

Plasma Proteomics Unveil Novel Immune Signatures and Biomarkers upon SARS-CoV-2 Infection
 , ,
, ,
Abstract
:1. Introduction
2. Results
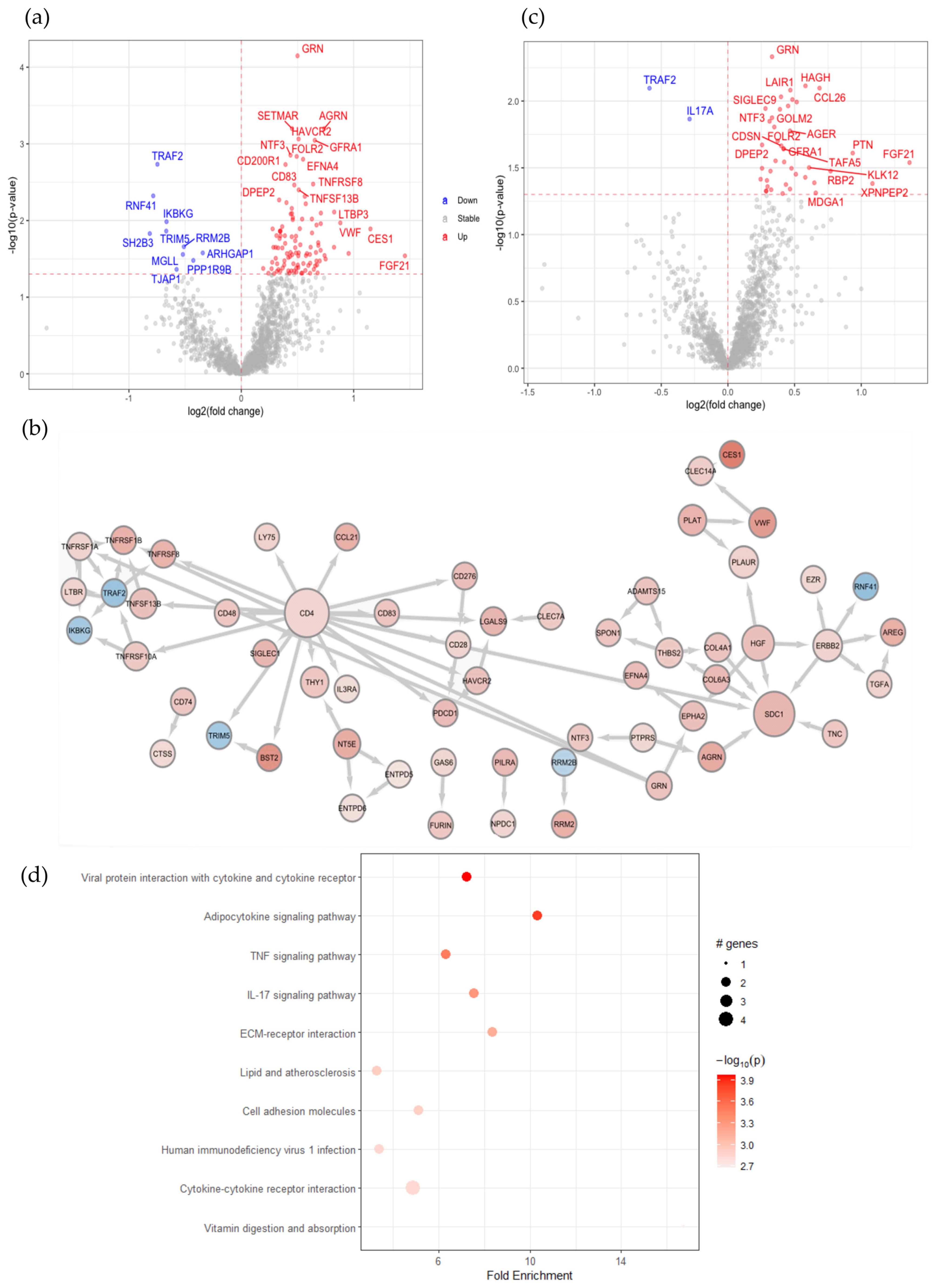
2.1. SARS-CoV-2 Infection in Patients with Comorbidities Causes Plasma Protein Changes with Enhanced Soluble CD4 and Associated Proteins
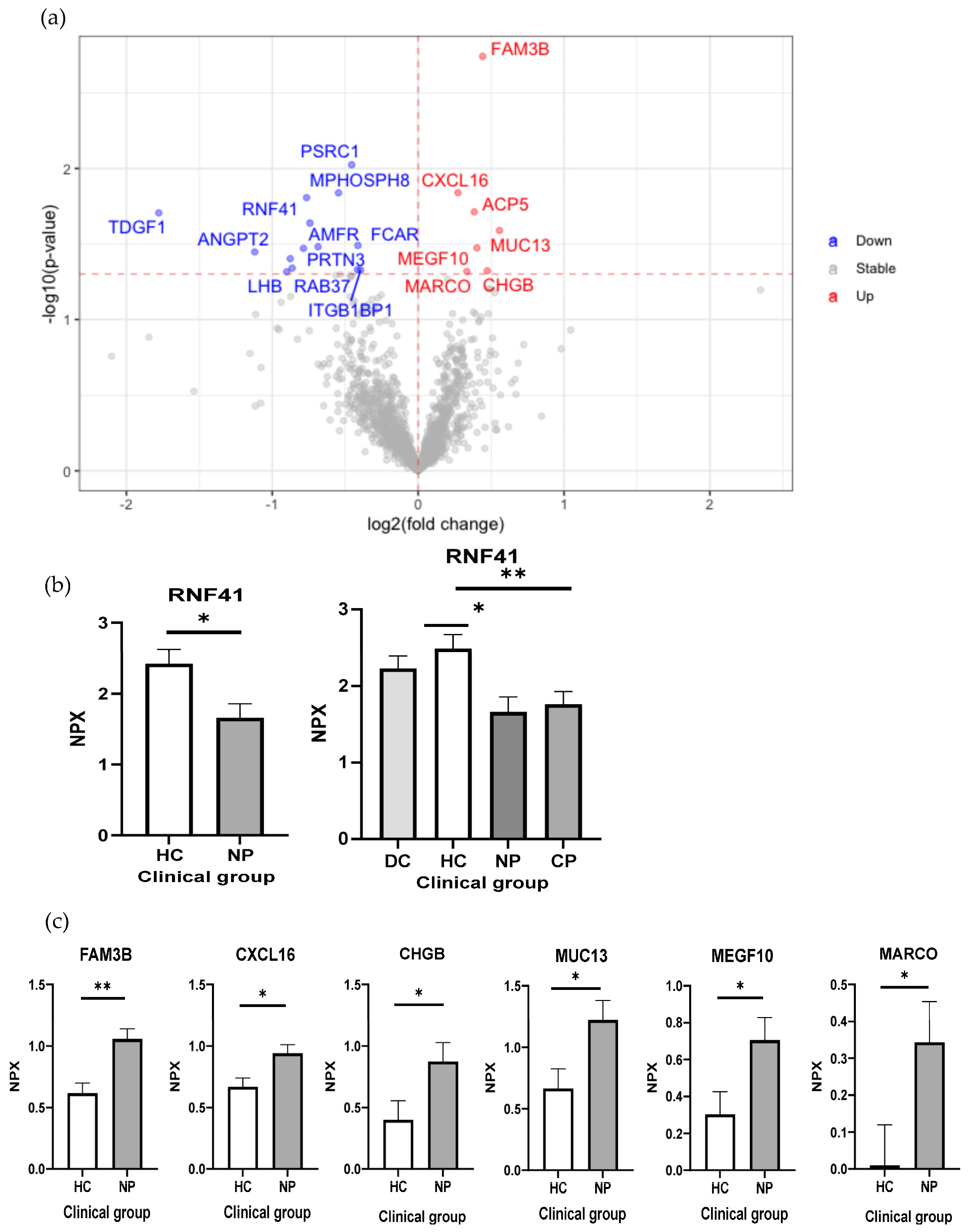
2.2. Reduced RNF41 in Plasma Is Associated with a SARS-CoV-2 Infection
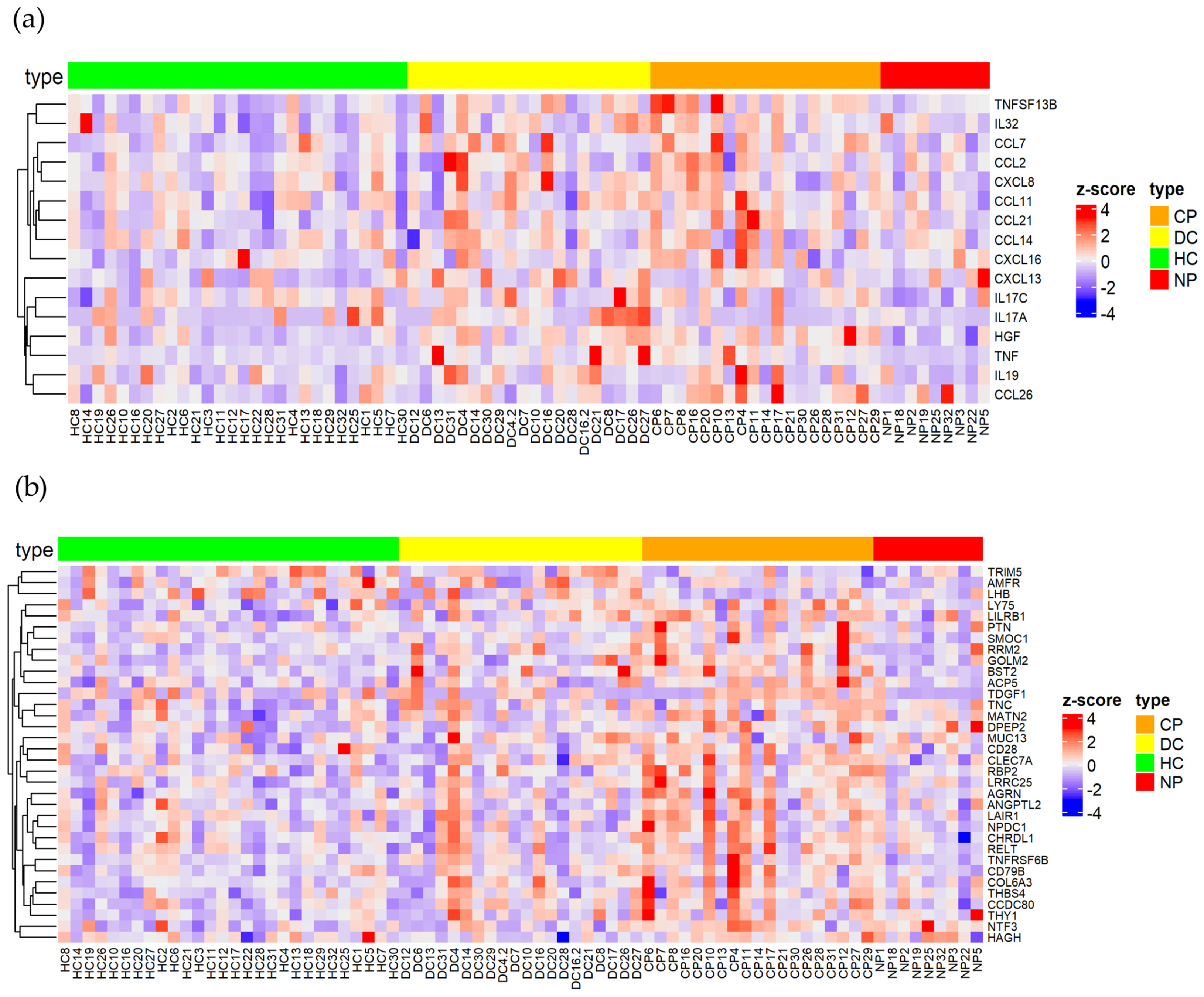
2.3. Characterization of Early and Long-Term Plasma Protein Responses Associated with SARS-CoV-2 Infection
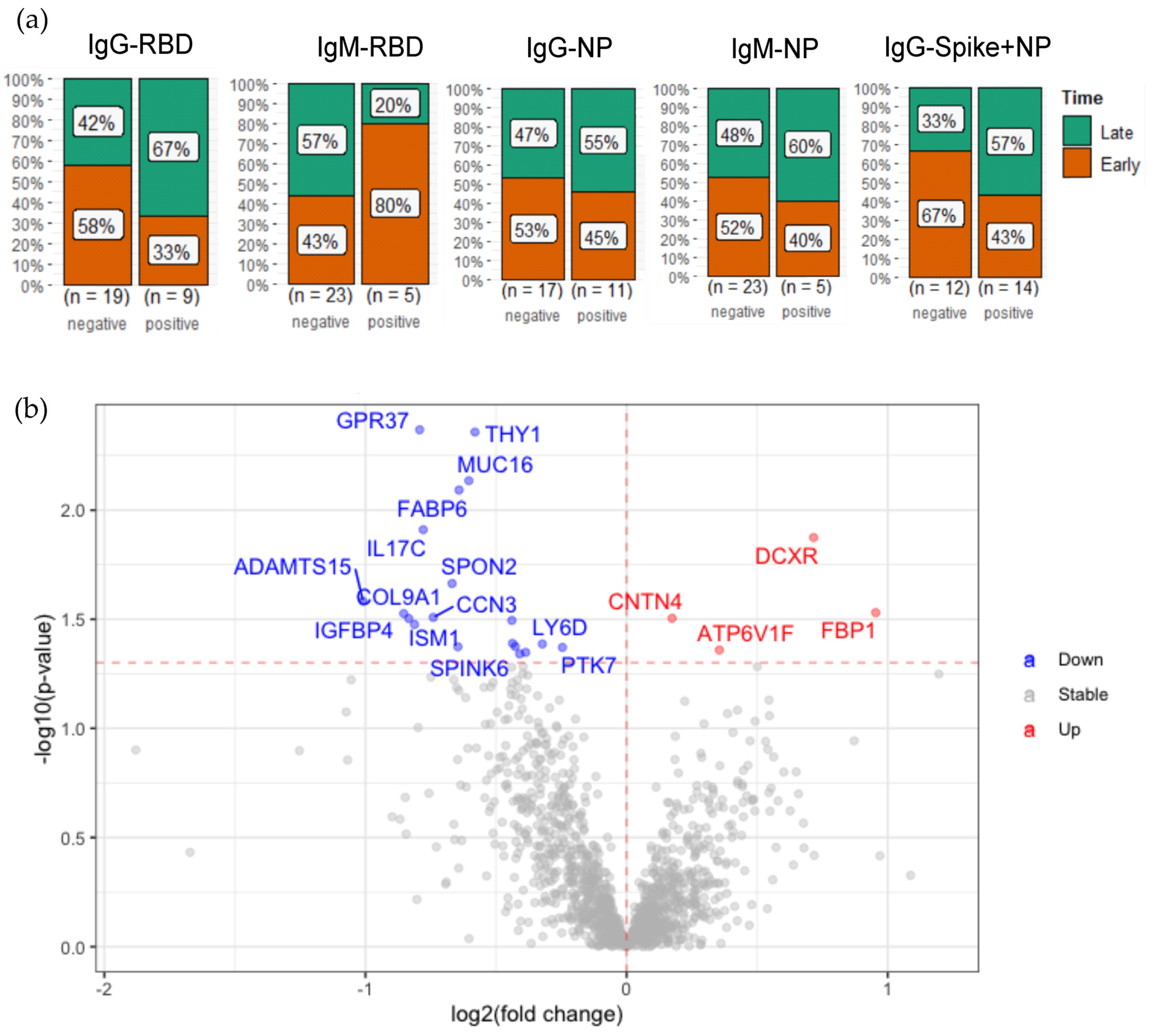
2.4. Plasma Protein Changes Associated with SARS-CoV-2 Antibody Generation
2.5. Identification of Key Immune Signatures and Novel Protein Changes Caused by a SARS-CoV-2 Infection
3. Discussion
4. Materials and Methods
4.1. Study Cohort
4.2. Proximity Extension Assay
4.3. Statistical Analyses
4.4. Enzyme-Linked Immunosorbent Assay (ELISA)
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WHO. COVID-19 Weekly Epidemiological Update; World Health Organization: Geneva, Switzerland, 2022; pp. 1–23. [Google Scholar]
- Pietro, R.D.; Calcagno, S.; Biondi-Zoccai, G.; Versaci, F. Is COVID-19 the deadliest event of the last century? Eur. Heart J. 2021, 42, 2876–2879. [Google Scholar] [CrossRef] [PubMed]
- Bergamaschi, L.; Mescia, F.; Turner, L.; Hanson, A.L.; Kotagiri, P.; Dunmore, B.J.; Ruffieux, H.; De Sa, A.; Huhn, O.; Morgan, M.D.; et al. Longitudinal analysis reveals that delayed bystander CD8+ T cell activation and early immune pathology distinguish severe COVID-19 from mild disease. Immunity 2021, 54, 1257–1275.e8. [Google Scholar] [CrossRef] [PubMed]
- Jamilloux, Y.; Henry, T.; Belot, A.; Viel, S.; Fauter, M.; El Jammal, T.; Walzer, T.; François, B.; Sève, P. Should we stimulate or suppress immune responses in COVID-19? Cytokine and anti-cytokine interventions. Autoimmun. Rev. 2020, 19, 102567. [Google Scholar] [CrossRef] [PubMed]
- Cao, X. COVID-19: Immunopathology and its implications for therapy. Nat. Rev. Immunol. 2020, 20, 269–270. [Google Scholar] [CrossRef] [Green Version]
- Selva, K.J.; Chung, A.W. Insights into how SARS-CoV2 infection induces cytokine storms. Trends Immunol. 2022, 43, 417–419. [Google Scholar] [CrossRef]
- Geng, J.; Chen, L.; Yuan, Y.; Wang, K.; Wang, Y.; Qin, C.; Wu, G.; Chen, R.; Zhang, Z.; Wei, D.; et al. CD147 antibody specifically and effectively inhibits infection and cytokine storm of SARS-CoV-2 and its variants delta, alpha, beta, and gamma. Signal Transduct. Target. Ther. 2021, 6, 347. [Google Scholar] [CrossRef]
- Marcelin, J.R.; Pettifor, A.; Janes, H.; Brown, E.R.; Kublin, J.G.; Stephenson, K.E. COVID-19 Vaccines and SARS-CoV-2 Transmission in the Era of New Variants: A Review and Perspective. Open Forum Infect. Dis. 2022, 9, ofac124. [Google Scholar] [CrossRef]
- Fathizadeh, H.; Afshar, S.; Masoudi, M.R.; Gholizadeh, P.; Asgharzadeh, M.; Ganbarov, K.; Köse, Ş.; Yousefi, M.; Kafil, H.S. SARS-CoV-2 (COVID-19) vaccines structure, mechanisms and effectiveness: A review. Int. J. Biol. Macromol. 2021, 188, 740–750. [Google Scholar] [CrossRef]
- Park, G.; Hwang, B.H. SARS-CoV-2 Variants: Mutations and Effective Changes. Biotechnol. Bioprocess Eng. 2021, 26, 859–870. [Google Scholar] [CrossRef]
- Fan, Y.; Li, X.; Zhang, L.; Wan, S.; Zhang, L.; Zhou, F. SARS-CoV-2 Omicron variant: Recent progress and future perspectives. Signal Transduct. Target. Ther. 2022, 7, 141. [Google Scholar] [CrossRef]
- Yong, S.J. Long COVID or post-COVID-19 syndrome: Putative pathophysiology, risk factors, and treatments. Infect. Dis. 2021, 53, 737–754. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Dong, X.; Liu, G.; Gao, Y. Risk and Protective Factors for COVID-19 Morbidity, Severity, and Mortality. Clin. Rev. Allergy Immunol. 2022, 64, 90–107. [Google Scholar] [CrossRef] [PubMed]
- Pennington, A.F.; Kompaniyets, L.; Summers, A.D.; Danielson, M.L.; Goodman, A.B.; Chevinsky, J.R.; Preston, L.E.; Schieber, L.Z.; Namulanda, G.; Courtney, J.; et al. Risk of Clinical Severity by Age and Race/Ethnicity Among Adults Hospitalized for COVID-19—United States, March–September 2020. Open Forum Infect. Dis. 2021, 8, ofaa638. [Google Scholar] [CrossRef]
- Liu, H.; Chen, S.; Liu, M.; Nie, H.; Lu, H. Comorbid Chronic Diseases are Strongly Correlated with Disease Severity among COVID-19 Patients: A Systematic Review and Meta-Analysis. Aging Dis. 2020, 11, 668–678. [Google Scholar] [CrossRef]
- Downes, D.J.; Cross, A.R.; Hua, P.; Roberts, N.; Schwessinger, R.; Cutler, A.J.; Munis, A.M.; Brown, J.; Mielczarek, O.; de Andrea, C.E.; et al. Identification of LZTFL1 as a candidate effector gene at a COVID-19 risk locus. Nat. Genet. 2021, 53, 1606–1615. [Google Scholar] [CrossRef] [PubMed]
- Pathania, A.S.; Prathipati, P.; Abdul, B.A.A.; Chava, S.; Katta, S.S.; Gupta, S.C.; Gangula, P.R.; Pandey, M.K.; Durden, D.L.; Byrareddy, S.N.; et al. COVID-19 and Cancer Comorbidity: Therapeutic Opportunities and Challenges. Theranostics 2021, 11, 731–753. [Google Scholar] [CrossRef]
- Li, G.; Chen, Z.; Lv, Z.; Li, H.; Chang, D.; Lu, J. Diabetes Mellitus and COVID-19: Associations and Possible Mechanisms. Int. J. Endocrinol. 2021, 2021, 7394378. [Google Scholar] [CrossRef]
- Nishiga, M.; Wang, D.W.; Han, Y.; Lewis, D.B.; Wu, J.C. COVID-19 and cardiovascular disease: From basic mechanisms to clinical perspectives. Nat. Rev. Cardiol. 2020, 17, 543–558. [Google Scholar] [CrossRef]
- Romagnolo, A.; Balestrino, R.; Imbalzano, G.; Ciccone, G.; Riccardini, F.; Artusi, C.A.; Bozzali, M.; Ferrero, B.; Montalenti, E.; Montanaro, E.; et al. Neurological comorbidity and severity of COVID-19. J. Neurol. 2021, 268, 762–769. [Google Scholar] [CrossRef]
- van Kessel, S.A.M.; Olde Hartman, T.C.; Lucassen, P.L.B.J.; van Jaarsveld, C.H.M. Post-acute and long-COVID-19 symptoms in patients with mild diseases: A systematic review. Fam. Pract. 2022, 39, 159–167. [Google Scholar] [CrossRef]
- Zhong, W.; Edfors, F.; Gummesson, A.; Bergström, G.; Fagerberg, L.; Uhlén, M. Next generation plasma proteome profiling to monitor health and disease. Nat. Commun. 2021, 12, 2493. [Google Scholar] [CrossRef] [PubMed]
- Perrotta, F.; Matera, M.G.; Cazzola, M.; Bianco, A. Severe respiratory SARS-CoV2 infection: Does ACE2 receptor matter? Respir. Med. 2020, 168, 105996. [Google Scholar] [CrossRef] [PubMed]
- Yeung, M.L.; Teng, J.L.L.; Jia, L.; Zhang, C.; Huang, C.; Cai, J.P.; Zhou, R.; Chan, K.H.; Zhao, H.; Zhu, L.; et al. Soluble ACE2-mediated cell entry of SARS-CoV-2 via interaction with proteins related to the renin-angiotensin system. Cell 2021, 184, 2212–2228.e12. [Google Scholar] [CrossRef] [PubMed]
- Jefferies, W.A.; Green, J.R.; Williams, A.F. Authentic T helper CD4 (W3/25) antigen on rat peritoneal macrophages. J. Exp. Med. 1985, 162, 117–127. [Google Scholar] [CrossRef] [PubMed]
- Peakman, M.; Senaldi, G.; Foote, N.; McManus, T.J.; Vergani, D. Naturally Occurring Soluble CD4 in Patients with Human Immunodeficiency Virus Infection. J. Infect. Dis. 1992, 165, 799–804. [Google Scholar] [CrossRef] [PubMed]
- Milani, D.; Caruso, L.; Zauli, E.; Al Owaifeer, A.M.; Secchiero, P.; Zauli, G.; Gemmati, D.; Tisato, V. p53/NF-kB Balance in SARS-CoV-2 Infection: From OMICs, Genomics and Pharmacogenomics Insights to Tailored Therapeutic Perspectives (COVIDomics). Front. Pharmacol. 2022, 13, 1754. [Google Scholar] [CrossRef]
- Tilborghs, S.; Corthouts, J.; Verhoeven, Y.; Arias, D.; Rolfo, C.; Trinh, X.B.; van Dam, P.A. The role of Nuclear Factor-kappa B signaling in human cervical cancer. Crit. Rev. Oncol. Hematol. 2017, 120, 141–150. [Google Scholar] [CrossRef]
- Wauman, J.; De Ceuninck, L.; Vanderroost, N.; Lievens, S.; Tavernier, J. RNF41 (Nrdp1) controls type 1 cytokine receptor degradation and ectodomain shedding. J. Cell Sci. 2011, 124, 921. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Li, L.F.; Munir, M.; Qiu, H.J. RING-domain E3 ligase-mediated host-virus interactions: Orchestrating immune responses by the host and antagonizing immune defense by viruses. Front. Immunol. 2018, 9, 1083. [Google Scholar] [CrossRef] [Green Version]
- Gordon, D.E.; Jang, G.M.; Bouhaddou, M.; Xu, J.; Obernier, K.; White, K.M.; O’Meara, M.J.; Rezelj, V.V.; Guo, J.Z.; Swaney, D.L.; et al. A SARS-CoV-2 protein interaction map reveals targets for drug repurposing. Nature 2020, 583, 459–468. [Google Scholar] [CrossRef]
- Ribero, M.S.; Jouvenet, N.; Dreux, M.; Nisole, S. Interplay between SARS-CoV-2 and the type I interferon response. PLoS Pathog. 2020, 16, e1008737. [Google Scholar] [CrossRef] [PubMed]
- Tullett, K.M.; Tan, P.S.; Park, H.Y.; Schittenhelm, R.B.; Michael, N.; Li, R.; Policheni, A.N.; Gruber, E.; Huang, C.; Fulcher, A.J.; et al. RNF41 regulates the damage recognition receptor Clec9a and antigen cross-presentation in mouse dendritic cells. eLife 2020, 9, e63452. [Google Scholar] [CrossRef] [PubMed]
- Kao, J.; Frankland, P.W. COVID fog demystified. Cell 2022, 185, 2391–2393. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Natesan, V.; Shi, H.; Hamik, A.; Kawanami, D.; Hao, C.; Mahabaleshwar, G.H.; Wang, W.; Jin, Z.G.; Atkins, G.B.; et al. A novel role of CCN3 in regulating endothelial inflammation. J. Cell Commun. Signal. 2010, 4, 141–153. [Google Scholar] [CrossRef] [Green Version]
- Gautier, T.; Deckert, V.; Nguyen, M.; Desrumaux, C.; Masson, D.; Lagrost, L. New therapeutic horizons for plasma phospholipid transfer protein (PLTP): Targeting endotoxemia, infection and sepsis. Pharmacol. Ther. 2022, 236, 108105. [Google Scholar] [CrossRef]
- Lauwers, M.; Au, M.; Yuan, S.; Wen, C. COVID-19 in Joint Ageing and Osteoarthritis: Current Status and Perspectives. Int. J. Mol. Sci. 2022, 23, 720. [Google Scholar] [CrossRef]
- Jia, W.; Li, H.; He, Y.-W. Pattern Recognition Molecule Mindin Promotes Intranasal Clearance of Influenza Viruses. J. Immunol. 2008, 180, 6255–6261. [Google Scholar] [CrossRef] [Green Version]
- Hou, Y.; Wang, T.; Ding, Y.; Yu, T.; Cui, Y.; Nie, H. Expression profiles of respiratory V-ATPase and calprotectin in SARS-CoV-2 infection. Cell Death Discov. 2022, 8, 362. [Google Scholar] [CrossRef]
- Jimenez, L.; Campos Codo, A.; de Souza Sampaio, V.; Oliveira, A.E.R.; Ferreira, L.K.K.; Davanzo, G.G.; de Brito Monteiro, L.; Victor Virgilio-da-Silva, J.; Borba, M.G.S.; Fabiano de Souza, G.; et al. Acid pH Increases SARS-CoV-2 Infection and the Risk of Death by COVID-19. Front. Med. 2021, 8, 637885. [Google Scholar] [CrossRef]
- Qi, M.; Liu, B.; Li, S.; Ni, Z.; Li, F. Construction and Investigation of Competing Endogenous RNA Networks and Candidate Genes Involved in SARS-CoV-2 Infection. Int. J. Gen. Med. 2021, 14, 6647–6659. [Google Scholar] [CrossRef]
- Laudanski, K.; Jihane, H.; Antalosky, B.; Ghani, D.; Phan, U.; Hernandez, R.; Okeke, T.; Wu, J.; Rader, D.; Susztak, K. Unbiased Analysis of Temporal Changes in Immune Serum Markers in Acute COVID-19 Infection With Emphasis on Organ Failure, Anti-Viral Treatment, and Demographic Characteristics. Front. Immunol. 2021, 12, 650465. [Google Scholar] [CrossRef] [PubMed]
- Ajaz, S.; McPhail, M.J.; Singh, K.K.; Mujib, S.; Trovato, F.M.; Napoli, S.; Agarwal, K. Mitochondrial metabolic manipulation by SARS-CoV-2 in peripheral blood mononuclear cells of patients with COVID-19. Am. J. Physiol.-Cell Physiol. 2021, 320, C57–C65. [Google Scholar] [CrossRef] [PubMed]
- Satış, H.; Özger, H.S.; Aysert Yıldız, P.; Hızel, K.; Gulbahar, Ö.; Erbaş, G.; Aygencel, G.; Guzel Tunccan, O.; Öztürk, M.A.; Dizbay, M.; et al. Prognostic value of interleukin-18 and its association with other inflammatory markers and disease severity in COVID-19. Cytokine 2021, 137, 155302. [Google Scholar] [CrossRef] [PubMed]
- Zaiss, D.M.W.; Gause, W.C.; Osborne, L.C.; Artis, D. Emerging functions of amphiregulin in orchestrating immunity, inflammation and tissue repair. Immunity 2015, 42, 216. [Google Scholar] [CrossRef] [Green Version]
- Harb, H.; Benamar, M.; Lai, P.S.; Contini, P.; Griffith, J.W.; Crestani, E.; Schmitz-Abe, K.; Chen, Q.; Fong, J.; Marri, L.; et al. Notch4 signaling limits regulatory T-cell-mediated tissue repair and promotes severe lung inflammation in viral infections. Immunity 2021, 54, 1186. [Google Scholar] [CrossRef]
- Feng, Y.; Duan, T.; Du, Y.; Jin, S.; Wang, M.; Cui, J.; Wang, R.F. LRRC25 Functions as an Inhibitor of NF-κB Signaling Pathway by Promoting p65/RelA for Autophagic Degradation. Sci. Rep. 2017, 7, 13448. [Google Scholar] [CrossRef] [Green Version]
- Du, Y.; Duan, T.; Feng, Y.; Liu, Q.; Lin, M.; Cui, J.; Wang, R.-F. LRRC25 inhibits type I IFN signaling by targeting ISG15-associated RIG-I for autophagic degradation. EMBO J. 2018, 37, 351–366. [Google Scholar] [CrossRef]
- Puig-Kröger, A.; Sierra-Filardi, E.; Domínguez-Soto, A.; Samaniego, R.; Corcuera, M.T.; Gómez-Aguado, F.; Ratnam, M.; Sánchez-Mateos, P.; Corbí, A.L. Folate Receptor β Is Expressed by Tumor-Associated Macrophages and Constitutes a Marker for M2 Anti-inflammatory/Regulatory Macrophages. Cancer Res. 2009, 69, 9395–9403. [Google Scholar] [CrossRef] [Green Version]
- Warmink, K.; Siebelt, M.; Low, P.S.; Riemers, F.M.; Wang, B.; Plomp, S.G.M.; Tryfonidou, M.A.; van Weeren, P.R.; Weinans, H.; Korthagen, N.M. Folate Receptor Expression by Human Monocyte–Derived Macrophage Subtypes and Effects of Corticosteroids. Cartilage 2022, 13, 194760352210814. [Google Scholar] [CrossRef]
- Ogilvie, P.; Paoletti, S.; Clark-Lewis, I.; Uguccioni, M. Eotaxin-3 is a natural antagonist for CCR2 and exerts a repulsive effect on human monocytes. Blood 2003, 102, 789–794. [Google Scholar] [CrossRef] [Green Version]
- Sugaya, M. Chemokines and Skin Diseases. Arch. Immunol. Ther. Exp. 2015, 63, 109–115. [Google Scholar] [CrossRef] [PubMed]
- Petkovic, V.; Moghini, C.; Paoletti, S.; Uguccioni, M.; Gerber, B. Eotaxin-3/CCL26 Is a Natural Antagonist for CC Chemokine Receptors 1 and 5: A Human Chemokine with a Regulatory Role. J. Biol. Chem. 2004, 279, 23357–23363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, Z.; Cao, L.; Guo, L.; Hong, Y.; Cao, J.; Chen, X. CCL26 regulates the proportion of CD4+CD25+FOXP3+ Tregs and the production of inflammatory factors in peripheral blood mononuclear cells following acute ischemic stroke via the STAT5 pathway. Exp. Ther. Med. 2020, 20, 3343–3351. [Google Scholar] [CrossRef] [PubMed]
- Su, C.M.; Wang, L.; Yoo, D. Activation of NF-κB and induction of proinflammatory cytokine expressions mediated by ORF7a protein of SARS-CoV-2. Sci. Rep. 2021, 11, 13464. [Google Scholar] [CrossRef] [PubMed]
- Vanderbeke, L.; Van Mol, P.; Van Herck, Y.; De Smet, F.; Humblet-Baron, S.; Martinod, K.; Antoranz, A.; Arijs, I.; Boeckx, B.; Bosisio, F.M.; et al. Monocyte-driven atypical cytokine storm and aberrant neutrophil activation as key mediators of COVID-19 disease severity. Nat. Commun. 2021, 12, 4117. [Google Scholar] [CrossRef] [PubMed]
- Abers, M.S.; Delmonte, O.M.; Ricotta, E.E.; Fintzi, J.; Fink, D.L.; Almeida de Jesus, A.A.; Zarember, K.A.; Alehashemi, S.; Oikonomou, V.; Desai, J.V.; et al. An immune-based biomarker signature is associated with mortality in COVID-19 patients. JCI Insight 2021, 6, e144455. [Google Scholar] [CrossRef]
- Kikly, K.; Liu, L.; Na, S.; Sedgwick, J.D. The IL-23/Th17 axis: Therapeutic targets for autoimmune inflammation. Curr. Opin. Immunol. 2006, 18, 670–675. [Google Scholar] [CrossRef]
- Blaner, W.S.; Brun, P.-J.; Calderon, R.M.; Golczak, M. Retinol-binding protein 2 (RBP2): Biology and pathobiology. Crit. Rev. Biochem. Mol. Biol. 2020, 55, 197–218. [Google Scholar] [CrossRef]
- Okada, I.; Hamanoue, H.; Terada, K.; Tohma, T.; Megarbane, A.; Chouery, E.; Abou-ghoch, J.; Jalkh, N.; Cogulu, O.; Ozkinay, F.; et al. SMOC1 Is Essential for Ocular and Limb Development in Humans and Mice. Am. J. Hum. Genet. 2011, 88, 30–41. [Google Scholar] [CrossRef] [Green Version]
- Duval, S.; Abu-Thuraia, A.; Elkholi, I.E.; Chen, R.; Seebun, D.; Mayne, J.; Côté, J.; Figeys, D.; Seidah, N.G. Shedding of cancer susceptibility candidate 4 by the convertases PC7/furin unravels a novel secretory protein implicated in cancer progression. Cell Death Dis. 2020, 11, 665. [Google Scholar] [CrossRef]
- da Silva Antunes, R.; Soldevila, F.; Pomaznoy, M.; Babor, M.; Bennett, J.; Tian, Y.; Khalil, N.; Qian, Y.; Mandava, A.; Scheuermann, R.H.; et al. A system-view of Bordetella pertussis booster vaccine responses in adults primed with whole-cell versus acellular vaccine in infancy. JCI Insight 2021, 6, e141023. [Google Scholar] [CrossRef] [PubMed]
- Gorreja, F.; Caër, C.; Rush, S.T.A.; Forsskål, S.K.; Härtlova, A.; Magnusson, M.K.; Bexe Lindskog, E.; Börjesson, L.G.; Block, M.; Wick, M.J. MEFV and NLRP3 Inflammasome Expression Is Attributed to Immature Macrophages and Correlates with Serum Inflammatory Proteins in Crohn’s Disease Patients. Inflammation 2022, 45, 1631–1650. [Google Scholar] [CrossRef] [PubMed]
- Henry, N.; Frank, J.; McLouth, C.; Trout, A.L.; Morris, A.; Chen, J.; Stowe, A.M.; Fraser, J.F.; Pennypacker, K. Short Chain Fatty Acids Taken at Time of Thrombectomy in Acute Ischemic Stroke Patients Are Independent of Stroke Severity But Associated With Inflammatory Markers and Worse Symptoms at Discharge. Front. Immunol. 2022, 12, 5883. [Google Scholar] [CrossRef] [PubMed]
- Heath, L.; Earls, J.C.; Magis, A.T.; Kornilov, S.A.; Lovejoy, J.C.; Funk, C.C.; Rappaport, N.; Logsdon, B.A.; Mangravite, L.M.; Kunkle, B.W.; et al. Manifestations of Alzheimer’s disease genetic risk in the blood are evident in a multiomic analysis in healthy adults aged 18 to 90. Sci. Rep. 2022, 12, 6117. [Google Scholar] [CrossRef] [PubMed]
- Dagnino, S.; Bodinier, B.; Guida, F.; Smith-Byrne, K.; Petrovic, D.; Whitaker, M.D.; Nøst, T.H.; Agnoli, C.; Palli, D.; Sacerdote, C.; et al. Prospective identification of elevated circulating CDCP1 in patients years before onset of lung cancer. Cancer Res. 2021, 81, 3738–3748. [Google Scholar] [CrossRef] [PubMed]
- Borges, L.; Kubin, M.; Kuhlman, T. LIR9, an immunoglobulin-superfamily-activating receptor, is expressed as a transmembrane and as a secreted molecule. Blood 2003, 101, 1484–1486. [Google Scholar] [CrossRef] [Green Version]
- Hammoudeh, S.M.; Hammoudeh, A.M.; Bhamidimarri, P.M.; Al Safar, H.; Mahboub, B.; Künstner, A.; Busch, H.; Halwani, R.; Hamid, Q.; Rahmani, M.; et al. Systems Immunology Analysis Reveals the Contribution of Pulmonary and Extrapulmonary Tissues to the Immunopathogenesis of Severe COVID-19 Patients. Front. Immunol. 2021, 12, 595150. [Google Scholar] [CrossRef]
- Kalia, N.; Singh, J.; Kaur, M. The role of dectin-1 in health and disease. Immunobiology 2021, 226, 152071. [Google Scholar] [CrossRef]
- Gadanec, L.K.; McSweeney, K.R.; Qaradakhi, T.; Ali, B.; Zulli, A.; Apostolopoulos, V. Can SARS-CoV-2 Virus Use Multiple Receptors to Enter Host Cells? Int. J. Mol. Sci. 2021, 22, 992. [Google Scholar] [CrossRef]
- Li, J.; Diao, B.; Guo, S.; Huang, X.; Yang, C.; Feng, Z.; Yan, W.; Ning, Q.; Zheng, L.; Chen, Y.; et al. VSIG4 inhibits proinflammatory macrophage activation by reprogramming mitochondrial pyruvate metabolism. Nat. Commun. 2017, 8, 1322. [Google Scholar] [CrossRef] [Green Version]
- Jung, K.; Seo, S.K.; Choi, I. Endogenous VSIG4 negatively regulates the helper T cell-mediated antibody response. Immunol. Lett. 2015, 165, 78–83. [Google Scholar] [CrossRef]
- Widyagarini, A.; Nishii, N.; Kawano, Y.; Zhang, C.; Azuma, M. VSIG4/CRIg directly regulates early CD8+ T cell activation through its counter-receptor in a narrow window. Biochem. Biophys. Res. Commun. 2022, 614, 100–106. [Google Scholar] [CrossRef]
- Saheb Sharif-Askari, N.; Saheb Sharif-Askari, F.; Mdkhana, B.; Al Heialy, S.; Alsafar, H.S.; Hamoudi, R.; Hamid, Q.; Halwani, R. Enhanced expression of immune checkpoint receptors during SARS-CoV-2 viral infection. Mol. Ther. Methods Clin. Dev. 2021, 20, 109–121. [Google Scholar] [CrossRef]
- Zheng, Y.; Ma, X.; Su, D.; Zhang, Y.; Yu, L.; Jiang, F.; Zhou, X.; Feng, Y.; Ma, F. The Roles of Siglec7 and Siglec9 on Natural Killer Cells in Virus Infection and Tumour Progression. J. Immunol. Res. 2020, 2020, 6243819. [Google Scholar] [CrossRef] [PubMed]
- Delaveris, C.S.; Wilk, A.J.; Riley, N.M.; Stark, J.C.; Yang, S.S.; Rogers, A.J.; Ranganath, T.; Nadeau, K.C.; Blish, C.A.; Bertozzi, C.R. Synthetic Siglec-9 Agonists Inhibit Neutrophil Activation Associated with COVID-19. ACS Cent. Sci. 2021, 7, 650–657. [Google Scholar] [CrossRef]
- Martin-Sancho, L.; Lewinski, M.K.; Pache, L.; Stoneham, C.A.; Yin, X.; Becker, M.E.; Pratt, D.; Churas, C.; Rosenthal, S.B.; Liu, S.; et al. Functional landscape of SARS-CoV-2 cellular restriction. Mol. Cell 2021, 81, 2656–2668.e8. [Google Scholar] [CrossRef] [PubMed]
- Altmann, D.M.; Boyton, R.J. SARS-CoV-2 T cell immunity: Specificity, function, durability, and role in protection. Sci. Immunol. 2020, 5, 6160. [Google Scholar] [CrossRef]
- Sica, G.L.; Zhu, G.; Tamada, K.; Liu, D.; Ni, J.; Chen, L. RELT, a new member of the tumor necrosis factor receptor superfamily, is selectively expressed in hematopoietic tissues and activates transcription factor NF-κB. Blood 2001, 97, 2702–2707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bradley, J.E.; Ramirez, G.; Hagood, J.S. Roles and Regulation of Thy-1, a Context-Dependent Modulator of Cell Phenotype. Biofactors 2009, 35, 258. [Google Scholar] [CrossRef] [Green Version]
- Furlong, S.; Coombs, M.R.P.; Ghassemi-Rad, J.; Hoskin, D.W. Thy-1 (CD90) Signaling Preferentially Promotes RORγt Expression and a Th17 Response. Front. Cell Dev. Biol. 2018, 6, 158. [Google Scholar] [CrossRef]
- Schröder, B. The multifaceted roles of the invariant chain CD74—More than just a chaperone. Biochim. Biophys. Acta Mol. Cell Res. 2016, 1863, 1269–1281. [Google Scholar] [CrossRef] [PubMed]
- Saverino, D.; Fabbi, M.; Merlo, A.; Ravera, G.; Grossi, C.E.; Ciccone, E. Surface density expression of the leukocyte-associated Ig-like receptor-1 is directly related to inhibition of human T-cell functions. Hum. Immunol. 2002, 63, 534–546. [Google Scholar] [CrossRef] [PubMed]
- Merlo, A.; Tenca, C.; Fais, F.; Battini, L.; Ciccone, E.; Grossi, C.E.; Saverino, D. Inhibitory receptors CD85j, LAIR-1, and CD152 down-regulate immunoglobulin and cytokine production by human B lymphocytes. Clin. Diagn. Lab. Immunol. 2005, 12, 705–712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Modabber, Z.; Shahbazi, M.; Akbari, R.; Bagherzadeh, M.; Firouzjahi, A.; Mohammadnia-Afrouzi, M. TIM-3 as a potential exhaustion marker in CD4+ T cells of COVID-19 patients. Immun. Inflamm. Dis. 2021, 9, 1707–1715. [Google Scholar] [CrossRef]
- Sansal, I.; Dupont, E.; Toru, D.; Evrard, C.; Rouget, P. NPDC-1, a regulator of neural cell proliferation and differentiation, interacts with E2F-1, reduces its binding to DNA and modulates its transcriptional activity. Oncogene 2000, 19, 5000–5009. [Google Scholar] [CrossRef] [Green Version]
- Rezzola, S.; Di Somma, M.; Corsini, M.; Leali, D.; Ravelli, C.; Polli, V.A.B.; Grillo, E.; Presta, M.; Mitola, S. VEGFR2 activation mediates the pro-angiogenic activity of BMP4. Angiogenesis 2019, 22, 521–533. [Google Scholar] [CrossRef]
- Sur, S.; Khatun, M.; Steele, R.; Isbell, T.S.; Ray, R.; Ray, R.B. Exosomes from COVID-19 Patients Carry Tenascin-C and Fibrinogen-β in Triggering Inflammatory Signals in Cells of Distant Organ. Int. J. Mol. Sci. 2021, 22, 3184. [Google Scholar] [CrossRef]
- Chi, Y.; Chai, J.; Xu, C.; Luo, H.; Zhang, Q. The extracellular matrix protein matrilin-2 induces post-burn inflammatory responses as an endogenous danger signal. Inflamm. Res. 2015, 64, 833–839. [Google Scholar] [CrossRef]
- Ladikou, E.E.; Sivaloganathan, H.; Milne, K.M.; Arter, W.E.; Ramasamy, R.; Saad, R.; Stoneham, S.M.; Philips, B.; Eziefula, A.C.; Chevassut, T. Von Willebrand factor (vWF): Marker of endothelial damage and thrombotic risk in COVID-19? Clin. Med. 2020, 20, e178–e182. [Google Scholar] [CrossRef]
- Babkina, A.S.; Ostrova, I.V.; Yadgarov, M.Y.; Kuzovlev, A.N.; Grechko, A.V.; Volkov, A.V.; Golubev, A.M. The Role of Von Willebrand Factor in the Pathogenesis of Pulmonary Vascular Thrombosis in COVID-19. Viruses 2022, 14, 211. [Google Scholar] [CrossRef]
- Olink Team. Olink; Assay Validation. Available online: https://olink.com/our-platform/assay-validation/ (accessed on 1 March 2023).
- R Core Team. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022; Available online: https://www.R-project.org/ (accessed on 12 December 2022).
- RStudio Team. RStudio: Integrated Development Environment for R; RStudio, PBC: Boston, MA, USA, 2022; Available online: http://www.rstudio.com/ (accessed on 12 December 2022).
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Ulgen, E.; Ozisik, O.; Sezerman, O.U. PathfindR: An R package for comprehensive identification of enriched pathways in omics data through active subnetworks. Front. Genet. 2019, 10, 858. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| COVID-19 Patients | Sex | Age (Years) | Disease Control (DC) | Healthy Control | Sampling <3 Months of Infection | Antibodies * | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| DC1 | DC2 | IgG RBD | IgG NP | IgG (Spike + NP) | IgM RBD | IgM NP | |||||
| Case 1 | F | 55 | + | ++ | + | ++ | – | + | |||
| Case 2 | M | 78 | + | ++ | + | +++ | – | – | |||
| Case 3 | F | 41 | + | + | – | + | + | + | – | ||
| Case 4 | F | 71 | + | + | + | – | – | – | – | – | |
| Case 5 | F | 36 | + | – | – | – | – | – | |||
| Case 6 | F | 71 | + | + | – | – | – | – | – | ||
| Case 7 | F | 54 | + | + | + | – | – | – | – | – | |
| Case 8 | F | 58 | + | + | +++ | ++ | +++ | – | – | ||
| Case 10 | F | 53 | + | + | + | – | – | – | – | ++ | |
| Case 11 | M | 64 | + | + | +++ | +++ | +++ | + | +++ | ||
| Case 12 | F | 61 | + | + | + | – | – | + | – | – | |
| Case 13 | F | 27 | + | + | + | – | + | + | – | – | |
| Case 14 | F | 45 | + | + | + | + | + | + | + | – | |
| Case 16 | M | 52 | + | + | + | + | – | + | – | – | |
| Case 17 | M | 64 | + | + | – | – | – | – | – | ||
| Case 18 | F | 35 | + | – | + | + | – | + | |||
| Case 19 | F | 31 | + | – | – | – | – | – | |||
| Case 20 | F | 23 | + | + | – | – | – | + | – | ||
| Case 21 | F | 53 | + | + | + | +++ | +++ | +++ | + | – | |
| Case 22 | F | 30 | + | – | – | + | – | – | |||
| Case 25 | F | 27 | + | + | – | – | – | – | – | ||
| Case 26 | M | 62 | + | + | + | – | – | – | – | – | |
| Case 27 | F | 79 | + | + | + | – | – | – | – | – | |
| Case 28 | F | 36 | + | + | + | – | – | – | – | – | |
| Case 29 | M | 74 | + | + | + | – | – | – | – | – | |
| Case 30 | F | 19 | + | + | + | – | – | – | – | – | |
| Case 31 | M | 72 | + | + | + | ++ | +++ | – | + | ||
| Case 32 | F | 53 | + | + | + | +++ | – | – | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Urbiola-Salvador, V.; Lima de Souza, S.; Grešner, P.; Qureshi, T.; Chen, Z. Plasma Proteomics Unveil Novel Immune Signatures and Biomarkers upon SARS-CoV-2 Infection. Int. J. Mol. Sci. 2023, 24, 6276. https://doi.org/10.3390/ijms24076276
Urbiola-Salvador V, Lima de Souza S, Grešner P, Qureshi T, Chen Z. Plasma Proteomics Unveil Novel Immune Signatures and Biomarkers upon SARS-CoV-2 Infection. International Journal of Molecular Sciences. 2023; 24(7):6276. https://doi.org/10.3390/ijms24076276
Chicago/Turabian StyleUrbiola-Salvador, Víctor, Suiane Lima de Souza, Peter Grešner, Talha Qureshi, and Zhi Chen. 2023. "Plasma Proteomics Unveil Novel Immune Signatures and Biomarkers upon SARS-CoV-2 Infection" International Journal of Molecular Sciences 24, no. 7: 6276. https://doi.org/10.3390/ijms24076276