Variability, Expression, and Methylation of IL-6 and IL-8 Genes in Bladder Cancer Pathophysiology


Abstract
:1. Introduction
2. Results
2.1. Characteristics of Study Participants
2.2. Single Nucleotide Polymorphisms of IL-6 and IL-8 as the Risk of BC Occurrence
2.3. Association between Combined Genotypes of IL-6 and IL-8 Polymorphisms and BC Risk—Gene–Gene Interaction
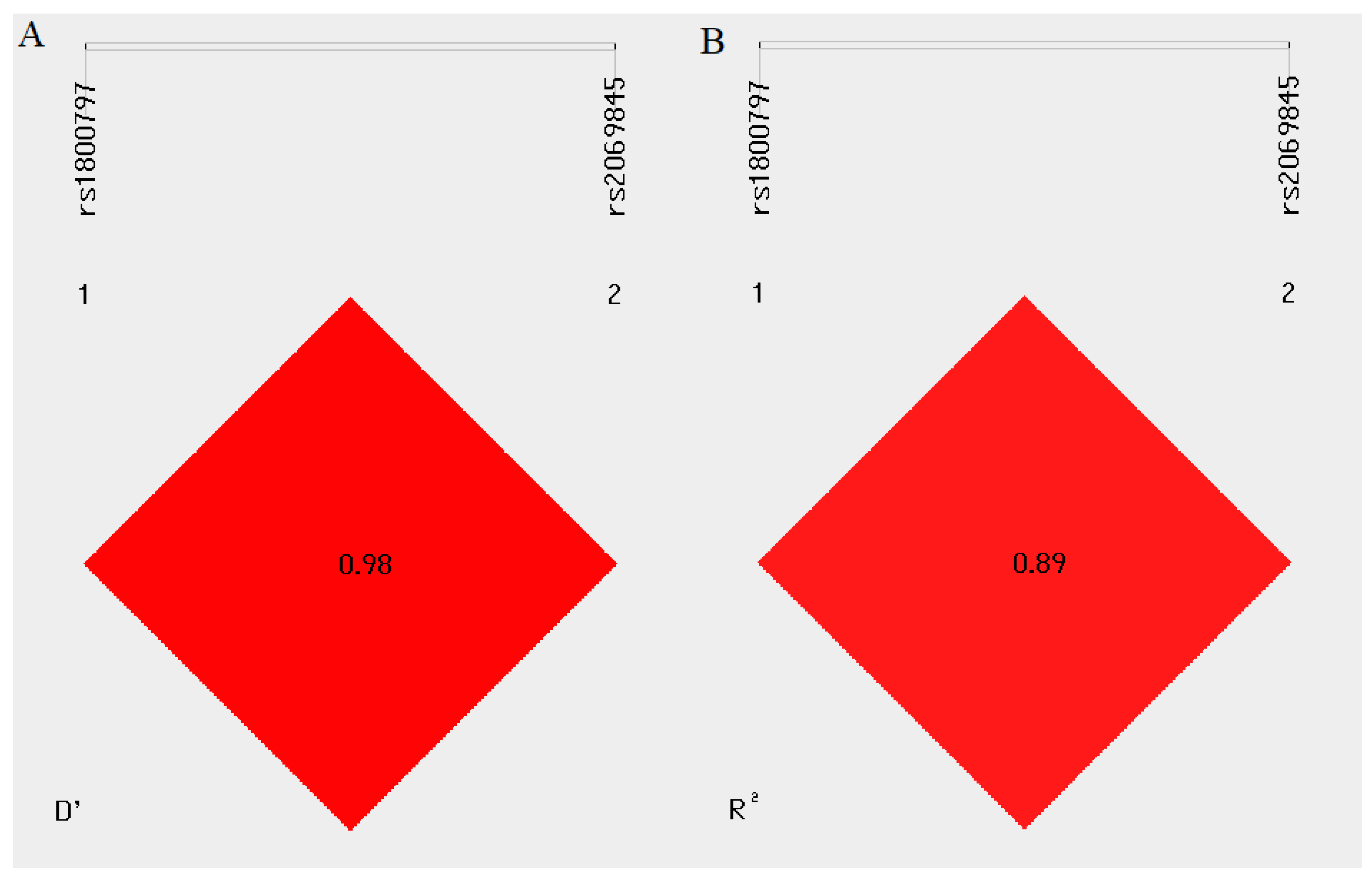
2.4. Linkage Disequilibrium and Haplotype Analysis
2.5. The Association between Studied Polymorphisms and the Clinical Histopathological Parameters of BC Patients
2.6. SNPs of IL-6 and IL-8 and BC Occurrence in the Male and Female Subpopulation
2.7. SNPs of Genes Encoding Interleukins and BC Occurrence in theNormal Body Weight/Overweight and Obesity Groups and in the Non-Smoker/Smoker Subpopulations
2.8. IL-6 and IL-8 mRNA Level Analysis
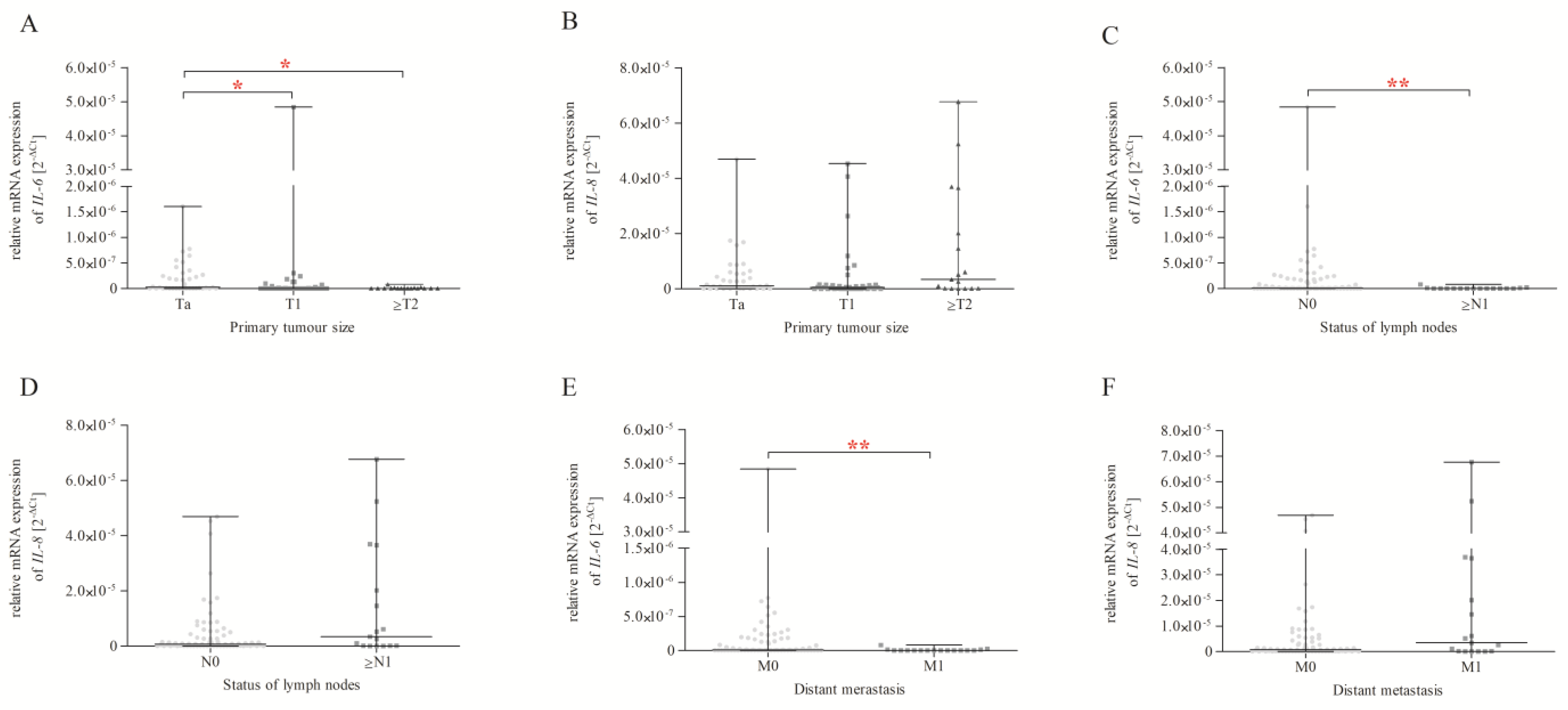
2.9. IL-6 and IL-8 Expression and Correlation with Size or the Direct Extent of the Primary Tumour, the Status of Lymph Node Metastasis, and Distant Metastasis according to the TNM Classification
2.10. IL-6 and IL-8 Expression and Correlation with the Grading of Histological Malignancy
2.11. IL-6 and IL-8 Expression in Genotype Subgroups
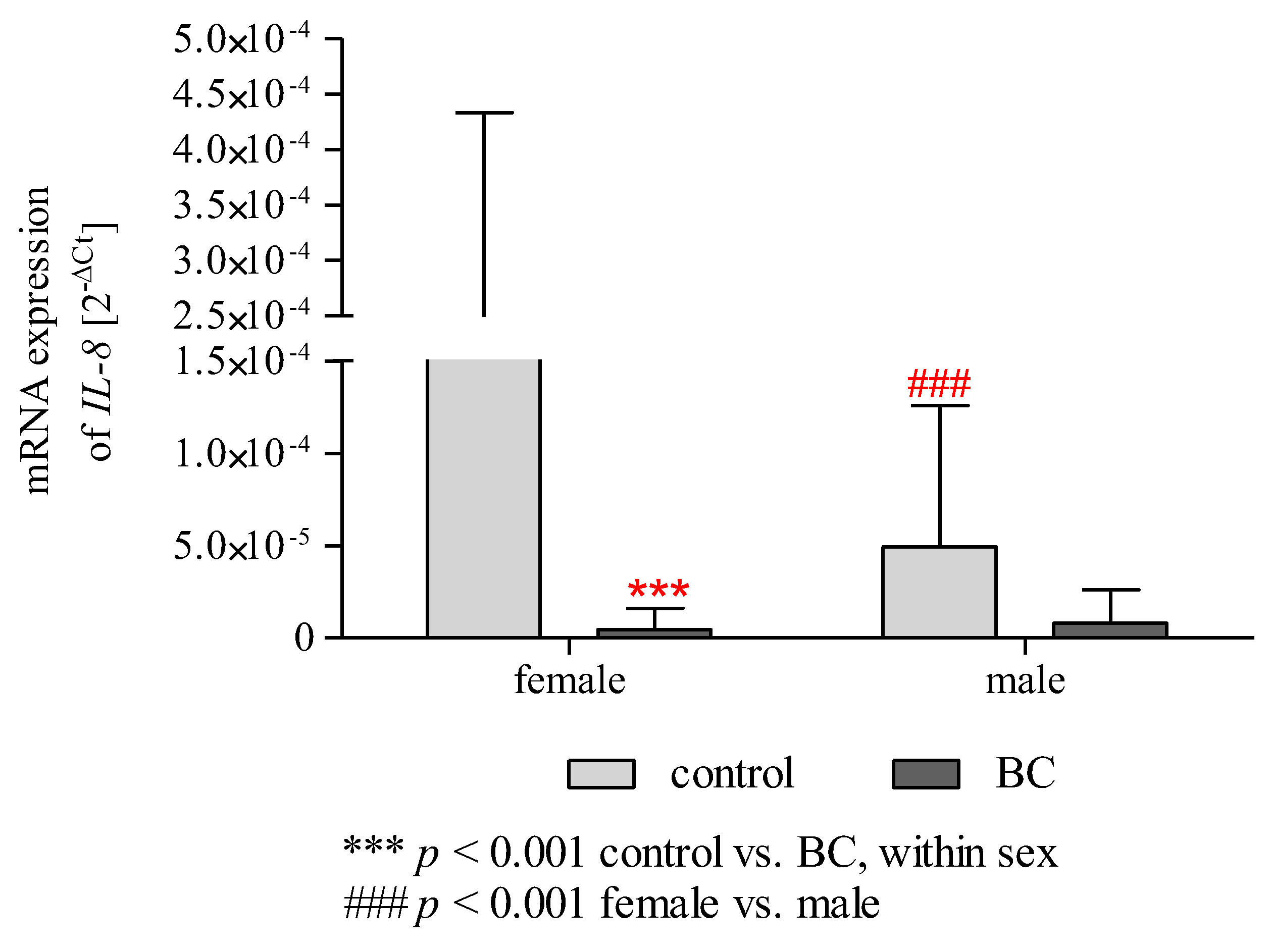
2.12. Effect of Gender/BMI/Cigarette Smoking and BC on the mRNA Expression of IL-6 and IL-8
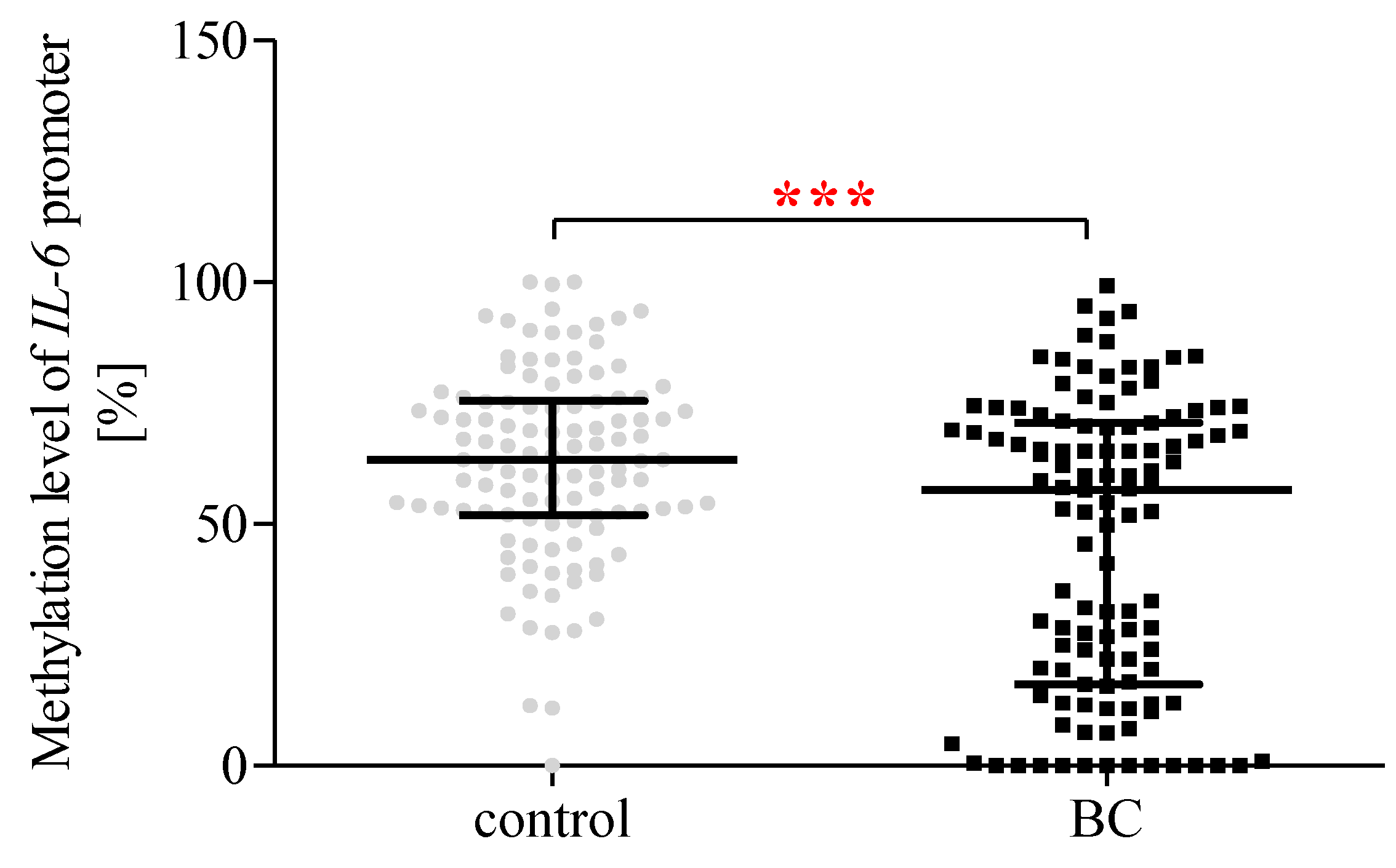
2.13. The Methylation Status of the IL-6 Promoter Region
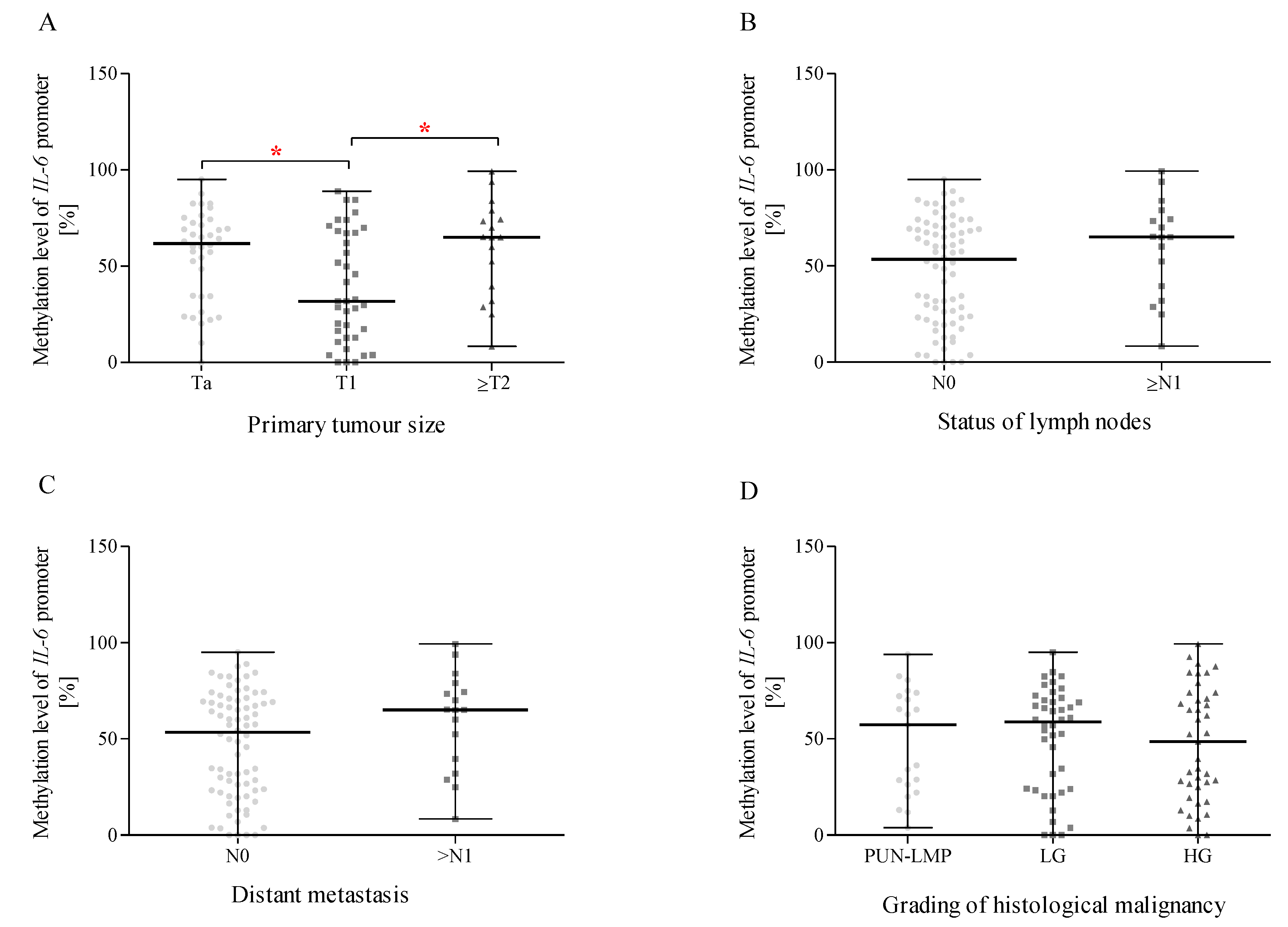
2.14. Methylation of the IL-6 Promoter Region and Correlation with Size or the Direct Extent of the Primary Tumour, the Status of Lymph Node Metastasis, Distant Metastasis According to the TNM Classification, and the Grading of Histological Malignancy
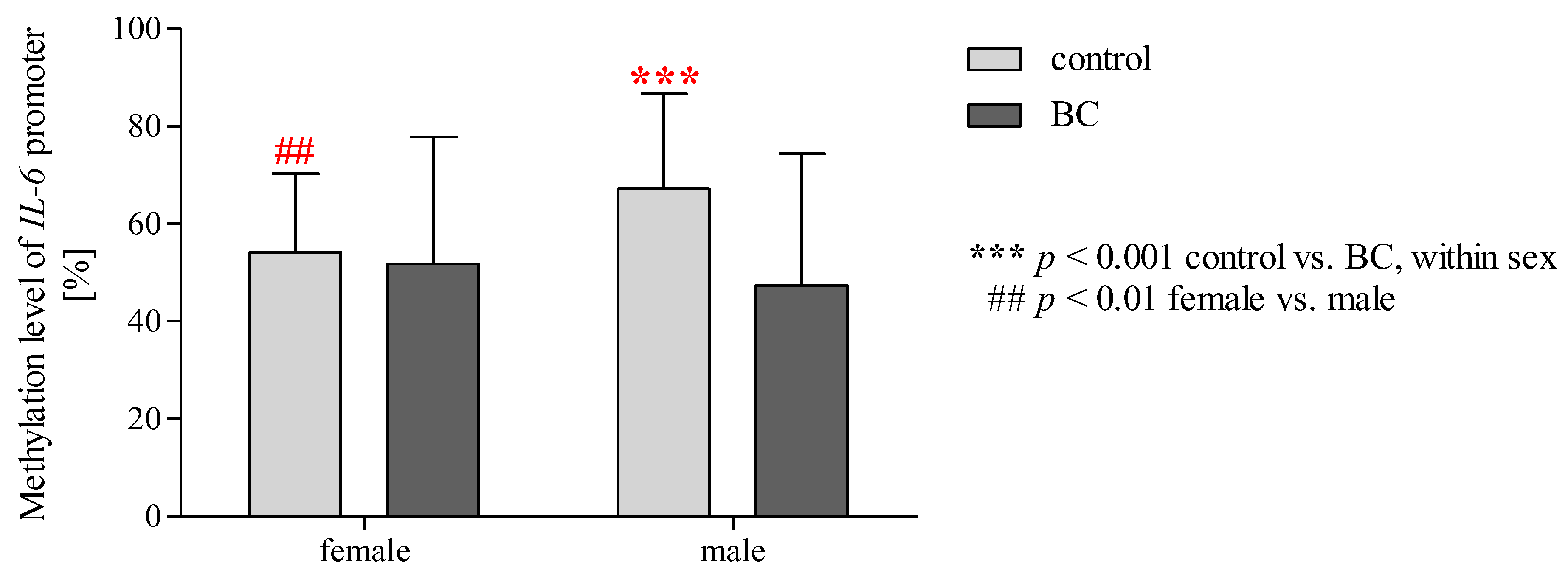
2.15. Effect of Gender/BMI/Cigarette Smoking and BC on the Methylation Level of the IL-6 Promoter Region
3. Discussion
4. Materials and Method
4.1. Participants
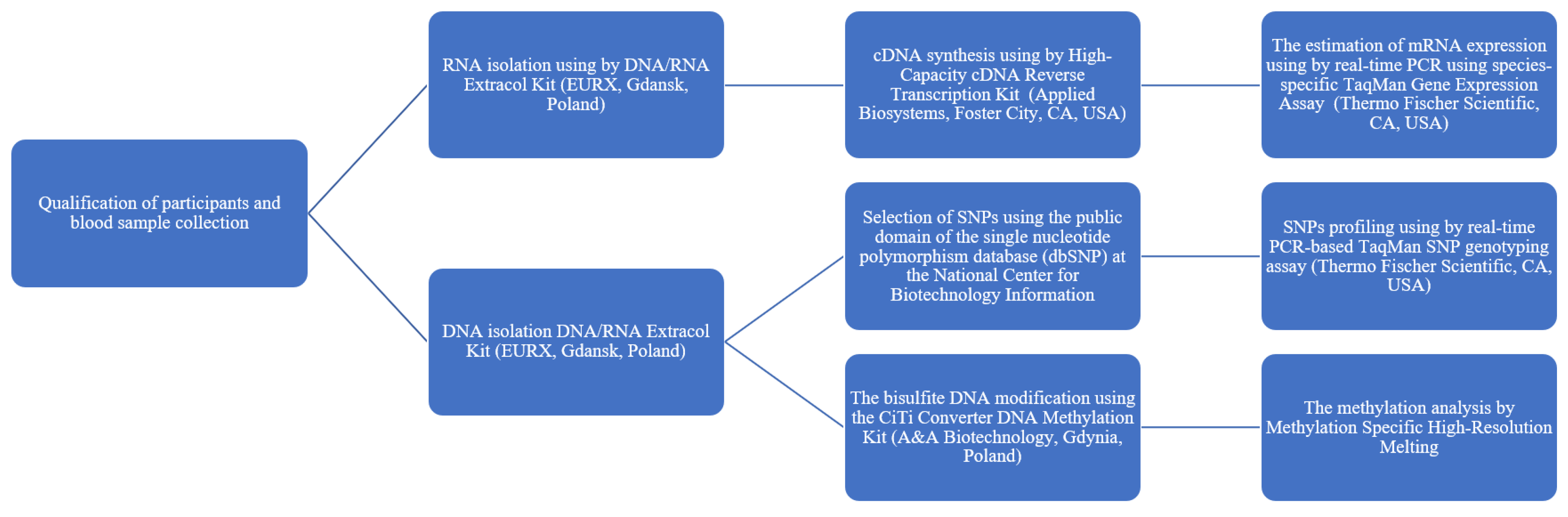
4.2. Blood Sample Collection and DNA and RNA Isolation
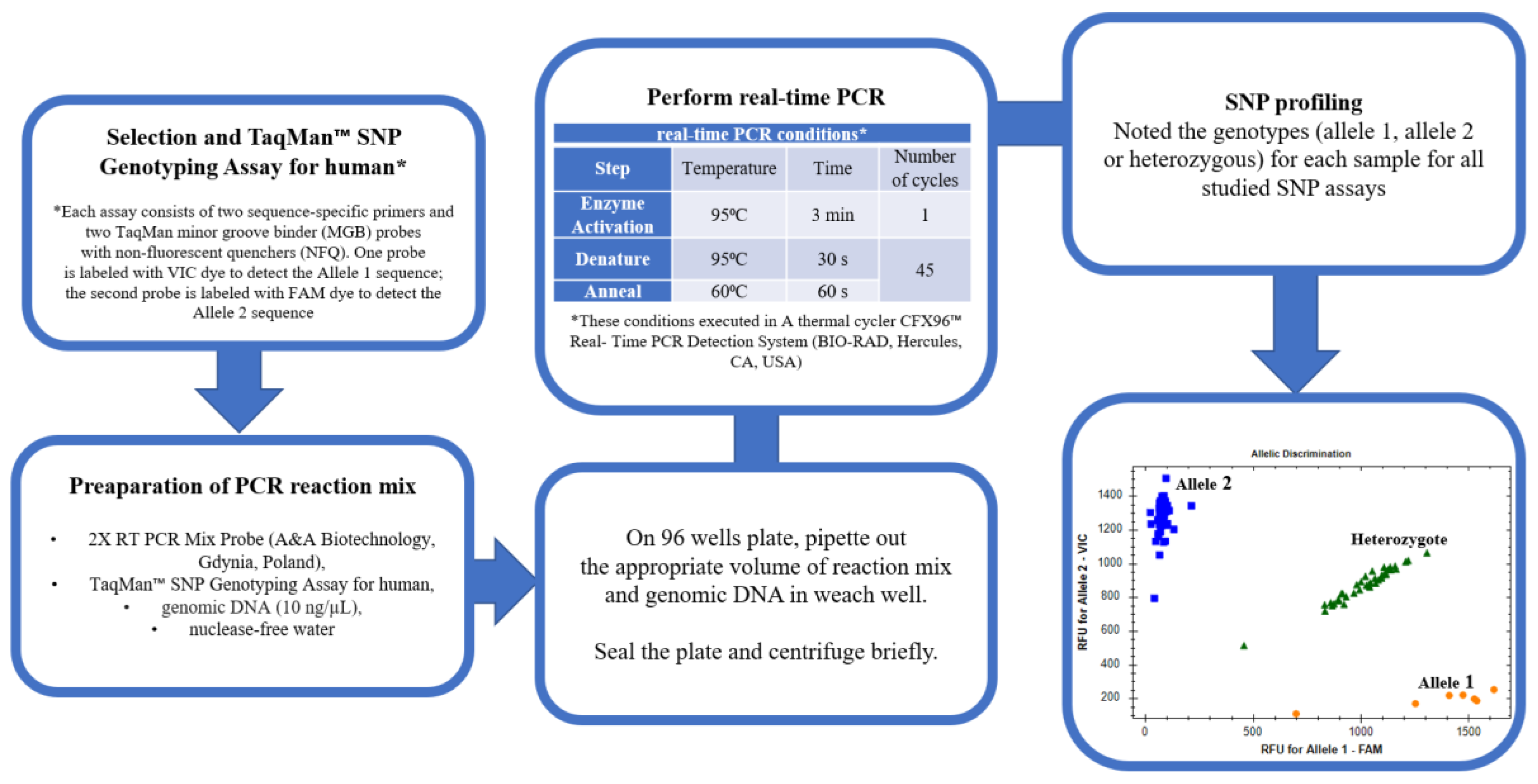
4.3. Selection of SNPs and Genotyping
4.4. cDNA Synthesis and mRNA Expression Levels
4.5. Bisulphite Treatment and Methylation Analysis by MS-HRM
4.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA A Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [Green Version]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA A Cancer J Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Mansour, B.; Monyók, Á.; Makra, N.; Gajdács, M.; Vadnay, I.; Ligeti, B.; Juhász, J.; Szabó, D.; Ostorházi, E. Bladder cancer-related microbiota: Examining differences in urine and tissue samples. Sci. Rep. 2020, 10, 11042. [Google Scholar] [CrossRef] [PubMed]
- Noon, A.P.; Albertsen, P.C.; Thomas, F.; Rosario, D.J.; Catto, J.W. Competing mortality in patients diagnosed with bladder cancer: Evidence of undertreatment in the elderly and female patients. Br. J. Cancer 2013, 108, 1534–1540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silverman, D.T.; Koutros, S.; Figueroa, J.D.; Prokunina-Olsson, L.; Rothman, N. Bladder cancer. Nat. Genet. 2008, 40, 1307–1312. [Google Scholar]
- Michaud, D.S.; Kogevinas, M.; Cantor, K.P.; Villanueva, C.M.; Garcia-Closas, M.; Rothman, N.; Malats, N.; Real, F.X.; Serra, C.; García-Closas, R.; et al. Total Fluid and Water Consumption and the Joint Effect of Exposure to Disinfection By-Products on Risk of Bladder Cancer. Environ. Health Perspect. 2007, 115, 1569–1572. [Google Scholar] [CrossRef] [Green Version]
- García-Pérez, J.; Pollán, M.; Boldo, E.; Pérez-Gómez, B.; Aragonés, N.; Lope, V.; Ramis, R.; Vidal, E.; López-Abente, G. Mortality due to lung, laryngeal and bladder cancer in towns lying in the vicinity of combustion installations. Sci. Total Environ. 2009, 407, 2593–2602. [Google Scholar] [CrossRef]
- Freedman, N.D.; Silverman, D.T.; Hollenbeck, A.R.; Schatzkin, A.; Abnet, C.C. Association between smoking and risk of bladder cancer among men and women. JAMA 2011, 306, 737–745. [Google Scholar] [CrossRef]
- Kuper, H.; Boffetta, P.; Adami, H.-O. Tobacco use and cancer causation: Association by tumour type. J. Intern. Med. 2002, 252, 206–224. [Google Scholar] [CrossRef] [Green Version]
- Brennan, P.; Bogillot, O.; Cordier, S.; Greiser, E.; Schill, W.; Vineis, P.; Lopez-Abente, G.; Tzonou, A.; Chang-Claude, J.; Bolm-Audorff, U.; et al. Cigarette smoking and bladder cancer in men: A pooled analysis of 11 case-control studies. Int. J. Cancer 2000, 86, 289–294. [Google Scholar] [CrossRef]
- Aben, K.K.; Witjes, J.A.; Schoenberg, M.P.; De Kaa, C.H.-V.; Kiemeney, L.A. Familial aggregation of urothelial cell carcinoma. Int. J. Cancer 2001, 98, 274–278. [Google Scholar] [CrossRef]
- Gu, J.; Wu, X. Genetic susceptibility to bladder cancer risk and outcome. Pers. Med. 2011, 8, 365–374. [Google Scholar] [CrossRef] [Green Version]
- Takeuchi, T.; Hattori-Kato, M.; Okuno, Y.; Zaitsu, M.; Azuma, T. Genome-Wide Association Study Adjusted for Occupational and Environmental Factors for Bladder Cancer Susceptibility. Genes 2022, 13, 448. [Google Scholar] [CrossRef] [PubMed]
- Thompson, D.B.; Siref, L.E.; Feloney, M.P.; Hauke, R.J.; Agrawal, D.K. Immunological basis in the pathogenesis and treatment of bladder cancer. Expert Rev. Clin. Immunol. 2015, 11, 265–279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joseph, M.; Enting, D. Immune Responses in Bladder Cancer-Role of Immune Cell Populations, Prognostic Factors and Therapeutic Implications. Front. Oncol. 2019, 9, 1270. [Google Scholar] [CrossRef] [Green Version]
- Milkovic, L.; Siems, W.; Siems, R.; Zarkovic, N. Oxidative Stress and Antioxidants in Carcinogenesis and Integrative Therapy of Cancer. Curr. Pharm. Des. 2014, 20, 6529–6542. [Google Scholar] [CrossRef]
- Mittal, M.; Siddiqui, M.R.; Tran, K.; Reddy, S.P.; Malik, A.B. Reactive oxygen species in inflammation and tissue injury. Antioxid. Redox Signal. 2014, 20, 1126–1167. [Google Scholar] [CrossRef] [Green Version]
- Chen, R.J.; Ho, Y.S.; Guo, H.R.; Wang, Y.J. Rapid activation of Stat3 and ERK1/2 by nicotine modulates cell proliferation in human bladder cancer cells. Toxicol. Sci. 2008, 104, 283–293. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.-L.; Cen, L.; Kohout, J.; Hutzen, B.; Chan, C.; Hsieh, F.-C.; Loy, A.; Huang, V.; Cheng, G.; Lin, J. Signal transducer and activator of transcription 3 activation is associated with bladder cancer cell growth and survival. Mol. Cancer 2008, 7, 78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Behzadi, P.; Sameer, A.S.; Nissar, S.; Banday, M.Z.; Gajdács, M.; García-Perdomo, H.A.; Akhtar, K.; Pinheiro, M.; Magnusson, P.; Sarshar, M.; et al. The Interleukin-1 (IL-1) Superfamily Cytokines and Their Single Nucleotide Polymorphisms (SNPs). J. Immunol. Res. 2022, 2022, 2054431. [Google Scholar] [CrossRef] [PubMed]
- Behzadi, P. The Role of Toll-Like Receptor (TLR) Polymorphisms in Urinary Bladder Cancer. In Genetic Polymorphism and Cancer Susceptibility; Sameer, A.S., Banday, M.Z., Nissar, S., Eds.; Springer: Singapore, 2021. [Google Scholar]
- Bhat, I.A.; Qasim, I.; Masoodi, K.Z.; Paul, S.A.; Bhat, B.A.; Rasool, R.; Aziz, S.A.; Shah, Z.A. Significant impact of IL-6 -174G/C but inverse relation with -634 C/G polymorphism in patients with non-small cell lung cancer in Kashmiri population. Immunol. Investig. 2015, 44, 349–360. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Huang, K.; Huang, Y.; Shiue, S.; Pu, S.; Su, T.; Lin, C.; Hsueh, M. Polymorphisms of TNF-α -308 G/A and IL-8 -251 T/A Genes Associated with Urothelial Carcinoma: A Case-Control Study. BioMed Res. Int. 2018, 8, 3148137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, Y.; Cao, H.; Huang, D.; Zheng, L.; Nie, Z.; Zhang, S. RNA-Binding Proteins in Bladder Cancer. Cancers 2023, 15, 1150. [Google Scholar] [CrossRef] [PubMed]
- Cortina-Borja, M.; Smith, A.D.; Combarros, O.; Lehmann, D.J. The synergy factor: A statistic to measure interactions in complex diseases. BMC Res. Notes 2009, 2, 105. [Google Scholar] [CrossRef] [Green Version]
- Brierley, J.D.; Gospodarowicz, M.K.; Wittekind, C. (Eds.) TNM Classification of Malignant Tumours; John Wiley & Sons: Hoboken, NJ, USA, 2017. [Google Scholar]
- Miyamoto, H.; Miller, J.S.; Fajardo, D.A.; Lee, T.K.; Netto, G.J.; Epstein, J.I. Non-invasive papillary urothelial neoplasms: The 2004 WHO/ISUP classification system. Pathol. Int. 2010, 60, 1–8. [Google Scholar] [CrossRef]
- Kluth, L.A.; Xylinas, E.; Crivelli, J.J.; Passoni, N.; Comploj, E.; Pycha, A.; Chrystal, J.; Sun, M.; Karakiewicz, P.I.; Gontero, P.; et al. Obesity is associated with worse outcomes in patients with T1 high grade urothe-lial carcinoma of the bladder. J. Urol. 2013, 190, 480–486. [Google Scholar] [CrossRef] [PubMed]
- Johnson, A.D.; Wang, D.; Sadee, W. Polymorphisms affecting gene regulation and mRNA processing: Broad implications for pharmacogenetics. Pharmacol. Ther. 2005, 106, 19–38. [Google Scholar] [CrossRef]
- Balkwill, F.; Mantovani, A. Inflammation and cancer: Back to Virchow? Lancet 2001, 357, 539–545. [Google Scholar] [CrossRef]
- Ames, B.N.; Gold, L.S.; Willett, W.C. The causes and prevention of cancer. Proc. Natl. Acad. Sci. USA 1995, 92, 5258–5265. [Google Scholar] [CrossRef] [Green Version]
- Zarogoulidis, P.; Tsakiridis, K.; Karapantzou, C.; Lampaki, S.; Kioumis, I.; Pitsiou, G.; Papaiwannou, A.; Hohenforst-Schmidt, W.; Huang, H.; Kesisis, G.; et al. Use of Proteins as Biomarkers and Their Role in Carcinogenesis. J. Cancer 2015, 6, 9–18. [Google Scholar] [CrossRef]
- Matsuda, T.; Hirano, T. IL-6. In Cytokine Reference; Oppenheim, J., Feldmann, M., Eds.; Academic Press: San Diego, CA, USA, 2000; Volume 1, p. 537563. [Google Scholar]
- Kidd, P. Th1/Th2 balance: The hypothesis, its limitations, and implications for health and disease. Altern. Med. Rev. 2003, 8, 223–246. [Google Scholar] [PubMed]
- Corre, I.; Pineau, D.; Hermouet, S. Interleukin-8: An Autocrine/Paracrine Growth Factor for Human Hematopoietic Progenitors Acting in Synergy with Colony Stimulating Factor-1 to Promote Monocyte-Macrophage Growth and Differentiation. Exp. Hematol. 1999, 27, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Li, A.; Dubey, S.; Varney, M.L.; Dave, B.J.; Singh, R.K. IL-8 Directly Enhanced Endothelial Cell Survival, Proliferation, and Matrix Metalloproteinases Production and Regulated Angiogenesis. J. Immunol. 2003, 170, 3369–3376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bréchard, S.; Bueb, J.-L.; Tschirhart, E.J. Interleukin-8 Primes Oxidative Burst in Neutrophil-like HL-60 through Changes in Cytosolic Calcium. Cell Calcium. 2005, 37, 531–540. [Google Scholar] [CrossRef]
- Lee, K.; Nelson, C.M. New insights into the regulation of epithelial-mesenchymal transition and tissue fibrosis. Int. Rev. Cell Mol. Biol. 2012, 294, 171–221. [Google Scholar]
- Hsu, P.C.; Chen, Y.H.; Cheng, C.F.; Kuom, C.Y.; Sytwu, H.K. Interleukin-6 and Interleukin-8 Regulate STAT3 Activation Migration/Invasion and EMT in Chrysophanol-Treated Oral Cancer Cell Lines. Life 2021, 11, 423. [Google Scholar] [CrossRef]
- Leibovici, D.; Grossman, H.B.; Dinney, C.P.; Millikan, R.E.; Lerner, S.; Wang, Y.; Gu, J.; Dong, Q.; Wu, X. Polymorphisms in inflammation genes and bladder cancer: From initiation to recurrence, progression, and survival. J. Clin. Oncol. 2005, 23, 5746–5756. [Google Scholar] [CrossRef]
- Ahirwar, D.; Kesarwani, P.; Manchanda, P.K.; Mandhani, A.; Mittal, R.D. Anti- and proinflammatory cytokine gene polymorphism and genetic predisposition: Association with smoking, tumor stage and grade, and bacillus Calmette-Guérin immunotherapy in bladder cancer. Cancer Genet. Cytogenet. 2008, 184, 1–8. [Google Scholar] [CrossRef]
- Ebadi, N.; Jahed, M.; Mivehchi, M.; Majidizadeh, T.; Asgary, M.; Hosseini, S.A. Interleukin-12 and interleukin-6 gene polymorphisms and risk of bladder cancer in the Iranian population. Asian Pac. J. Cancer Prev. 2014, 15, 7869–7873. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.C.; Huang, Y.K.; Chung, C.J.; Huang, C.Y.; Pu, Y.S.; Shiue, H.S.; Lai, L.A.; Lin, Y.C.; Su, C.T.; Hsueh, Y.M. Polymorphism of inflammatory genes and arsenic methylation capacity are associated with urothelial carcinoma. Toxicol. Appl. Pharmacol. 2013, 272, 30–36. [Google Scholar] [CrossRef]
- Bennermo, M.; Held, C.; Stemme, S.; Ericsson, C.G.; Silveira, A.; Green, F.; Tornvall, P. Genetic predisposition of the interleukin6 response to inflammation: Implications for a variety of major diseases? Clin. Chem. 2004, 50, 2136–2140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sales-Marques, C.; Cardoso, C.C.; Alvarado-Arnez, L.E.; Illaramendi, X.; Sales, A.M.; Hacker, M.A.; Barbosa, M.G.M.; Nery, J.A.D.C.; Pinheiro, R.O.; Sarno, E.N.; et al. Genetic polymorphisms of the IL6 and NOD2 genes are risk factors for inflammatory reactions in leprosy. PloS Negl. Trop. Dis. 2017, 11, e0005754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, M.F.; Lin, P.Y.; Wu, C.F.; Chen, W.C.; Wu, C.T. IL-6 expression regulates tumorigenicity and correlates with prognosis in bladder cancer. PLoS ONE 2013, 8, e61901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hatina, J.; Kripnerová, M.; Houfková, K.; Hajdůšková, M.; Navvabi, N.; Havlíčková, N. Recent Progress in Urothelial Bladder Carcinoma: Basic Biology, Molecular Characterization, Conventional and Innovative Therapies and Tumour–Stroma Interactions. In Interdisciplinary Cancer Research; Springer: Cham, Switzerland, 2022. [Google Scholar]
- Sehgal, P.B. Interleukin-6 at the host-tumor interface: STAT3 in biomolecular condensates in cancer cells. Cells 2022, 11, 1164. [Google Scholar] [CrossRef] [PubMed]
- Chotirmall, S.H.; Greene, C.M.; Oglesby, I.K.; Thomas, W.; O’Neill, S.J.; Harvey, B.J.; McElvaney, N.G. 17Beta-estradiol inhibits IL-8 in cystic fibrosis by up-regulating secretory leucoprotease inhibitor. Am. J. Respir. Crit. Care Med. 2010, 182, 62–72. [Google Scholar] [CrossRef]
- Goulet, C.R.; Champagne, A.; Bernard, G.; Vandal, D.; Chabaud, S.; Pouliot, F.; Bolduc, S. Cancer-associated fibroblasts induce epithelial–mesenchymal transition of bladder cancer cells through paracrine IL-6 signalling. BMC Cancer 2019, 19, 137. [Google Scholar] [CrossRef] [Green Version]
- Reis, S.T.; Leite, K.R.; Piovesan, L.F.; Pontes-Junior, J.; Viana, N.I.; Abe, D.K.; Crippa, A.; Moura, C.M.; Adonias, S.P.; Srougi, M.; et al. Increased expression of MMP-9 and IL-8 are correlated with poor prognosis of Bladder Cancer. BMC Urol. 2012, 12, 18. [Google Scholar] [CrossRef] [Green Version]
- Ou, Z.; Wang, Y.; Liu, L.; Li, L.; Yeh, S.; Qi, L.; Chang, C. Tumor microenvironment B cells increase bladder cancer metastasis via modulation of the IL-8/androgen receptor (AR)/MMPs signals. Oncotarget 2015, 6, 26065–26078. [Google Scholar] [CrossRef] [Green Version]
- Portela, A.; Esteller, M. Epigenetic modifications and human disease. Nat. Biotechnol. 2010, 28, 1057–1068. [Google Scholar] [CrossRef]
- Muhammad, S.B.; Hassan, F.; Bhowmik, K.K.; Millat, M.S.; Sarwar, M.S.; Aziz, M.A.; Barek, M.A.; Sarowar Uddin, M.; Ferdous, M.; Safiqul Islam, M. Detection of association of IL1β, IL4R, and IL6 gene polymorphisms with cervical cancer in the Bangladeshi women by tetra-primer ARMS-PCR method. Int. Immunopharmacol. 2021, 90, 107131. [Google Scholar] [CrossRef]
- Korobeinikova, E.; Ugenskiene, R.; Insodaite, R.; Rudzianskas, V.; Jaselske, E.; Poskiene, L.; Juozaityte, E. Association of angiogenesis and inflammation-related gene functional polymorphisms with early-stage breast cancer prognosis. Oncol. Lett. 2020, 19, 3687–3700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winchester, D.A.; Till, C.; Goodman, P.J.; Tangen, C.M.; Santella, R.M.; Johnson-Pais, T.L.; Leach, R.J.; Xu, J.; Zheng, S.L.; Thompson, I.M.; et al. Association between variants in genes involved in the immune response and prostate cancer risk in men randomized to the finasteride arm in the Prostate Cancer Prevention Trial. Prostate 2017, 77, 908–919. [Google Scholar] [CrossRef] [PubMed]
- Taheri, M.; Noroozi, R.; Rakhshan, A.; Ghanbari, M.; Omrani, M.D.; Ghafouri-Fard, S. IL-6 Genomic Variants and Risk of Prostate Cancer. Urol. J. 2019, 16, 463–468. [Google Scholar] [PubMed]
- Bondurant, K.L.; Lundgreen, A.; Herrick, J.S.; Kadlubar, S.; Wolff, R.K.; Slattery, M.L. Interleukin genes and associations with colon and rectal cancer risk and overall survival. Int. J. Cancer 2013, 132, 905–915. [Google Scholar] [CrossRef] [Green Version]
- Srivastava, K.; Srivastava, A.; Sharma, K.L.; Mittal, B. Candidate gene studies in gallbladder cancer: A systematic review and meta-analysis. Mutat. Res. 2011, 728, 67–79. [Google Scholar] [CrossRef] [Green Version]
- Silver, N.; Best, S.; Jiang, J.; Thein, S.L. Selection of housekeeping genes for gene expression studies in human reticulocytes using real-time PCR. BMC Mol. Biol. 2006, 7, 33. [Google Scholar] [CrossRef] [Green Version]
- Wojdacz, T.K.; Borgbo, T.; Hansen, L.L. Primer design versus PCR bias in methylation independent PCR amplifications. Epigenetics 2009, 4, 231–234. [Google Scholar] [CrossRef] [Green Version]
- Qin, Z.S.; Niu, T.; Liu, J.S. Partition-ligation-expectation-maximization algorithm for haplotype inference with single-nucleotide polymorphisms. Am. J. Hum. Genet. 2002, 71, 1242–1247. [Google Scholar] [CrossRef] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Demographic Characteristics of the Study Participants | |||||
|---|---|---|---|---|---|
| Feature | Controls (n = 114) Frequency | Patients with BC (n = 116) Frequency | p * | ||
| Gender | females | 0.34 | 0.28 | 0.278 | |
| males | 0.66 | 0.72 | |||
| Age | mean ± SD | 66.71 ± 11.76 | 69.67 ± 11.26 | 0.031 | |
| range | 28–91 | 20–92 | |||
| Education | primary (basic) education | 0.16 | 0.30 | 0.087 | |
| vocational education | 0.39 | 0.25 | |||
| high school education | 0.33 | 0.42 | |||
| university degree | 0.12 | 0.03 | |||
| Residency | village | 0.36 | 0.46 | 0.805 | |
| a city with a population under 50 thou. residents | 0.39 | 0.16 | |||
| a city with a population over 50 thou. residents | 0.25 | 0.39 | |||
| Marital status | single | 0.25 | 0.13 | 0.003 | |
| married | 0.67 | 0.70 | |||
| widow/widower | 0.08 | 0.17 | |||
| Professional activity | physical work | 0.28 | 0.20 | 0.031 | |
| mental work | 0.17 | 0.05 | |||
| unemployment | 0.01 | 0.04 | |||
| pension | 0.64 | 0.71 | |||
| Smoking | never | 0.56 | 0.31 | 0.367 | |
| former | 0.24 | 0.38 | |||
| current | 0.20 | 0.31 | |||
| BMI [kg/m2] | mean ± SD | 27.69 ± 3.51 | 27.12 ± 4.83 | 0.305 | |
| <25 | 0.31 | 0.36 | |||
| 25–30 | 0.42 | 0.37 | |||
| >30 | 0.27 | 0.27 | |||
| Daily fluid intake | <2 L/day | 0.52 | 0.50 | 0.791 | |
| >2 L/day | 0.48 | 0.50 | |||
| Daily coffee consumption (number of cups, one cup has a capacity of 200 mL) | 0 | 0.21 | 0.27 | 0.369 | |
| 1 | 0.44 | 0.41 | |||
| 2–3 | 0.32 | 0.30 | |||
| >4 | 0.03 | 0.02 | |||
| Total blood count of the study participants | |||||
| Feature/Parameters | Controls (n = 114) mean ± SD | Patients with BC (n = 116) mean ± SD | p | ||
| Red blood cells—RBC (×1012/L) | 4.49 ± 0.61 | 4.29 ± 0.71 | 0.039 | ||
| Haematocrit—HCT (%) | 40.91 ± 5.26 | 38.80 ± 6.58 | 0.014 | ||
| Haemoglobin—HGB (g/L) | 13.56 ± 2.44 | 12.91 ± 2.33 | 0.030 | ||
| Mean corpuscular volume—MCV (fL) | 91.25 ± 5.34 | 90.49 ±5.97 | 0.135 | ||
| Mean cell haemoglobin—MCH (pg/cell) | 30.20 ± 3.20 | 30.24 ± 2.74 | 0.290 | ||
| Mean corpuscular haemoglobin concentration—MCHC (g/L) | 33.27 ± 1.22 | 33.11 ±1.52 | 0.207 | ||
| Red cell distribution width—RDW (%) | 13.50 ± 1.49 | 13.90 ± 1.32 | 0.010 | ||
| Haemoglobin distribution width—HDW (g/L) | 2.53 ± 0.25 | 2.56 ± 0.40 | 0.467 | ||
| White blood cells—WBC (×109/L) | 9.23 ± 18.67 | 8.88 ± 5.07 | 0.032 | ||
| %HYPO | 2.34 ± 4.23 | 3.63 ± 7.10 | 0.157 | ||
| %MIKRO | 1.35 ± 2.76 | 1.32 ± 2.56 | 0.903 | ||
| %MAKRO | 1.44 ± 1.85 | 1.77 ± 4.05 | 0.978 | ||
| %HYPER | 0.73 ± 0.54 | 0.72 ± 0.85 | 0.091 | ||
| Blood platelets—PLT (×109/L) | 234.95 ± 70.80 | 259.25 ± 106.83 | 0.127 | ||
| Mean platelet volume—MPV (fL) | 8.68 ± 1.53 | 8.98 ±1.04 | 0.081 | ||
| Blood biochemical parameters | |||||
| Feature/Parameters | Controls (n = 114) mean ± SD | Patients with BC (n = 116) mean ± SD | p | ||
| Glucose (mmol/L) | 5.92 ± 2.16 | 6.64 ± 2.29 | <0.001 | ||
| Creatinine (µmol/L) | 106.94 ± 12.14 | 127.02 ± 156.58 | 0.005 | ||
| Sodium (mmol/L) | 139.93 ± 2.75 | 139.48 ± 4.25 | 0.391 | ||
| Potassium (mmol/L) | 5.57 ± 9.42 | 4.60 ± 0.56 | 0.043 | ||
| Coagulation panel | |||||
| Prothrombin time (s) | 12.68 ± 2.48 | 12.19 ± 1.20 | 0.285 | ||
| Prothrombin index (%) | 92.90 ± 11.89 | 95.30 ± 8.59 | 0.237 | ||
| International normalised ratio (INR) | 1.13 ± 0.23 | 1.09 ± 0.11 | 0.883 | ||
| Activated partial thromboplastin time (APTT, s) | 30.67 ± 3.91 | 30.00 ± 3.11 | 0.609 | ||
| Fibrinogen (mg/L) | 400.24 ± 145.26 | 426.59 ±178.41 | 0.362 | ||
| Dipstick urinalysis | |||||
| Feature/Parameters | Controls (n = 114) mean ± SD | Patients with BC (n = 116) mean ± SD | p | ||
| pH | 5.84 ± 0.88 | 5.89 ± 1.01 | 0.911 | ||
| Specific gravity | 1.02 ± 0.01 | 1.02 ± 0.01 | 0.842 | ||
| Feature/Parameters | Controls (n = 114) Frequency | Patients with BC (n = 116) Frequency | p | ||
| WBC | absence per high-power field | 0.68 | 0.45 | 0.077 | |
| single per high-power field | 0.29 | 0.25 | |||
| numerous per high-power field | 0.04 | 0.30 | |||
| Nitrite | negative | 0.88 | 0.88 | 0.658 | |
| positive | 0.12 | 0.12 | |||
| Glucose | negative | 0.98 | 0.95 | 0.262 | |
| positive | 0.02 | 0.05 | |||
| Protein | negative | 0.76 | 0.44 | <0.001 | |
| positive | 0.24 | 0.56 | |||
| Ketones | negative | 0.90 | 0.90 | 0.964 | |
| positive | 0.10 | 0.10 | |||
| Bilirubin | negative | 0.98 | 0.89 | 0.006 | |
| positive | 0.02 | 0.11 | |||
| Urobilinogen | normal level | 1.00 | 0.94 | 0.711 | |
| above normal | 0.00 | 0.06 | |||
| Colour | pale yellow | 0.05 | 0.11 | 0.218 | |
| straw/yellow | 0.82 | 0.63 | |||
| dark yellow | 0.02 | 0.02 | |||
| amber | 0.05 | 0.10 | |||
| brown | 0.01 | 0.06 | |||
| red | 0.04 | 0.08 | |||
| Clarity | clear | 0.74 | 0.48 | <0.001 | |
| slightly cloudy | 0.12 | 0.25 | |||
| cloudy | 0.06 | 0.07 | |||
| very cloudy | 0.08 | 0.20 | |||
| Urine microscopy | |||||
| Feature/Parameters | Controls (n = 114) Frequency | Patients with BC (n = 116) Frequency | p | ||
| RBC | 0–3/high power field | 0.78 | 0.33 | <0.001 | |
| 3–5/high power field | 0.07 | 0.03 | |||
| 5–10//high power field | 0.03 | 0.12 | |||
| 10–15/high power field | 0.03 | 0.22 | |||
| 15–20/high power field | 0.03 | 0.04 | |||
| 20–25/high power field | 0.06 | 0.26 | |||
| WBC | 1–3/high power field | 0.66 | 0.45 | 0.006 | |
| 3–5/high power field | 0.05 | 0.14 | |||
| 5–10/high power field | 0.19 | 0.20 | |||
| 10–15/high power field | 0.01 | 0.05 | |||
| 20–25/high power field | 0.08 | 0.15 | |||
| Squamous epithelial cells | single per high power field | 0.82 | 0.69 | 0.177 | |
| sparse per high power field | 0.10 | 0.23 | |||
| many per high power field | 0.08 | 0.08 | |||
| Mucus thread | single per high power field | 0.00 | 0.38 | 0.851 | |
| sparse per high power field | 0.80 | 0.24 | |||
| many per high power field | 0.20 | 0.38 | |||
| Bacteria | lack of/single per high power field | 0.65 | 0.01 | <0.001 | |
| sparse per high power field | 0.27 | 0.39 | |||
| many per high power field | 0.08 | 0.60 | |||
| Additional information | |||||
| Feature/Parameters | Controls (n = 114) Frequency | Patients with BC (n = 116) Frequency | p | ||
| Comorbidities | hypertension | yes | 0.53 | 0.31 | <0.001 |
| no | 0.47 | 0.69 | |||
| diabetes | yes | 0.17 | 0.16 | 0.955 | |
| no | 0.83 | 0.84 | |||
| hypercholesterolaemia | yes | 0.22 | 0.10 | 0.017 | |
| no | 0.78 | 0.90 | |||
| Family history of bladder cancer | yes | n/d | 0.10 | n/d | |
| no | n/d | 0.90 | |||
| Actual therapy for bladder cancer | surgical treatment—TURBT (transurethral resection of bladder tumour) | n/d | 0.91 | n/d | |
| chemotherapy | n/d | 0.03 | |||
| cystectomy | n/d | 0.06 | |||
| Symptoms accompanying BC | |||||
| Haematuria with clots | yes | n/d | 0.68 | n/d | |
| no | n/d | 0.32 | |||
| Dysuria | yes | n/d | 0.65 | n/d | |
| no | n/d | 0.35 | |||
| Recurrent urinary tract infections | yes | n/d | 0.63 | n/d | |
| no | n/d | 0.37 | |||
| Pollakiuria | yes | n/d | 0.61 | n/d | |
| no | n/d | 0.39 | |||
| Urgent pressures | yes | n/d | 0.68 | n/d | |
| no | n/d | 0.32 | |||
| The feeling of something left behind after voiding | yes | n/d | 0.08 | n/d | |
| no | n/d | 0.92 | |||
| Waiting for micturition | yes | n/d | 0.05 | n/d | |
| no | n/d | 0.95 | |||
| Urinary incontinence problem | yes | n/d | 0.54 | n/d | |
| no | n/d | 0.46 | |||
| Lower abdominal pain | yes | n/d | 0.51 | n/d | |
| no | n/d | 0.49 | |||
| Weight loss | yes | n/d | 0.41 | n/d | |
| no | n/d | 0.59 | |||
| Zubrod’s fitness level (ECOG scale) | 0 | n/d | 0.84 | n/d | |
| 1 | n/d | 0.09 | |||
| 2 | n/d | 0.02 | |||
| 3 | n/d | 0.04 | |||
| 4 | n/d | 0.01 | |||
| 5 | n/d | 0 | |||
| TNM Classification of Malignant Tumours (TNM) | Tx | n/d | 0.01 | n/d | |
| T0 | n/d | 0.01 | |||
| Ta | n/d | 0.38 | |||
| Tis | n/d | 0.01 | |||
| T1 | n/d | 0.41 | |||
| T2 | n/d | 0.17 | |||
| T3 | n/d | 0.01 | |||
| T4 | n/d | 0.00 | |||
| Status of regional lymph nodes | N0–N1 | n/d | 0.82 | n/d | |
| ≥N2 | n/d | 0.18 | |||
| The presence of distant metastases | M0 | n/d | 0.82 | n/d | |
| M1 | n/d | 0.18 | |||
| Pathomorphology of nonmuscle-invasive tumours | urothelial papilloma | n/d | 0.02 | n/d | |
| inverted papilloma | n/d | 0.00 | |||
| papillary urothelial neoplasm of low malignant potential (PUN-LMP) | n/d | 0.16 | |||
| low-grade papillary urothelial carcinoma (LG) | n/d | 0.42 | |||
| high-grade papillary urothelial carcinoma (HG) | n/d | 0.36 | |||
| Pathomorphology of tumours infiltrating the muscle membrane | invasive urothelial carcinoma | n/d | 0.00 | n/d | |
| squamous cell carcinoma | n/d | 0.04 | |||
| glandular carcinoma | n/d | 0.00 | |||
| small cell carcinoma | n/d | 0.00 | |||
| undifferentiated carcinoma | n/d | 0.00 | |||
| other | n/d | 0.02 | |||
| Genotype/Allele | Control (n = 114) | BC (n = 116) | Crude OR (95% CI) * | p | Adjusted OR (95% CI) * | p | ||
|---|---|---|---|---|---|---|---|---|
| Number | Frequency | Number | Frequency | |||||
| -597 A>G—IL-6 (rs1800797) | ||||||||
| Frequencies | ||||||||
| A/A | 25 | 0.219 | 24 | 0.207 | 0.929 (0.494–1.746) | 0.818 | 0.905 (0.480–1.707) | 0.757 |
| A/G | 57 | 0.500 | 61 | 0.526 | 1.109 (0.661–1.861) | 0.695 | 1.113 (0.663–1.870) | 0.685 |
| G/G | 32 | 0.281 | 31 | 0.267 | 0.935 (0.523–1.669) | 0.819 | 0.950 (0.531–1.701) | 0.864 |
| χ2 = 230.000; p = 0.432 | ||||||||
| A | 107 | 0.469 | 109 | 0.470 | 1.002 (0.691–1.454) | 0.991 | 0.986 (0.679–1.433) | 0.942 |
| G | 121 | 0.531 | 123 | 0.530 | 0.998 (0.688–1.447) | 0.991 | 1.014 (0.698–1.474) | 0.942 |
| Carriage rates | ||||||||
| A (+) | 81 | 0.355 | 85 | 0.366 | 1.117 (0.627–1.989) | 0.707 | 1.101 (0.617–1.965) | 0.744 |
| A (−) | 33 | 0.145 | 31 | 0.134 | 0.895 (0.503–1.594) | 0.707 | 0.908 (0.509–1.610) | 0.744 |
| G (+) | 91 | 0.399 | 92 | 0.397 | 0.969 (0.510–1.840) | 0.923 | 0.989 (0.520–1.883) | 0.974 |
| G (−) | 23 | 0.101 | 24 | 0.103 | 1.032 (0.544–1.960) | 0.923 | 1.011 (0.531–1.924) | 0.974 |
| c.3331 G>A—IL-6 (rs2069845) | ||||||||
| Frequencies | ||||||||
| G/G | 26 | 0.228 | 25 | 0.216 | 0.930 (0.499–1.733) | 0.819 | 0.917 (0.491–1.712) | 0.785 |
| G/A | 56 | 0.491 | 63 | 0.543 | 1.231 (0.733–2.067) | 0.431 | 1.228 (0.730–2.063) | 0.439 |
| A/A | 32 | 0.281 | 28 | 0.241 | 0.815 (0.452–1.470) | 0.497 | 0.828 (0.458–1.497) | 0.533 |
| χ2 = 230.000; p = 0.432 | ||||||||
| G | 108 | 0.474 | 113 | 0.487 | 1.057 (0.728–1.500) | 0.770 | 1.045 (0.719–1.520) | 0.817 |
| A | 120 | 0.526 | 119 | 0.513 | 0.946 (0.652–1.373) | 0.770 | 0.957 (0.658–1.391) | 0.817 |
| Carriage rates | ||||||||
| G (+) | 82 | 0.360 | 88 | 0.379 | 1.226 (0.680–2.212) | 0.497 | 1.207 (0.668–2.182) | 0.533 |
| G (−) | 32 | 0.140 | 28 | 0.121 | 0.815 90.452–1.470) | 0.497 | 0.828 (0.458–1.497) | 0.533 |
| A (+) | 88 | 0.386 | 91 | 0.392 | 1.075 (0.577–2.004) | 0.819 | 1.091 (0.584–2.037) | 0.785 |
| A (−) | 26 | 0.114 | 25 | 0.108 | 0.930 (0.499–1.733) | 0.819 | 0.917 (0.491–1.712) | 0.785 |
| c.+396 T>G—IL-8 (rs2227307) | ||||||||
| Frequencies | ||||||||
| T/T | 27 | 0.237 | 30 | 0.259 | 1.124 (0.617–2.047) | 0.702 | 1.120 (0.614–2.042) | 0.712 |
| T/G | 61 | 0.535 | 61 | 0.526 | 0.964 (0.574–1.618) | 0.889 | 0.965 (0.574–1.622) | 0.894 |
| G/G | 26 | 0.228 | 25 | 0.216 | 0.930 (0.499–1.733) | 0.819 | 0.931 (0.499–1.738) | 0.823 |
| χ2 = 230.000; p = 0.432 | ||||||||
| T | 115 | 0.504 | 121 | 0.522 | 1.076 (0.738–1.570) | 0.704 | 1.074 (0.735–1.568) | 0.713 |
| G | 113 | 0.496 | 111 | 0.478 | 0.929 (0.637–1.356) | 0.704 | 0.931 (0.638–1.360) | 0.713 |
| Carriage rates | ||||||||
| T (+) | 88 | 0.386 | 91 | 0.392 | 1.075 (0.577–2.004) | 0.819 | 1.074 (0.575–2.004) | 0.823 |
| T (−) | 26 | 0.114 | 25 | 0.108 | 0.930 (0.499–1.733) | 0.819 | 0.931 (0.499–1.738) | 0.823 |
| G (+) | 87 | 0.382 | 86 | 0.371 | 0.890 (0.489–1.620) | 0.702 | 0.893 (0.490–1.628) | 0.712 |
| G (−) | 27 | 0.118 | 30 | 0.129 | 1.124 (0.617–2.047) | 0.702 | 1.120 (0.614–2.042) | 0.712 |
| Factor | Gene | Study Groups | Gender/BMI/Smoking | Interaction | |||
|---|---|---|---|---|---|---|---|
| F | p | F | p | F | p * | ||
| Gender | IL-6 | 0.714 | 0.399 | 0.717 | 0.398 | 0.710 | 0.400 |
| IL-8 | 36.725 | <0.001 | 11.674 | <0.001 | 13.151 | <0.001 | |
| BMI | IL-6 | 1.045 | 0.308 | 0.166 | 0.684 | 0.162 | 0.688 |
| IL-8 | 24.560 | <0.001 | 0.135 | 0.713 | 0.196 | 0.658 | |
| Smoking | IL-6 | 0.932 | 0.336 | 0.926 | 0.337 | 0.935 | 0.335 |
| IL-8 | 25.526 | <0.001 | 1.059 | 0.305 | 0.400 | 0.528 | |
| Factor | Study Groups | Gender/BMI/Smoking | Interaction | |||
|---|---|---|---|---|---|---|
| F | p | F | p | F | p * | |
| Gender | 10.844 | 0.001 | 1.682 | 0.196 | 6.783 | 0.010 |
| BMI | 13.810 | <0.001 | 0.827 | 0.364 | 0.026 | 0.871 |
| Smoking | 20.848 | <0.001 | 0.681 | 0.410 | 0.355 | 0.552 |
| Gene | SNPs | NCBI db SNP ID | Region | MAF in European Population | Assay ID of TaqMan™ SNP Genotyping Assay | Location | Function | Ref. |
|---|---|---|---|---|---|---|---|---|
| IL-6 | -597 A>G | rs1800797 | Intron | G: 0.609 | C___1839695_20 | Chr.7: 22726602 | The G allele was associated with increased inflammatory responses | [22] |
| c.3331 G>A | rs2069845 | Intron | A: 0.590 | C___1839699_10 | Chr.7: 22730530 | G carriers were characterised by high secretion of IL-6 in serum | [44] | |
| IL-8 | c.+396 T>G | rs2227307 | Intron | G: 0.447 | C__11748168_10 | Chr.4: 73740952 | This polymorphism may regulate the protein production of IL-8 | [45] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grębowski, R.; Saluk, J.; Bijak, M.; Szemraj, J.; Wigner, P. Variability, Expression, and Methylation of IL-6 and IL-8 Genes in Bladder Cancer Pathophysiology. Int. J. Mol. Sci. 2023, 24, 6266. https://doi.org/10.3390/ijms24076266
Grębowski R, Saluk J, Bijak M, Szemraj J, Wigner P. Variability, Expression, and Methylation of IL-6 and IL-8 Genes in Bladder Cancer Pathophysiology. International Journal of Molecular Sciences. 2023; 24(7):6266. https://doi.org/10.3390/ijms24076266
Chicago/Turabian StyleGrębowski, Radosław, Joanna Saluk, Michał Bijak, Janusz Szemraj, and Paulina Wigner. 2023. "Variability, Expression, and Methylation of IL-6 and IL-8 Genes in Bladder Cancer Pathophysiology" International Journal of Molecular Sciences 24, no. 7: 6266. https://doi.org/10.3390/ijms24076266