
Specialized Ribosomes in Health and Disease
 , , , and
, , , and
Abstract
:1. Introduction
2. The Mammalian Ribosome
2.1. Eukaryotic Ribosomes Represent the Expansion of Bacteria and Archaeal Ribosomes
2.2. Eukaryotic Ribosome Subunit Assembly Occurs in the Nucleolus
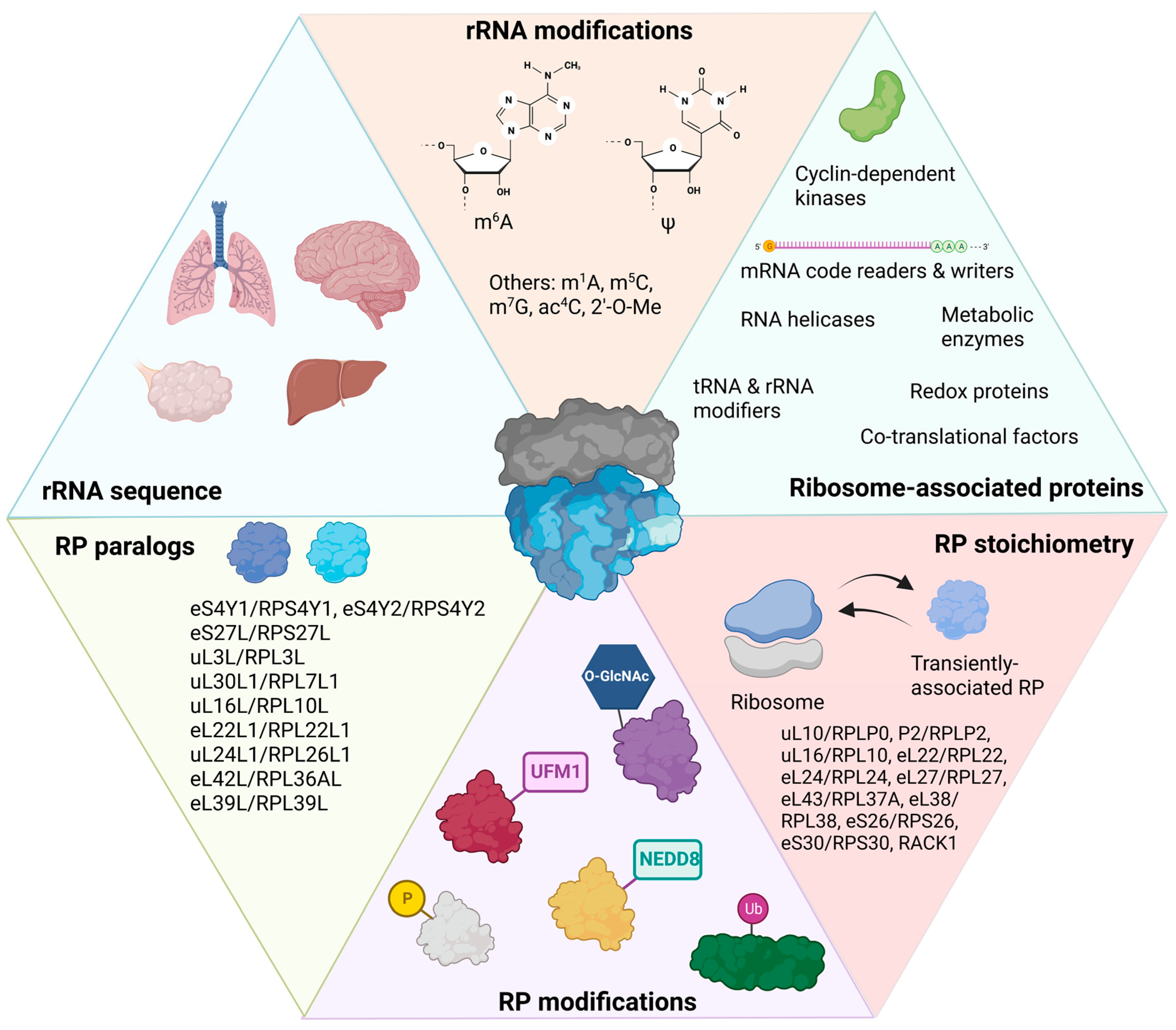
2.3. RPs and rRNAs Contain Many Post-Translational or Post-Transcriptional Modifications
2.3.1. Ribosomal RNA Post-Transcriptional Modifications
2.3.2. Ribosomal Protein Post-Translational Modifications
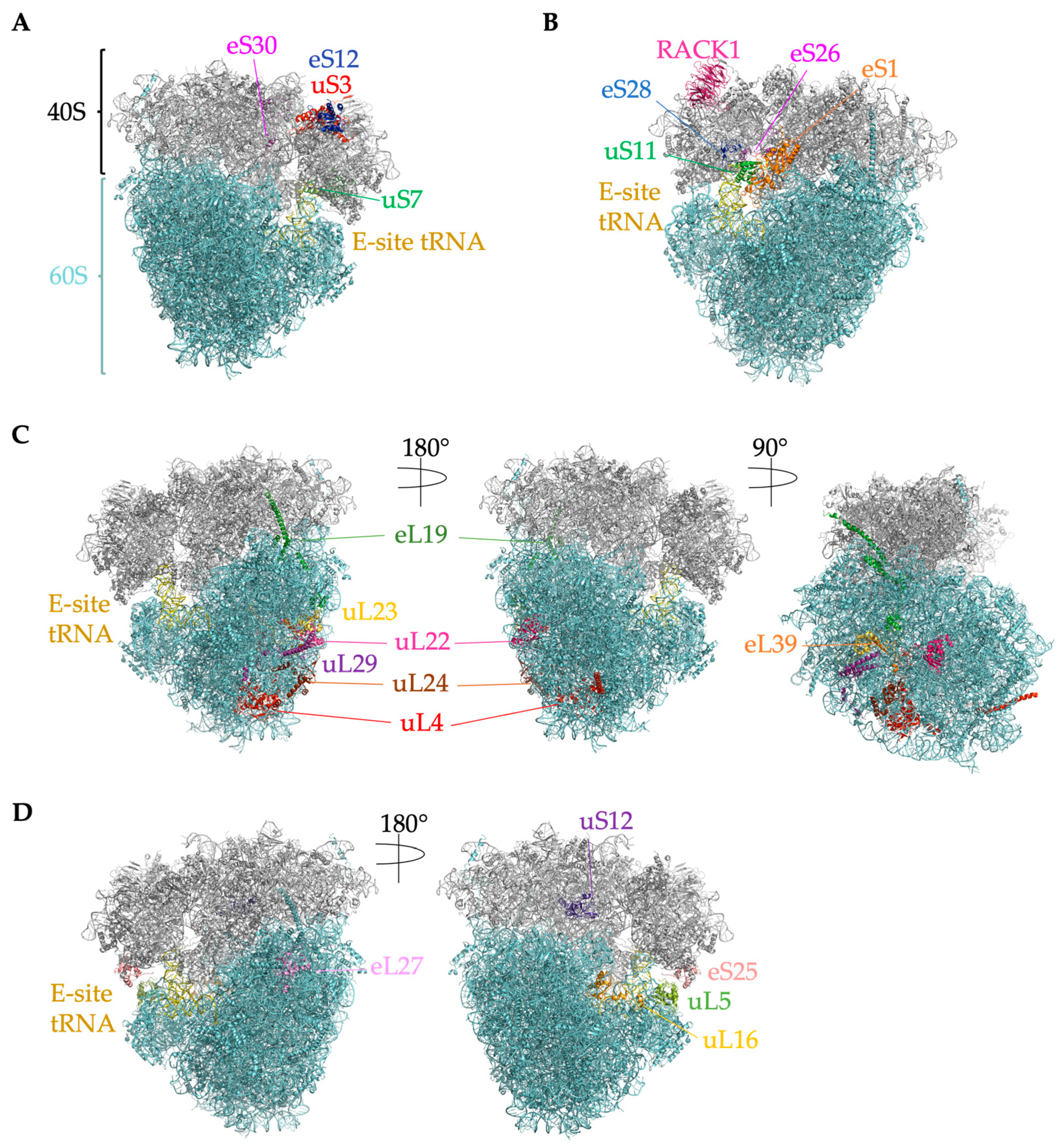
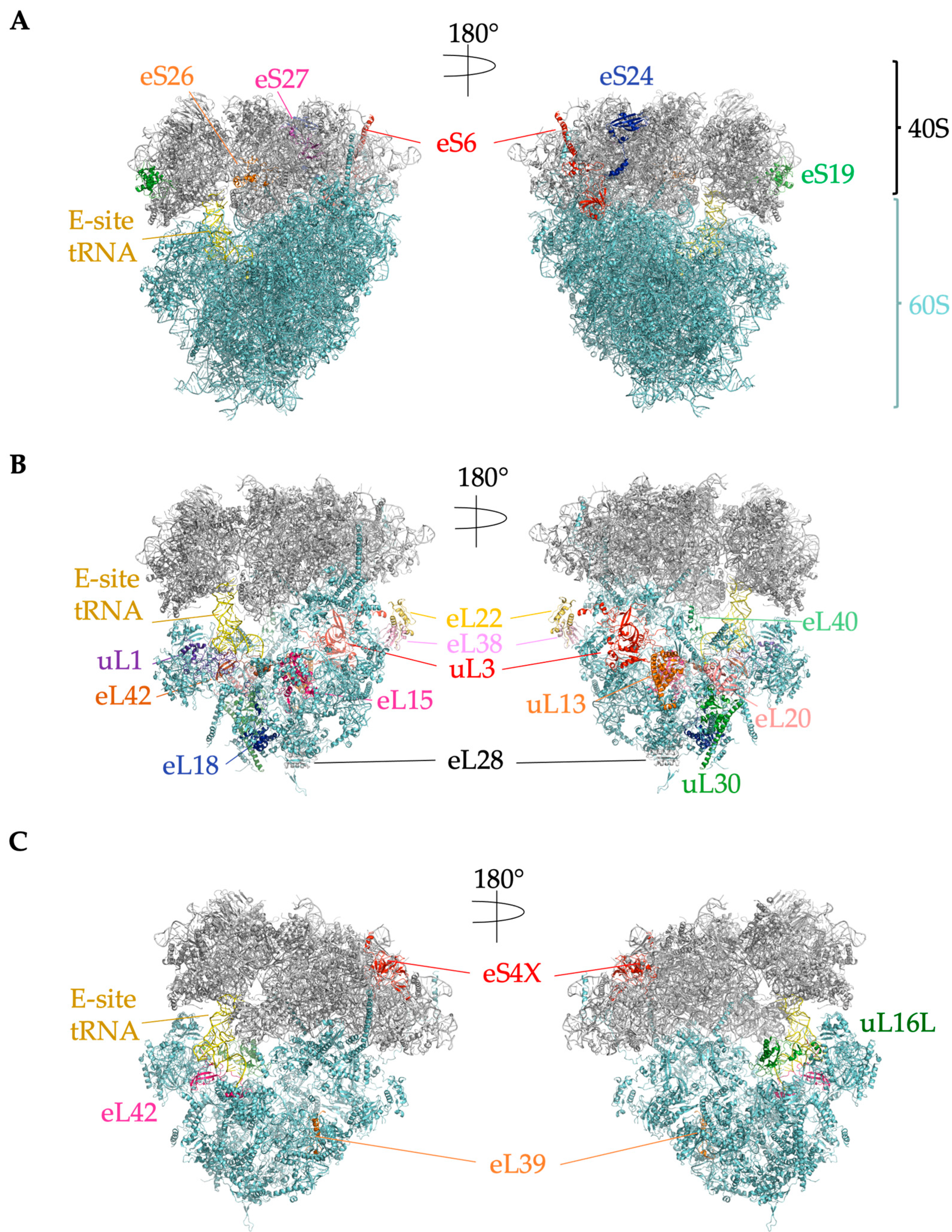
2.4. Ribosome-Associated Proteins
3. Endoplasmic Reticulum-Associated Ribosomes Are Different from Cytosolic Ribosomes
4. Cell and Tissue Ribosomal Heterogeneity in Mammals
4.1. Ribosomal Heterogeneity in Adult Tissues
4.1.1. RP mRNA Signatures in the Gastrointestinal Tract
4.1.2. RP mRNA Signatures in the Endocrine System
4.1.3. RP mRNA Signatures in the Reproductive System
4.1.4. RP mRNA Signatures in the Respiratory and Circulatory Systems
4.1.5. RP mRNA Signature in Skeletal Muscle
4.1.6. RP mRNA Signatures in the Immune System
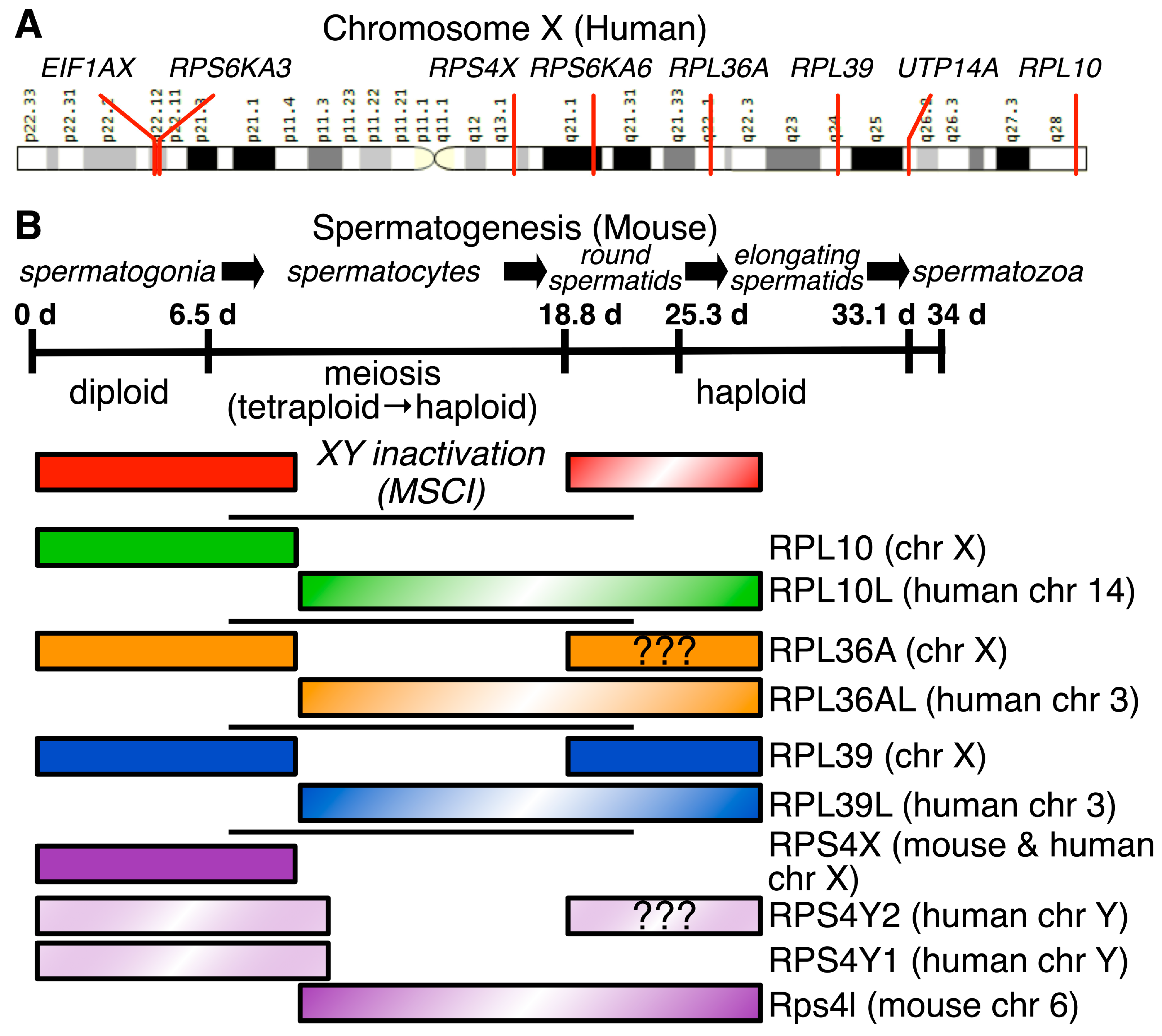
4.2. X-Linked Genes Influence Ribosomal Structure in Males
4.2.1. Male Germ Cells in Mammals Express Paralogs of Several X-Linked Ribosomal Protein Genes
4.2.2. uL16/RPL10, eL42/RPL36A, and eL39/RPL39 Paralogs Are Expressed during and after Male Meiosis
4.2.3. eL39L/RPL39L Is a Component of Specialized Male Germ Cell Ribosomes That Guides Proper Folding of Nascent Germ Cell Proteins
4.2.4. RPS4X Has Two Y-Chromosomal Paralogs in Primates, but Only a Single Autosomal Paralog in Other Mammals
4.2.5. RPS6KA3, RPS6KA6, UTP14A and EIF1AX Are X-Linked Ribosome-Modifying Protein Genes That Have Important Paralogs Expressed in Male Germ Cells
5. Disease-Associated Ribosomal Heterogeneity and Specialization
5.1. Ribosomopathies
5.2. Ribosome Heterogeneity in the Nervous System and in Neurological Diseases
5.3. Ribosomal Proteins and Cancer
5.3.1. Glioblastoma
5.3.2. Neuroblastoma
5.3.3. Breast Cancer
5.3.4. Colorectal Cancer
5.3.5. Liver Cancer
5.3.6. Melanoma
5.3.7. Rhabdomyosarcoma
5.4. The Immune System and Viral Ribosome Remodeling
5.4.1. Viral Proteins Bind to Ribosomes
5.4.2. Viral Translation Depends on RP Modifications
6. Perspectives on Ribosome Specialization in Human Disease
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| rRNA | ribosomal RNA |
| RP | ribosomal protein |
| RPS | small subunit ribosomal protein in mammals (old naming system) |
| RPL | large subunit ribosomal protein in mammals (old naming system) |
| uL | large subunit ribosomal protein with bacteria/archaea and eukarya homologs |
| uS | small subunit ribosomal protein with bacteria/archaea and eukarya homologs |
| eL | large subunit ribosomal protein in eukaryotes |
| eS | small subunit ribosomal protein in eukaryotes |
| ER | endoplasmic reticulum |
References
- Geiger, T.; Wehner, A.; Schaab, C.; Cox, J.; Mann, M. Comparative Proteomic Analysis of Eleven Common Cell Lines Reveals Ubiquitous but Varying Expression of Most Proteins. Mol. Cell. Proteom. 2012, 11, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Genuth, N.R.; Barna, M. The Discovery of Ribosome Heterogeneity and Its Implications for Gene Regulation and Organismal Life. Mol. Cell 2018, 71, 364–374. [Google Scholar] [CrossRef] [Green Version]
- Kampen, K.R.; Sulima, S.O.; Vereecke, S.; De Keersmaecker, K. Hallmarks of ribosomopathies. Nucleic Acids Res. 2020, 48, 1013–1028. [Google Scholar] [CrossRef] [PubMed]
- Guimaraes, J.C.; Zavolan, M. Patterns of ribosomal protein expression specify normal and malignant human cells. Genome Biol. 2016, 17, 236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panda, A.; Yadav, A.; Yeerna, H.; Singh, A.; Biehl, M.; Lux, M.; Schulz, A.; Klecha, T.; Doniach, S.; Khiabanian, H.; et al. Tissue- and development-stage-specific mRNA and heterogeneous CNV signatures of human ribosomal proteins in normal and cancer samples. Nucleic Acids Res. 2020, 48, 7079–7098. [Google Scholar] [CrossRef]
- Li, H.; Huo, Y.; He, X.; Yao, L.; Zhang, H.; Cui, Y.; Xiao, H.; Xie, W.; Zhang, D.; Wang, Y.; et al. A male germ-cell-specific ribosome controls male fertility. Nature 2022, 612, 725–731. [Google Scholar] [CrossRef]
- Barna, M.; Karbstein, K.; Tollervey, D.; Ruggero, D.; Brar, G.; Greer, E.L.; Dinman, J.D. The promises and pitfalls of specialized ribosomes. Mol. Cell 2022, 82, 2179–2184. [Google Scholar] [CrossRef]
- Genuth, N.R.; Barna, M. Heterogeneity and specialized functions of translation machinery: From genes to organisms. Nat. Rev. Genet. 2018, 19, 431–452. [Google Scholar] [CrossRef]
- Gerst, J.E. Pimp My Ribosome: Ribosomal Protein Paralogs Specify Translational Control. Trends Genet. 2018, 34, 832–845. [Google Scholar] [CrossRef]
- Mauro, V.P.; Matsuda, D. Translation regulation by ribosomes: Increased complexity and expanded scope. RNA Biol. 2016, 13, 748–755. [Google Scholar] [CrossRef] [Green Version]
- Dinman, J.D. Pathways to Specialized Ribosomes: The Brussels Lecture. J. Mol. Biol. 2016, 428, 2186–2194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferretti, M.B.; Karbstein, K. Does functional specialization of ribosomes really exist? Rna 2019, 25, 521–538. [Google Scholar] [CrossRef] [PubMed]
- Barna, M. Specialized Ribosomes: A New Frontier in Gene Regulation, Organismal Biology, & Evolution. FASEB J. 2017, 31, 387.1. [Google Scholar]
- Petelski, A.A.; Slavov, N. Analyzing Ribosome Remodeling in Health and Disease. Proteomics 2020, 20, e2000039. [Google Scholar] [CrossRef]
- Slavov, N. Counting protein molecules for single-cell proteomics. Cell 2022, 185, 232–234. [Google Scholar] [CrossRef] [PubMed]
- Melnikov, S.; Ben-Shem, A.; Garreau de Loubresse, N.; Jenner, L.; Yusupova, G.; Yusupov, M. One core, two shells: Bacterial and eukaryotic ribosomes. Nat. Struct. Mol. Biol. 2012, 19, 560–567. [Google Scholar] [CrossRef] [PubMed]
- Ben-Shem, A.; Garreau de Loubresse, N.; Melnikov, S.; Jenner, L.; Yusupova, G.; Yusupov, M. The structure of the eukaryotic ribosome at 3.0 A resolution. Science 2011, 334, 1524–1529. [Google Scholar] [CrossRef] [Green Version]
- Wilson, D.N.; Doudna Cate, J.H. The structure and function of the eukaryotic ribosome. Cold Spring Harb. Perspect. Biol. 2012, 4, a011536. [Google Scholar] [CrossRef]
- Hariharan, N.; Ghosh, S.; Palakodeti, D. The story of rRNA expansion segments: Finding functionality amidst diversity. Wiley Interdiscip. Rev.-Rna 2022, 14, e1732. [Google Scholar] [CrossRef]
- Parks, M.M.; Kurylo, C.M.; Dass, R.A.; Bojmar, L.; Lyden, D.; Vincent, C.T.; Blanchard, S.C. Variant ribosomal RNA alleles are conserved and exhibit tissue-specific expression. Sci. Adv. 2018, 4, eaao0665. [Google Scholar] [CrossRef] [Green Version]
- Leppek, K.; Fujii, K.; Quade, N.; Susanto, T.T.; Boehringer, D.; Lenarcic, T.; Xue, S.; Genuth, N.R.; Ban, N.; Barna, M. Gene- and Species-Specific Hox mRNA Translation by Ribosome Expansion Segments. Mol. Cell 2020, 80, 980–995. [Google Scholar] [CrossRef] [PubMed]
- Khatter, H.; Myasnikov, A.G.; Natchiar, S.K.; Klaholz, B.P. Structure of the human 80S ribosome. Nature 2015, 520, 640–645. [Google Scholar] [CrossRef]
- Gulen, B.; Petrov, A.S.; Okafor, C.D.; Wood, D.V.; O’Neill, E.B.; Hud, N.V.; Williams, L.D. Ribosomal small subunit domains radiate from a central core. Sci. Rep. 2016, 6, 20885. [Google Scholar] [CrossRef] [Green Version]
- Ban, N.; Beckmann, R.; Cate, J.H.; Dinman, J.D.; Dragon, F.; Ellis, S.R.; Lafontaine, D.L.; Lindahl, L.; Liljas, A.; Lipton, J.M.; et al. A new system for naming ribosomal proteins. Curr. Opin. Struct. Biol. 2014, 24, 165–169. [Google Scholar] [CrossRef] [Green Version]
- Shi, Z.; Fujii, K.; Kovary, K.M.; Genuth, N.R.; Röst, H.L.; Teruel, M.N.; Barna, M. Heterogeneous Ribosomes Preferentially Translate Distinct Subpools of mRNAs Genome-wide. Mol. Cell 2017, 67, 71–83.e77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barros-Silva, D.; Klavert, J.; Jenster, G.; Jerónimo, C.; Lafontaine, D.L.J.; Martens-Uzunova, E.S. The role of OncoSnoRNAs and Ribosomal RNA 2′-O-methylation in Cancer. RNA Biol. 2021, 18, 61–74. [Google Scholar] [CrossRef] [PubMed]
- Karamysheva, Z.N.; Tikhonova, E.B.; Grozdanov, P.N.; Huffman, J.C.; Baca, K.R.; Karamyshev, A.; Denison, R.B.; MacDonald, C.C.; Zhang, K.; Karamyshev, A.L. Polysome Profiling in Leishmania, Human Cells and Mouse Testis. Jove-J. Vis. Exp. 2018, 2018, e57600. [Google Scholar]
- Sugihara, Y.; Honda, H.; Iida, T.; Morinaga, T.; Hino, S.; Okajima, T.; Matsuda, T.; Nadano, D. Proteomic analysis of rodent ribosomes revealed heterogeneity including ribosomal proteins L10-like, L22-like 1, and L39-like. J. Proteome Res. 2010, 9, 1351–1366. [Google Scholar] [CrossRef]
- Gupta, V.; Warner, J.R. Ribosome-omics of the human ribosome. Rna 2014, 20, 1004–1013. [Google Scholar] [CrossRef] [Green Version]
- Chaillou, T.; Zhang, X.P.; McCarthy, J.J. Expression of Muscle-Specific Ribosomal Protein L3-Like Impairs Myotube Growth. J. Cell. Physiol. 2016, 231, 1894–1902. [Google Scholar] [CrossRef] [Green Version]
- Uechi, T.; Maeda, N.; Tanaka, T.; Kenmochi, N. Functional second genes generated by retrotransposition of the X-linked ribosomal protein genes. Nucleic Acids Res. 2002, 30, 5369–5375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larionova, T.D.; Bastola, S.; Aksinina, T.E.; Anufrieva, K.S.; Wang, J.; Shender, V.O.; Andreev, D.E.; Kovalenko, T.F.; Arapidi, G.P.; Shnaider, P.V.; et al. Alternative RNA splicing modulates ribosomal composition and determines the spatial phenotype of glioblastoma cells. Nat. Cell Biol. 2022, 24, 1541–1557. [Google Scholar] [CrossRef] [PubMed]
- Zou, Q.X.; Qi, H.Y. Deletion of ribosomal paralogs Rpl39 and Rpl39l compromises cell proliferation via protein synthesis and mitochondrial activity. Int. J. Biochem. Cell Biol. 2021, 139, 106070. [Google Scholar] [CrossRef] [PubMed]
- Baßler, J.; Hurt, E. Eukaryotic Ribosome Assembly. Annu. Rev. Biochem. 2019, 88, 281–306. [Google Scholar] [CrossRef]
- Pena, C.; Hurt, E.; Panse, V.G. Eukaryotic ribosome assembly, transport and quality control. Nat. Struct. Mol. Biol. 2017, 24, 689–699. [Google Scholar] [CrossRef]
- Natchiar, S.K.; Myasnikov, A.G.; Kratzat, H.; Hazemann, I.; Klaholz, B.P. Visualization of chemical modifications in the human 80S ribosome structure. Nature 2017, 551, 472–477. [Google Scholar] [CrossRef]
- Taoka, M.; Nobe, Y.; Yamaki, Y.; Sato, K.; Ishikawa, H.; Izumikawa, K.; Yamauchi, Y.; Hirota, K.; Nakayama, H.; Takahashi, N.; et al. Landscape of the complete RNA chemical modifications in the human 80S ribosome. Nucleic Acids Res. 2018, 46, 9289–9298. [Google Scholar] [CrossRef]
- Motorin, Y.; Quinternet, M.; Rhalloussi, W.; Marchand, V. Constitutive and variable 2′-O-methylation (Nm) in human ribosomal RNA. Rna Biol. 2021, 18, 88–97. [Google Scholar] [CrossRef]
- Cui, L.; Ma, R.; Cai, J.; Guo, C.; Chen, Z.; Yao, L.; Wang, Y.; Fan, R.; Wang, X.; Shi, Y. RNA modifications: Importance in immune cell biology and related diseases. Signal Transduct. Target. Ther. 2022, 7, 334. [Google Scholar] [CrossRef]
- Schosserer, M.; Minois, N.; Angerer, T.B.; Amring, M.; Dellago, H.; Harreither, E.; Calle-Perez, A.; Pircher, A.; Gerstl, M.P.; Pfeifenberger, S.; et al. Methylation of ribosomal RNA by NSUN5 is a conserved mechanism modulating organismal lifespan. Nat. Commun. 2015, 6, 6158. [Google Scholar] [CrossRef] [Green Version]
- Simsek, D.; Tiu, G.C.; Flynn, R.A.; Byeon, G.W.; Leppek, K.; Xu, A.F.; Chang, H.Y.; Barna, M. The Mammalian Ribo-interactome Reveals Ribosome Functional Diversity and Heterogeneity. Cell 2017, 169, 1051–1065. [Google Scholar] [CrossRef] [Green Version]
- Bohlen, J.; Roiuk, M.; Teleman, A.A. Phosphorylation of ribosomal protein S6 differentially affects mRNA translation based on ORF length. Nucleic Acids Res. 2021, 49, 13062–13074. [Google Scholar] [CrossRef] [PubMed]
- Ruvinsky, I.; Sharon, N.; Lerer, T.; Cohen, H.; Stolovich-Rain, M.; Nir, T.; Dor, Y.; Zisman, P.; Meyuhas, O. Ribosomal protein S6 phosphorylation is a determinant of cell size and glucose homeostasis. Genes Dev. 2005, 19, 2199–2211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruvinsky, I.; Katz, M.; Dreazen, A.; Gielchinsky, Y.; Saada, A.; Freedman, N.; Mishani, E.; Zimmerman, G.; Kasir, J.; Meyuhas, O. Mice deficient in ribosomal protein S6 phosphorylation suffer from muscle weakness that reflects a growth defect and energy deficit. PLoS ONE 2009, 4, e5618. [Google Scholar] [CrossRef] [Green Version]
- Karagöz, G.E.; Acosta-Alvear, D.; Walter, P. The Unfolded Protein Response: Detecting and Responding to Fluctuations in the Protein-Folding Capacity of the Endoplasmic Reticulum. Cold Spring Harb. Perspect. Biol. 2019, 11, a033886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Komander, D.; Rape, M. The ubiquitin code. Annu. Rev. Biochem. 2012, 81, 203–229. [Google Scholar] [CrossRef] [Green Version]
- Higgins, R.; Gendron, J.M.; Rising, L.; Mak, R.; Webb, K.; Kaiser, S.E.; Zuzow, N.; Riviere, P.; Yang, B.; Fenech, E.; et al. The Unfolded Protein Response Triggers Site-Specific Regulatory Ubiquitylation of 40S Ribosomal Proteins. Mol. Cell 2015, 59, 35–49. [Google Scholar] [CrossRef] [Green Version]
- Walczak, C.P.; Leto, D.E.; Zhang, L.; Riepe, C.; Muller, R.Y.; DaRosa, P.A.; Ingolia, N.T.; Elias, J.E.; Kopito, R.R. Ribosomal protein RPL26 is the principal target of UFMylation. Proc. Natl. Acad. Sci. USA 2019, 116, 1299–1308. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Xu, Y.; Rogers, H.; Saidi, L.; Noguchi, C.T.; Li, H.; Yewdell, J.W.; Guydosh, N.R.; Ye, Y. UFMylation of RPL26 links translocation-associated quality control to endoplasmic reticulum protein homeostasis. Cell Res. 2020, 30, 5–20. [Google Scholar] [CrossRef]
- Xirodimas, D.P.; Sundqvist, A.; Nakamura, A.; Shen, L.; Botting, C.; Hay, R.T. Ribosomal proteins are targets for the NEDD8 pathway. EMBO Rep. 2008, 9, 280–286. [Google Scholar] [CrossRef] [Green Version]
- Ohn, T.; Kedersha, N.; Hickman, T.; Tisdale, S.; Anderson, P. A functional RNAi screen links O-GlcNAc modification of ribosomal proteins to stress granule and processing body assembly. Nat. Cell Biol. 2008, 10, 1224–1231. [Google Scholar] [CrossRef] [Green Version]
- Zachara, N.E.; Hart, G.W. Cell signaling, the essential role of O-GlcNAc! Biochim. Biophys. Acta 2006, 1761, 599–617. [Google Scholar] [CrossRef]
- Chen, E.; Sharma, M.R.; Shi, X.; Agrawal, R.K.; Joseph, S. Fragile X mental retardation protein regulates translation by binding directly to the ribosome. Mol. Cell 2014, 54, 407–417. [Google Scholar] [CrossRef] [Green Version]
- Lauria, F.; Bernabo, P.; Tebaldi, T.; Groen, E.J.N.; Perenthaler, E.; Maniscalco, F.; Rossi, A.; Donzel, D.; Clamer, M.; Marchioretto, M.; et al. SMN-primed ribosomes modulate the translation of transcripts related to spinal muscular atrophy. Nat. Cell Biol. 2020, 22, 1239–1251. [Google Scholar] [CrossRef]
- Shao, S.; Brown, A.; Santhanam, B.; Hegde, R.S. Structure and assembly pathway of the ribosome quality control complex. Mol. Cell 2015, 57, 433–444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wild, K.; Aleksic, M.; Lapouge, K.; Juaire, K.D.; Flemming, D.; Pfeffer, S.; Sinning, I. MetAP-like Ebp1 occupies the human ribosomal tunnel exit and recruits flexible rRNA expansion segments. Nat. Commun. 2020, 11, 776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karamyshev, A.L.; Patrick, A.E.; Karamysheva, Z.N.; Griesemer, D.S.; Hudson, H.; Tjon-Kon-Sang, S.; Nilsson, I.; Otto, H.; Liu, Q.; Rospert, S.; et al. Inefficient SRP Interaction with a Nascent Chain Triggers a mRNA Quality Control Pathway. Cell 2014, 156, 146–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinarbasi, E.S.; Karamyshev, A.L.; Tikhonova, E.B.; Wu, I.H.; Hudson, H.; Thomas, P.J. Pathogenic Signal Sequence Mutations in Progranulin Disrupt SRP Interactions Required for mRNA Stability. Cell Rep. 2018, 23, 2844–2851. [Google Scholar] [CrossRef]
- Tikhonova, E.B.; Karamysheva, Z.N.; von Heijne, G.; Karamyshev, A.L. Silencing of Aberrant Secretory Protein Expression by Disease-Associated Mutations. J. Mol. Biol. 2019, 431, 2567–2580. [Google Scholar] [CrossRef]
- Tikhonova, E.B.; Gutierrez Guarnizo, S.A.; Kellogg, M.K.; Karamyshev, A.; Dozmorov, I.M.; Karamysheva, Z.N.; Karamyshev, A.L. Defective Human SRP Induces Protein Quality Control and Triggers Stress Response. J. Mol. Biol. 2022, 434, 167832. [Google Scholar] [CrossRef]
- Karamyshev, A.L.; Karamysheva, Z.N. Lost in Translation: Ribosome-Associated mRNA and Protein Quality Controls. Front. Genet. 2018, 9, 431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karamyshev, A.L.; Tikhonova, E.B.; Karamysheva, Z.N. Translational Control of Secretory Proteins in Health and Disease. Int. J. Mol. Sci. 2020, 21, 2538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, I.J.; Wang, I.A.; Tai, L.R.; Lin, A. The role of expansion segment of human ribosomal protein L35 in nuclear entry, translation activity, and endoplasmic reticulum docking. Biochem. Cell Biol. 2008, 86, 271–277. [Google Scholar] [CrossRef] [PubMed]
- Bhaskar, V.; Desogus, J.; Graff-Meyer, A.; Schenk, A.D.; Cavadini, S.; Chao, J.A. Dynamic association of human Ebp1 with the ribosome. Rna 2021, 27, 411–419. [Google Scholar] [CrossRef]
- Jiang, N.; Hu, L.J.; Liu, C.N.; Gao, X.L.; Zheng, S.M. 60S ribosomal protein L35 regulates beta-casein translational elongation and secretion in bovine mammary epithelial cells. Arch. Biochem. Biophys. 2015, 583, 130–139. [Google Scholar] [CrossRef] [PubMed]
- Zinnall, U.; Milek, M.; Minia, I.; Vieira-Vieira, C.H.; Müller, S.; Mastrobuoni, G.; Hazapis, O.-G.; Del Giudice, S.; Schwefel, D.; Bley, N.; et al. HDLBP binds ER-targeted mRNAs by multivalent interactions to promote protein synthesis of transmembrane and secreted proteins. Nat. Commun. 2022, 13, 2727. [Google Scholar] [CrossRef]
- Bachtrog, D.; Kirkpatrick, M.; Mank, J.E.; McDaniel, S.F.; Pires, J.C.; Rice, W.; Valenzuela, N. Are all sex chromosomes created equal? Trends Genet. TIG 2011, 27, 350–357. [Google Scholar] [CrossRef]
- Posynick, B.J.; Brown, C.J. Escape From X-Chromosome Inactivation: An Evolutionary Perspective. Front. Cell Dev. Biol. 2019, 7, 241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.E.; Vibranovski, M.D.; Landback, P.; Marais, G.A.; Long, M. Chromosomal redistribution of male-biased genes in mammalian evolution with two bursts of gene gain on the X chromosome. PLoS Biol. 2010, 8, e1000494. [Google Scholar] [CrossRef]
- Lee, J.T. The X as model for RNA’s niche in epigenomic regulation. Cold Spring Harb. Perspect. Biol. 2010, 2, a003749. [Google Scholar] [CrossRef] [Green Version]
- Handel, M.A. The XY body: An attractive chromatin domain. Biol. Reprod. 2020, 102, 985–987. [Google Scholar] [CrossRef]
- Yan, W.; McCarrey, J.R. Sex chromosome inactivation in the male. Epigenetics 2009, 4, 452–456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, P.J. X chromosomes, retrogenes and their role in male reproduction. Trends Endocrinol. Metab. 2004, 15, 79–83. [Google Scholar] [CrossRef] [PubMed]
- Lahn, B.T.; Page, D.C. Four evolutionary strata on the human X chromosome. Science 1999, 286, 964–967. [Google Scholar] [CrossRef] [Green Version]
- Namekawa, S.H.; Park, P.J.; Zhang, L.F.; Shima, J.E.; McCarrey, J.R.; Griswold, M.D.; Lee, J.T. Postmeiotic sex chromatin in the male germline of mice. Curr. Biol. 2006, 16, 660–667. [Google Scholar] [CrossRef] [Green Version]
- Vemuganti, S.A.; de Villena, F.P.; O’Brien, D.A. Frequent and recent retrotransposition of orthologous genes plays a role in the evolution of sperm glycolytic enzymes. BMC Genom. 2010, 11, 285. [Google Scholar] [CrossRef] [Green Version]
- Lopes, A.M.; Miguel, R.N.; Sargent, C.A.; Ellis, P.J.; Amorim, A.; Affara, N.A. The human RPS4 paralogue on Yq11.223 encodes a structurally conserved ribosomal protein and is preferentially expressed during spermatogenesis. BMC Mol. Biol. 2010, 11, 33. [Google Scholar] [CrossRef] [Green Version]
- Fisher, E.M.; Beer-Romero, P.; Brown, L.G.; Ridley, A.; McNeil, J.A.; Lawrence, J.B.; Willard, H.F.; Bieber, F.R.; Page, D.C. Homologous ribosomal protein genes on the human X and Y chromosomes: Escape from X inactivation and possible implications for Turner syndrome. Cell 1990, 63, 1205–1218. [Google Scholar] [CrossRef] [PubMed]
- Andres, O.; Kellermann, T.; Lopez-Giraldez, F.; Rozas, J.; Domingo-Roura, X.; Bosch, M. RPS4Y gene family evolution in primates. BMC Evol. Biol. 2008, 8, 142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, L.; Li, T.; Zhang, X.; Zhang, B.; Yu, C.; Li, Y.; Fan, S.; Jiang, X.; Khan, T.; Hao, Q.; et al. RPL10L Is Required for Male Meiotic Division by Compensating for RPL10 during Meiotic Sex Chromosome Inactivation in Mice. Curr. Biol. 2017, 27, 1498–1505.e6. [Google Scholar] [CrossRef] [Green Version]
- Shi, K.; Wang, B.; Dou, L.; Wang, S.; Fu, X.; Yu, H. Integrated bioinformatics analysis of the transcription factor-mediated gene regulatory networks in the formation of spermatogonial stem cells. Front. Physiol. 2022, 13, 949486. [Google Scholar] [CrossRef]
- Nadano, D.; Notsu, T.; Matsuda, T.; Sato, T. A human gene encoding a protein homologous to ribosomal protein L39 is normally expressed in the testis and derepressed in multiple cancer cells. Biochim. Biophys. Acta 2002, 1577, 430–436. [Google Scholar] [CrossRef] [PubMed]
- Rohozinski, J.; Lamb, D.J.; Bishop, C.E. UTP14c is a recently acquired retrogene associated with spermatogenesis and fertility in man. Biol. Reprod. 2006, 74, 644–651. [Google Scholar] [CrossRef] [PubMed]
- Rohozinski, J. Lineage-independent retrotransposition of UTP14 associated with male fertility has occurred multiple times throughout mammalian evolution. R. Soc. Open Sci. 2017, 4, 171049. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Ray, D.; Ye, P. Identification of germ cell-specific genes in mammalian meiotic prophase. BMC Bioinform. 2013, 14, 72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hughes, J.F.; Skaletsky, H.; Koutseva, N.; Pyntikova, T.; Page, D.C. Sex chromosome-to-autosome transposition events counter Y-chromosome gene loss in mammals. Genome Biol. 2015, 16, 104. [Google Scholar] [CrossRef] [Green Version]
- Godfrey, A.K.; Naqvi, S.; Chmatal, L.; Chick, J.M.; Mitchell, R.N.; Gygi, S.P.; Skaletsky, H.; Page, D.C. Quantitative analysis of Y-Chromosome gene expression across 36 human tissues. Genome Res. 2020, 30, 860–873. [Google Scholar] [CrossRef]
- Kenmochi, N.; Kawaguchi, T.; Rozen, S.; Davis, E.; Goodman, N.; Hudson, T.J.; Tanaka, T.; Page, D.C. A map of 75 human ribosomal protein genes. Genome Res. 1998, 8, 509–523. [Google Scholar] [CrossRef] [Green Version]
- Uechi, T.; Tanaka, T.; Kenmochi, N. A complete map of the human ribosomal protein genes: Assignment of 80 genes to the cytogenetic map and implications for human disorders. Genomics 2001, 72, 223–230. [Google Scholar] [CrossRef]
- Pollutri, D.; Penzo, M. Ribosomal Protein L10: From Function to Dysfunction. Cells 2020, 9, 2503. [Google Scholar] [CrossRef]
- Brooks, S.S.; Wall, A.L.; Golzio, C.; Reid, D.W.; Kondyles, A.; Willer, J.R.; Botti, C.; Nicchitta, C.V.; Katsanis, N.; Davis, E.E. A novel ribosomopathy caused by dysfunction of RPL10 disrupts neurodevelopment and causes X-linked microcephaly in humans. Genetics 2014, 198, 723–733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bourque, D.K.; Hartley, T.; Nikkel, S.M.; Pohl, D.; Tetreault, M.; Kernohan, K.D.; Care4Rare Canada Consortium; Dyment, D.A. A de novo mutation in RPL10 causes a rare X-linked ribosomopathy characterized by syndromic intellectual disability and epilepsy: A new case and review of the literature. Eur. J. Med. Genet. 2018, 61, 89–93. [Google Scholar] [CrossRef] [PubMed]
- Tu, C.; Meng, L.; Nie, H.; Yuan, S.; Wang, W.; Du, J.; Lu, G.; Lin, G.; Tan, Y.Q. A homozygous RPL10L missense mutation associated with male factor infertility and severe oligozoospermia. Fertil. Steril. 2020, 113, 561–568. [Google Scholar] [CrossRef]
- Baouz, S.; Woisard, A.; Sinapah, S.; Le Caer, J.P.; Argentini, M.; Bulygin, K.; Aguie, G.; Hountondji, C. The human large subunit ribosomal protein L36A-like contacts the CCA end of P-site bound tRNA. Biochimie 2009, 91, 1420–1425. [Google Scholar] [CrossRef] [PubMed]
- Hountondji, C.; Bulygin, K.; Woisard, A.; Tuffery, P.; Crechet, J.B.; Pech, M.; Nierhaus, K.H.; Karpova, G.; Baouz, S. Lys53 of ribosomal protein L36AL and the CCA end of a tRNA at the P/E hybrid site are in close proximity on the human ribosome. Chembiochem 2012, 13, 1791–1797. [Google Scholar] [CrossRef] [PubMed]
- Myasnikov, A.G.; Kundhavai Natchiar, S.; Nebout, M.; Hazemann, I.; Imbert, V.; Khatter, H.; Peyron, J.F.; Klaholz, B.P. Structure-function insights reveal the human ribosome as a cancer target for antibiotics. Nat. Commun. 2016, 7, 12856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Obaide, M.A.; Al, O., II; Vasylyeva, T.L. The potential consequences of bidirectional promoter methylation on GLA and HNRNPH2 expression in Fabry disease phenotypes in a family of patients carrying a GLA deletion variant. Biomed. Rep. 2022, 17, 71. [Google Scholar] [CrossRef] [PubMed]
- Rohozinski, J.; Anderson, M.L.; Broaddus, R.E.; Edwards, C.L.; Bishop, C.E. Spermatogenesis associated retrogenes are expressed in the human ovary and ovarian cancers. PLoS ONE 2009, 4, e5064. [Google Scholar] [CrossRef] [Green Version]
- Wong, Q.W.; Li, J.; Ng, S.R.; Lim, S.G.; Yang, H.; Vardy, L.A. RPL39L is an example of a recently evolved ribosomal protein paralog that shows highly specific tissue expression patterns and is upregulated in ESCs and HCC tumors. RNA Biol. 2014, 11, 33–41. [Google Scholar] [CrossRef] [Green Version]
- Zou, Q.; Yang, L.; Shi, R.; Qi, Y.; Zhang, X.; Qi, H. Proteostasis regulated by testis-specific ribosomal protein RPL39L maintains mouse spermatogenesis. iScience 2021, 24, 103396. [Google Scholar] [CrossRef]
- Micic, J.; Rodriguez-Galan, O.; Babiano, R.; Fitzgerald, F.; Fernandez-Fernandez, J.; Zhang, Y.; Gao, N.; Woolford, J.L.; de la Cruz, J. Ribosomal protein eL39 is important for maturation of the nascent polypeptide exit tunnel and proper protein folding during translation. Nucleic Acids Res. 2022, 50, 6453–6473. [Google Scholar] [CrossRef] [PubMed]
- Skaletsky, H.; Kuroda-Kawaguchi, T.; Minx, P.J.; Cordum, H.S.; Hillier, L.; Brown, L.G.; Repping, S.; Pyntikova, T.; Ali, J.; Bieri, T.; et al. The male-specific region of the human Y chromosome is a mosaic of discrete sequence classes. Nature 2003, 423, 825–837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trombetta, B.; D’Atanasio, E.; Cruciani, F. Patterns of Inter-Chromosomal Gene Conversion on the Male-Specific Region of the Human Y Chromosome. Front. Genet. 2017, 8, 54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sugihara, Y.; Sadohara, E.; Yonezawa, K.; Kugo, M.; Oshima, K.; Matsuda, T.; Nadano, D. Identification and expression of an autosomal paralogue of ribosomal protein S4, X-linked, in mice: Potential involvement of testis-specific ribosomal proteins in translation and spermatogenesis. Gene 2013, 521, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Chauvin, C.; Koka, V.; Nouschi, A.; Mieulet, V.; Hoareau-Aveilla, C.; Dreazen, A.; Cagnard, N.; Carpentier, W.; Kiss, T.; Meyuhas, O.; et al. Ribosomal protein S6 kinase activity controls the ribosome biogenesis transcriptional program. Oncogene 2014, 33, 474–483. [Google Scholar] [CrossRef]
- Merienne, K.; Jacquot, S.; Pannetier, S.; Zeniou, M.; Bankier, A.; Gecz, J.; Mandel, J.L.; Mulley, J.; Sassone-Corsi, P.; Hanauer, A. A missense mutation in RPS6KA3 (RSK2) responsible for non-specific mental retardation. Nat. Genet. 1999, 22, 13–14. [Google Scholar] [CrossRef]
- Yntema, H.G.; van den Helm, B.; Kissing, J.; van Duijnhoven, G.; Poppelaars, F.; Chelly, J.; Moraine, C.; Fryns, J.P.; Hamel, B.C.; Heilbronner, H.; et al. A novel ribosomal S6-kinase (RSK4; RPS6KA6) is commonly deleted in patients with complex X-linked mental retardation. Genomics 1999, 62, 332–343. [Google Scholar] [CrossRef]
- Abel, M.H.; Baban, D.; Lee, S.; Charlton, H.M.; O’Shaughnessy, P.J. Effects of FSH on testicular mRNA transcript levels in the hypogonadal mouse. J. Mol. Endocrinol. 2009, 42, 291–303. [Google Scholar] [CrossRef] [Green Version]
- Black, J.J.; Wang, Z.; Goering, L.M.; Johnson, A.W. Utp14 interaction with the small subunit processome. RNA 2018, 24, 1214–1228. [Google Scholar] [CrossRef]
- Rohozinski, J.; Bishop, C.E. The mouse juvenile spermatogonial depletion (jsd) phenotype is due to a mutation in the X-derived retrogene, mUtp14b. Proc. Natl. Acad. Sci. USA 2004, 101, 11695–11700. [Google Scholar] [CrossRef] [Green Version]
- Bradley, J.; Baltus, A.; Skaletsky, H.; Royce-Tolland, M.; Dewar, K.; Page, D.C. An X-to-autosome retrogene is required for spermatogenesis in mice. Nat. Genet. 2004, 36, 872–876. [Google Scholar] [CrossRef] [PubMed]
- Köferle, A.; Schlattl, A.; Hormann, A.; Thatikonda, V.; Popa, A.; Spreitzer, F.; Ravichandran, M.C.; Supper, V.; Oberndorfer, S.; Puchner, T.; et al. Interrogation of cancer gene dependencies reveals paralog interactions of autosome and sex chromosome-encoded genes. Cell Rep. 2022, 39, 110636. [Google Scholar] [CrossRef] [PubMed]
- Khajuria, R.K.; Munschauer, M.; Ulirsch, J.C.; Fiorini, C.; Ludwig, L.S.; McFarland, S.K.; Abdulhay, N.J.; Specht, H.; Keshishian, H.; Mani, D.R.; et al. Ribosome Levels Selectively Regulate Translation and Lineage Commitment in Human Hematopoiesis. Cell 2018, 173, 90–103.e19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mills, E.W.; Green, R. Ribosomopathies: There’s strength in numbers. Science 2017, 358, eaan2755. [Google Scholar] [CrossRef] [Green Version]
- Lebaron, S.; O’Donohue, M.F.; Smith, S.C.; Engleman, K.L.; Juusola, J.; Safina, N.P.; Thiffault, I.; Saunders, C.J.; Gleizes, P.E. Functionally impaired RPL8 variants associated with Diamond-Blackfan anemia and a Diamond-Blackfan anemia-like phenotype. Hum. Mutat. 2022, 43, 389–402. [Google Scholar] [CrossRef]
- Tiu, G.C.; Kerr, C.H.; Forester, C.M.; Krishnarao, P.S.; Rosenblatt, H.D.; Raj, N.; Lantz, T.C.; Zhulyn, O.; Bowen, M.E.; Shokat, L.; et al. A p53-dependent translational program directs tissue-selective phenotypes in a model of ribosomopathies. Dev. Cell 2021, 56, 2089–2102.e11. [Google Scholar] [CrossRef]
- Hannan, K.M.; Soo, P.; Wong, M.S.; Lee, J.K.; Hein, N.; Poh, P.; Wysoke, K.D.; Williams, T.D.; Montellese, C.; Smith, L.K.; et al. Nuclear stabilization of p53 requires a functional nucleolar surveillance pathway. Cell Rep. 2022, 41, 111571. [Google Scholar] [CrossRef]
- Nicolas, E.; Parisot, P.; Pinto-Monteiro, C.; de Walque, R.; De Vleeschouwer, C.; Lafontaine, D.L.J. Involvement of human ribosomal proteins in nucleolar structure and p53-dependent nucleolar stress. Nat. Commun. 2016, 7, 11390. [Google Scholar] [CrossRef] [Green Version]
- Takagi, M.; Absalon, M.J.; McLure, K.G.; Kastan, M.B. Regulation of p53 translation and induction after DNA damage by ribosomal protein L26 and nucleolin. Cell 2005, 123, 49–63. [Google Scholar] [CrossRef] [Green Version]
- Perez, J.D.; Fusco, C.M.; Schuman, E.M. A Functional Dissection of the mRNA and Locally Synthesized Protein Population in Neuronal Dendrites and Axons. Annu. Rev. Genet. 2021, 55, 183–207. [Google Scholar] [CrossRef]
- Hafner, A.S.; Donlin-Asp, P.G.; Leitch, B.; Herzog, E.; Schuman, E.M. Local protein synthesis is a ubiquitous feature of neuronal pre- and postsynaptic compartments. Science 2019, 364, eaau3644. [Google Scholar] [CrossRef] [PubMed]
- Biever, A.; Glock, C.; Tushev, G.; Ciirdaeva, E.; Dalmay, T.; Langer, J.D.; Schuman, E.M. Monosomes actively translate synaptic mRNAs in neuronal processes. Science 2020, 367, eaay4991. [Google Scholar] [CrossRef] [PubMed]
- Donlin-Asp, P.G.; Polisseni, C.; Klimek, R.; Heckel, A.; Schuman, E.M. Differential regulation of local mRNA dynamics and translation following long-term potentiation and depression. Proc. Natl. Acad. Sci. USA 2021, 118, e2017578118. [Google Scholar] [CrossRef]
- Fusco, C.M.; Desch, K.; Dorrbaum, A.R.; Wang, M.; Staab, A.; Chan, I.C.W.; Vail, E.; Villeri, V.; Langer, J.D.; Schuman, E.M. Neuronal ribosomes exhibit dynamic and context-dependent exchange of ribosomal proteins. Nat. Commun. 2021, 12, 6127. [Google Scholar] [CrossRef] [PubMed]
- de la Cruz, J.; Karbstein, K.; Woolford, J.L., Jr. Functions of ribosomal proteins in assembly of eukaryotic ribosomes in vivo. Annu. Rev. Biochem. 2015, 84, 93–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puighermanal, E.; Biever, A.; Pascoli, V.; Melser, S.; Pratlong, M.; Cutando, L.; Rialle, S.; Severac, D.; Boubaker-Vitre, J.; Meyuhas, O.; et al. Ribosomal Protein S6 Phosphorylation Is Involved in Novelty-Induced Locomotion, Synaptic Plasticity and mRNA Translation. Front. Mol. Neurosci. 2017, 10, 419. [Google Scholar] [CrossRef] [Green Version]
- Ascano, M.; Mukherjee, N.; Bandaru, P.; Miller, J.B.; Nusbaum, J.D.; Corcoran, D.L.; Langlois, C.; Munschauer, M.; Dewell, S.; Hafner, M.; et al. FMRP targets distinct mRNA sequence elements to regulate protein expression. Nature 2012, 492, 382–386. [Google Scholar] [CrossRef] [Green Version]
- Hale, C.R.; Sawicka, K.; Mora, K.; Fak, J.J.; Kang, J.J.; Cutrim, P.; Cialowicz, K.; Carroll, T.S.; Darnell, R.B. FMRP regulates mRNAs encoding distinct functions in the cell body and dendrites of CA1 pyramidal neurons. Elife 2021, 10, e71892. [Google Scholar] [CrossRef]
- Seo, S.S.; Louros, S.R.; Anstey, N.; Gonzalez-Lozano, M.A.; Harper, C.B.; Verity, N.C.; Dando, O.; Thomson, S.R.; Darnell, J.C.; Kind, P.C.; et al. Excess ribosomal protein production unbalances translation in a model of Fragile X Syndrome. Nat. Commun. 2022, 13, 3236. [Google Scholar] [CrossRef]
- Romano, N.; Di Giacomo, B.; Nobile, V.; Borreca, A.; Willems, D.; Tilesi, F.; Catalani, E.; Agrawal, M.; Welshhans, K.; Ricciardi, S.; et al. Ribosomal RACK1 Regulates the Dendritic Arborization by Repressing FMRP Activity. Int. J. Mol. Sci. 2022, 23, 11857. [Google Scholar] [CrossRef]
- Gabanella, F.; Onori, A.; Ralli, M.; Greco, A.; Passananti, C.; Di Certo, M.G. SMN protein promotes membrane compartmentalization of ribosomal protein S6 transcript in human fibroblasts. Sci. Rep. 2020, 10, 19000. [Google Scholar] [CrossRef] [PubMed]
- Pelletier, J.; Thomas, G.; Volarevic, S. Ribosome biogenesis in cancer: New players and therapeutic avenues. Nat. Rev. Cancer 2018, 18, 51–63. [Google Scholar] [CrossRef]
- Warner, J.R.; McIntosh, K.B. How Common Are Extraribosomal Functions of Ribosomal Proteins? Mol. Cell 2009, 34, 3–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bastide, A.; David, A. The ribosome, (slow) beating heart of cancer (stem) cell. Oncogenesis 2018, 7, 34. [Google Scholar] [CrossRef] [Green Version]
- Babaian, A.; Rothe, K.; Girodat, D.; Minia, I.; Djondovic, S.; Milek, M.; Miko, S.E.S.; Wieden, H.J.; Landthaler, M.; Morin, G.B.; et al. Loss of m(1)acp(3)Psi Ribosomal RNA Modification Is a Major Feature of Cancer. Cell Rep. 2020, 31, 107611. [Google Scholar] [CrossRef] [PubMed]
- Kampen, K.R.; Sulima, S.O.; Verbelen, B.; Girardi, T.; Vereecke, S.; Rinaldi, G.; Verbeeck, J.; Op de Beeck, J.; Uyttebroeck, A.; Meijerink, J.P.P.; et al. The ribosomal RPL10 R98S mutation drives IRES-dependent BCL-2 translation in T-ALL. Leukemia 2019, 33, 319–332. [Google Scholar] [CrossRef]
- Genuth, N.R.; Barna, M. Ribosome specialization in glioblastoma. Nat. Cell Biol. 2022, 24, 1451–1453. [Google Scholar] [CrossRef]
- Brumwell, A.; Fell, L.; Obress, L.; Uniacke, J. Hypoxia influences polysome distribution of human ribosomal protein S12 and alternative splicing of ribosomal protein mRNAs. Rna 2020, 26, 361–371. [Google Scholar] [CrossRef]
- Wu, W.D.; Yu, N.D.; Li, F.; Gao, P.Q.; Lin, S.Y.; Zhu, Y. RPL35 promotes neuroblastoma progression via the enhanced aerobic glycolysis. Am. J. Cancer Res. 2021, 11, 5701–5714. [Google Scholar]
- van Riggelen, J.; Yetil, A.; Felsher, D.W. MYC as a regulator of ribosome biogenesis and protein synthesis. Nat. Rev. Cancer 2010, 10, 301–309. [Google Scholar] [CrossRef]
- Ebright, R.Y.; Lee, S.; Wittner, B.S.; Niederhoffer, K.L.; Nicholson, B.T.; Bardia, A.; Truesdell, S.; Wiley, D.F.; Wesley, B.; Li, S.; et al. Deregulation of ribosomal protein expression and translation promotes breast cancer metastasis. Science 2020, 367, 1468–1473. [Google Scholar] [CrossRef] [PubMed]
- Rong, B.; Zhang, Q.; Wan, J.; Xing, S.; Dai, R.; Li, Y.; Cai, J.; Xie, J.; Song, Y.; Chen, J.; et al. Ribosome 18S m(6)A Methyltransferase METTL5 Promotes Translation Initiation and Breast Cancer Cell Growth. Cell Rep. 2020, 33, 108544. [Google Scholar] [CrossRef] [PubMed]
- Longley, D.B.; Harkin, D.P.; Johnston, P.G. 5-Fluorouracil: Mechanisms of action and clinical strategies. Nat. Rev. Cancer 2003, 3, 330–338. [Google Scholar] [CrossRef] [PubMed]
- Therizols, G.; Bash-Imam, Z.; Panthu, B.; Machon, C.; Vincent, A.; Ripoll, J.; Nait-Slimane, S.; Chalabi-Dchar, M.; Gaucherot, A.; Garcia, M.; et al. Alteration of ribosome function upon 5-fluorouracil treatment favors cancer cell drug-tolerance. Nat. Commun. 2022, 13, 173. [Google Scholar] [CrossRef]
- Peng, H.; Chen, B.; Wei, W.; Guo, S.; Han, H.; Yang, C.; Ma, J.; Wang, L.; Peng, S.; Kuang, M.; et al. N(6)-methyladenosine (m(6)A) in 18S rRNA promotes fatty acid metabolism and oncogenic transformation. Nat. Metab. 2022, 4, 1041–1054. [Google Scholar] [CrossRef]
- Zhang, T.; Sun, L.; Hao, Y.; Suo, C.; Shen, S.; Wei, H.; Ma, W.; Zhang, P.; Wang, T.; Gu, X.; et al. ENO1 suppresses cancer cell ferroptosis by degrading the mRNA of iron regulatory protein 1. Nat. Cancer 2022, 3, 75–89. [Google Scholar] [CrossRef]
- Chang, C.H.; Curtis, J.D.; Maggi, L.B.; Faubert, B.; Villarino, A.V.; O’Sullivan, D.; Huang, S.C.C.; van der Windt, G.J.W.; Blagih, J.; Qiu, J.; et al. Posttranscriptional Control of T Cell Effector Function by Aerobic Glycolysis. Cell 2013, 153, 1239–1251. [Google Scholar] [CrossRef] [Green Version]
- Castello, A.; Fischer, B.; Eichelbaum, K.; Horos, R.; Benedikt, B.; Strein, C.; Davey, N.; Humphreys, D.; Preiss, T.; Steinmetz, L.; et al. Insights into RNA Biology from an Atlas of Mammalian mRNA-Binding Proteins. Cell 2012, 149, 1393–1406. [Google Scholar] [CrossRef] [Green Version]
- Yamada, S.; Kitai, Y.; Tadokoro, T.; Takahashi, R.; Shoji, H.; Maemoto, T.; Ishiura, M.; Muromoto, R.; Kashiwakura, J.; Ishii, K.J.; et al. Identification of RPL15 60S Ribosomal Protein as a Novel Topotecan Target Protein That Correlates with DAMP Secretion and Antitumor Immune Activation. J. Immunol. 2022, 209, 171–179. [Google Scholar] [CrossRef]
- Lui, G.Y.L.; Grandori, C.; Kemp, C.J. CDK12: An emerging therapeutic target for cancer. J. Clin. Pathol. 2018, 71, 957–962. [Google Scholar] [CrossRef] [Green Version]
- Shaikho, S.; Dobson, C.C.; Naing, T.; Samanfar, B.; Moteshareie, H.; Hajikarimloo, M.; Golshani, A.; Holcik, M. Elevated levels of ribosomal proteins eL36 and eL42 control expression of Hsp90 in rhabdomyosarcoma. Translation 2016, 4, e1244395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, J.J.; Yewdell, J.W. Immunoribosomes: Where’s there’s fire, there’s fire. Mol. Immunol. 2019, 113, 38–42. [Google Scholar] [CrossRef] [PubMed]
- Marchingo, J.M.; Cantrell, D.A. Protein synthesis, degradation, and energy metabolism in T cell immunity. Cell. Mol. Immunol. 2022, 19, 303–315. [Google Scholar] [CrossRef] [PubMed]
- Burgess, H.M.; Vink, E.I.; Mohr, I. Minding the message: Tactics controlling RNA decay, modification, and translation in virus-infected cells. Gene Dev. 2022, 36, 108–132. [Google Scholar] [CrossRef]
- Wang, X.; Zhu, J.; Zhang, D.; Liu, G.Q. Ribosomal control in RNA virus-infected cells. Front. Microbiol. 2022, 13, 1026887. [Google Scholar] [CrossRef]
- Miller, C.M.; Selvam, S.; Fuchs, G. Fatal attraction: The roles of ribosomal proteins in the viral life cycle. Wiley Interdiscip. Rev.-Rna 2021, 12, e1613. [Google Scholar] [CrossRef]
- Komov, L.; Kadosh, D.M.; Barnea, E.; Admon, A. The Effect of Interferons on Presentation of Defective Ribosomal Products as HLA Peptides. Mol. Cell. Proteom. 2021, 20, 100105. [Google Scholar] [CrossRef]
- Le Page, C.; Genin, P.; Baines, M.G.; Hiscott, J. Interferon activation and innate immunity. Rev. Immunogenet. 2000, 2, 374–386. [Google Scholar]
- Wei, J.J.; Kishton, R.J.; Angel, M.; Conn, C.S.; Dalla-Venezia, N.; Marcel, V.; Vincent, A.; Catez, F.; Ferre, S.; Ayadi, L.; et al. Ribosomal Proteins Regulate MHC Class I Peptide Generation for Immunosurveillance. Mol. Cell 2019, 73, 1162–1173.e5. [Google Scholar] [CrossRef] [Green Version]
- Thoms, M.; Buschauer, R.; Ameismeier, M.; Koepke, L.; Denk, T.; Hirschenberger, M.; Kratzat, H.; Hayn, M.; Mackens-Kiani, T.; Cheng, J.; et al. Structural basis for translational shutdown and immune evasion by the Nsp1 protein of SARS-CoV-2. Science 2020, 369, 1249–1255. [Google Scholar] [CrossRef]
- Cervantes-Salazar, M.; Angel-Ambrocio, A.H.; Soto-Acosta, R.; Bautista-Carbajal, P.; Hurtado-Monzon, A.M.; Alcaraz-Estrada, S.L.; Ludert, J.E.; Del Angel, R.M. Dengue virus NS1 protein interacts with the ribosomal protein RPL18: This interaction is required for viral translation and replication in Huh-7 cells. Virology 2015, 484, 113–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamitani, W.; Huang, C.; Narayanan, K.; Lokugamage, K.G.; Makino, S. A two-pronged strategy to suppress host protein synthesis by SARS coronavirus Nsp1 protein. Nat. Struct. Mol. Biol. 2009, 16, 1134–1140. [Google Scholar] [CrossRef]
- Mendez, A.S.; Ly, M.; Gonzalez-Sanchez, A.M.; Hartenian, E.; Ingolia, N.T.; Cate, J.H.; Glaunsinger, B.A. The N-terminal domain of SARS-CoV-2 nsp1 plays key roles in suppression of cellular gene expression and preservation of viral gene expression. Cell Rep. 2021, 37, 109841. [Google Scholar] [CrossRef]
- Tidu, A.; Janvier, A.; Schaeffer, L.; Sosnowski, P.; Kuhn, L.; Hammann, P.; Westhof, E.; Eriani, G.; Martin, F. The viral protein NSP1 acts as a ribosome gatekeeper for shutting down host translation and fostering SARS-CoV-2 translation. Rna 2020, 27, 253–264. [Google Scholar] [CrossRef] [PubMed]
- Rao, S.; Hoskins, I.; Tonn, T.; Garcia, P.D.; Ozadam, H.; Sarinay Cenik, E.; Cenik, C. Genes with 5’ terminal oligopyrimidine tracts preferentially escape global suppression of translation by the SARS-CoV-2 Nsp1 protein. Rna 2021, 27, 1025–1045. [Google Scholar] [CrossRef] [PubMed]
- DiGiuseppe, S.; Rollins, M.G.; Astar, H.; Khalatyan, N.; Savas, J.N.; Walsh, D. Proteomic and mechanistic dissection of the poxvirus-customized ribosome. J. Cell Sci. 2020, 134, jcs246603. [Google Scholar] [CrossRef]
- Rollins, M.G.; Shasmal, M.; Meade, N.; Astar, H.; Shen, P.S.; Walsh, D. Negative charge in the RACK1 loop broadens the translational capacity of the human ribosome. Cell Rep. 2021, 36, 109663. [Google Scholar] [CrossRef]
- DiGiuseppe, S.; Rollins, M.G.; Bartom, E.T.; Walsh, D. ZNF598 Plays Distinct Roles in Interferon-Stimulated Gene Expression and Poxvirus Protein Synthesis. Cell Rep. 2018, 23, 1249–1258. [Google Scholar] [CrossRef]
- Kulsuptrakul, J.; Wang, R.; Meyers, N.L.; Ott, M.; Puschnik, A.S. A genome-wide CRISPR screen identifies UFMylation and TRAMP-like complexes as host factors required for hepatitis A virus infection. Cell Rep. 2021, 34, 108859. [Google Scholar] [CrossRef]
- Mazumder, B.; Sampath, P.; Seshadri, V.; Maitra, R.K.; DiCorleto, P.E.; Fox, P.L. Regulated release of L13a from the 60S ribosomal subunit as a mechanism of transcript-specific translational control. Cell 2003, 115, 187–198. [Google Scholar] [CrossRef] [Green Version]
- Sampath, P.; Mazumder, B.; Seshadri, V.; Fox, P.L. Transcript-selective translational silencing by gamma interferon is directed by a novel structural element in the ceruloplasmin mRNA 3’ untranslated region. Mol. Cell. Biol. 2003, 23, 1509–1519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poddar, D.; Basu, A.; Baldwin, W.M., 3rd; Kondratov, R.V.; Barik, S.; Mazumder, B. An extraribosomal function of ribosomal protein L13a in macrophages resolves inflammation. J. Immunol. 2013, 190, 3600–3612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basu, A.; Penumutchu, S.; Nguyen, K.; Mbonye, U.; Tolbert, B.S.; Karn, J.; Komar, A.A.; Mazumder, B. A Structurally Conserved RNA Element within SARS-CoV-2 ORF1a RNA and S mRNA Regulates Translation in Response to Viral S Protein-Induced Signaling in Human Lung Cells. J. Virol. 2022, 96, e0167821. [Google Scholar] [CrossRef] [PubMed]
- Johnson, A.G.; Flynn, R.A.; Lapointe, C.P.; Ooi, Y.S.; Zhao, M.L.; Richards, C.M.; Qiao, W.J.; Yamada, S.B.; Couthouis, J.; Gitler, A.D.; et al. A memory of eS25 loss drives resistance phenotypes. Nucleic Acids Res. 2020, 48, 7279–7297. [Google Scholar] [CrossRef]
- Pink, R.C.; Wicks, K.; Caley, D.P.; Punch, E.K.; Jacobs, L.; Carter, D.R. Pseudogenes: Pseudo-functional or key regulators in health and disease? Rna 2011, 17, 792–798. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Harrison, P.; Gerstein, M. Identification and analysis of over 2000 ribosomal protein pseudogenes in the human genome. Genome Res. 2002, 12, 1466–1482. [Google Scholar] [CrossRef] [Green Version]
- Tonner, P.; Srinivasasainagendra, V.; Zhang, S.; Zhi, D. Detecting transcription of ribosomal protein pseudogenes in diverse human tissues from RNA-seq data. BMC Genom. 2012, 13, 412. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Chr. | Protein Name | Description | Paralog(s) | References |
|---|---|---|---|---|---|
| RPS4X | Xq13.1 | eS4/RPS4 | 40S ribosomal protein S4X | RPS4Y1 RPS4Y2 | [4] |
| RPS27 | 1q21.3 | eS27/RPS27 | 40S ribosomal protein S27 | RPS27L | [4] |
| RPL3 | 22q13.1 | uL3/RPL3 | 60S ribosomal protein L3 | RPL3L | [4,6,30] |
| RPL7 | 8q21.11 | uL30/RPL7 | 60S ribosomal protein L7 | RPL7L1 | [4] |
| RPL10 | Xq28 | uL16/RPL10 | 60S ribosomal protein L10 | RPL10L | [6,28,31] |
| RPL22 | 1p36.31 | eL22/RPL22 | 60S ribosomal protein 22 | RPL22L1 | [4,28,32] |
| RPL26 | 17p13.1 | uL24/RPL26 | 60S ribosomal protein 26 | RPL26L1 | [4] |
| RPL36A | Xq22.1 | eL42/RPL36A | 60S ribosomal protein L36a | RPL36AL | [4,31] |
| RPL39 | Xq24 | eL39/RPL39 | 60S ribosomal protein L39 | RPL39L | [6,28,31,33] |
| Gene Name | Chr. | Protein Name | Description | Paralog(s) | References |
|---|---|---|---|---|---|
| RPS4X | Xq13.1 | eS4/RPS4 | 40S Ribosomal Protein S4, X Isoform | RPS4Y1, RPS4Y2, Rps4l | [77,78,79] |
| RPL10 | Xq28 | uL16/RPL10 | 60S Ribosomal Protein L10 | RPL10L | [31,80] |
| RPL36A | Xq22.1 | eL42/RPL36A | 60S Ribosomal Protein L36a | RPL36AL | [31,81] |
| RPL39 | Xq24 | eL39/RPL39 | 60S Ribosomal Protein L39 | RPL39L | [28,33,82] |
| UTP14A | Xq26.1 | N.A. 1 | UTP14A Small Subunit (SSU) Processome Component | UTP14C | [83,84] |
| RPS6KA3 | Xp22.12 | N.A. | Ribosomal Protein S6 Kinase A3 | None reported | [85] |
| EIF1AX | Xp22.12 | N.A. | Eukaryotic Translation Initiation Factor 1A X-Linked | EIF1AY | [86,87] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miller, S.C.; MacDonald, C.C.; Kellogg, M.K.; Karamysheva, Z.N.; Karamyshev, A.L. Specialized Ribosomes in Health and Disease. Int. J. Mol. Sci. 2023, 24, 6334. https://doi.org/10.3390/ijms24076334
Miller SC, MacDonald CC, Kellogg MK, Karamysheva ZN, Karamyshev AL. Specialized Ribosomes in Health and Disease. International Journal of Molecular Sciences. 2023; 24(7):6334. https://doi.org/10.3390/ijms24076334
Chicago/Turabian StyleMiller, Sarah C., Clinton C. MacDonald, Morgana K. Kellogg, Zemfira N. Karamysheva, and Andrey L. Karamyshev. 2023. "Specialized Ribosomes in Health and Disease" International Journal of Molecular Sciences 24, no. 7: 6334. https://doi.org/10.3390/ijms24076334