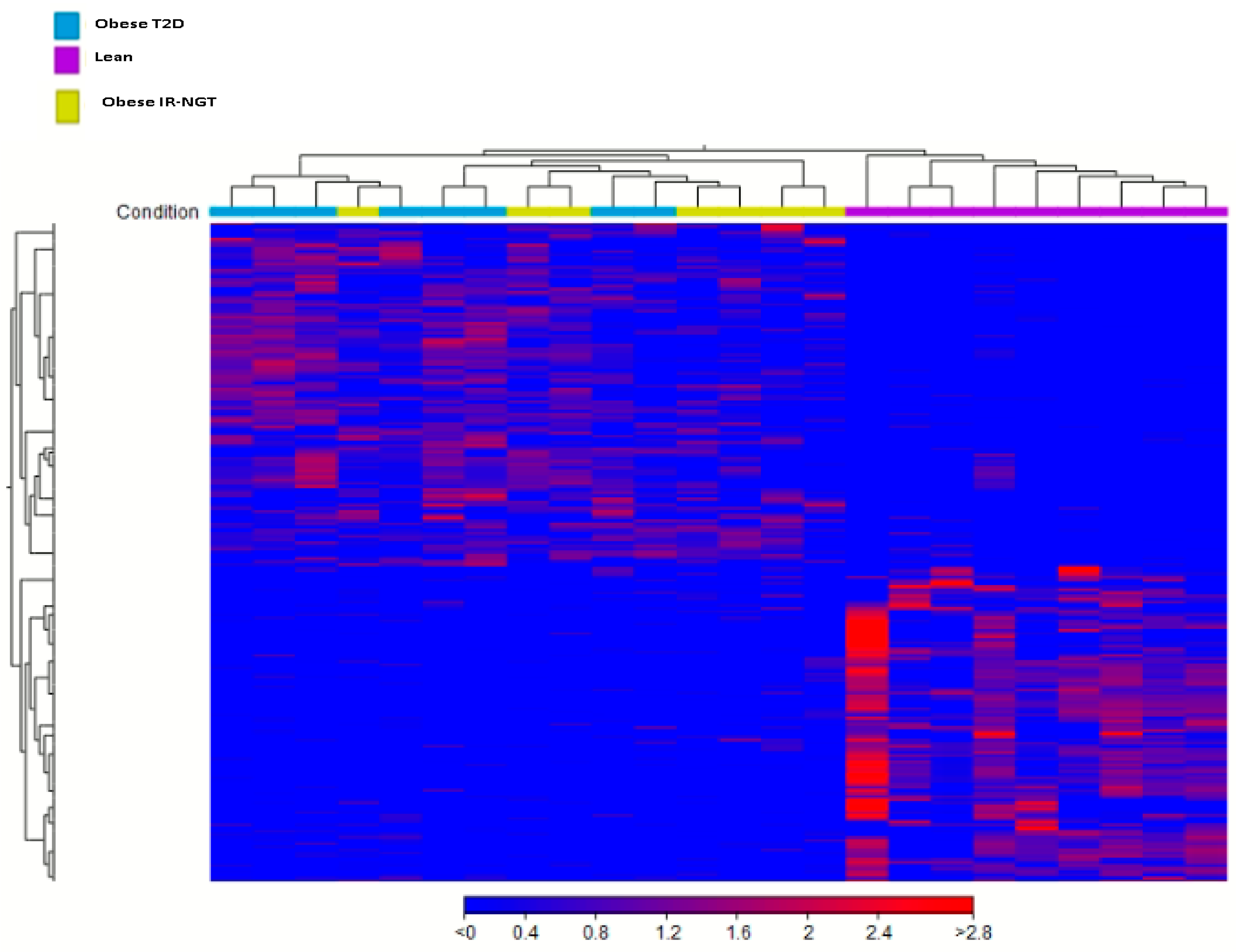
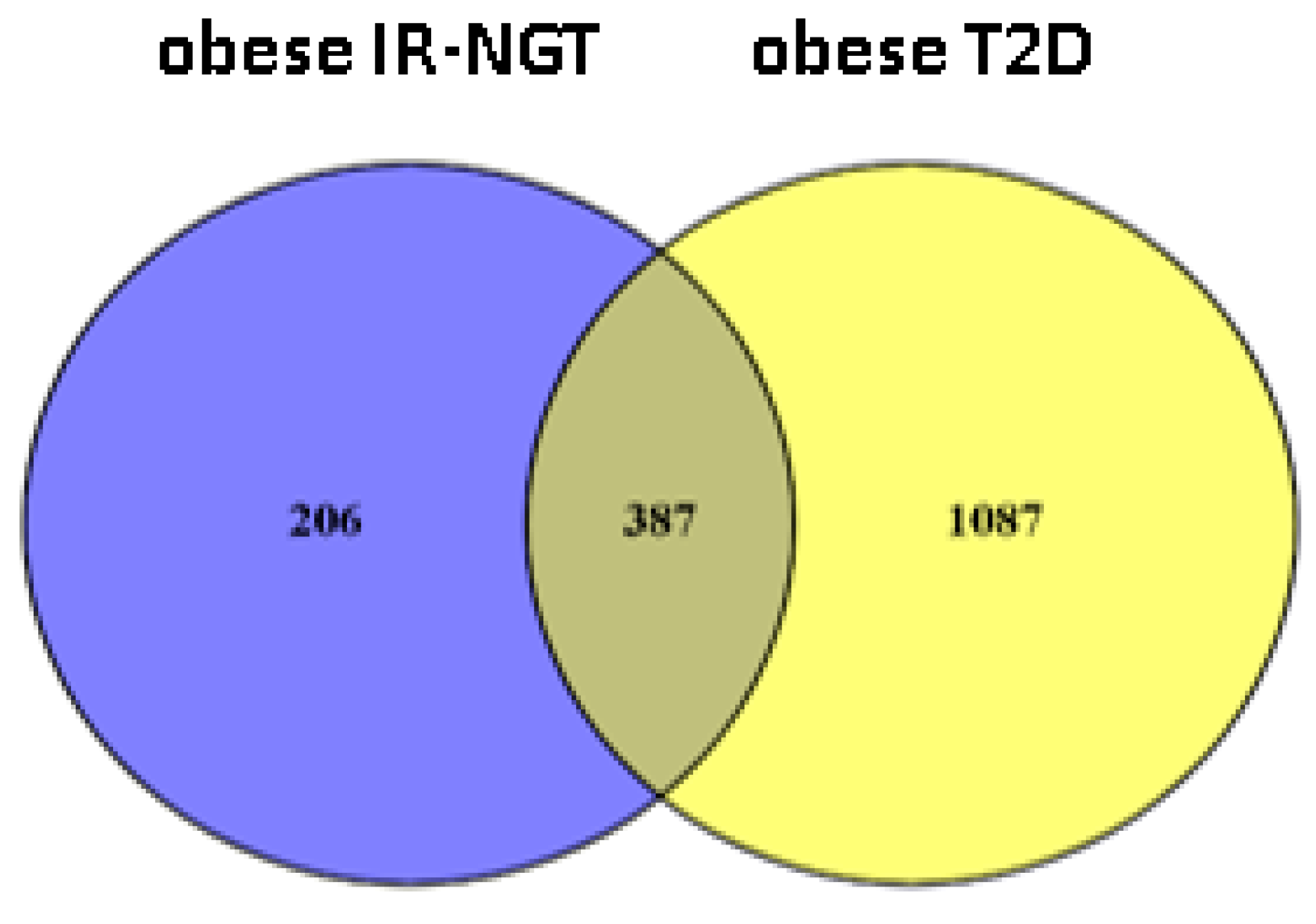
2.1. Identification of DEGs in Obese IR-NGT and Obese T2D
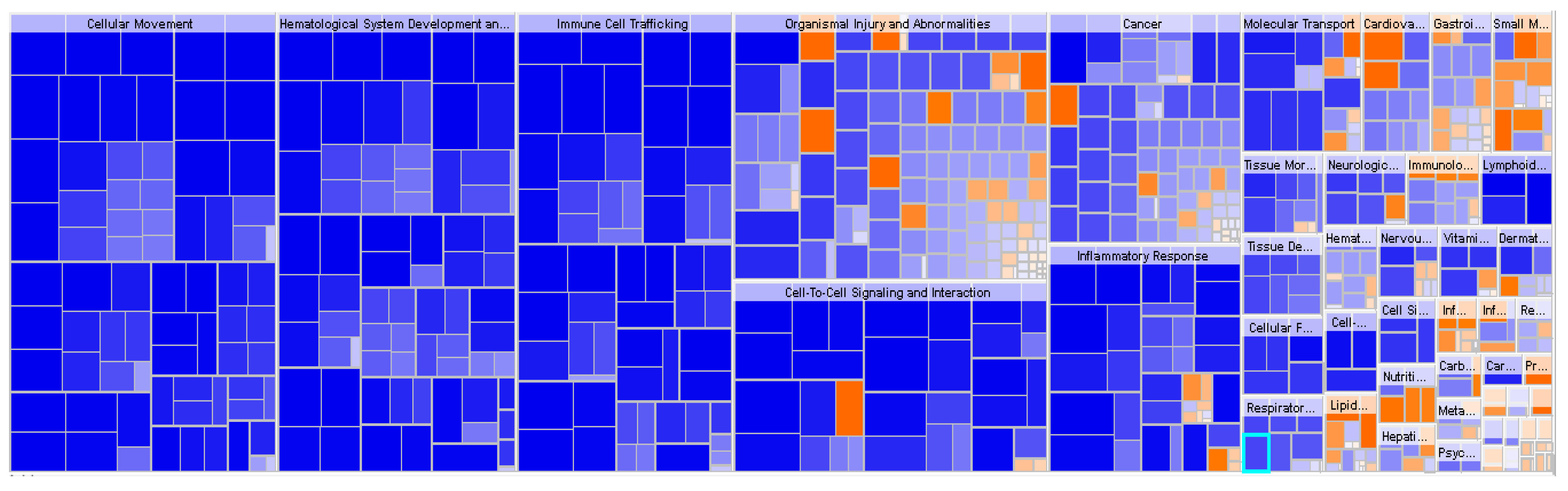
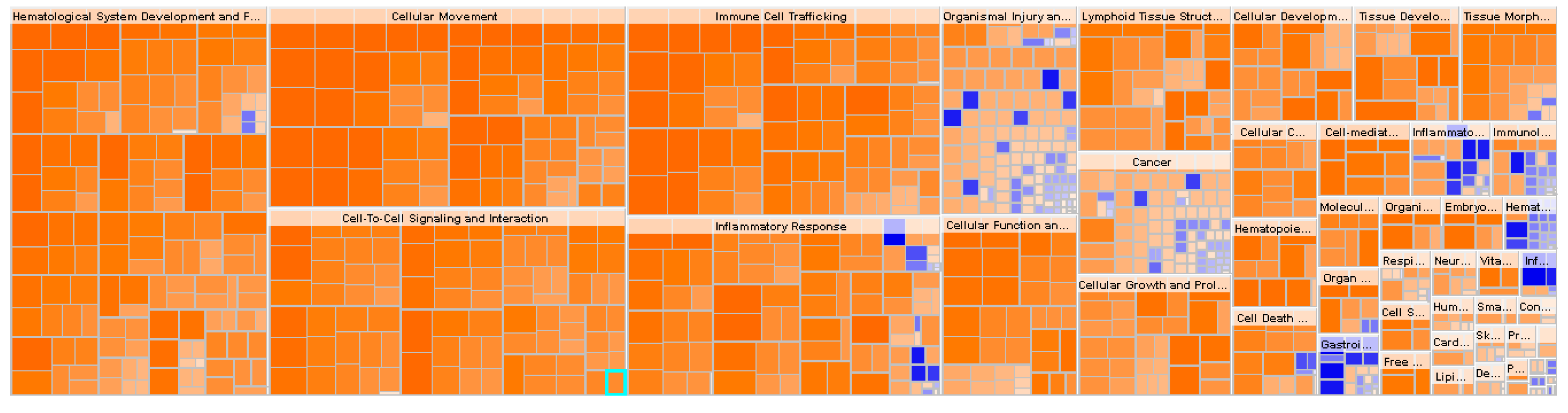
2.2. Functional Enrichment Analysis Highlights Inhibition of Inflammatory Responses in Obese IR-NGT versus an Activation of Inflammatory Response and Immune Cell Trafficking of WAT in Obese T2D Patients
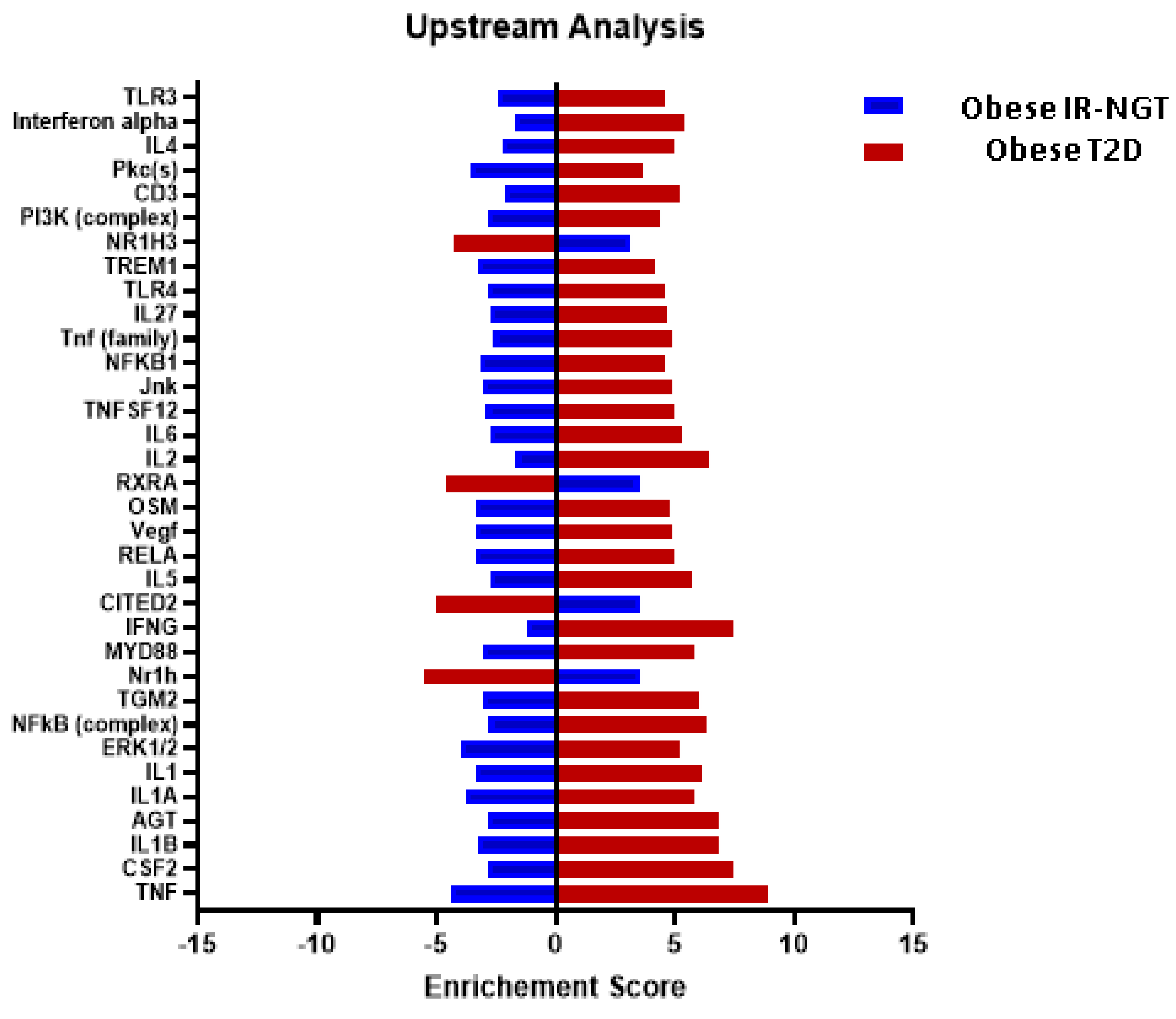
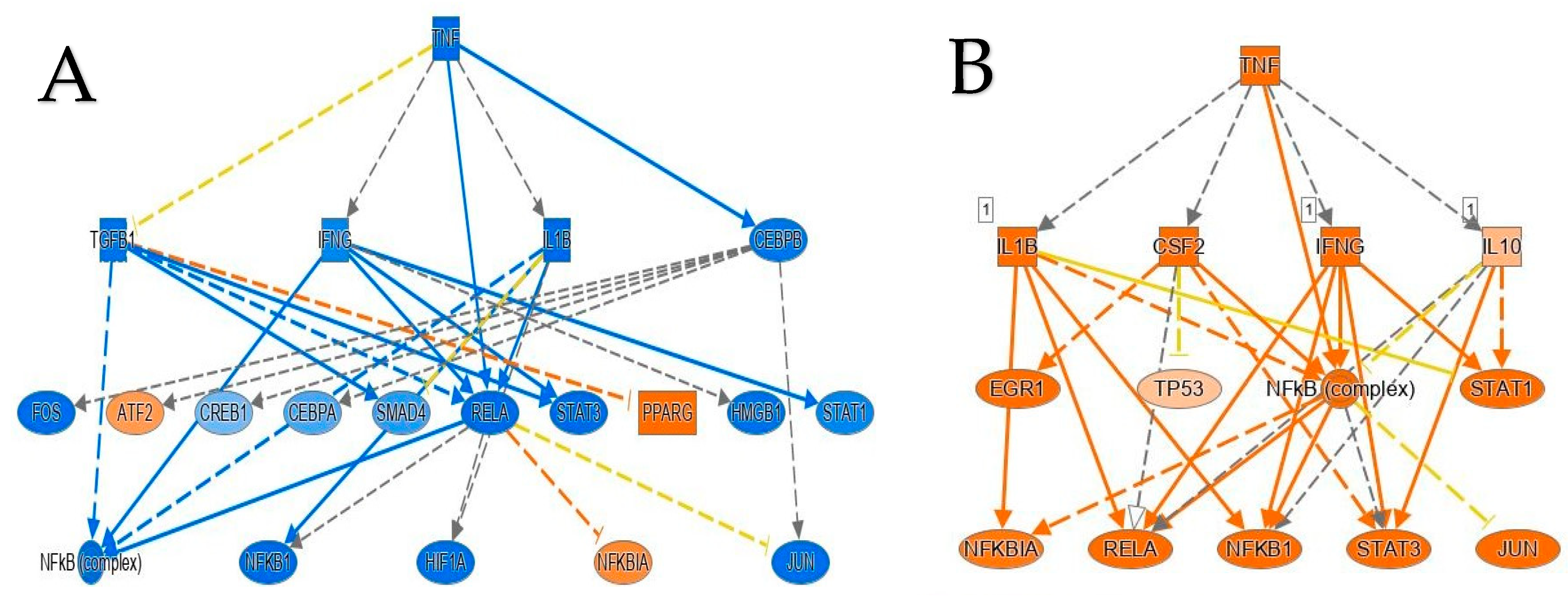
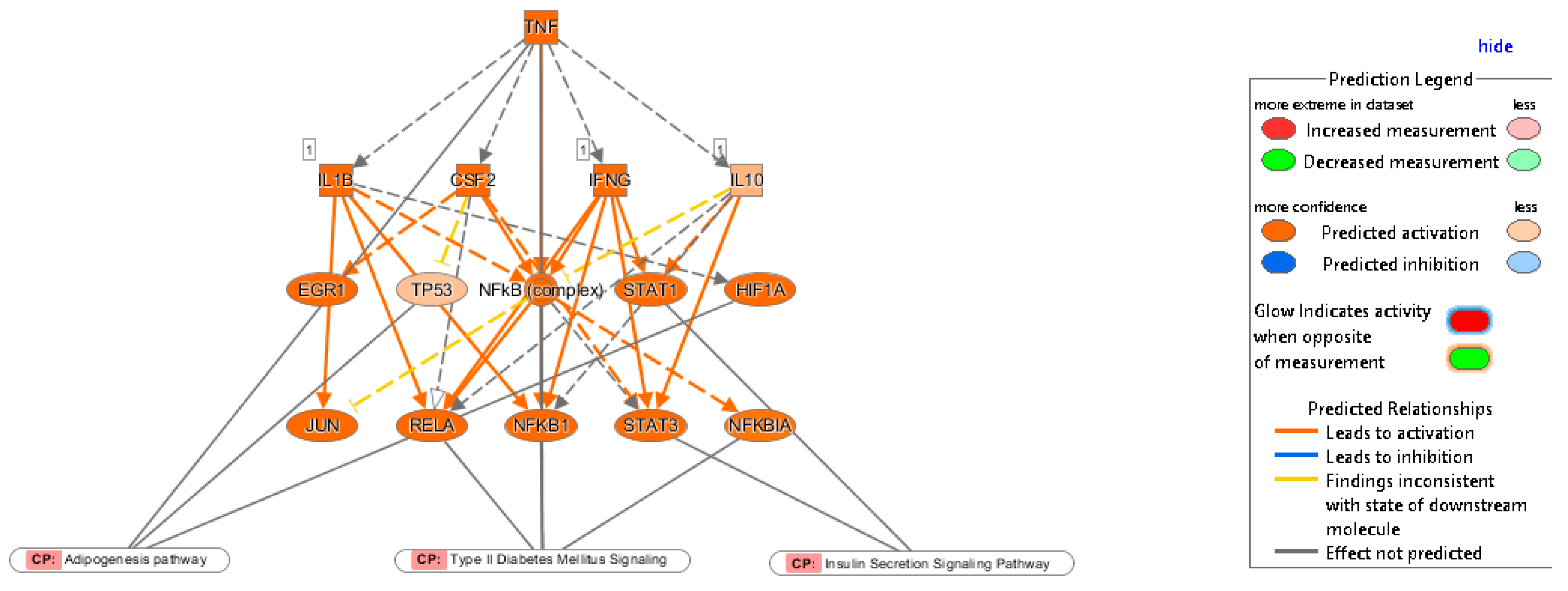
2.3. Multiple Affected Signaling Network in Obese IR-NGT and Obese T2D Compared to Lean Subjects
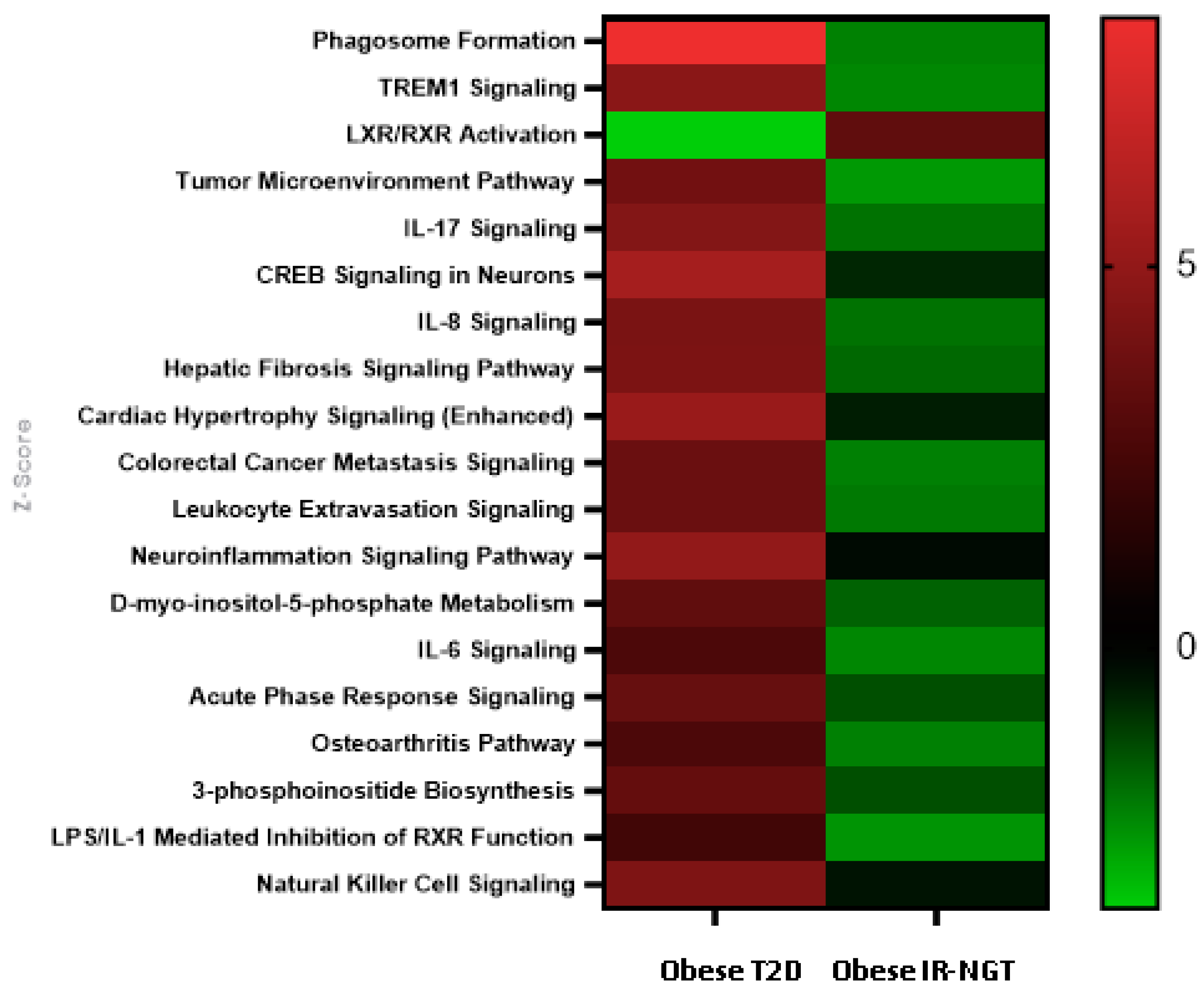
2.4. Identification of Significantly Enriched Canonical Signaling Pathways in Obese IR-NGT and Obese T2D in Comparison with Lean
3. Discussion
4. Materials and Methods
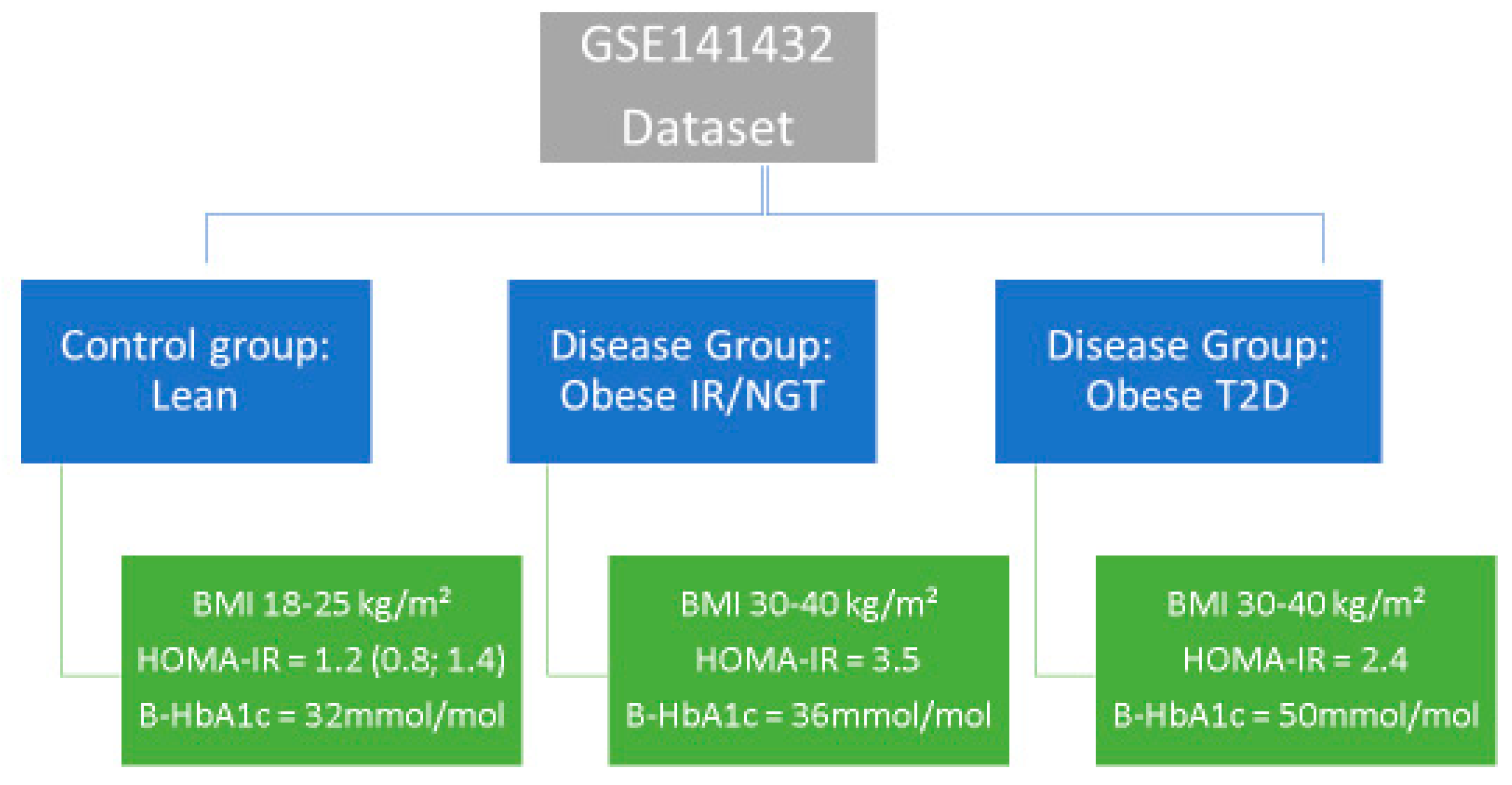
4.1. Source of Data
4.2. Next-Generation Sequencing Data
4.3. IPA Pathway Enrichment Analysis of DEGs
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sarma, S.; Sockalingam, S.; Dash, S. Obesity as a multisystem disease: Trends in obesity rates and obesity-related complications. Diabetes Obes. Metab. 2021, 23, 3–16. [Google Scholar] [CrossRef] [PubMed]
- Castorani, V.; Polidori, N.; Giannini, C.; Blasetti, A.; Chiarelli, F. Insulin resistance and type 2 diabetes in children. Ann. Pediatr. Endocrinol. Metab. 2020, 25, 217–226. [Google Scholar] [CrossRef] [PubMed]
- Dilworth, L.; Facey, A.; Omoruyi, F. Diabetes Mellitus and Its Metabolic Complications: The Role of Adipose Tissues. Int. J. Mol. Sci. 2021, 22, 7644. [Google Scholar] [CrossRef] [PubMed]
- Czaja-Stolc, S.; Potrykus, M.; Stankiewicz, M.; Kaska, Ł.; Małgorzewicz, S. Pro-Inflammatory Profile of Adipokines in Obesity Contributes to Pathogenesis, Nutritional Disorders, and Cardiovascular Risk in Chronic Kidney Disease. Nutrients 2022, 14, 1457. [Google Scholar] [CrossRef] [PubMed]
- Thouvenot, K.; Turpin, T.; Taïlé, J.; Clément, K.; Meilhac, O.; Gonthier, M.P. Links between Insulin Resistance and Periodontal Bacteria: Insights on Molecular Players and Therapeutic Potential of Polyphenols. Biomolecules 2022, 12, 378. [Google Scholar] [CrossRef]
- Nedosugova, L.V.; Markina, Y.V.; Bochkareva, L.A.; Kuzina, I.A.; Petunina, N.A.; Yudina, I.Y.; Kirichenko, T.V. Inflammatory Mechanisms of Diabetes and Its Vascular Complications. Biomedicines 2022, 10, 1168. [Google Scholar] [CrossRef]
- Kim, J.; Lee, J. Role of obesity-induced inflammation in the development of insulin resistance and type 2 diabetes: History of the research and remaining questions. Ann. Pediatr. Endocrinol. Metab. 2021, 26, 1. [Google Scholar] [CrossRef]
- Feng, Z.; Du, Z.; Shu, X.; Zhu, L.; Wu, J.; Gao, Q.; Wang, L.; Chen, N.; Li, Y.; Luo, M.; et al. Role of RAGE in obesity-induced adipose tissue inflammation and insulin resistance. Cell Death Discov. 2021, 7, 305. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wang, F.; Imani, S.; Tao, L.; Deng, Y.; Cai, Y. Natural Killer Cells: Friend or Foe in Metabolic Diseases? Front. Immunol. 2021, 12, 614429. [Google Scholar] [CrossRef]
- Li, M.; Chi, X.; Wang, Y.; Setrerrahmane, S.; Xie, W.; Xu, H. Trends in insulin resistance: Insights into mechanisms and therapeutic strategy. Signal Transduct. Target. Ther. 2022, 7, 216. [Google Scholar] [CrossRef]
- Gómez-Hernández, A.; Beneit, N.; Díaz-Castroverde, S.; Escribano, Ó. Differential Role of Adipose Tissues in Obesity and Related Metabolic and Vascular Complications. Int. J. Endocrinol. 2016, 2016, 1216783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Machado, S.A.; Pasquarelli-do-Nascimento, G.; da Silva, D.S.; Farias, G.R.; de Oliveira Santos, I.; Baptista, L.B.; Magalhães, K.G. Browning of the white adipose tissue regulation: New insights into nutritional and metabolic relevance in health and diseases. Nutr. Metab. 2022, 19, 61. [Google Scholar] [CrossRef] [PubMed]
- Majnarić, L.T.; Bosnić, Z.; Štefanić, M.; Wittlinger, T. Cross-Talk between the Cytokine IL-37 and Thyroid Hormones in Modulating Chronic Inflammation Associated with Target Organ Damage in Age-Related Metabolic and Vascular Conditions. Int. J. Mol. Sci. 2022, 23, 6456. [Google Scholar] [CrossRef]
- Chan, P.C.; Hsieh, P.S. The Role and Regulatory Mechanism of Brown Adipose Tissue Activation in Diet-Induced Thermogenesis in Health and Diseases. Int. J. Mol. Sci. 2022, 23, 9448. [Google Scholar] [CrossRef] [PubMed]
- Hayden, M.R. The Mighty Mitochondria Are Unifying Organelles and Metabolic Hubs in Multiple Organs of Obesity, Insulin Resistance, Metabolic Syndrome, and Type 2 Diabetes: An Observational Ultrastructure Study. Int. J. Mol. Sci. 2022, 23, 4820. [Google Scholar] [CrossRef]
- Zhang, K.; Pan, H.; Wang, L.; Yang, H.; Zhu, H.; Gong, F. Adipose Tissue Insulin Resistance is Closely Associated with Metabolic Syndrome in Northern Chinese Populations. Diabetes Metab. Syndr. Obes. Targets Ther. 2021, 14, 1117–1128. [Google Scholar] [CrossRef]
- Guilherme, A.; Virbasius, J.V.; Puri, V.; Czech, M.P. Adipocyte dysfunctions linking obesity to insulin resistance and type 2 diabetes. Nat. Rev. Mol. Cell Biol. 2008, 9, 367–377. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Wu, Y.; Rong, X.; Zheng, C.; Guo, J. Anti-Lipolysis Induced by Insulin in Diverse Pathophysiologic Conditions of Adipose Tissue. Diabetes Metab. Syndr. Obes. 2020, 13, 1575–1585. [Google Scholar] [CrossRef] [PubMed]
- Richard, A.J.; White, U.; Elks, C.M.; Stephens, J.M. Adipose Tissue: Physiology to Metabolic Dysfunction; Endotext [Internet]; MDText.com, Inc.: South Dartmouth, MA, USA, 2020. [Google Scholar]
- Schmitz-Peiffer, C.; Biden, T.J. Protein kinase C function in muscle, liver, and beta-cells and its therapeutic implications for type 2 diabetes. Diabetes 2008, 57, 1774–1783. [Google Scholar] [CrossRef] [Green Version]
- Boden, G. Fatty acid—Induced inflammation and insulin resistance in skeletal muscle and liver. Curr. Diabetes Rep. 2006, 6, 177–181. [Google Scholar] [CrossRef]
- Jornayvaz, F.R.; Shulman, G.I. Diacylglycerol activation of protein kinase Cε and hepatic insulin resistance. Cell Metab. 2012, 15, 574–584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tonyan, Z.N.; Nasykhova, Y.A.; Danilova, M.M.; Barbitoff, Y.A.; Changalidi, A.I.; Mikhailova, A.A.; Glotov, A.S. Overview of Transcriptomic Research on Type 2 Diabetes: Challenges and Perspectives. Genes 2022, 13, 1176. [Google Scholar] [CrossRef] [PubMed]
- Clough, E.; Barrett, T. The Gene Expression Omnibus Database. Methods Mol. Biol. 2016, 1418, 93–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.; Domrachev, M.; Lash, A.E. Gene Expression Omnibus: NCBI gene expression and hybridization array data repository. Nucleic Acids Res. 2002, 30, 207–210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fryk, E.; Olausson, J.; Mossberg, K.; Strindberg, L.; Schmelz, M.; Brogren, H.; Gan, L.-M.; Piazza, S.; Provenzani, A.; Becattini, B.; et al. Hyperinsulinemia and insulin resistance in the obese may develop as part of a homeostatic response to elevated free fatty acids: A mechanistic case-control and a population-based cohort study. EBioMedicine 2021, 65, 103264. [Google Scholar] [CrossRef]
- Bódis, K.; Roden, M. Energy metabolism of white adipose tissue and insulin resistance in humans. Eur. J. Clin. Investig. 2018, 48, e13017. [Google Scholar] [CrossRef] [Green Version]
- Smith, U.; Kahn, B.B. Adipose tissue regulates insulin sensitivity: Role of adipogenesis, de novo lipogenesis and novel lipids. J. Intern. Med. 2016, 280, 465–475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vargas, E.; Joy, N.V.; Sepulveda, M.A.C. Biochemistry, Insulin Metabolic Effects; StatPearls Publishing: Treasure Island, FL, USA, 2021. [Google Scholar]
- Laffitte, B.A.; Chao, L.C.; Li, J.; Walczak, R.; Hummasti, S.; Joseph, S.B.; Castrillo, A.; Wilpitz, D.C.; Mangelsdorf, D.J.; Collins, J.L.; et al. Activation of liver X receptor improves glucose tolerance through coordinate regulation of glucose metabolism in liver and adipose tissue. Proc. Natl. Acad. Sci. USA 2003, 100, 5419–5424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaillant, A.A.J.; Qurie, A. Interleukin. In StatPearls; StatPearls Publishing LLC: Treasure Island, FL, USA, 2021. [Google Scholar]
- Pan, P.; Liu, X.; Wu, L.; Li, X.; Wang, K.; Wang, X.; Zhou, X.; Long, Y.; Liu, D.; Xie, L.; et al. TREM-1 promoted apoptosis and inhibited autophagy in LPS-treated HK-2 cells through the NF-κB pathway. Int. J. Med. Sci. 2021, 18, 8–17. [Google Scholar] [CrossRef]
- Baranov, M.V.; Kumar, M.; Sacanna, S.; Thutupalli, S.; van den Bogaart, G. Modulation of Immune Responses by Particle Size and Shape. Front. Immunol. 2021, 11, 3854. [Google Scholar] [CrossRef]
- Berbudi, A.; Rahmadika, N.; Tjahjadi, A.I.; Ruslami, R. Type 2 Diabetes and its Impact on the Immune System. Curr. Diabetes Rev. 2020, 16, 442–449. [Google Scholar] [CrossRef] [PubMed]
- Eckel, R.H.; Kahn, S.E.; Ferrannini, E.; Goldfine, A.B.; Nathan, D.M.; Schwartz, M.W.; Smith, R.J.; Smith, S.R. Obesity and type 2 diabetes: What can be unified and what needs to be individualized? J. Clin. Endocrinol. Metab. 2011, 96, 1654–1663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Golay, A.; Ybarra, J. Link between obesity and type 2 diabetes. Best Pr. Res. Clin. Endocrinol. Metab. 2005, 19, 649–663. [Google Scholar] [CrossRef] [PubMed]
- Kern, L.; Mittenbühler, M.J.; Vesting, A.J.; Ostermann, A.L.; Wunderlich, C.M.; Wunderlich, F.T. Obesity-Induced TNFα and IL-6 Signaling: The Missing Link between Obesity and Inflammation—Driven Liver and Colorectal Cancers. Cancers 2018, 11, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Howard, B.V. Insulin resistance and lipid metabolism. Am. J. Cardiol. 1999, 84, 28–32. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Gu, X.; Yi, S. Ingenuity Pathway Analysis of Gene Expression Profiles in Distal Nerve Stump following Nerve Injury: Insights into Wallerian Degeneration. Front. Cell. Neurosci. 2016, 10, 274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shao, Z.; Wang, K.; Zhang, S.; Yuan, J.; Liao, X.; Wu, C.; Zou, Y.; Ha, Y.; Shen, Z.; Guo, J.; et al. Ingenuity pathway analysis of differentially expressed genes involved in signaling pathways and molecular networks in RhoE gene-edited cardiomyocytes. Int. J. Mol. Med. 2020, 46, 1225–1238. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Top Upregulated DEGs Obese IR-NGT | Top Downregulated DEGs Obese IR-NGT | ||||
|---|---|---|---|---|---|
| Name | Log2 Fold Change | Description | Name | Log2 Fold Change | Description |
| RNF17 | 5.66 | ring finger protein 17 | DCD | −7.77 | dermcidin |
| CTAG2 | 4.83 | cancer/testis antigen 2 | MUCL1 | −6.66 | mucin-like 1 |
| TBC1D3K | 4.69 | TBC1 domain family member 3K | SCGB2A2 | −6.66 | secretoglobin family 2A member 2 |
| TBC1D3E | 4.13 | TBC1 domain family member 3E | SCGB1D2 | −6.19 | secretoglobin family 1D member 2 |
| DEUP1 | 3.95 | deuterosome assembly protein 1 | MSLN | −5.09 | mesothelin |
| TMEM215 | 3.77 | transmembrane protein 215 | BORCS7-ASMT | −4.76 | BORCS7-ASMT readthrough (NMD candidate) |
| CEACAM20 | 3.66 | CEA cell adhesion molecule 20 | NTSR2 | −4.66 | neurotensin receptor 2 |
| C2orf83 | 3.42 | chromosome 2 open reading frame 83 | CA6 | −4.66 | carbonic anhydrase 6 |
| DPYSL4 | 3.37 | dihydropyrimidinase-like 4 | EEF1E1-BLOC1S5 | −4.08 | EEF1E1-BLOC1S5 readthrough (NMD candidate) |
| ZNF723 | 3.24 | zinc finger protein 723 | ZPBP | −3.78 | zona pellucida binding protein |
| PMCH | 3.23 | pro-melanin concentrating hormone | ABHD16B | −3.57 | abhydrolase domain containing 16B |
| RPP21 | 3.2 | ribonuclease P/MRP subunit p21 | GABRR3 | −3.53 | gamma-aminobutyric acid type A receptor subunit rho3 |
| TRIM39-RPP21 | 3.15 | TRIM39-RPP21 readthrough | RASSF6 | −3.38 | Ras association domain family member 6 |
| COMP | 3.14 | cartilage oligomeric matrix protein | SPX | −3.27 | spexin hormone |
| HOXD10 | 3.07 | homeobox D10 | IQCA1L | −3.24 | IQ motif containing AAA domain 1-like |
| CCDC54 | 3.06 | coiled-coil domain containing 54 | CYP1A2 | −3.12 | cytochrome P450 family 1 subfamily A member 2 |
| Top Upregulated DEGs Obese T2D | Top Downregulated DEGs Obese T2D | ||||
|---|---|---|---|---|---|
| Name | Log2 Fold Change | Description | Name | Log2 Fold Change | Description |
| HBG2 | 5.14 | hemoglobin subunit gamma 2 | TMEM52 | −1.05 | transmembrane protein 52 |
| AC008763.3 | 5.04 | alpha hemoglobin stabilizing protein | CFAP74 | −2.84 | cilia and flagella associated protein 74 |
| HBD | 4.45 | hemoglobin subunit delta | GABRD | −3.07 | gamma-aminobutyric acid type A receptor subunit delta |
| DEFA1B | 4.1 | defensin alpha 1B | ARHGEF16 | −1.26 | Rho guanine nucleotide exchange factor 16 |
| AC139530.2 | 4.07 | 5′-aminolevulinate synthase 2 | TAS1R1 | −1.11 | taste 1 receptor member 1 |
| ARMH2 | 3.8 | armadillo-like helical domain containing 2 | CA6 | −2.62 | carbonic anhydrase 6 |
| MAG | 3.7 | myelin-associated glycoprotein | SLC25A34 | −1.4 | solute carrier family 25 member 34 |
| TMEM215 | 3.62 | transmembrane protein 215 | CLCNKB | −1.31 | chloride voltage-gated channel Kb |
| AC104389.6 | 3.62 | prokineticin 2 | PLA2G5 | −1.05 | phospholipase A2 group V |
| PTPRN | 3.59 | protein tyrosine phosphatase receptor type N | GRIK3 | −1.53 | glutamate ionotropic receptor kainate type subunit 3 |
| FAM72C | 3.53 | family with sequence similarity 72 member C | GJA9 | −1.87 | gap junction protein alpha 9 |
| POPDC3 | 3.53 | popeye domain containing 3 | NT5C1A | −1.34 | 5′-nucleotidase, cytosolic IA |
| LCN1 | 3.53 | lipocalin 1 | HYI | −1.12 | hydroxypyruvate isomerase (putative) |
| NPFF | 3.53 | neuropeptide FF-amide peptide precursor | AGBL4 | −1.47 | ATP/GTP binding protein-like 4 |
| HBA1 | 3.51 | hemoglobin subunit alpha 2 | ELAVL4 | −1.25 | ELAV-like RNA binding protein 4 |
| IZUMO3 | 3.48 | IZUMO family member 3 | CDKN2C | −1.62 | cyclin-dependent kinase inhibitor 2C |
| LY6G6F | 3.42 | lymphocyte antigen 6 family member G6F | GLIS1 | −1.41 | GLIS family zinc finger 1 |
| EGFL6 | 3.38 | EGF-like domain multiple 6 | FOXD3 | −1.77 | forkhead box D3 |
| CAMP | 3.37 | cathelicidin antimicrobial peptide | TTLL7 | −1.1 | tubulin tyrosine ligase-like 7 |
| HBB | 3.37 | hemoglobin subunit beta | MCOLN3 | −1.01 | mucolipin TRP cation channel 3 |
| AC034102.2 | 3.37 | chromosome 2 open reading frame 83 | GBP7 | −2.33 | guanylate-binding protein 7 |
| ITLN1 | 3.29 | intelectin 1 | UBL4B | −1.3 | ubiquitin-like 4B |
| COMP | 3.21 | cartilage oligomeric matrix protein | CHIA | −2.71 | chitinase acidic |
| GYPB | 3.2 | glycophorin B (MNS blood group) | CASQ2 | −1.77 | calsequestrin 2 |
| SLC4A1 | 3.19 | solute carrier family 4 member 1 (Diego blood group) | PHGDH | −1.49 | phosphoglycerate dehydrogenase |
| TRIM10 | 3.18 | tripartite motif containing 10 | CIART | −1.01 | circadian-associated repressor of transcription |
| HBQ1 | 3.17 | hemoglobin subunit theta 1 | RORC | −1.43 | RAR-related orphan receptor C |
| DUSP13 | 3.16 | dual specificity phosphatase 13 | S100A1 | −1.74 | S100 calcium-binding protein A1 |
| KLF1 | 3.13 | Kruppel-like factor 1 | NUP210L | −1.13 | nucleoporin 210-like |
| KCNA10 | 3.13 | potassium voltage-gated channel subfamily A member 10 | DCST2 | −1.03 | DC-STAMP domain containing 2 |
| JCHAIN | 3.1 | joining chain of multimeric IgA and IgM | NHLH1 | −2.31 | nescient helix-loop-helix 1 |
| CMTM2 | 3.1 | CKLF-like MARVEL transmembrane domain containing 2 | TSTD1 | −1.09 | thiosulfate sulfurtransferase-like domain containing 1 |
| CA1 | 3.07 | carbonic anhydrase 1 | SPATA46 | −1.57 | spermatogenesis associated 46 |
| KRT72 | 3.05 | keratin 72 | FAM78B | −1.41 | family with sequence similarity 78 member B |
| HBM | 3.05 | hemoglobin subunit mu | MAEL | −2.1 | maelstrom spermatogenic transposon silencer |
| S100P | 3.01 | S100 calcium-binding protein P | SLC19A2 | −1.05 | solute carrier family 19 member 2 |
| MMP7 | 3.01 | matrix metallopeptidase 7 | AXDND1 | −1.4 | axonemal dynein light-chain domain containing 1 |
| RIPPLY2 | 3.01 | ripply transcriptional repressor 2 | GLUL | −1.53 | glutamate-ammonia ligase |
| CBLIF | 3.01 | cobalamin binding intrinsic factor | ADORA1 | −1.22 | adenosine A1 receptor |
| TMEM132D | 3 | transmembrane protein 132D | LEFTY2 | −1.91 | left-right determination factor 2 |
| ZNF723 | 2.99 | zinc finger protein 723 | COQ8A | −1.1 | coenzyme Q8A |
| KLRC4 | 2.99 | killer cell lectin-like receptor C4 | TRIM67 | −1.35 | Novel protein |
| FCGR3B | 2.98 | Fc fragment of IgG receptor IIIb | NTSR2 | −4.63 | tripartite motif containing 67 |
| IFIT1B | 2.97 | interferon-induced protein with tetratricopeptide repeats 1B | LPIN1 | −1.59 | neurotensin receptor 2 |
| FUT7 | 2.96 | fucosyltransferase 7 | VSNL1 | −1.79 | lipin 1 |
| S100A8 | 2.95 | S100 calcium-binding protein A8 | APOB | −2.26 | visinin-like 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Errafii, K.; Boujraf, S.; Chikri, M. Transcriptomic Analysis from Normal Glucose Tolerance to T2D of Obese Individuals Using Bioinformatic Tools. Int. J. Mol. Sci. 2023, 24, 6337. https://doi.org/10.3390/ijms24076337
Errafii K, Boujraf S, Chikri M. Transcriptomic Analysis from Normal Glucose Tolerance to T2D of Obese Individuals Using Bioinformatic Tools. International Journal of Molecular Sciences. 2023; 24(7):6337. https://doi.org/10.3390/ijms24076337
Chicago/Turabian StyleErrafii, Khaoula, Said Boujraf, and Mohamed Chikri. 2023. "Transcriptomic Analysis from Normal Glucose Tolerance to T2D of Obese Individuals Using Bioinformatic Tools" International Journal of Molecular Sciences 24, no. 7: 6337. https://doi.org/10.3390/ijms24076337