
Different miRNAs Related to FBXW7 Mutations or High Mitotic Indices Contribute to Rectal Neuroendocrine Tumors: A Pilot Study
 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Baseline Characteristics and Genetic Alterations of Rectal NETs
2.2. Screening of Candidate miRNAs Associated with Specific Genetic Mutations
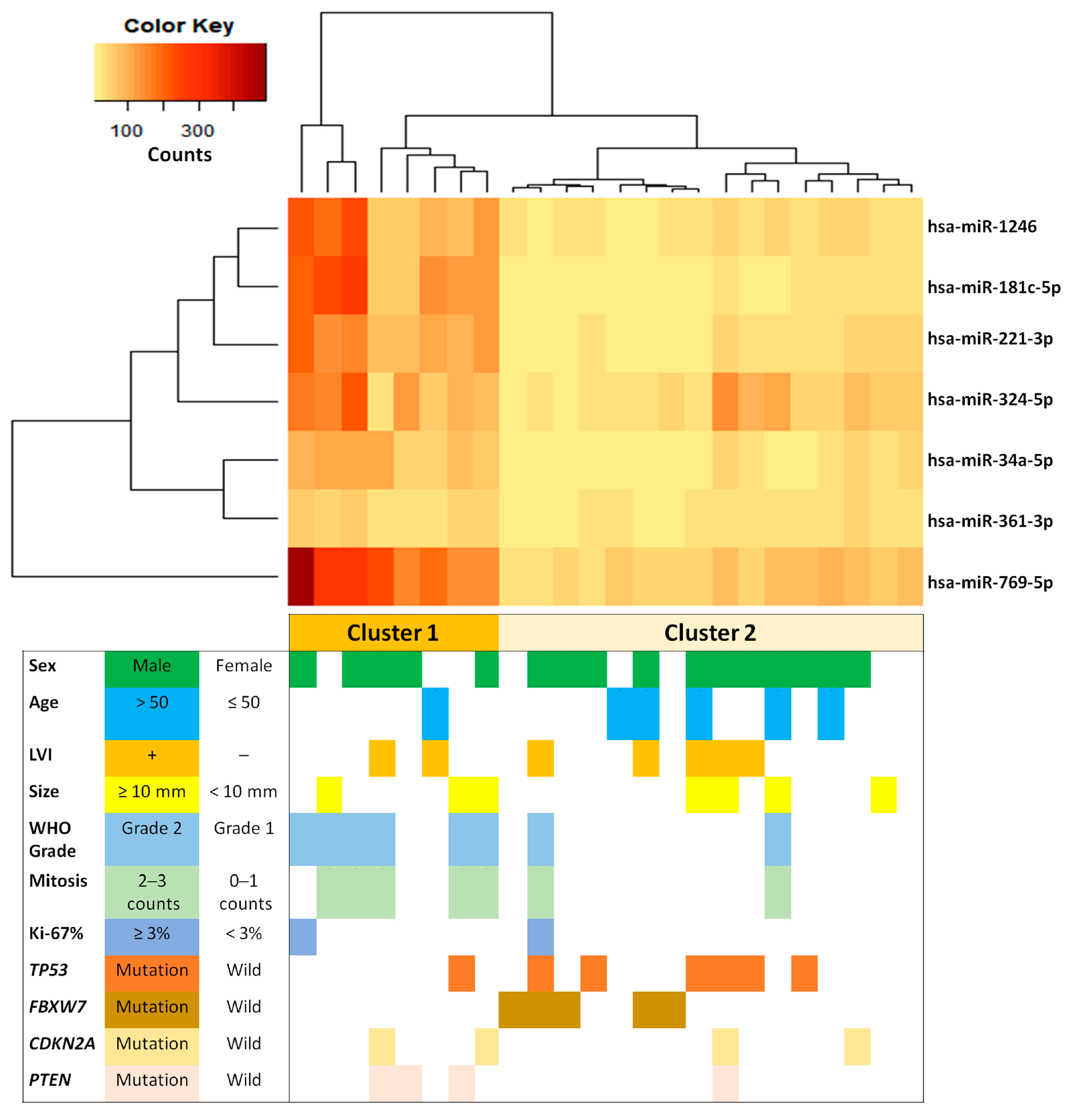
2.3. miRNAs Related to Clinical and Pathological Characteristics
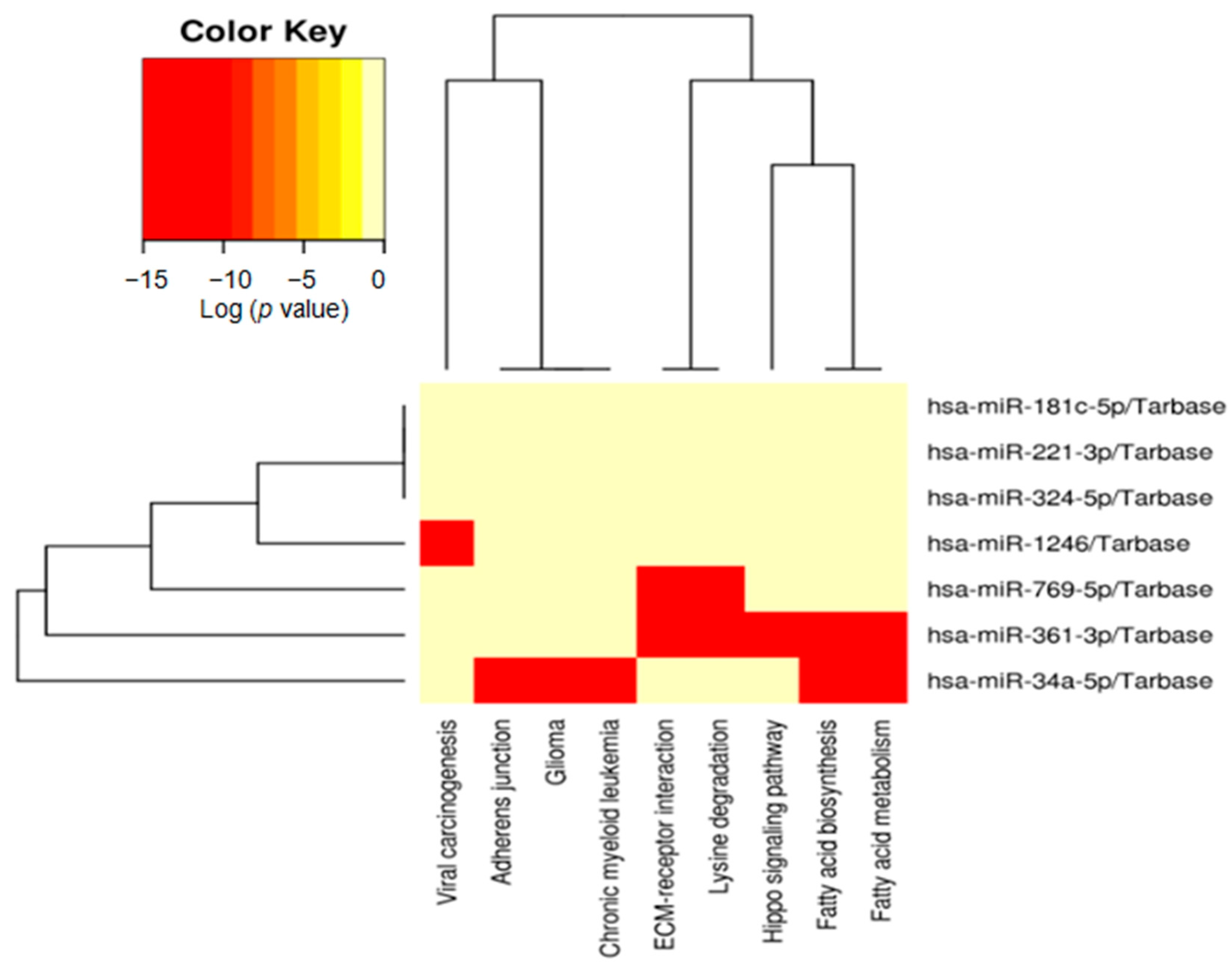
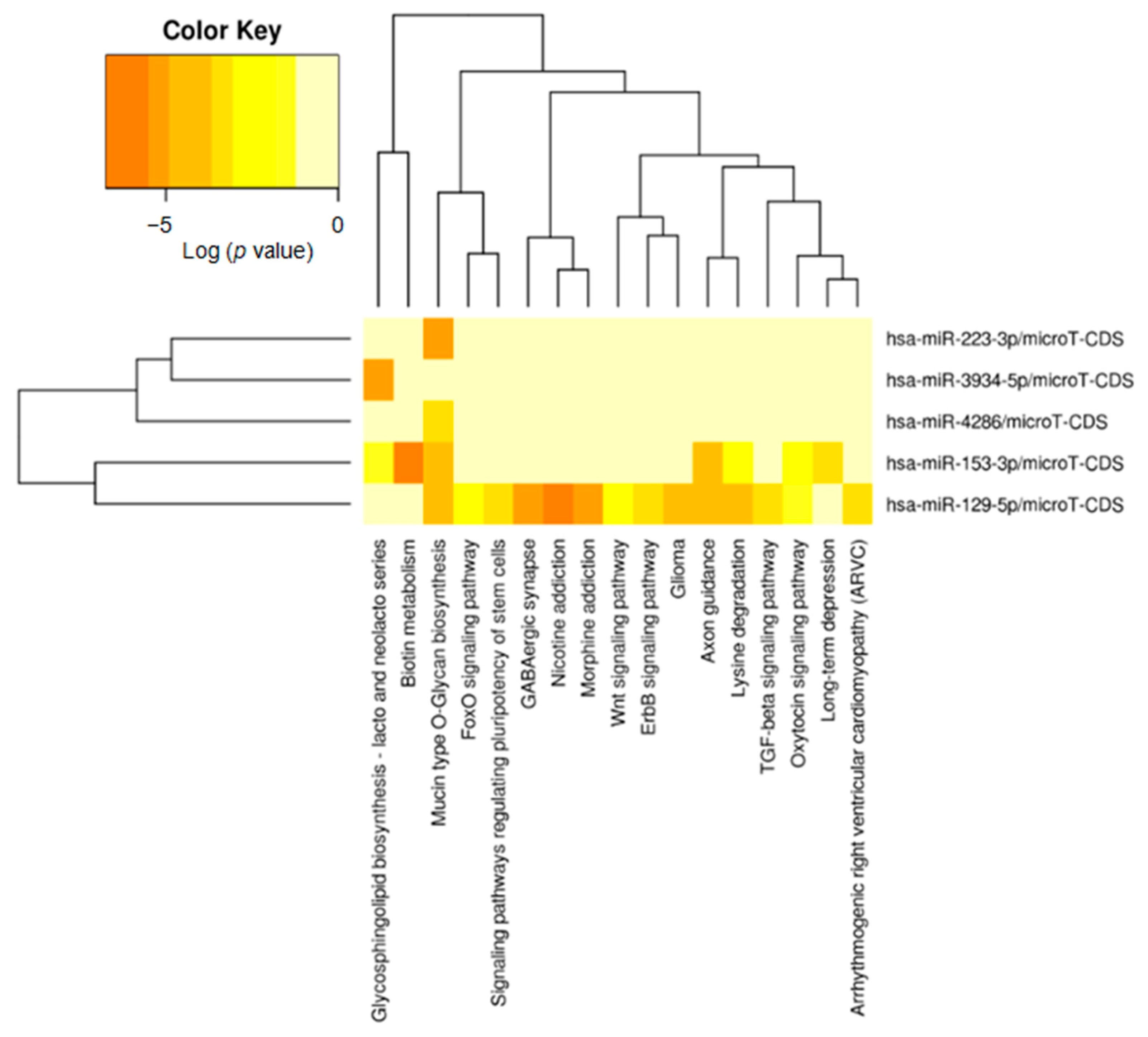
2.4. Identification of Potential Functional Pathways Related to miRNAs
3. Discussion
4. Materials and Methods
4.1. Patients
4.2. Histologic Evaluation
4.3. DNA Extraction and NGS
4.4. miRNA Extraction and Nanostring nCounter miRNA Expression Assay
4.5. DNA Intelligent Analysis (DIANA)-miRPath for miRNA Pathway Analysis
4.6. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Soga, J. Carcinoids of the rectum: An evaluation of 1271 reported cases. Surg. Today. 1997, 27, 112–119. [Google Scholar] [CrossRef]
- Modlin, I.M.; Oberg, K.; Chung, D.C.; Jensen, R.T.; de Herder, W.W.; Thakker, R.V.; Caplin, M.; Delle Fave, G.; Kaltsas, G.A.; Krenning, E.P.; et al. Gastroenteropancreatic neuroendocrine tumours. Lancet Oncol. 2008, 9, 61–72. [Google Scholar] [CrossRef] [PubMed]
- Jung, E.S.; Kang, Y.K.; Cho, M.Y.; Kim, J.M.; Lee, W.A.; Lee, H.E.; Park, S.; Sohn, J.H.; Jin, S.Y. Update on the proposal for creating a guideline for cancer registration of the gastrointestinal tumors (I-2). Korean J. Pathol. 2012, 46, 443–453. [Google Scholar] [CrossRef] [PubMed]
- Gao, W.; Liu, S.M.; Lu, H.Z.; Liang, J.; Yuan, Y.L.; Liu, X.Y. Analysis of clinicopathological features of intestinal neuroendocrine neoplasms. Zhonghua Zhong Liu Za Zhi 2012, 34, 450–456. [Google Scholar] [CrossRef] [PubMed]
- Gastrointestinal Pathology Study Group of Korean Society of Pathologists; Cho, M.Y.; Kim, J.M.; Sohn, J.H.; Kim, M.J.; Kim, K.M.; Kim, W.H.; Kim, H.; Kook, M.C.; Park, D.Y.; et al. Current Trends of the Incidence and Pathological Diagnosis of Gastroenteropancreatic Neuroendocrine Tumors (GEP-NETs) in Korea 2000–2009: Multicenter Study. Cancer Res. Treat. 2012, 44, 157–165. [Google Scholar] [CrossRef]
- Kwon, M.J.; Kang, H.S.; Soh, J.S.; Lim, H.; Kim, J.H.; Park, C.K.; Park, H.R.; Nam, E.S. Lymphovascular invasion in more than one-quarter of small rectal neuroendocrine tumors. World J. Gastroenterol. 2016, 22, 9400–9410. [Google Scholar] [CrossRef]
- Fahy, B.N.; Tang, L.H.; Klimstra, D.; Wong, W.D.; Guillem, J.G.; Paty, P.B.; Temple, L.K.; Shia, J.; Weiser, M.R. Carcinoid of the rectum risk stratification (CaRRs): A strategy for preoperative outcome assessment. Ann. Surg. Oncol. 2007, 14, 396–404. [Google Scholar] [CrossRef]
- Shields, C.J.; Tiret, E.; Winter, D.C.; International Rectal Carcinoid Study Group. Carcinoid tumors of the rectum: A multi-institutional international collaboration. Ann. Surg. 2010, 252, 750–755. [Google Scholar] [CrossRef]
- Yoon, S.N.; Yu, C.S.; Shin, U.S.; Kim, C.W.; Lim, S.B.; Kim, J.C. Clinicopathological characteristics of rectal carcinoids. Int. J. Color. Dis. 2010, 25, 1087–1092. [Google Scholar] [CrossRef]
- Kasuga, A.; Chino, A.; Uragami, N.; Kishihara, T.; Igarashi, M.; Fujita, R.; Yamamoto, N.; Ueno, M.; Oya, M.; Muto, T. Treatment strategy for rectal carcinoids: A clinicopathological analysis of 229 cases at a single cancer institution. J. Gastroenterol. Hepatol. 2012, 27, 1801–1807. [Google Scholar] [CrossRef]
- Yamagishi, D.; Matsubara, N.; Noda, M.; Yamano, T.; Tsukamoto, K.; Kuno, T.; Hamanaka, M.; Kobayashi, M.; Ikeuchi, H.; Matsuda, I.; et al. Clinicopathological characteristics of rectal carcinoid patients undergoing surgical resection. Oncol. Lett. 2012, 4, 910–914. [Google Scholar] [CrossRef] [PubMed]
- Caplin, M.; Sundin, A.; Nillson, O.; Baum, R.P.; Klose, K.J.; Kelestimur, F.; Plöckinger, U.; Papotti, M.; Salazar, R.; Pascher, A.; et al. Enets Consensus Guidelines for the management of patients with digestive neuroendocrine neoplasms: Colorectal neuroendocrine neoplasms. Neuroendocrinology 2012, 95, 88–97. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Xie, H.; Xie, L.; Li, J.; Fu, W. Factors associated with lymph node metastasis in radically resected rectal carcinoids: A systematic review and meta-analysis. J. Gastrointest. Surg. 2013, 17, 1689–1697. [Google Scholar] [CrossRef] [PubMed]
- Han, D.S.; Sohn, J.H.; Byeon, J.S.; Choi, H.; Kim, J.M. Diagnostic coding for intramucosal carcinoma and neuroendocrine tumor in the colorectum: Proposal for avoiding confusing coding in Korea. Clin. Endosc. 2015, 48, 216–220. [Google Scholar] [CrossRef]
- Venizelos, A.; Elvebakken, H.; Perren, A.; Nikolaienko, O.; Deng, W.; Lothe, I.M.B.; Couvelard, A.; Hjortland, G.O.; Sundlöv, A.; Svensson, J.; et al. The molecular characteristics of high-grade gastroenteropancreatic neuroendocrine neoplasms. Endocr. Relat. Cancer 2021, 29, 1–14. [Google Scholar] [CrossRef]
- Tang, L.H.; Basturk, O.; Sue, J.J.; Klimstra, D.S. A practical approach to the classification of WHO Grade 3 (G3) well-differentiated neuroendocrine tumor (WD-NET) and poorly differentiated neuroendocrine carcinoma (PD-NEC) of the pancreas. Am. J. Surg. Pathol. 2016, 40, 1192–1202. [Google Scholar] [CrossRef] [Green Version]
- Capuano, F.; Grami, O.; Pugliese, L.; Paulli, M.; Pietrabissa, A.; Solcia, E.; Vanoli, A. Grade 3 neuroendocrine tumor (G3 NET) in a background of multiple serotonin cell neoplasms of the ileum associated with carcinoid syndrome and aggressive behavior. Endocr. Pathol. 2018, 29, 369–373. [Google Scholar] [CrossRef]
- Park, H.Y.; Kwon, M.J.; Kang, H.S.; Kim, Y.J.; Kim, N.Y.; Kim, M.J.; Min, K.W.; Choi, K.C.; Nam, E.S.; Cho, S.J.; et al. Targeted next-generation sequencing of well-differentiated rectal, gastric, and appendiceal neuroendocrine tumors to identify potential targets. Hum. Pathol. 2019, 87, 83–94. [Google Scholar] [CrossRef]
- Kagohara, L.T.; Stein-O’Brien, G.L.; Kelley, D.; Flam, E.; Wick, H.C.; Danilova, L.V.; Easwaran, H.; Favorov, A.V.; Qian, J.; Gaykalova, D.A.; et al. Epigenetic regulation of gene expression in cancer: Techniques, resources and analysis. Brief. Funct. Genom. 2018, 17, 49–63. [Google Scholar] [CrossRef] [Green Version]
- Ying, S.Y.; Chang, D.C.; Lin, S.L. The microRNA (miRNA): Overview of the RNA genes that modulate gene function. Mol. Biotechnol. 2008, 38, 257–268. [Google Scholar] [CrossRef]
- Mitsuhashi, K.; Yamamoto, I.; Kurihara, H.; Kanno, S.; Ito, M.; Igarashi, H.; Ishigami, K.; Sukawa, Y.; Tachibana, M.; Takahashi, H.; et al. Analysis of the molecular features of rectal carcinoid tumors to identify new biomarkers that predict biological malignancy. Oncotarget 2015, 6, 22114–22125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murayama, N.; Okamoto, K.; Nakagawa, T.; Miyoshi, J.; Nishida, K.; Kawaguchi, T.; Kagemoto, K.; Kitamura, S.; Ma, B.; Miyamoto, H.; et al. miR-144-3p/miR-451a promotes lymphovascular invasion through repression of PTEN/p19 in rectal neuroendocrine tumors. J. Gastroenterol. Hepatol. 2022, 37, 919–927. [Google Scholar] [CrossRef]
- Vega Neira, S.J.; Torres Carvajal, M.M.; Barrera Herrera, L.; Andrade Perez, R.; Lopez Panqueva, R. P-017 Analysis of microRNA expression in gastroenteropancreatic Neuroendocrine Tumor. Ann Oncol. 2016, 27, II5. [Google Scholar] [CrossRef] [Green Version]
- Malczewska, A.; Kidd, M.; Matar, S.; Kos-Kudla, B.; Modlin, I.M. A comprehensive assessment of the role of miRNAs as biomarkers in gastroenteropancreatic neuroendocrine tumors. Neuroendocrinology 2018, 107, 73–90. [Google Scholar] [CrossRef] [PubMed]
- Precazzini, F.; Detassis, S.; Imperatori, A.S.; Denti, M.A.; Campomenosi, P. Measurements methods for the development of microRNA-based tests for cancer diagnosis. Int. J. Mol. Sci. 2021, 22, 1176. [Google Scholar] [CrossRef]
- Benjamini, Y.; Krieger, A.M.; Yekutieli, D. Adaptive linear step-up procedures that control the false discovery rate. Biometrika 2006, 93, 491–507. [Google Scholar] [CrossRef]
- Welcker, M.; Clurman, B.E. FBW7 ubiquitin ligase: A tumour suppressor at the crossroads of cell division, growth and differentiation. Nat. Rev. Cancer 2008, 8, 83–93. [Google Scholar] [CrossRef]
- Davis, R.J.; Welcker, M.; Clurman, B.E. Tumor suppression by the Fbw7 ubiquitin ligase: Mechanisms and opportunities. Cancer Cell. 2014, 26, 455–464. [Google Scholar] [CrossRef] [Green Version]
- Lan, H.; Sun, Y. Tumor suppressor FBXW7 and its regulation of DNA damage response and repair. Front. Cell Dev. Biol. 2021, 9, 751574. [Google Scholar] [CrossRef]
- Jiang, Y.; Qi, X.; Liu, X.; Zhang, J.; Ji, J.; Zhu, Z.; Ren, J.; Yu, Y. Fbxw7 haploinsufficiency loses its protection against DNA damage and accelerates MNU-induced gastric carcinogenesis. Oncotarget 2017, 8, 33444–33456. [Google Scholar] [CrossRef] [Green Version]
- Galindo-Moreno, M.; Giráldez, S.; Limón-Mortés, M.C.; Belmonte-Fernández, A.; Reed, S.I.; Sáez, C.; Japón, M.Á.; Tortolero, M.; Romero, F. SCF(FBXW7)-mediated degradation of p53 promotes cell recovery after UV-induced DNA damage. FASEB J. 2019, 33, 11420–11430. [Google Scholar] [CrossRef]
- Zhang, Q.; Mady, A.S.A.; Ma, Y.; Ryan, C.; Lawrence, T.S.; Nikolovska-Coleska, Z.; Sun, Y.; Morgan, M.A. The WD40 domain of FBXW7 is a poly(ADP-ribose)-binding domain that mediates the early DNA damage response. Nucleic Acids Res. 2019, 47, 4039–4053. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Guo, Y.; Liang, X.; Sun, M.; Wang, G.; De, W.; Wu, W. MicroRNA-223 functions as an oncogene in human gastric cancer by targeting FBXW7/hCdc. J. Cancer Res. Clin. Oncol. 2012, 138, 763–774. [Google Scholar] [CrossRef]
- Kumar, V.; Palermo, R.; Talora, C.; Campese, A.F.; Checquolo, S.; Bellavia, D.; Tottone, L.; Testa, G.; Miele, E.; Indraccolo, S.; et al. Notch and NF-kB signaling pathways regulate miR-223/FBXW7 axis in T-cell acute lymphoblastic leukemia. Leukemia 2014, 28, 2324–2335. [Google Scholar] [CrossRef] [PubMed]
- Xia, W.; Zhou, J.; Luo, H.; Liu, Y.; Peng, C.; Zheng, W.; Ma, W. MicroRNA-32 promotes cell proliferation, migration and suppresses apoptosis in breast cancer cells by targeting FBXW. Cancer Cell Int. 2017, 17, 14. [Google Scholar] [CrossRef] [Green Version]
- Zhou, C.; Shen, L.; Mao, L.; Wang, B.; Li, Y.; Yu, H. miR-92a is upregulated in cervical cancer and promotes cell proliferation and invasion by targeting FBXW. Biochem. Biophys. Res. Commun. 2015, 458, 63–69. [Google Scholar] [CrossRef]
- Chang, H.; Liu, Y.H.; Wang, L.L.; Wang, J.; Zhao, Z.H.; Qu, J.F.; Wang, S.F. MiR-182 promotes cell proliferation by suppressing FBXW7 and FBXW11 in non-small cell lung cancer. Am. J. Transl. Res. 2018, 10, 1131–1142. [Google Scholar] [PubMed]
- Yue, B. Biology of the extracellular matrix: An overview. J. Glaucoma. 2014, 23 (Suppl. 1), S20–S23. [Google Scholar] [CrossRef] [Green Version]
- Saupe, F.; Schwenzer, A.; Jia, Y.; Gasser, I.; Spenlé, C.; Langlois, B.; Kammerer, M.; Lefebvre, O.; Hlushchuk, R.; Rupp, T.; et al. Tenascin-C downregulates wnt inhibitor dickkopf-1, promoting tumorigenesis in a neuroendocrine tumor model. Cell Rep. 2013, 5, 482–492. [Google Scholar] [CrossRef] [Green Version]
- Cives, M.; Pelle’, E.; Quaresmini, D.; Rizzo, F.M.; Tucci, M.; Silvestris, F. The Tumor Microenvironment in Neuroendocrine Tumors: Biology and Therapeutic Implications. Neuroendocrinology 2019, 109, 83–99. [Google Scholar] [CrossRef]
- Prieto, T.G.; Machado-Rugolo, J.; Baldavira, C.M.; Velosa, A.P.P.; Teodoro, W.R.; Saber, A.M.A.; Capelozzi, V.L. The fibrosis-targeted collagen/integrins gene profile predicts risk of metastasis in pulmonary neuroendocrine neoplasms. Front. Oncol. 2021, 11, 706141. [Google Scholar] [CrossRef] [PubMed]
- Dai, Z.; Yang, S.; Xu, L.; Hu, H.; Liao, K.; Wang, J.; Wang, Q.; Gao, S.; Li, B.; Lai, L. Identification of Cancer–associated metabolic vulnerabilities by modeling multi-objective optimality in metabolism. Cell Commun. Signal. 2019, 17, 124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, W.; Liu, S.; Zhang, J.; Pei, M.; Xiao, Y.; Li, J.; Hong, L.; Lin, J.; Wang, J.; Wu, X.; et al. Vorinostat triggers miR-769-5p/3p-mediated suppression of proliferation and induces apoptosis via the STAT3-IGF1R-HDAC3 complex in human gastric cancer. Cancer Lett. 2021, 521, 196–209. [Google Scholar] [CrossRef]
- Sun, Y.; Li, J.; Zheng, S. MiR-769-5p, Which Targets HDGF, Inhibits Cell Proliferation and Invasion in Nonsmall Cell Lung Cancer. Cancer Biotherapy Radiopharm. 2021. [Google Scholar] [CrossRef]
- Gao, T.; Zhao, L.; Zhang, F.; Cao, C.; Fan, S.; Shi, X. Evaluate the diagnostic and prognostic value of NUSAP1 in papillary thyroid carcinoma and identify the relationship with genes, proteins, and immune factors. World J. Surg. Oncol. 2022, 20, 207. [Google Scholar] [CrossRef]
- Corti, A. Chromogranin A and the tumor microenvironment. Cell Mol. Neurobiol. 2010, 30, 1163–1170. [Google Scholar] [CrossRef]
- Foye, C.; Yan, I.K.; David, W.; Shukla, N.; Habboush, Y.; Chase, L.; Ryland, K.; Kesari, V.; Patel, T. Comparison of miRNA quantitation by Nanostring in serum and plasma samples. PLoS ONE 2017, 12, e0189165. [Google Scholar] [CrossRef] [Green Version]
- Géczi, D.; Nagy, B.; Szilágyi, M.; Penyige, A.; Klekner, Á.; Jenei, A.; Virga, J.; Birkó, Z. Analysis of circulating miRNA profile in plasma samples of glioblastoma patients. Int. J. Mol. Sci. 2021, 22, 5058. [Google Scholar] [CrossRef] [PubMed]
- Cui, C.; Cui, Q. The relationship of human tissue microRNAs with those from body fluids. Sci Rep. 2020, 10, 5644. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.J.; Kwon, M.J.; Kang, H.S.; Choi, K.C.; Nam, E.S.; Cho, S.J.; Park, H.R.; Min, S.K.; Seo, J.; Choe, J.Y.; et al. Identification of phosphohistone H3 cutoff values corresponding to original WHO grades but distinguishable in well-differentiated gastrointestinal neuroendocrine tumors. BioMed. Res. Int. 2018, 2018, 1013640. [Google Scholar] [CrossRef] [Green Version]
- Denaro, M.; Ugolini, C.; Poma, A.M.; Borrelli, N.; Materazzi, G.; Piaggi, P.; Chiarugi, M.; Miccoli, P.; Vitti, P.; Basolo, F. Differences in miRNA expression profiles between wild-type and mutated NIFTPs. Endocr. Relat. Cancer 2017, 24, 543–553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vlachos, I.S.; Zagganas, K.; Paraskevopoulou, M.D.; Georgakilas, G.; Karagkouni, D.; Vergoulis, T.; Dalamagas, T.; Hatzigeorgiou, A.G. Diana-miRPath v3.0: Deciphering microRNA function with experimental support. Nucleic Acids Res. 2015, 43, W460–W466. [Google Scholar] [CrossRef] [PubMed]
- Clarke, R.; Ressom, H.W.; Wang, A.; Xuan, J.; Liu, M.C.; Gehan, E.A.; Wang, Y. The properties of high-dimensional data spaces: Implications for exploring gene and protein expression data. Nat. Rev. Cancer. 2008, 8, 37–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Characteristic | N = 24 |
|---|---|
| Sex | |
| Male | 17 (70.8%) |
| Female | 7 (29.2%) |
| Age, mean ± SD (years) | 48.5 ± 9.51 (range, 34–73) |
| ≤50 | 16 (66.7%) |
| >50 | 8 (33.3%) |
| Tumor size, mean ± SD (mm) | 7.62 ± 2.81 (range, 5–15) |
| <10 mm | 17 (70.8%) |
| ≥10 mm | 7 (29.2%) |
| WHO grade | |
| Grade 1 | 16 (66.7%) |
| Grade 2 | 8 (33.3%) |
| Mitotic count/10 HPFs | |
| 0–1 | 18 (75.0%) |
| 2–3 | 6 (25.0%) |
| Ki-67 labeling index | |
| <3% | 22 (91.6%) |
| ≥3% | 2 (8.3%) |
| Lymphovascular invasion | |
| Negative | 17 (70.8%) |
| Positive | 7 (29.2%) |
| Mutated detected tumors | |
| No detected | 5 (20.8%) |
| Detected | 19 (79.2%) |
| TP53 | |
| Wild-type | 17 (70.8%) |
| Mutated | 7 (29.2%) |
| FBXW7 | |
| Wild-type | 19 (79.2%) |
| Mutated | 5 (20.8%) |
| CDKN2A | |
| Wild-type | 20 (83.3%) |
| Mutated | 4 (16.7%) |
| PTEN | |
| Wild-type | 20 (83.3%) |
| Mutated | 4 (16.7%) |
| Dysregulated miRNAs | FBXW7 Wild-Type vs. Mutated | Expression in FBXW7- Mutated Tumors | |
|---|---|---|---|
| p-Value * | Adjusted p-Value ** | ||
| hsa-miR-769-5p | 0.000047 | 0.000047 | down-regulated |
| hsa-miR-221-3p | 0.000565 | 0.000565 | down-regulated |
| hsa-miR-34a-5p | 0.000565 | 0.000565 | down-regulated |
| hsa-miR-181c-5p | 0.000847 | 0.000847 | down-regulated |
| hsa-miR-1246 | 0.000894 | 0.000894 | down-regulated |
| hsa-miR-324-5p | 0.000894 | 0.000894 | down-regulated |
| hsa-miR-361-3p | 0.000894 | 0.000894 | down-regulated |
| Dysregulated miRNAs | Mitotic Index (Low vs. High) | Expression in Tumors with High Mitotic Index | |
|---|---|---|---|
| p-Value * | Adjusted p-Value ** | ||
| hsa-miR-3934-5p | 0.000040 | 0.000047 | up–regulated |
| hsa-miR-153-3p | 0.003259 | 0.366977 | down-regulated |
| hsa-miR-4286 | 0.004207 | 0.001365 | down-regulated |
| hsa-miR-223-3p | 0.006796 | 0.023951 | down-regulated |
| hsa-miR-129-5p | 0.008518 | 0.235225 | down-regulated |
| Signaling Pathway | p-Value | Target Genes | miRNAs |
|---|---|---|---|
| Fatty acid biosynthesis | 1 × 10−325 | FASN, ACSL4, ACSL1, ACACA | hsa-miR-34a-5p hsa-miR-361-3p |
| ECM-receptor interaction | 1 × 10−325 | LAMB2, LAMB1, THBS1, COL4A5, COL4A2, COL6A2, COL5A1, COL1A1, DAG1, COL1A2, COL4A6, TNC, SDC4 | hsa-miR-769-5p hsa-miR-361-3p |
| Fatty acid metabolism | 1.67 × 10−11 | FASN, SCD5, ACOX1, ACOX3, ACAA2, PPT1, PTPLA, CPT2, ACADVL, PPT2, SCD, ACSL4, ACSL1, HSD17B12, MECR, ACACA | hsa-miR-34a-5p hsa-miR-361-3p |
| Lysine degradation | 2.49 × 10−8 | SETD1B, NSD1, ASH1L, KMT2D, DOT1L, WHSC1, KMT2A, KMT2E, SETD1A | hsa-miR-769-5p hsa-miR-361-3p |
| Viral carcinogenesis | 8.07 × 10−8 | PIK3CB, BAX, CDK6, TP53, KAT2B, CCNE1 | hsa-miR-1246 |
| Hippo signaling pathway | 3.76 × 10−5 | PPP1CA, YAP1, YWHAG, DLG4, CCND1, PPP2R1A, TEAD2, PARD6B | hsa-miR-361-3p |
| Glioma | 4.73 × 10−5 | BRAF, PDGFRA, E2F1, CDK4, E2F2, MAP2K2, TGFA, PIK3R2, RAF1, EGFR, CDKN2A, CDK6, ARAF, TP53, AKT2, PLCG1, CCND1, E2F3, PRKCB, IGF1, PIK3CA, CDKN1A, MAP2K1, PTEN, MAPK1, PDGFRB | hsa-miR-34a-5p |
| Chronic myeloid leukemia | 3.82 × 10−4 | BRAF, E2F1, NFKB1, CDK4, E2F2, CRKL, MAP2K2, CRK, RUNX1, HDAC1, PIK3R2, RAF1, CDKN2A, CDK6, TGFB1, ARAF, TP53, AKT2, CCND1, SMAD4, E2F3, MYC, NFKBIA, PTPN11, PIK3CA, CDKN1A, MAP2K1, MAPK1, TGFBR2, BAD, TGFB3 | hsa-miR-34a-5p |
| Adherens junction | 4.54 × 10−4 | ACTB, CSNK2A2, MET, RAC2, CTNND1, PVRL2, ACTG1, TCF7L1, IQGAP1, PTPRM, EGFR, VCL, SNAI1, TJP1, SORBS1, FYN, MLLT4, CDH1, SMAD4, CTNNB1, WASF2, CSNK2A1, FARP2, SRC, EP300, LEF1, PARD3, FGFR1, MAPK1, TGFBR2, PVRL1 | hsa-miR-34a-5p |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kang, H.S.; Park, H.Y.; Lim, H.; Son, I.T.; Kim, M.-J.; Kim, N.Y.; Kim, M.J.; Nam, E.S.; Cho, S.J.; Kwon, M.J. Different miRNAs Related to FBXW7 Mutations or High Mitotic Indices Contribute to Rectal Neuroendocrine Tumors: A Pilot Study. Int. J. Mol. Sci. 2023, 24, 6329. https://doi.org/10.3390/ijms24076329
Kang HS, Park HY, Lim H, Son IT, Kim M-J, Kim NY, Kim MJ, Nam ES, Cho SJ, Kwon MJ. Different miRNAs Related to FBXW7 Mutations or High Mitotic Indices Contribute to Rectal Neuroendocrine Tumors: A Pilot Study. International Journal of Molecular Sciences. 2023; 24(7):6329. https://doi.org/10.3390/ijms24076329
Chicago/Turabian StyleKang, Ho Suk, Ha Young Park, Hyun Lim, Il Tae Son, Min-Jeong Kim, Nan Young Kim, Min Jeong Kim, Eun Sook Nam, Seong Jin Cho, and Mi Jung Kwon. 2023. "Different miRNAs Related to FBXW7 Mutations or High Mitotic Indices Contribute to Rectal Neuroendocrine Tumors: A Pilot Study" International Journal of Molecular Sciences 24, no. 7: 6329. https://doi.org/10.3390/ijms24076329