
Physiological Rhythms and Biological Variation of Biomolecules: The Road to Personalized Laboratory Medicine


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Biological Rhythms
2.1. Ultradian Rhythms
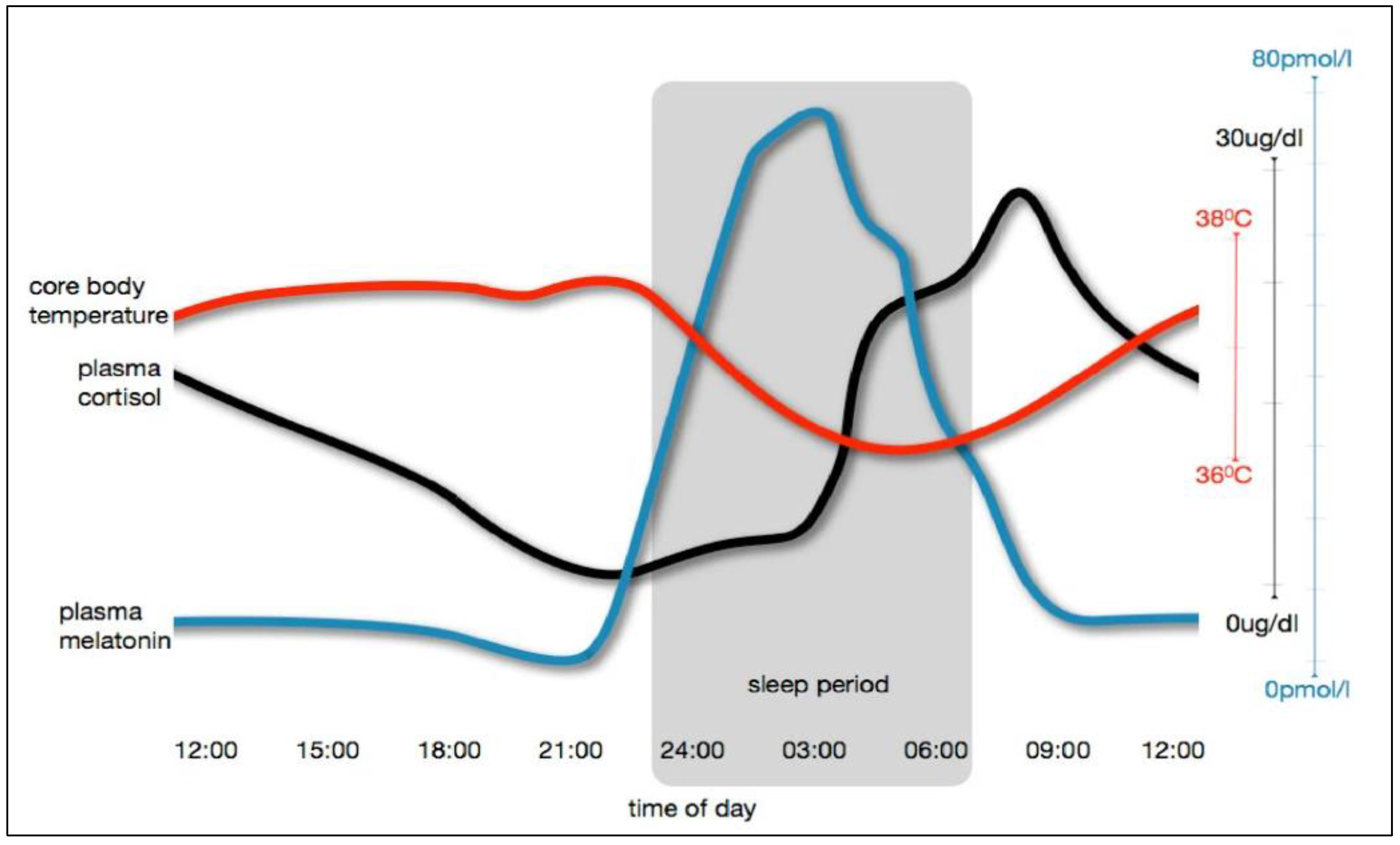
2.2. Circadian Rhythm

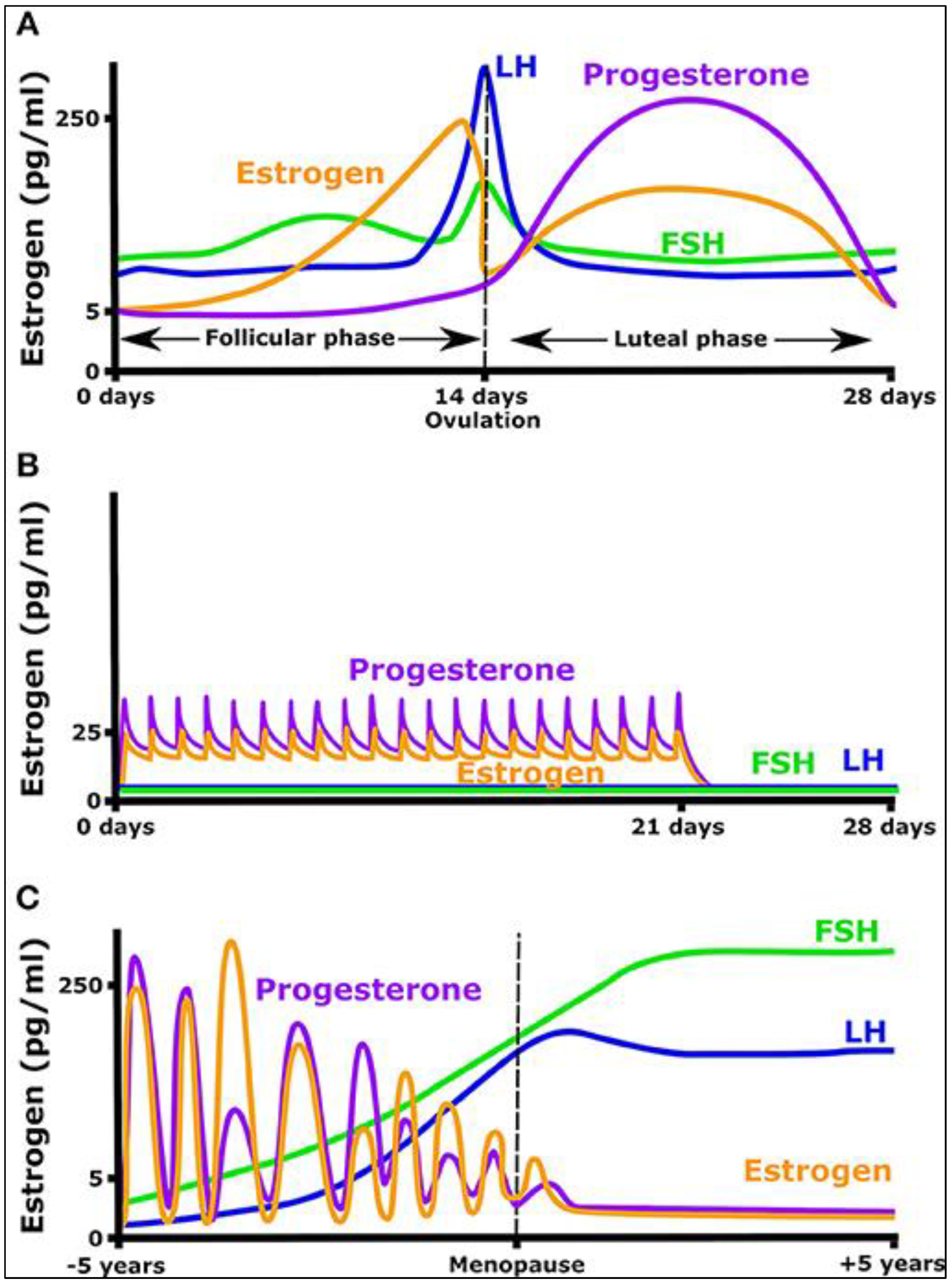
2.3. Infradian Rhythms
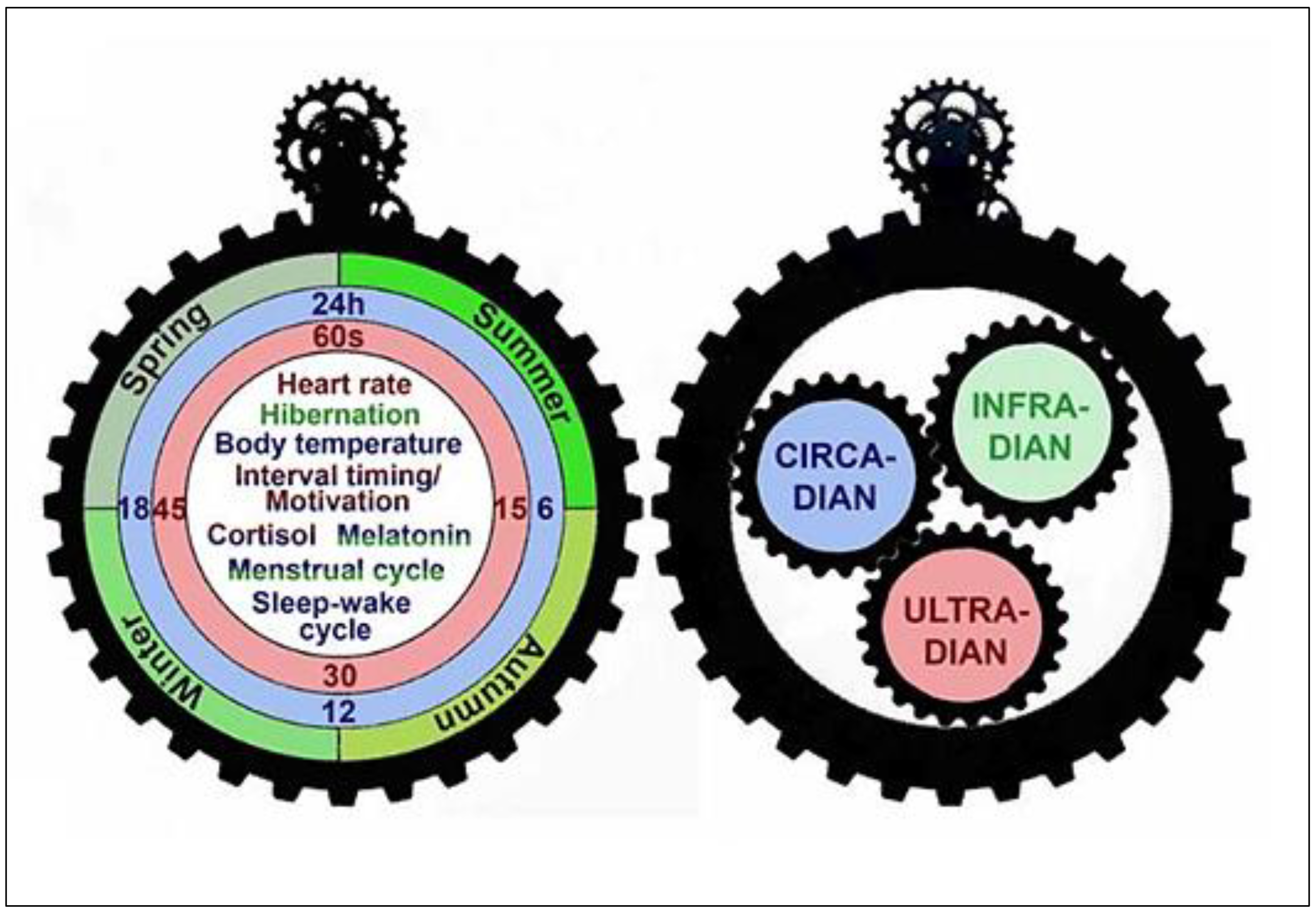
2.4. Interactions among Ultradian, Circadian, and Infradian Rhythms
2.5. Disruption of Biological Rhythms
3. Biological Variation
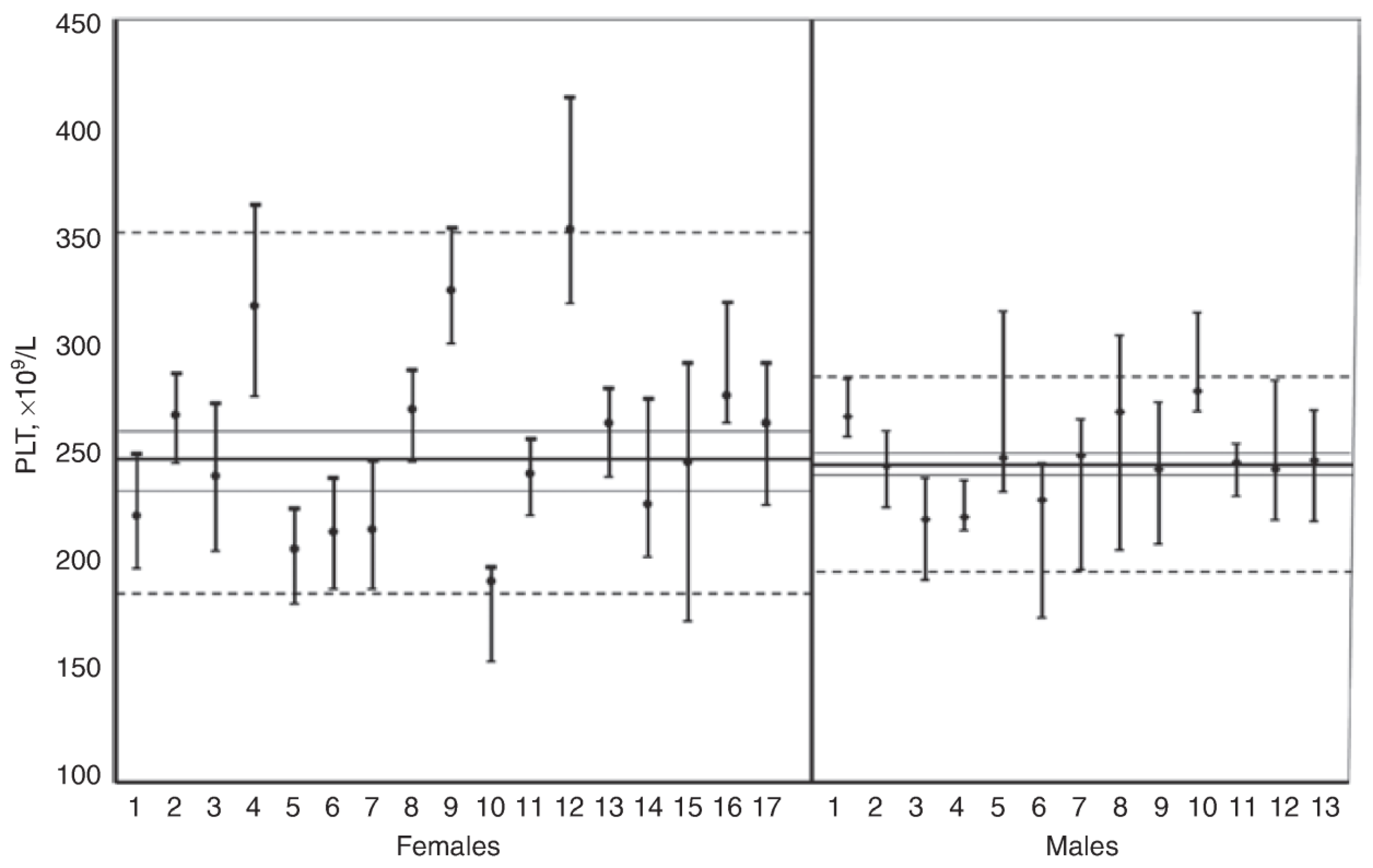
3.1. Between-Subject Biological Variation
3.2. Within-Subject Biological Variation
3.3. Within-Person Biological Variation
3.4. Clinical Applications of Biological Variation Data

3.5. Reliability of Biological Variation Data
3.6. Biological Variation in Diseases
4. Physiological Rhythms and Reference Intervals
5. Physiological Rhythms and Reference Change Value
6. Physiological Rhythms and Chronotypes
7. 3P Medicine and Variations
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ade, P.A.; Aghanim, N.; Arnaud, M.; Ashdown, M.; Aumont, J.; Baccigalupi, C.; Banday, A.J.; Barreiro, R.B.; Bartlett, J.G.; Bartolo, N.; et al. Planck 2015 results-xiii. cosmological parameters. Astron. Astrophys. 2016, 594, A13. [Google Scholar]
- Graham, D.W. Heraclitus: Flux, order, and knowledge. In The Oxford Handbook of Presocratic Philosophy; Curd, P., Graham, D.W., Eds.; Oxford University Press: Oxford, UK, 2008; pp. 169–188. [Google Scholar]
- Rosen, I.M. Change is the Only Constant in Life (and in Sleep Medicine). J. Clin. Sleep Med. 2018, 14, 1025–1030. [Google Scholar] [CrossRef] [PubMed]
- Tu, Y.; Rappel, W.J. Adaptation in Living Systems. Annu. Rev. Condens. Matter Phys. 2018, 9, 183–205. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, T. Biologically inspired information theory: Adaptation through construction of external reality models by living systems. Prog. Biophys. Mol. Biol. 2015, 119, 634–648. [Google Scholar] [CrossRef]
- Chatterjee, A.; Georgiev, G.; Iannacchione, G. Aging and efficiency in living systems: Complexity, adaptation and self-organization. Mech. Ageing Dev. 2017, 163, 2–7. [Google Scholar] [CrossRef]
- Buijink, M.R.; Olde Engberink, A.H.; Wit, C.B.; Almog, A.; Meijer, J.H.; Rohling, J.H.; Michel, S. Aging Affects the Capacity of Photoperiodic Adaptation Downstream from the Central Molecular Clock. J. Biol. Rhythm. 2020, 35, 167–179. [Google Scholar] [CrossRef] [Green Version]
- Saderi, N.; Escobar, C.; Salgado-Delgado, R. Alteration of biological rhythms causes metabolic diseases and obesity. Rev. Neurol. 2013, 57, 71–78. [Google Scholar]
- Smith, S.K.; Musiek, E.S. Impact of circadian and diurnal rhythms on cellular metabolic function and neurodegenerative diseases. Int. Rev. Neurobiol. 2020, 154, 393–412. [Google Scholar]
- Tong, X.; Yin, L. Circadian Rhythms in Liver Physiology and Liver Diseases. Compr. Physiol. 2013, 3, 917–940. [Google Scholar]
- Feriel, J.; Tchipeva, D.; Depasse, F. Effects of circadian variation, lifestyle and environment on hematological parameters: A narrative review. Int. J. Lab. Hematol. 2021, 43, 917–926. [Google Scholar] [CrossRef]
- Hernández-García, J.; Navas-Carrillo, D.; Orenes-Piñero, E. Alterations of circadian rhythms and their impact on obesity, metabolic syndrome and cardiovascular diseases. Crit. Rev. Food Sci. Nutr. 2020, 60, 1038–1047. [Google Scholar] [CrossRef] [PubMed]
- Seifalian, A.; Hart, A. Circadian Rhythms: Will It Revolutionise the Management of Diseases? J. Lifestyle Med. 2019, 9, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Prendergast, B.J.; Zucker, I. Ultradian rhythms in mammalian physiology and behavior. Curr. Opin. Neurobiol. 2016, 40, 150–154. [Google Scholar] [CrossRef] [PubMed]
- Laerum, O.D.; Smaaland, R. Circadian and infradian aspects of the cell cycle: From past to future. Chronobiologia 1989, 16, 441–453. [Google Scholar]
- Lamont, E.W.; Amir, S. Circadian and ultradian clocks/rhythms. In Encyclopedia of Behavioral Neuroscience, 2nd ed.; Della Sala, S., Ed.; Academic Press: London, UK, 2017; pp. 746–751. [Google Scholar]
- Moser, M.; Penter, R.; Fruehwirth, M.; Kenner, T. Why life oscillates-Biological rhythms and health. In Proceedings of the 2006 International Conference of the IEEE Engineering in Medicine and Biology Society, New York, NY, USA, 30 August–3 September 2006. [Google Scholar]
- Harmer, S.L.; Panda, S.; Kay, S.A. Molecular Bases of Circadian Rhythms. Annu. Rev. Cell Dev. Biol. 2001, 17, 215–253. [Google Scholar] [CrossRef] [Green Version]
- Fagiani, F.; Di Marino, D.; Romagnoli, A.; Travelli, C.; Voltan, D.; Di Cesare Mannelli, L.; Racchi, M.; Govoni, S.; Lanni, C. Molecular regulations of circadian rhythm and implications for physiology and diseases. Signal Transduct. Target. Ther. 2022, 7, 41. [Google Scholar] [CrossRef]
- Demas, G.E.; Kriegsfeld, L.J. Biological Rhythms. Encycl. Endocr. Dis. 2004, 7, 345–351. [Google Scholar]
- Golombek, D.A.; Bussi, I.L.; Agostino, P.V. Minutes, days and years: Molecular interactions among different scales of biological timing. Philos. Trans. R. Soc. B Biol. Sci. 2014, 369, 20120465. [Google Scholar] [CrossRef] [Green Version]
- Tabibzadeh, S. CircadiOmic medicine and aging. Ageing Res. Rev. 2021, 71, 101424. [Google Scholar] [CrossRef]
- Aschoff, J. Aschoff, J. A Survey on Biological Rhythms. In Biological Rhythms, 1st ed.; Aschoff, J., Ed.; Springer: Boston, MA, USA, 1981; pp. 3–10. [Google Scholar]
- Goh, G.H.; Maloney, S.K.; Mark, P.J.; Blache, D. Episodic Ultradian Events—Ultradian Rhythms. Biology 2019, 8, 15. [Google Scholar] [CrossRef] [Green Version]
- Kippert, F.; Hunt, P. Ultradian clocks in eukaryotic microbes: From behavioural observation to functional genomics. Bioessays 2000, 22, 16–22. [Google Scholar] [CrossRef]
- Kippert, F. The ultradian clocks of eukaryotic microbes: Timekeeping devices displaying a homeostasis of the period. Chronobiol. Int. 1997, 14, 469–479. [Google Scholar] [CrossRef] [PubMed]
- Brodsky, V.Y. Circahoralian (Ultradian) metabolic rhythms. Biochemistry 2014, 79, 483–495. [Google Scholar] [CrossRef]
- Lloyd, D.; Murray, D.B. Ultradian metronome: Timekeeper for orchestration of cellular coherence. Trends Biochem. Sci. 2005, 30, 373–377. [Google Scholar] [CrossRef]
- López-Otín, C.; Kroemer, G. Hallmarks of Health. Cell 2021, 184, 33–63. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Irusta, A.; Kembro, J.M.; Domínguez, E.M.; Matamoros-Volante, A.; Gallea, M.N.; Molina, R.; Guidobaldi, H.A.; Treviño, C.L.; Figueras, M.J.; Babini, A.; et al. Sperm physiology varies according to ultradian and infradian rhythms. Sci. Rep. 2019, 9, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blessing, W.W. Thermoregulation and the ultradian basic rest-activity cycle. Handb. Clin. Neurol. 2018, 156, 367–375. [Google Scholar] [PubMed]
- Lindsley, G.; Dowse, H.B.; Burgoon, P.W.; Kolka, M.A.; Stephenson, L.A. A persistent circhoral ultradian rhythm is identified in human core temperature. Chronobiol. Int. 1999, 16, 69–78. [Google Scholar] [CrossRef]
- Morf, J.; Schibler, U. Body temperature cycles: Gatekeepers of circadian clocks. Cell Cycle 2013, 12, 539–540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moszczynski, A.; Murray, B.J. Neurobiological aspects of sleep physiology. Neurol. Clin. 2012, 30, 963–985. [Google Scholar] [CrossRef]
- Isomura, A.; Kageyama, R. Ultradian oscillations and pulses: Coordinating cellular responses and cell fate decisions. Development 2014, 141, 3627–3636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilbert, D.; Lloyd, D. The living cell: A complex autodynamic multi-oscillator system? Cell Biol. Int. 2000, 24, 569–580. [Google Scholar] [CrossRef] [PubMed]
- Lloyd, D.; Aon, M.A.; Cortassa, S. Why homeodynamics, not homeostasis? Sci. World J. 2001, 1, 133–145. [Google Scholar] [CrossRef] [Green Version]
- Nunemaker, C.S.; Satin, L.S. Episodic hormone secretion: A comparison of the basis of pulsatile secretion of insulin and GnRH. Endocrine 2014, 47, 49–63. [Google Scholar] [CrossRef] [Green Version]
- Nuñez, L.; Faught, W.J.; Frawley, L.S. Episodic gonadotropin-releasing hormone gene expression revealed by dynamic monitoring of luciferase reporter activity in single, living neurons. Proc. Natl. Acad. Sci. USA 1998, 95, 9648–9653. [Google Scholar] [CrossRef] [Green Version]
- Rizi, E.P.; Baig, S.; Toh, S.A.; Loh, T.P.; Khoo, C.M. Biological variation of glucose, insulin and lipids in lean, insulin-sensitive and obese, insulin-resistant Chinese males without diabetes. Pathology 2016, 48, 510–512. [Google Scholar] [CrossRef] [PubMed]
- Simon, C. Ultradian pulsatility of plasma glucose and insulin secretion rate: Circadian and sleep modulation. Horm. Res. 1998, 49, 185–190. [Google Scholar] [CrossRef]
- Halberg, F. Physiologic 24-hour periodicity; general and procedural considerations with reference to the adrenal cycle. Int. Z. Vitam. Beih 1959, 10, 225–296. [Google Scholar]
- Skeldon, A.C.; Phillips, A.J.K.; Dijk, D.J. The effects of self-selected light-dark cycles and social constraints on human sleep and circadian timing: A modeling approach. Sci. Rep. 2017, 7, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Scheer, F.A.; Wright, K.P., Jr.; Kronauer, R.E.; Czeisler, C.A. Plasticity of the intrinsic period of the human circadian timing system. PLoS ONE 2007, 2, 721. [Google Scholar] [CrossRef] [Green Version]
- Escobar, C.; Rojas-Granados, A.; Angeles-Castellanos, M. Development of the circadian system and relevance of periodic signals for neonatal development. Handb. Clin. Neurol. 2021, 179, 249–258. [Google Scholar] [PubMed]
- Peek, C.B.; Ramsey, K.M.; Marcheva, B.; Bass, J. Nutrient sensing and the circadian clock. Trends Endocrinol. Metab. 2012, 23, 312–318. [Google Scholar] [CrossRef] [Green Version]
- Hughes, S.; Jagannath, A.; Hankins, M.W.; Foster, R.G.; Peirson, S.N. Photic Regulation of Clock Systems. Methods Enzym. 2015, 552, 125–143. [Google Scholar]
- Patton, A.P.; Hastings, M.H. The suprachiasmatic nucleus. Curr. Biol. 2018, 28, R816–R822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosenwasser, A.M.; Turek, F.W. Neurobiology of Circadian Rhythm Regulation. Sleep Med. Clin. 2015, 10, 403–412. [Google Scholar] [CrossRef]
- Pacheco-Bernal, I.; Becerril-Pérez, F.; Aguilar-Arnal, L. Circadian rhythms in the three-dimensional genome: Implications of chromatin interactions for cyclic transcription. Clin. Epigenet. 2019, 11, 79. [Google Scholar] [CrossRef]
- Kalsbeek, A.; La Fleur, S.; Fliers, E. Circadian control of glucose metabolism. Mol. Metab. 2014, 3, 372–383. [Google Scholar] [CrossRef]
- Pickel, L.; Sung, H.K. Feeding Rhythms and the Circadian Regulation of Metabolism. Front. Nutr. 2020, 7, 39. [Google Scholar] [CrossRef]
- Zhang, Z.; Shui, G.; Li, M.D. Time to eat reveals the hierarchy of peripheral clocks. Trends Cell Biol. 2021, 31, 869–872. [Google Scholar] [CrossRef]
- Kuehn, B.M. Resetting the Circadian Clock Might Boost Metabolic Health. JAMA 2017, 317, 1303–1305. [Google Scholar] [CrossRef]
- Astiz, M.; Heyde, I.; Oster, H. Mechanisms of communication in the mammalian circadian timing system. Int. J. Mol. Sci. 2019, 20, 343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamont, E.W.; Renteria Diaz, L.; Barry-Shaw, J.; Stewart, J.; Amir, S. Daily restricted feeding rescues a rhythm of period2 expression in the arrhythmic suprachiasmatic nucleus. Neuroscience 2005, 132, 245–248. [Google Scholar] [CrossRef] [Green Version]
- Ledford, H.; Callaway, E. Circadian clocks scoop Nobel prize. Nature 2017, 550, 18. [Google Scholar] [CrossRef] [Green Version]
- Lande-Diner, L.; Boyault, C.; Kim, J.Y.; Weitz, C.J. A positièe feedback loop links circadian clock factor CLOCK-BMAL1 to the basic transcriptional machinery. Proc. Natl. Acad. Sci. USA 2013, 110, 16021–16026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pett, J.P.; Korenčič, A.; Wesener, F.; Kramer, A.; Herzel, H. Feedback Loops of the Mammalian Circadian Clock Constitute Repressilator. PLoS Comput. Biol. 2016, 12, e1005266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, L.S.; Doyle, F.J. A dual-feedback loop model of the mammalian circadian clock for multi-input control of circadian phase. PLoS Comput. Biol. 2020, 16, e1008459. [Google Scholar] [CrossRef] [PubMed]
- Gaucher, J.; Montellier, E.; Sassone-Corsi, P. Molecular Cogs: Interplay between Circadian Clock and Cell Cycle. Trends Cell Biol. 2018, 28, 368–379. [Google Scholar] [CrossRef]
- Takahashi, J.S. Transcriptional architecture of the mammalian circadian clock. Nat. Rev. Genet. 2016, 18, 164–179. [Google Scholar] [CrossRef] [Green Version]
- Reddy, A.B.; O’Neill, J.S. Healthy clocks, healthy body, healthy mind. Trends Cell Biol. 2010, 20, 36–44. [Google Scholar] [CrossRef] [Green Version]
- Tomita, J.; Nakajima, M.; Kondo, T.; Iwasaki, H. No Transcription-Translation Feedback in Circadian Rhythm of KaiC Phosphorylation. Science 2005, 307, 251–254. [Google Scholar] [CrossRef]
- Lakin-Thomas, P.L. Transcriptional feedback oscillators: Maybe, maybe not…. J. Biol. Rhythm. 2006, 21, 83–92. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, J.S.; Reddy, A.B. Circadian clocks in human red blood cells. Nature 2011, 469, 498–503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beale, A.D.; Labeed, F.H.; Kitcatt, S.J.; O’Neill, J.S. Detecting Circadian Rhythms in Human Red Blood Cells by Dielectrophoresis. Methods Mol. Biol. 2022, 2482, 255–264. [Google Scholar] [PubMed]
- Hall, A.; Karplus, P.A.; Poole, L.B. Typical 2-Cys peroxiredoxins–structures, mechanisms and functions. FEBS J. 2009, 276, 2469–2477. [Google Scholar] [CrossRef] [Green Version]
- Perkins, A.; Nelson, K.J.; Parsonage, D.; Poole, L.B.; Karplus, P.A. Peroxiredoxins: Guardians against oxidative stress and modulators of peroxide signaling. Trends Biochem. Sci. 2015, 40, 435–445. [Google Scholar] [CrossRef] [Green Version]
- Ceglia, N.; Liu, Y.; Chen, S.; Agostinelli, F.; Eckel-Mahan, K.; Sassone-Corsi, P.; Baldi, P. CircadiOmics: Circadian omic web portal. Nucleic Acids Res. 2018, 46, W157–W162. [Google Scholar] [CrossRef] [Green Version]
- Haus, E. Chronobiology in the endocrine system. Adv. Drug Deliv. Rev. 2007, 59, 985–1014. [Google Scholar] [CrossRef]
- Loef, B.; Nanlohy, N.M.; Jacobi, R.H.; van de Ven, C.; Mariman, R.; van der Beek, A.J.; Proper, K.I.; van Baarle, D. Immunological effects of shift work in healthcare workers. Sci. Rep. 2019, 9, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Souto Filho, J.T.D.; Portugal, R.D.; Nucci, M. Effect of circadian variation on neutrophil mobilization to the peripheral blood in benign constitutional neutropenia. Exp. Hematol. 2019, 69, 22–26. [Google Scholar] [CrossRef] [Green Version]
- Ackermann, K.; Revell, V.L.; Lao, O.; Rombouts, E.J.; Skene, D.J.; Kayser, M. Diurnal rhythms in blood cell populations and the effect of acute sleep deprivation in healthy young men. Sleep 2012, 35, 933–940. [Google Scholar] [CrossRef] [Green Version]
- Sennels, H.P.; Jørgensen, H.L.; Hansen, A.L.S.; Goetze, J.P.; Fahrenkrug, J. Diurnal variation of hematology parameters in healthy young males: The Bispebjerg study of diurnal variations. Scand. J. Clin. Lab. Investig. 2011, 71, 532–541. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Pollock, D.M. Diurnal Regulation of Renal Electrolyte Excretion: The Role of Paracrine Factors. Annu. Rev. Physiol. 2020, 82, 343–363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hongo, T.; Suzuki, T.; Ishida, H.; Kabuto, M.; Neriishi, K. Diurnal variation of plasma minerals and trace elements in a group of Japanese male adults. J. Nutr. Sci. Vitaminol. 1993, 39, 33–46. [Google Scholar] [CrossRef] [Green Version]
- Scales, W.E.; Vander, A.J.; Brown, M.B.; Kluger, M.J. Human circadian rhythms in temperature, trace metals, and blood variables. J. Appl. Physiol. 1988, 65, 1840–1846. [Google Scholar] [CrossRef]
- Zhao, L.; Hutchison, A.T.; Heilbronn, L.K. Carbohydrate intake and circadian synchronicity in the regulation of glucose homeostasis. Curr. Opin. Clin. Nutr. Metab. Care 2021, 24, 342–348. [Google Scholar] [CrossRef] [PubMed]
- Sato, K.; Meng, F.; Francis, H.; Wu, N.; Chen, L.; Kennedy, L.; Zhou, T.; Franchitto, A.; Onori, P.; Gaudio, E.; et al. Melatonin and circadian rhythms in liver diseases: Functional roles and potential therapies. J. Pineal Res. 2020, 68, e12639. [Google Scholar] [CrossRef] [PubMed]
- Gnocchi, D.; Bruscalupi, G. Circadian Rhythms and Hormonal Homeostasis: Pathophysiological Implications. Biology 2017, 6, 10. [Google Scholar] [CrossRef] [Green Version]
- Nava Zisapel, C.; Zisapel, N. New perspectives on the role of melatonin in human sleep, circadian rhythms and their regulation. Br. J. Pharmacol. 2018, 175, 3190–3199. [Google Scholar] [CrossRef] [Green Version]
- Chan, S.; Debono, M. Replication of cortisol circadian rhythm: New advances in hydrocortisone replacement therapy. Ther. Adv. Endocrinol. Metab. 2010, 1, 129–138. [Google Scholar] [CrossRef]
- Hickie, I.B.; Naismith, S.L.; Robillard, R.; Scott, E.M.; Hermens, D.F. Manipulating the sleep-wake cycle and circadian rhythms to improve clinical management of major depression. BMC Med. 2013, 11, 79. [Google Scholar] [CrossRef] [Green Version]
- Laje, R.; Agostino, P.V.; Golombek, D.A. The Times of Our Lives: Interaction Among Different Biological Periodicities. Front. Integr. Neurosci. 2018, 12, 10. [Google Scholar] [CrossRef] [Green Version]
- Goldman, B.D. Mammalian photoperiodic system: Formal properties and neuroendocrine mechanisms of photoperiodic time measurement. J. Biol. Rhythm. 2001, 16, 283–301. [Google Scholar] [CrossRef]
- Postolache, T.T.; Raheja, U.K. Body Rhythms/Biological Clocks. In Encyclopedia of Mental Health, 2nd ed.; Friedman, H.S., Ed.; University of California: Berkeley, CA, USA, 2016; pp. 193–203. [Google Scholar]
- Draper, C.F.; Duisters, K.; Weger, B.; Chakrabarti, A.; Harms, A.C.; Brennan, L.; Hankemeier, T.; Goulet, L.; Konz, T.; Martin, F.P.; et al. Menstrual cycle rhythmicity: Metabolic patterns in healthy women. Sci. Rep. 2018, 8, 14568. [Google Scholar] [CrossRef] [Green Version]
- Baker, F.C.; Siboza, F.; Fuller, A. Temperature regulation in women: Effects of the menstrual cycle. Temperature 2020, 7, 226–262. [Google Scholar] [CrossRef]
- de Mouzon, J.; Testart, J.; Lefevre, B.; Pouly, J.L.; Frydman, R. Time relationships between basal body temperature and ovulation or plasma progestins. Fertil. Steril. 1984, 41, 254–259. [Google Scholar] [CrossRef]
- Barton, M.; Wiesner, B.P. Thermogenic effect of progesterone. Lancet 1945, 24, 671–672. [Google Scholar] [CrossRef] [PubMed]
- Ekhart, D.; Wicht, H.; Kersken, T.; Ackermann, H.; Kaczmarczyk, M.; Pretzsch, G.; Alexander, H.; Korf, H.W. Dynamics of core body temperature cycles in long-term measurements under real life conditions in women. Chronobiol. Int. 2018, 35, 8–23. [Google Scholar] [CrossRef]
- Refinetti, R. Circadian rhythmicity of body temperature and metabolism. Temperature 2020, 7, 321–362. [Google Scholar] [CrossRef]
- Chidi-Ogbolu, N.; Baar, K. Effect of estrogen on musculoskeletal performance and injury risk. Front. Physiol. 2019, 10, 1834. [Google Scholar] [CrossRef] [Green Version]
- Korf, H.W. Signaling pathways to and from the hypophysial pars tuberalis, an important center for the control of seasonal rhythms. Gen. Comp. Endocrinol. 2018, 258, 236–243. [Google Scholar] [CrossRef]
- Dopico, X.C.; Evangelou, M.; Ferreira, R.C.; Guo, H.; Pekalski, M.L.; Smyth, D.J.; Cooper, N.; Burren, O.S.; Fulford, A.J.; Hennig, B.J.; et al. Widespread seasonal gene expression reveals annual differences in human immunity and physiology. Nat. Commun. 2015, 6, 7000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valdebenito, J.O.; Halimubieke, N.; Lendvai, Á.Z.; Figuerola, J.; Eichhorn, G.; Székely, T. Seasonal variation in sex-specific immunity in wild birds. Sci. Rep. 2021, 11, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Pierre, K.; Schlesinger, N.; Androulakis, I.P. The role of the hypothalamic-pituitary-adrenal axis in modulating seasonal changes in immunity. Physiol. Genom. 2016, 48, 719–738. [Google Scholar] [CrossRef] [Green Version]
- Khandaker, G.M.; Pearson, R.M.; Zammit, S.; Lewis, G.; Jones, P.B. Association of serum interleukin 6 and C-reactive protein in childhood with depression and psychosis in young adult life: A population-based longitudinal study. JAMA Psychiatry 2014, 71, 1121–1128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wium-Andersen, M.K.; Ørsted, D.D.; Nielsen, S.F.; Nordestgaard, B.G. Elevated C-reactive protein levels, psychological distress, and depression in 73, 131 individuals. JAMA Psychiatry 2013, 70, 176–184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferreira, R.C.; Freitag, D.F.; Cutler, A.J.; Howson, J.M.; Rainbow, D.B.; Smyth, D.J.; Kaptoge, S.; Clarke, P.; Boreham, C.; Coulson, R.M.; et al. Functional IL6R 358Ala Allele Impairs Classical IL-6 Receptor Signaling and Influences Risk of Diverse Inflammatory Diseases. PLoS Genet. 2013, 9, e1003444. [Google Scholar] [CrossRef] [PubMed]
- Pell, J.P.; Cobbe, S.M. Seasonal variations in coronary heart disease. QJM 1999, 92, 689–696. [Google Scholar] [CrossRef]
- Tsao, C.W.; Aday, A.W.; Almarzooq, Z.I.; Alonso, A.; Beaton, A.Z.; Bittencourt, M.S.; Boehme, A.K.; Buxton, A.E.; Carson, A.P.; Commodore-Mensah, Y.; et al. Heart Disease and Stroke Statistics-2022 Update: A Report from the American Heart Association. Circulation 2022, 145, E153–E639. [Google Scholar] [CrossRef]
- Nevo-Shor, A.; Kogan, S.; Joshua, B.Z.; Bahat-Dinur, A.; Novack, V.; Fraenkel, M. Seasonal changes in serum calcium, PTH and vitamin D levels in patients with primary hyperparathyroidism. Bone 2016, 89, 59–63. [Google Scholar] [CrossRef]
- Fyfe, T.; Dunnigan, M.G.; Hamilton, E.; Rae, R.J. Seasonal variation in serum lipids, and incidence and mortality of ischaemic heart disease. J. Atheroscler. Res. 1968, 8, 591–596. [Google Scholar] [CrossRef]
- Fuller, J.H.; Grainger, S.L.; Jarrett, R.J.; Keen, H. Possible seasonal variation of plasma lipids in a healthy population. Clin. Chim. Acta 1974, 52, 305–310. [Google Scholar] [CrossRef] [PubMed]
- Woodhouse, P.R.; Khaw, K.T.; Plummer, M. Seasonal variation of serum lipids in an elderly population. Age Ageing 1993, 22, 273–278. [Google Scholar] [CrossRef] [PubMed]
- Manfredini, R.; Salmi, R.; Boari, B.; Manfredini, F. Seasonal Variation in Lipids: Should We Consider It More? Am. J. Cardiol. 2009, 104, 739–740. [Google Scholar] [CrossRef] [PubMed]
- Donahoo, W.T.; Jensen, D.R.; Shepard, T.Y.; Eckel, R.H. Seasonal variation in lipoprotein lipase and plasma lipids in physically active, normal weight humans. J. Clin. Endocrinol. Metab. 2000, 85, 3065–3068. [Google Scholar] [PubMed] [Green Version]
- Cambras, T.; Baena-Fustegueras, J.A.; Pardina, E.; Ricart-Jané, D.; Rossell, J.; Díez-Noguera, A.; Peinado-Onsurbe, J. Seasonal variation in plasma lipids and lipases in young healthy humans. Chronobiol. Int. 2017, 34, 1248–1258. [Google Scholar] [CrossRef]
- Ma, X.; Yan, H.; Zhang, H.; Wang, M.; Zhang, Q.; Zhou, X. Progress in the seasonal variations of blood lipids: A mini-review. Lipids Health Dis. 2020, 19, 108. [Google Scholar] [CrossRef]
- Bellastella, G.; Maiorino, M.I.; Scappaticcio, L.; De Bellis, A.; Mercadante, S.; Esposito, K.; Bellastella, A. Chronothyroidology: Chronobiological Aspects in Thyroid Function and Diseases. Life 2021, 11, 426. [Google Scholar] [CrossRef]
- Cargnelutti, F.; Pallotti, F.; Carlini, T.; Faja, F.; Vestri, A.R.; Fegatelli, D.A.; Lenzi, A.; Lombardo, F.; Paoli, D. A decade of WHO 2010: Total sperm number temporal trend and role of lifestyle factors. Asian J. Androl. 2023, 10-4103. [Google Scholar] [CrossRef]
- Thalange, N.K.; Gill, M.S.; Gill, L.; Whatmore, A.J.; Addison, G.M.; Price, D.A.; Clayton, P.E. Infradian rhythms in urinary growth hormone excretion. J. Clin. Endocrinol. Metab. 1996, 81, 100–106. [Google Scholar]
- Gerasch, A.; Burchardt, U.; Klagge, M.; Balschun, D. Age dependency of infradian rhythms in enzymuria of female volunteers. Eur. J. Clin. Chem. Clin. Biochem. 1997, 35, 281–286. [Google Scholar] [CrossRef] [Green Version]
- Burchardt, U.; Winkler, K.; Klagge, M.; Balschun, D.; Barth, A. Infradian biorhythms of enzymuria in man? J. Clin. Chem. Clin. Biochem. 1988, 26, 491–496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klingman, K.M.; Marsh, E.E.; Klerman, E.B.; Anderson, E.J.; Hall, J.E. Absence of circadian rhythms of gonadotropin secretion in women. J. Clin. Endocrinol. Metab. 2011, 96, 1456–1461. [Google Scholar] [CrossRef] [PubMed]
- Rossmanith, W.G. Ultradian and circadian patterns in luteinizing hormone secretion during reproductive life in women. Hum. Reprod. 1993, 8 (Suppl. S2), 77–83. [Google Scholar] [CrossRef] [PubMed]
- Mortola, J.F.; Laughlin, G.A.; Yen, S.S.C. A circadian rhythm of serum follicle-stimulating hormone in women. J. Clin. Endocrinol. Metab. 1992, 75, 861–864. [Google Scholar] [PubMed]
- Kanabrocki, E.L.; Sothern, R.B.; Ryan, M.D.; Kahn, S.; Augustine, G.; Johnson, C.; Foley, S.; Gathing, A.; Eastman, G.; Friedman, N.; et al. Circadian characteristics of serum calcium, magnesium and eight trace elements and of their metallo-moieties in urine of healthy middle-aged men. Clin. Ter. 2008, 159, 329–346. [Google Scholar]
- Micozkadioglu, H.; Ozelsancak, R.; Yildiz, I.; Erken, E.; Zumrutdal, A.; Torun, D.; Haberal, M. Circadian rhythm of serum phosphate, calcium and parathyroid hormone levels in hemodialysis patients. Clin. Lab. 2013, 59, 79–84. [Google Scholar] [CrossRef]
- Choi, J.H.; Lee, B.; Lee, J.Y.; Kim, C.H.; Park, B.; Kim, D.Y.; Kim, H.J.; Park, D.Y. Relationship between Sleep Duration, Sun Exposure, and Serum 25-Hydroxyvitamin D Status: A Cross-sectional Study. Sci. Rep. 2020, 10, 4168. [Google Scholar] [CrossRef] [Green Version]
- Masood, T.; Kushwaha, R.S.; Singh, R.; Sailwal, S.; Pandey, H.; Varma, A.; Singh, R.K.; Cornelissen, G. Circadian rhythm of serum 25 (OH) vitamin D, calcium and phosphorus levels in the treatment and management of type-2 diabetic patients. Drug Discov. Ther. 2015, 9, 70–74. [Google Scholar] [CrossRef] [Green Version]
- Zhu, B.; Dacso, C.C.; O’Malley, B.W. Unveiling “Musica Universalis” of the Cell: A Brief History of Biological 12-Hour Rhythms. J. Endocr. Soc. 2018, 2, 727–752. [Google Scholar] [CrossRef]
- James, S.M.; Honn, K.A.; Gaddameedhi, S.; Van Dongen, H.P.A. Shift Work: Disrupted Circadian Rhythms and Sleep—Implications for Health and Well-Being. Curr. Sleep Med. Rep. 2017, 3, 104–112. [Google Scholar] [CrossRef]
- Vosko, A.M.; Colwell, C.S.; Avidan, A.Y. Jet lag syndrome: Circadian organization, pathophysiology, and management strategies. Nat. Sci. Sleep 2010, 2, 187–198. [Google Scholar] [PubMed] [Green Version]
- Caliandro, R.; Streng, A.A.; van Kerkhof, L.W.M.; van der Horst, G.T.J.; Chaves, I. Social Jetlag and Related Risks for Human Health: A Timely Review. Nutrients 2021, 13, 4543. [Google Scholar] [CrossRef]
- Hebl, J.T.; Velasco, J.; McHill, A.W. Work Around the Clock: How Work Hours Induce Social Jetlag and Sleep Deficiency. Clin. Chest Med. 2022, 43, 249–259. [Google Scholar] [CrossRef]
- Blume, C.; Garbazza, C.; Spitschan, M. Effects of light on human circadian rhythms, sleep and mood. Somnologie 2019, 23, 147. [Google Scholar] [CrossRef] [Green Version]
- Maury, E. Off the Clock: From Circadian Disruption to Metabolic Disease. Int. J. Mol. Sci. 2019, 20, 1597. [Google Scholar] [CrossRef] [Green Version]
- Pan, A.; Schernhammer, E.S.; Sun, Q.; Hu, F.B. Rotating Night Shift Work and Risk of Type 2 Diabetes: Two Prospective Cohort Studies in Women. PLoS Med. 2011, 8, e1001141. [Google Scholar] [CrossRef] [Green Version]
- Haupt, C.M.; Alte, D.; Dörr, M.; Robinson, D.M.; Felix, S.B.; John, U.; Völzke, H. The relation of exposure to shift work with atherosclerosis and myocardial infarction in a general population. Atherosclerosis 2008, 201, 205–211. [Google Scholar] [CrossRef]
- Xiang, K.; Xu, Z.; Hu, Y.Q.; He, Y.S.; Wu, G.C.; Li, T.Y.; Wang, X.R.; Ding, L.H.; Zhang, Q.; Tao, S.S.; et al. Circadian clock genes as promising therapeutic targets for autoimmune diseases. Autoimmun. Rev. 2021, 20, 102866. [Google Scholar] [CrossRef]
- Li, Y.; Ma, J.; Yao, K.; Su, W.; Tan, B.; Wu, X.; Huang, X.; Li, T.; Yin, Y.; Tosini, G.; et al. Circadian rhythms and obesity: Timekeeping governs lipid metabolism. J. Pineal Res. 2020, 69, e12682. [Google Scholar] [CrossRef]
- Shafi, A.A.; Knudsen, K.E. Cancer and the Circadian Clock. Cancer Res. 2019, 79, 3806–3814. [Google Scholar] [CrossRef] [Green Version]
- Kamdar, B.B.; Needham, D.M.; Collop, N.A. Sleep Deprivation in Critical Illness: Its Role in Physical and Psychological Recovery. J. Intensive Care Med. 2012, 27, 97–111. [Google Scholar] [CrossRef] [PubMed]
- Mingrone, G.; Granato, L.; Valera-Mora, E.; Iaconelli, A.; Calvani, M.F.; Bracaglia, R.; Manco, M.; Nanni, G.; Castagneto, M. Ultradian ghrelin pulsatility is disrupted in morbidly obese subjects after weight loss induced by malabsorptive bariatric surgery. Am. J. Clin. Nutr. 2006, 83, 1017–1024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarabdjitsingh, R.A.; Spiga, F.; Oitzl, M.S.; Kershaw, Y.; Meijer, O.C.; Lightman, S.L.; De Kloet, E.R. Recovery from Disrupted Ultradian Glucocorticoid Rhythmicity Reveals a Dissociation Between Hormonal and Behavioural Stress Responsiveness. J. Neuroendocr. 2010, 22, 862–871. [Google Scholar]
- Lieberman, J.L.; De Souza, M.J.; Wagstaff, D.A.; Williams, N.I. Menstrual Disruption with Exercise is not Linked to an Energy Availability Threshold. Med. Sci. Sport. Exerc. 2018, 50, 551–561. [Google Scholar] [CrossRef] [PubMed]
- Monje, F.J.; Cicvaric, A.; Acevedo Aguilar, J.P.; Elbau, I.; Horvath, O.; Diao, W.; Glat, M.; Pollak, D.D. Disrupted ultradian activity rhythms and differential expression of several clock genes in interleukin-6-deficient mice. Front. Neurol. 2017, 8, 99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fraser, C.G. Biological Variation: From Principles to Practice; Amer. Assoc. for Clinical Chemistry: Washington, DC, USA, 2001. [Google Scholar]
- Fraser, C.G. Age-Related Changes in Laboratory Test Results. Drugs Aging 1993, 3, 246–257. [Google Scholar] [CrossRef] [PubMed]
- EFLM Biological Variation. Available online: https://biologicalvariation.eu/ (accessed on 24 February 2022).
- Coskun, A.; Sandberg, S.; Unsal, I.; Yavuz, F.G.; Cavusoglu, C.; Serteser, M.; Kilercik, M.; Aarsand, A.K. Personalized reference intervals–statistical approaches and considerations. Clin. Chem. Lab. Med. 2021, 60, 629–635. [Google Scholar] [CrossRef]
- Coskun, A.; Sandberg, S.; Unsal, I.; Cavusoglu, C.; Serteser, M.; Kilercik, M.; Aarsand, A.K. Personalized reference intervals: Using estimates of within-subject or within-person biological variation requires different statistical approaches. Clin. Chim. Acta 2022, 524, 201–202. [Google Scholar] [CrossRef]
- Coskun, A.; Sandberg, S.; Unsal, I.; Serteser, M.; Aarsand, A.K. Personalized reference intervals: From theory to practice. Crit. Rev. Clin. Lab. Sci. 2022, 59, 501–516. [Google Scholar] [CrossRef]
- Coşkun, A.; Sandberg, S.; Unsal, I.; Cavusoglu, C.; Serteser, M.; Kilercik, M.; Aarsand, A.K. Personalized Reference Intervals in Laboratory Medicine: A New Model Based on Within-Subject Biological Variation. Clin. Chem. 2021, 67, 374–384. [Google Scholar] [CrossRef]
- Carobene, A.; Graziani, M.S.; Cascio, C.L.; Tretti, L.; Cremonese, E.; Yabarek, T.; Gambaro, G.; Ceriotti, F. Age dependence of within-subject biological variation of nine common clinical chemistry analytes. Clin. Chem. Lab. Med. 2012, 50, 841–844. [Google Scholar] [CrossRef] [PubMed]
- Jones, G.R.D. Estimates of Within-Subject Biological Variation Derived from Pathology Databases: An Approach to Allow Assessment of the Effects of Age, Sex, Time between Sample Collections, and Analyte Concentration on Reference Change Values. Clin. Chem. 2019, 65, 579–588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ricós, C.; Iglesias, N.; García-Lario, J.V.; Simón, M.; Cava, F.; Hernández, A.; Perich, C.; Minchinela, J.; Alvarez, V.; Doménech, M.V.; et al. Within-subject biological variation in disease: Collated data and clinical consequences. Ann. Clin. Biochem. 2007, 44, 343–352. [Google Scholar] [CrossRef] [PubMed]
- Ricós, C.; Álvarez, V.; Perich, C.; Fernández-Calle, P.; Minchinela, J.; Cava, F.; Biosca, C.; Boned, B.; Doménech, M.; García-Lario, J.V.; et al. Rationale for using data on biological variation. Clin. Chem. Lab. Med. 2015, 53, 863–870. [Google Scholar] [CrossRef] [PubMed]
- Coşkun, A.; Carobene, A.; Kilercik, M.; Serteser, M.; Sandberg, S.; Aarsand, A.K.; Fernandez-Calle, P.; Jonker, N.; Bartlett, W.A.; Díaz-Garzón, J.; et al. Within-subject and between-subject biological variation estimates of 21 hematological parameters in 30 healthy subjects. Clin. Chem. Lab. Med. 2018, 56, 1309–1318. [Google Scholar] [CrossRef]
- Carobene, A. Reliability of biological variation data available in an online database: Need for improvement. Clin. Chem. Lab. Med. 2015, 53, 871–877. [Google Scholar] [CrossRef]
- Carobene, A.; Braga, F.; Roraas, T.; Sandberg, S.; Bartlett, W.A. A systematic review of data on biological variation for alanine aminotransferase, aspartate aminotransferase and γ-glutamyl transferase. Clin. Chem. Lab. Med. 2013, 51, 1997–2007. [Google Scholar] [CrossRef]
- Carobene, A.; Strollo, M.; Jonker, N.; Barla, G.; Bartlett, W.A.; Sandberg, S.; Sylte, M.S.; Røraas, T.; Sølvik, U.Ø.; Fernandez-Calle, P.; et al. Sample collections from healthy volunteers for biological variation estimates’ update: A new project undertaken by the Working Group on Biological Variation established by the European Federation of Clinical Chemistry and Laboratory Medicine. Clin. Chem. Lab. Med. 2016, 54, 1599–1608. [Google Scholar] [CrossRef]
- Carobene, A.; Aarsand, A.K.; Bartlett, W.A.; Coskun, A.; Diaz-Garzon, J.; Fernandez-Calle, P.; Guerra, E.; Jonker, N.; Locatelli, M.; Plebani, M.; et al. The European Biological Variation Study (EuBIVAS): A summary report. Clin. Chem. Lab. Med. 2022, 60, 505–517. [Google Scholar] [CrossRef]
- Aarsand, A.K.; Røraas, T.; Fernandez-Calle, P.; Ricos, C.; Díaz-Garzón, J.; Jonker, N.; Perich, C.; González-Lao, E.; Carobene, A.; Minchinela, J.; et al. The Biological Variation Data Critical Appraisal Checklist: A Standard for Evaluating Studies on Biological Variation. Clin. Chem. 2018, 64, 501–514. [Google Scholar] [CrossRef] [Green Version]
- Desirable Biological Variation Database Specifications-Westgard. Available online: https://www.westgard.com/biodatabase1.htm (accessed on 24 February 2022).
- Perich, C.; Minchinela, J.; Ricós, C.; Fernández-Calle, P.; Alvarez, V.; Doménech, M.V.; Simón, M.; Biosca, C.; Boned, B.; García-Lario, J.V.; et al. Biological variation database: Structure and criteria used for generation and update. Clin. Chem. Lab. Med. 2015, 53, 299–305. [Google Scholar] [CrossRef] [PubMed]
- Şahin, T.; Tozcu, D. Circadian rhythm and obesity. Int. J. Sci. Lett. 2022, 4, 202–219. [Google Scholar] [CrossRef]
- Parrinello, C.M.; Lutsey, P.L.; Couper, D.; Eckfeldt, J.H.; Steffes, M.W.; Coresh, J.; Selvin, E. Total Short-term Variability in Biomarkers of Hyperglycemia in Older Adults. Clin. Chem. 2015, 61, 1540–1541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ozarda, Y.; Sikaris, K.; Streichert, T.; Macri, J. Distinguishing reference intervals and clinical decision limits-A review by the IFCC Committee on Reference Intervals and Decision Limits. Crit. Rev. Clin. Lab. Sci. 2018, 55, 420–431. [Google Scholar] [CrossRef] [PubMed]
- Horowitz, G.L.; Altaie, S.; Boyd, J.C.; Ceriotti, F.; Garg, U.; Horn, P. EP28-A3c Defining, Establishing, and Verifying Reference Intervals in the Clinical Laboratory; Approved Guideline, 3rd ed.; Clinical And Laboratory Standards İnstitute: Wayne, PA, USA, 2008. [Google Scholar]
- Pelsers, M.M.; Chapelle, J.P.; Knapen, M.; Vermeer, C.; Muijtjens, A.M.; Hermens, W.T.; Glatz, J.F. Influence of age and sex and day-to-day and within-day biological variation on plasma concentrations of fatty acid-binding protein and myoglobin in healthy subjects. Clin. Chem. 1999, 45, 441–443. [Google Scholar] [PubMed]
- Hilderink, J.M.; Klinkenberg, L.J.; Aakre, K.M.; de Wit, N.C.; Henskens, Y.M.; van der Linden, N.; Bekers, O.; Rennenberg, R.J.; Koopmans, R.P.; Meex, S.J. Within-day biological variation and hour-to-hour reference change values for hematological parameters. Clin. Chem. Lab. Med. 2017, 55, 1013–1024. [Google Scholar] [CrossRef]
- Madsen, A.T.; Hojbjerg, J.A.; Sorensen, B.S.; Winther-Larsen, A. Day-to-day and within-day biological variation of cell-free DNA. EBioMedicine 2019, 49, 284–290. [Google Scholar] [CrossRef] [Green Version]
- Bailey, D.; Bevilacqua, V.; Colantonio, D.A.; Pasic, M.D.; Perumal, N.; Chan, M.K.; Adeli, K. Pediatric within-day biological variation and quality specifications for 38 biochemical markers in the CALIPER cohort. Clin. Chem. 2014, 60, 518–529. [Google Scholar] [CrossRef] [Green Version]
- Baskin, L.; Chin, A.; Abdullah, A.; Naugler, C. Errors in patient preparation, specimen collection, anticoagulant and preservative use: How to avoid such pre-analytical errors. In Accurate Results in the Clinical Laboratory, 2nd ed.; Dasgupta, A., Sepulveda, J.L., Eds.; Elsevier: Amsterdam, The Netherlands, 2019; pp. 11–26. [Google Scholar]
- Schüring, A.N.; Kelsch, R.; Pierściński, G.; Nofer, J.-R. Brief Communication Clinical Chemistry Establishing Reference Intervals for Sex Hormones on the Analytical Platforms Advia Centaur and Immulite 2000XP. Ann. Lab. Med. 2016, 36, 55–59. [Google Scholar]
- Verdonk, S.J.E.; Vesper, H.W.; Martens, F.; Sluss, P.M.; Hillebrand, J.J.; Heijboer, A.C. Estradiol reference intervals in women during the menstrual cycle, postmenopausal women and men using an LC-MS/MS method. Clin. Chim. Acta 2019, 495, 198–204. [Google Scholar] [CrossRef]
- Stricker, R.; Eberhart, R.; Chevailler, M.C.; Quinn, F.A.; Bischof, P.; Stricker, R. Establishment of detailed reference values for luteinizing hormone, follicle stimulating hormone, estradiol, and progesterone during different phases of the menstrual cycle on the Abbott ARCHITECT analyzer. Clin. Chem. Lab. Med. 2006, 44, 883–887. [Google Scholar] [CrossRef] [PubMed]
- Fraser, C.G. Reference change values: The way forward in monitoring. Ann. Clin. Biochem. 2009, 46, 264–265. [Google Scholar] [CrossRef] [PubMed]
- Dittadi, R.; Fabricio, A.S.C.; Gion, M. Biological variation and reference change value as decision criteria in clinical use of tumor biomarkers. Are they really useful? Clin. Chem. Lab. Med. 2022, 60, e136–e137. [Google Scholar] [CrossRef]
- Mazri, F.H.; Manaf, Z.A.; Shahar, S.; Ludin, A.F.M. The Association between Chronotype and Dietary Pattern among Adults: A Scoping Review. Int. J. Environ. Res. Public Health 2019, 17, 68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalmbach, D.A.; Schneider, L.D.; Cheung, J.; Bertrand, S.J.; Kariharan, T.; Pack, A.I.; Gehrman, P.R. Genetic Basis of Chronotype in Humans: Insights From Three Landmark GWAS. Sleep 2017, 40, zsw048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montaruli, A.; Castelli, L.; Mulè, A.; Scurati, R.; Esposito, F.; Galasso, L.; Roveda, E. Biological Rhythm and Chronotype: New Perspectives in Health. Biomolecules 2021, 11, 487. [Google Scholar] [CrossRef] [PubMed]
- Sempere-Rubio, N.; Aguas, M.; Faubel, R. Association between Chronotype, Physical Activity and Sedentary Behaviour: A Systematic Review. Int. J. Environ. Res. Public Health 2022, 19, 9646. [Google Scholar] [CrossRef] [PubMed]
- Chauhan, S.; Norbury, R.; Faßbender, K.C.; Ettinger, U.; Kumari, V. Beyond Sleep: A Multidimensional Model of Chronotype. Neurosci. Biobehav. Rev. 2023, 148, 105114. [Google Scholar] [CrossRef]
- Horne, J.A.; Östberg, O. Individual differences in human circadian rhythms. Biol. Psychol. 1977, 5, 179–190. [Google Scholar] [CrossRef] [PubMed]
- van der Merwe, C.; Münch, M.; Kruger, R. Chronotype Differences in Body Composition, Dietary Intake and Eating Behavior Outcomes: A Scoping Systematic Review. Adv. Nutr. 2022, 13, 2357–2405. [Google Scholar] [CrossRef]
- Baron, K.G.; Reid, K.J.; Kern, A.S.; Zee, P.C. Role of sleep timing in caloric intake and BMI. Obesity 2011, 19, 1374–1381. [Google Scholar] [CrossRef] [PubMed]
- Merikanto, I.; Lahti, T.; Puolijoki, H.; Vanhala, M.; Peltonen, M.; Laatikainen, T.; Vartiainen, E.; Salomaa, V.; Kronholm, E.; Partonen, T. Associations of chronotype and sleep with cardiovascular diseases and type 2 diabetes. Chronobiol. Int. 2013, 30, 470–477. [Google Scholar] [CrossRef]
- Merikanto, I.; Lahti, T.; Kronholm, E.; Peltonen, M.; Laatikainen, T.; Vartiainen, E.; Salomaa, V.; Partonen, T. Evening types are prone to depression. Chronobiol. Int. 2013, 30, 719–725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morera-Fumero, A.L. Chronotype as modulator of morning serum melatonin levels. Actas Españolas Psiquiatr. 2013, 41, 149–153. [Google Scholar]
- Adan, A.; Archer, S.N.; Hidalgo, M.P.; Milia LDi Natale, V.; Randler, C. Circadian Typology: A Comprehensive Review. Chronobiol. Int. 2012, 29, 1153–1175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vera, B.; Dashti, H.S.; Gómez-Abellán, P.; Hernández-Martínez, A.M.; Esteban, A.; Scheer, F.A.; Saxena, R.; Garaulet, M. Modifiable lifestyle behaviors, but not a genetic risk score, associate with metabolic syndrome in evening chronotypes. Sci. Rep. 2018, 8, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Lucassen, E.A.; Zhao, X.; Rother, K.I.; Mattingly, M.S.; Courville, A.B.; De Jonge, L.; Csako, G.; Cizza, G. Sleep Extension Study Group. Evening Chronotype Is Associated with Changes in Eating Behavior, More Sleep Apnea, and Increased Stress Hormones in Short Sleeping Obese Individuals. PLoS ONE 2013, 8, e56519. [Google Scholar] [CrossRef] [Green Version]
- Mormont, M.C.; Waterhouse, J. Contribution of the rest–activity circadian rhythm to quality of life in cancer patients. Chronobiol. Int. 2002, 19, 313–323. [Google Scholar] [CrossRef]
- Cohen, L.; Cole, S.W.; Sood, A.K.; Prinsloo, S.; Kirschbaum, C.; Arevalo, J.M.; Jennings, N.B.; Scott, S.; Vence, L.; Wei, Q.; et al. Depressive Symptoms and Cortisol Rhythmicity Predict Survival in Patients with Renal Cell Carcinoma: Role of Inflammatory Signaling. PLoS ONE 2012, 7, e42324. [Google Scholar] [CrossRef]
- Sephton, S.E.; Lush, E.; Dedert, E.A.; Floyd, A.R.; Rebholz, W.N.; Dhabhar, F.S.; Spiegel, D.; Salmon, P. Diurnal cortisol rhythm as a predictor of lung cancer survival. Brain Behav. Immun. 2013, 30, S163–S170. [Google Scholar] [CrossRef] [PubMed]
- Lévi, F.; Dugué, P.A.; Innominato, P.; Karaboué, A.; Dispersyn, G.; Parganiha, A.; Giacchetti, S.; Moreau, T.; Focan, C.; Waterhouse, J.; et al. Wrist actimetry circadian rhythm as a robust predictor of colorectal cancer patients survival. Chronobiol. Int. 2014, 31, 891–900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sephton, S.E.; Sapolsky, R.M.; Kraemer, H.C.; Spiegel, D. Diurnal cortisol rhythm as a predictor of breast cancer survival. J. Natl. Cancer Inst. 2000, 92, 994–1000. [Google Scholar] [CrossRef] [PubMed]
- Galasso, L.; Montaruli, A.; Mulè, A.; Castelli, L.; Bruno, E.; Pasanisi, P.; Caumo, A.; Esposito, F.; Roveda, E. Rest-activity rhythm in breast cancer survivors: An update based on non-parametric indices. Chronobiol. Int. 2020, 37, 946–951. [Google Scholar] [CrossRef] [PubMed]
- Allison, K.C.; Goel, N.; Ahima, R.S. Delayed Timing of Eating: Impact on Weight and Metabolism. Curr. Obes. Rep. 2014, 3, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Spiegel, K.; Tasali, E.; Leproult, R.; Van Cauter, E. Effects of poor and short sleep on glucose metabolism and obesity risk. Nat. Rev. Endocrinol. 2009, 5, 253–561. [Google Scholar] [CrossRef] [PubMed]
- Spiegel, K.; Tasali, E.; Penev, P.; Van Cauter, E. Brief communication: Sleep curtailment in healthy young men is associated with decreased leptin levels, elevated ghrelin levels, and increased hunger and appetite. Ann. Intern. Med. 2004, 141, 846–850. [Google Scholar] [CrossRef] [PubMed]
- Taheri, S.; Lin, L.; Austin, D.; Young, T.; Mignot, E. Short sleep duration is associated with reduced leptin, elevated ghrelin, and increased body mass index. PLoS Med. 2004, 1, 210–217. [Google Scholar] [CrossRef] [PubMed]
- Amiama-Roig, A.; Verdugo-Sivianes, E.M.; Carnero, A.; Blanco, J.R. Chronotherapy: Circadian Rhythms and Their Influence in Cancer Therapy. Cancers 2022, 14, 5071. [Google Scholar] [CrossRef] [PubMed]
- Greco, C.M.; Sassone-Corsi, P. Personalized medicine and circadian rhythms: Opportunities for modern society. J. Exp. Med. 2020, 217, e20200702. [Google Scholar] [CrossRef]
- Chang, A.M.; Aeschbach, D.; Duffy, J.F.; Czeisler, C.A. Evening use of light-emitting eReaders negatively affects sleep, circadian timing, and next-morning alertness. Proc. Natl. Acad. Sci. USA 2015, 112, 1232–1237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koike, N.; Yoo, S.H.; Huang, H.C.; Kumar, V.; Lee, C.; Kim, T.K.; Takahashi, J.S. Transcriptional architecture and chromatin landscape of the core circadian clock in mammals. Science 2012, 338, 349–354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mure, L.S.; Le, H.D.; Benegiamo, G.; Chang, M.W.; Rios, L.; Jillani, N.; Ngotho, M.; Kariuki, T.; Dkhissi-Benyahya, O.; Cooper, H.M.; et al. Diurnal transcriptome atlas of a primate across major neural and peripheral tissues. Science 2018, 359, eaao0318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rana, S.; Prabhu, S.D.; Young, M.E. Chronobiological Influence Over Cardiovascular Function: The Good, the Bad, and the Ugly. Circ. Res. 2020, 126, 258–279. [Google Scholar] [CrossRef] [PubMed]
- Ezagouri, S.; Zwighaft, Z.; Sobel, J.; Baillieul, S.; Doutreleau, S.; Ladeuix, B.; Golik, M.; Verges, S.; Asher, G. Physiological and Molecular Dissection of Daily Variance in Exercise Capacity. Cell Metab. 2019, 30, 78–91. [Google Scholar] [CrossRef] [PubMed]
- Asher, G.; Sassone-Corsi, P. Time for food: The intimate interplay between nutrition, metabolism, and the circadian clock. Cell 2015, 161, 84–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guan, D.; Xiong, Y.; Borck, P.C.; Jang, C.; Doulias, P.T.; Papazyan, R.; Fang, B.; Jiang, C.; Zhang, Y.; Briggs, E.R.; et al. Diet-Induced Circadian Enhancer Remodeling Synchronizes Opposing Hepatic Lipid Metabolic Processes. Cell 2018, 174, 831–842. [Google Scholar] [CrossRef] [Green Version]
- Qin, X.; Li, Q.; Chen, W.; Bai, Y.; Baban, B.; Mao, J. The circadian expression of osteogenic factors in periodontal tissue loading mechanical force: New concepts of the personalized orthodontic care. EPMA J. 2019, 10, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Golubnitschaja, O. Feeling cold and other underestimated symptoms in breast cancer: Anecdotes or individual profiles for advanced patient stratification? EPMA J. 2017, 8, 17–22. [Google Scholar] [CrossRef] [Green Version]
- Masri, S.; Sassone-Corsi, P. The emerging link between cancer, metabolism, and circadian rhythms. Nat. Med. 2018, 24, 1795–1803. [Google Scholar] [CrossRef]
- Acker, J.; Golubnitschaja, O.; Büttner-Teleaga, A.; Richter, K. Wrist actigraphic approach in primary, secondary and tertiary care based on the principles of predictive, preventive and personalised (3P) medicine. EPMA J. 2021, 12, 349–363. [Google Scholar] [CrossRef] [PubMed]
- Pearson, J.A.; Wong, F.S.; Wen, L. Crosstalk between circadian rhythms and the microbiota. Immunology 2020, 161, 278–290. [Google Scholar] [CrossRef] [PubMed]
- Leng, Y.; Musiek, E.S.; Hu, K.; Cappuccio, F.P.; Yaffe, K. Association between circadian rhythms and neurodegenerative diseases. Lancet Neurol. 2019, 18, 307–318. [Google Scholar] [CrossRef] [PubMed]
- Portaluppi, F.; Tiseo, R.; Smolensky, M.H.; Hermida, R.C.; Ayala, D.E.; Fabbian, F. Circadian rhythms and cardiovascular health. Sleep Med. Rev. 2012, 16, 151–166. [Google Scholar] [CrossRef] [PubMed]
- Griffis, C.G.; Mistry, J.; Islam, K.; Cutler, T.; Colwell, C.S.; Garfinkel, A. Circadian and ultradian rhythms in normal mice and in a mouse model of Huntington’s disease. Chronobiol. Int. 2022, 39, 513–524. [Google Scholar] [CrossRef] [PubMed]
- Hartsock, M.J.; Strnad, H.K.; Spencer, R.L. Iterative Metaplasticity Across Timescales: How Circadian, Ultradian, and Infradian Rhythms Modulate Memory Mechanisms. J. Biol. Rhythm. 2022, 37, 29–42. [Google Scholar] [CrossRef] [PubMed]
- Olovnikov, A. Lunasensor, infradian rhythms, telomeres, and the chronomere program of aging. Ann. N. Y. Acad. Sci. 2005, 1057, 112–132. [Google Scholar] [CrossRef] [PubMed]
- Turbill, C.; Ruf, T.; Smith, S.; Bieber, C. Seasonal variation in telomere length of a hibernating rodent. Biol. Lett. 2013, 9, 20121095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rehkopf, D.H.; Dow, W.H.; Rosero-Bixby, L.; Lin, J.; Epel, E.S.; Blackburn, E.H. Seasonal variation of peripheral blood leukocyte telomere length in Costa Rica: A population-based observational study. Am. J. Hum. Biol. 2014, 26, 367–375. [Google Scholar] [CrossRef] [Green Version]
- Beaulieu, M.; Benoit, L.; Abaga, S.; Kappeler, P.M.; Charpentier, M.J.E. Mind the cell: Seasonal variation in telomere length mirrors changes in leucocyte profile. Mol. Ecol. 2017, 26, 5603–5613. [Google Scholar] [CrossRef]
- Ledda, C.; Loreto, C.; Rapisarda, V. Telomere Length as a Biomarker of Biological Aging in Shift Workers. Appl. Sci. 2020, 10, 2764. [Google Scholar] [CrossRef]
- Daios, S.; Anogeianaki, A.; Kaiafa, G.; Kontana, A.; Veneti, S.; Gogou, C.; Karlafti, E.; Pilalas, D.; Kanellos, I.; Savopoulos, C. Telomere Length as a Marker of Biological Aging: A Critical Review of Recent Literature. Curr. Med. Chem. 2022, 29, 5478–5495. [Google Scholar] [PubMed]
- Bittman, E.L. Timing in the Testis. J. Biol. Rhythm. 2016, 31, 12–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mao, H.; Feng, L.; Yang, W.X. Environmental factors contributed to circannual rhythm of semen quality. Chronobiol. Int. 2017, 34, 411–425. [Google Scholar] [CrossRef] [PubMed]
- Xue, T.; Song, C.; Wang, Q.; Wang, Y.; Chen, G. Investigations of the CLOCK and BMAL1 Proteins Binding to DNA: A Molecular Dynamics Simulation Study. PLoS ONE 2016, 11, e0155105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González-Arto, M.; Vicente-Carrillo, A.; Martínez-Pastor, F.; Fernández-Alegre, E.; Roca, J.; Miró, J.; Rigau, T.; Rodríguez-Gil, J.E.; Pérez-Pé, R.; Muiño-Blanco, T.; et al. Melatonin receptors MT1 and MT2 are expressed in spermatozoa from several seasonal and nonseasonal breeder species. Theriogenology 2016, 86, 1958–1968. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Coskun, A.; Zarepour, A.; Zarrabi, A. Physiological Rhythms and Biological Variation of Biomolecules: The Road to Personalized Laboratory Medicine. Int. J. Mol. Sci. 2023, 24, 6275. https://doi.org/10.3390/ijms24076275
Coskun A, Zarepour A, Zarrabi A. Physiological Rhythms and Biological Variation of Biomolecules: The Road to Personalized Laboratory Medicine. International Journal of Molecular Sciences. 2023; 24(7):6275. https://doi.org/10.3390/ijms24076275
Chicago/Turabian StyleCoskun, Abdurrahman, Atefeh Zarepour, and Ali Zarrabi. 2023. "Physiological Rhythms and Biological Variation of Biomolecules: The Road to Personalized Laboratory Medicine" International Journal of Molecular Sciences 24, no. 7: 6275. https://doi.org/10.3390/ijms24076275