
iE-DAP Induced Inflammatory Response and Tight Junction Disruption in Bovine Mammary Epithelial Cells via NOD1-Dependent NF-κB and MLCK Signaling Pathway
 , and
, and
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
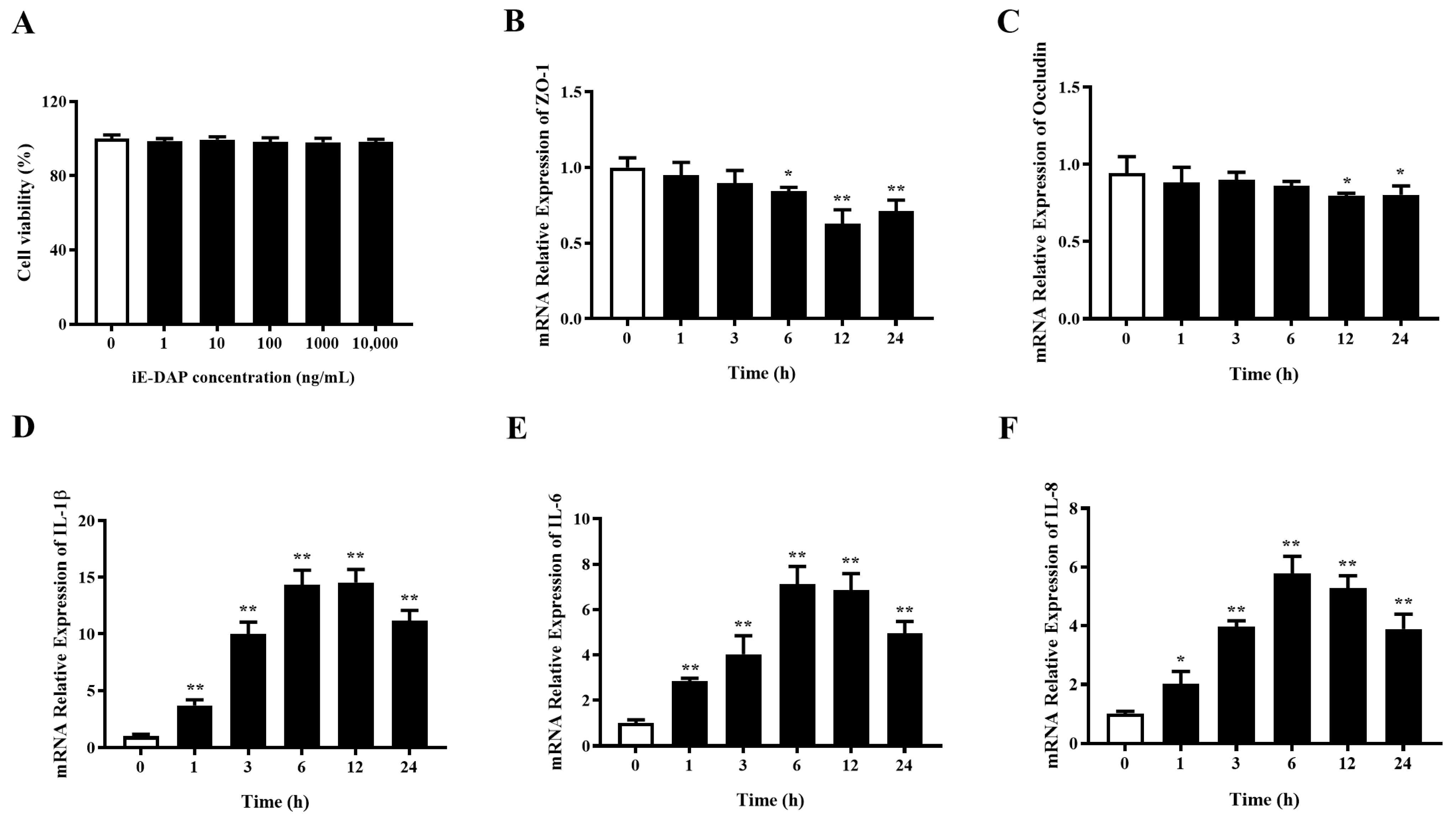
2.1. Time Gradient Effects of iE-DAP Treatment on the mRNA Expression of Inflammatory Cytokines and Tight Junction Protein Genes in BMECs
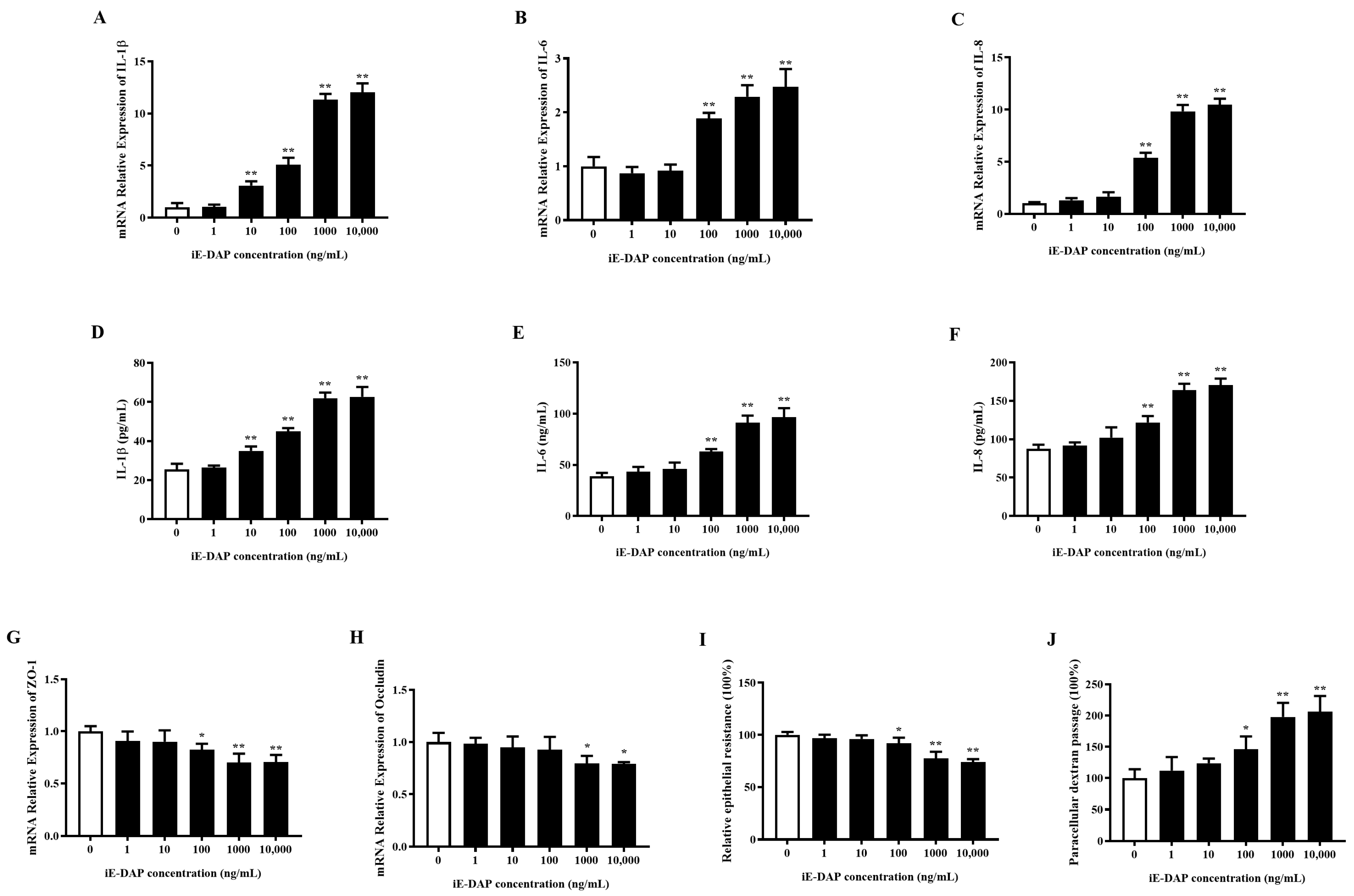
2.2. Gradient Effects of iE-DAP Treatment on the Inflammatory Response and Tight Junction Integrity in BMECs
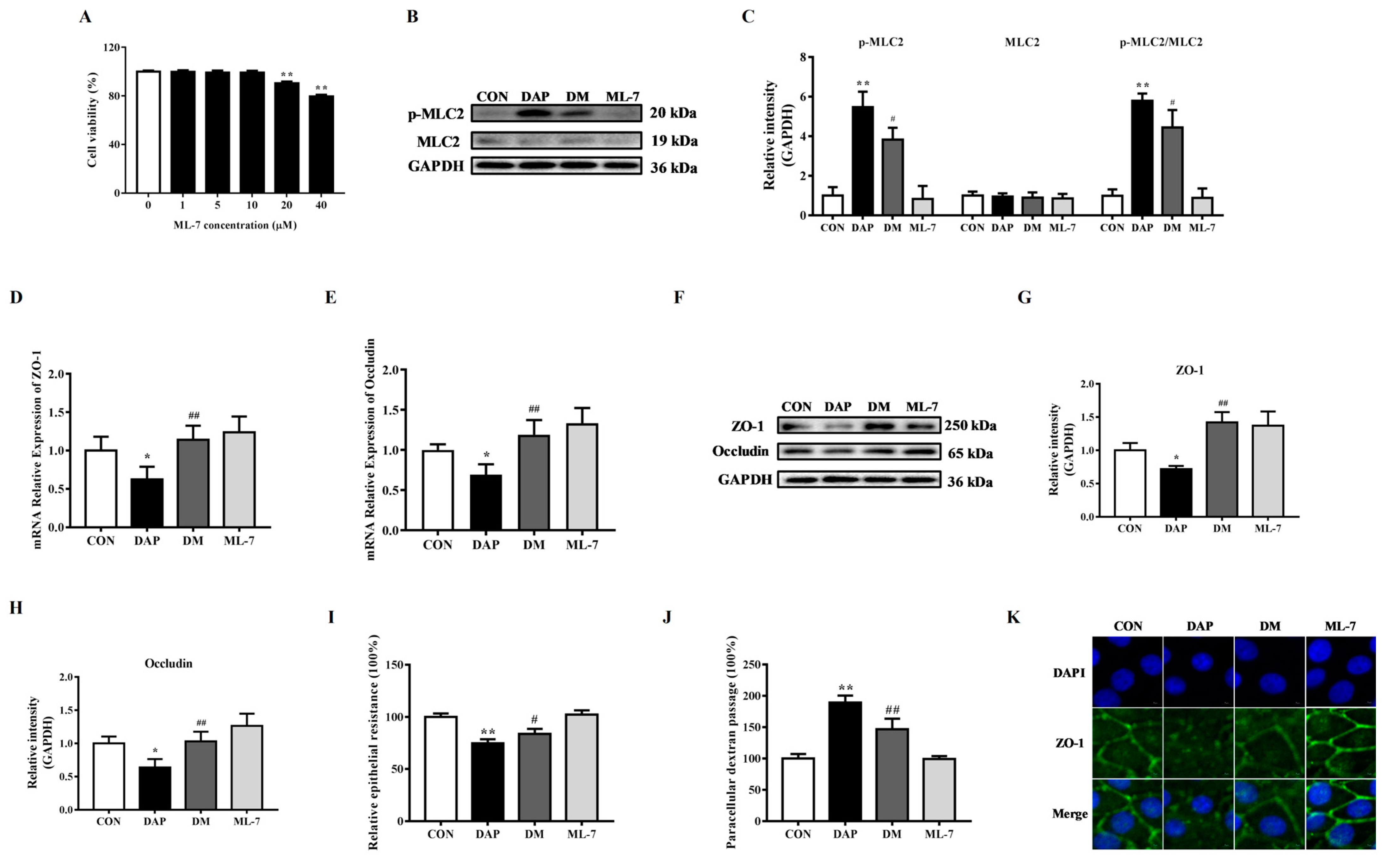
2.3. Effects of ML-7 Pretreatment on Tight Junction Disruption in BMECs Induced by iE-DAP
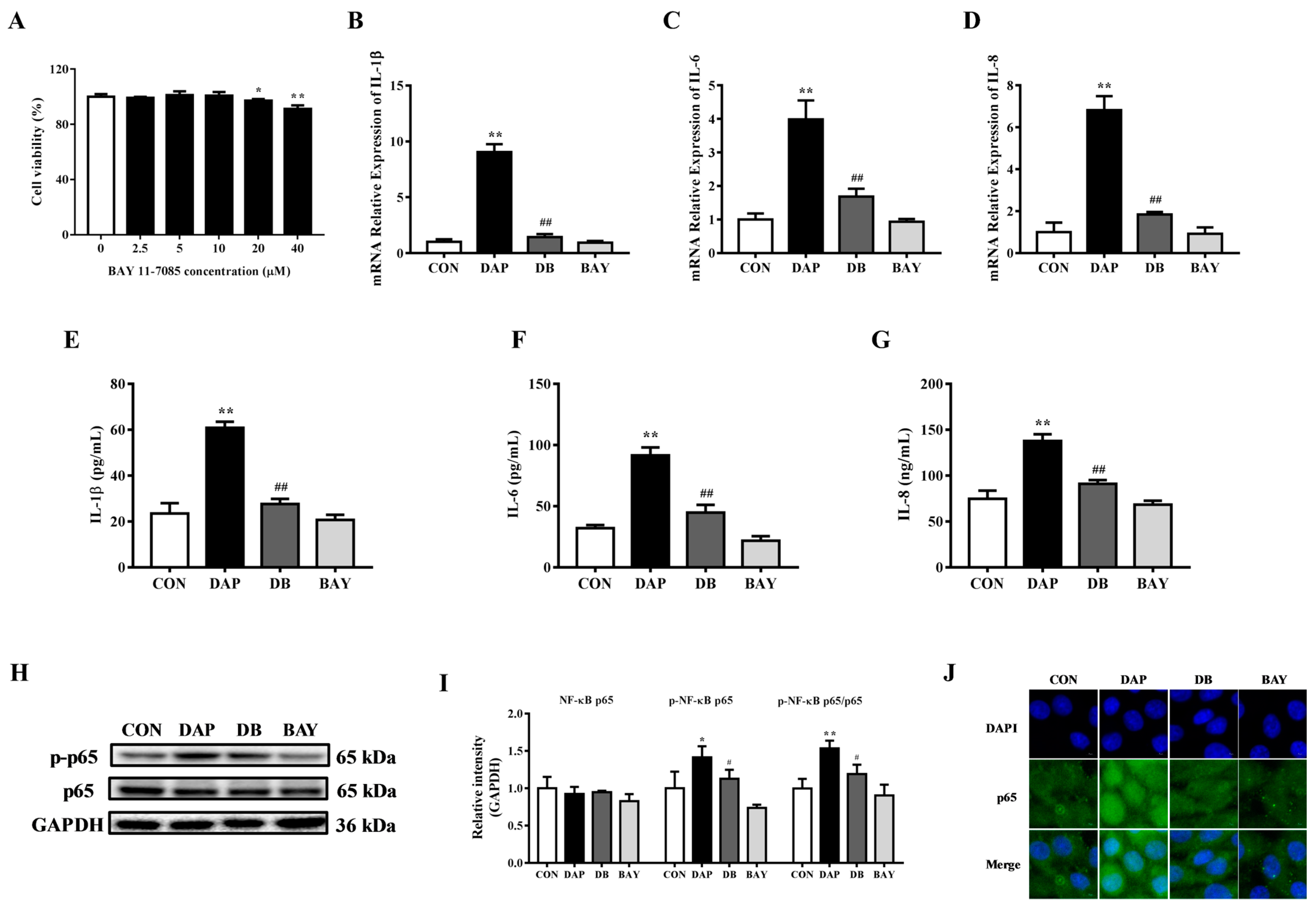
2.4. Effects of BAY 11-7085 Pretreatment on the Inflammatory Response in BMECs Induced by iE-DAP
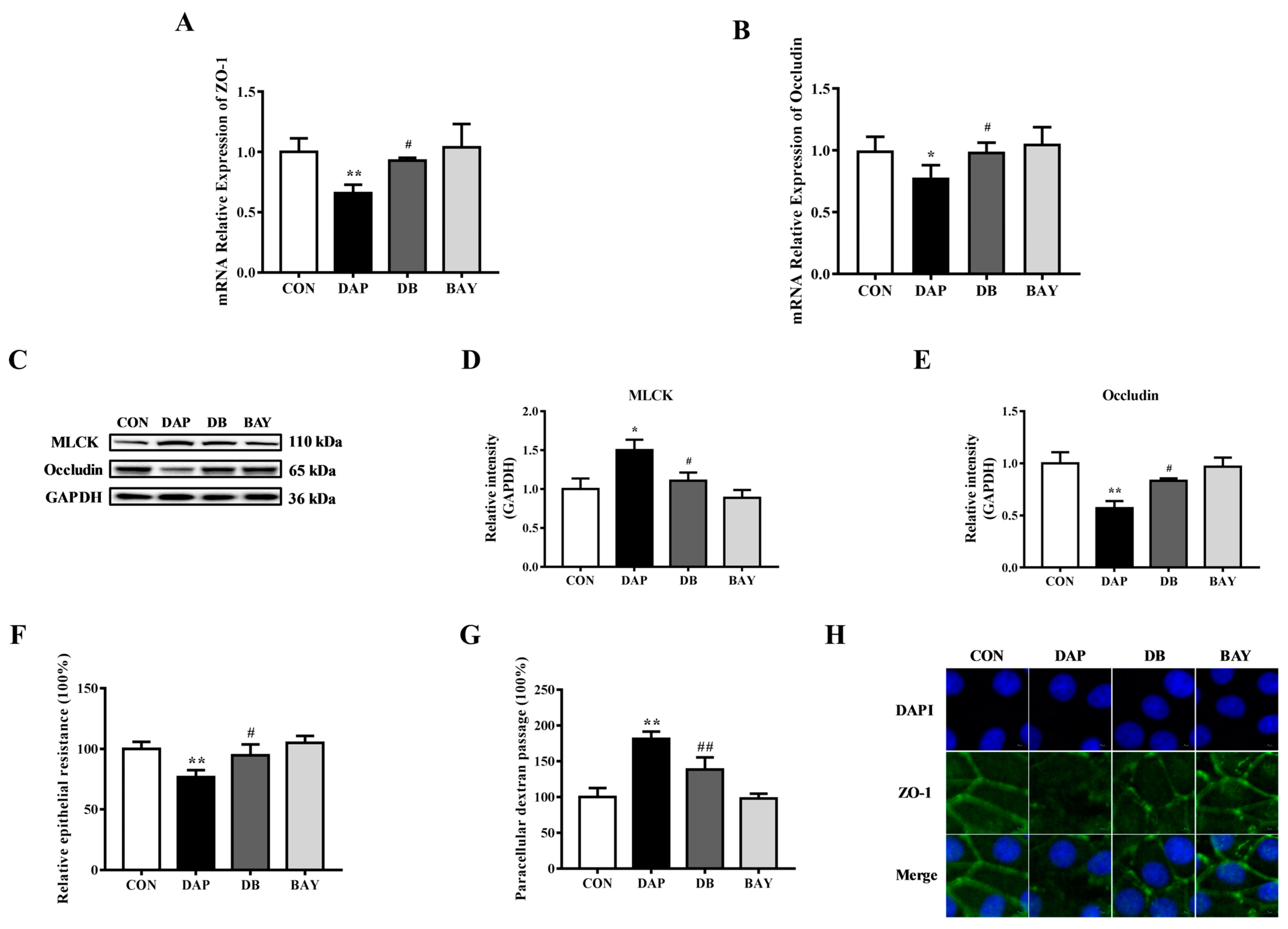
2.5. Effects of BAY 11-7085 Pretreatment on the Tight Junction Disruption in BMECs Induced by iE-DAP
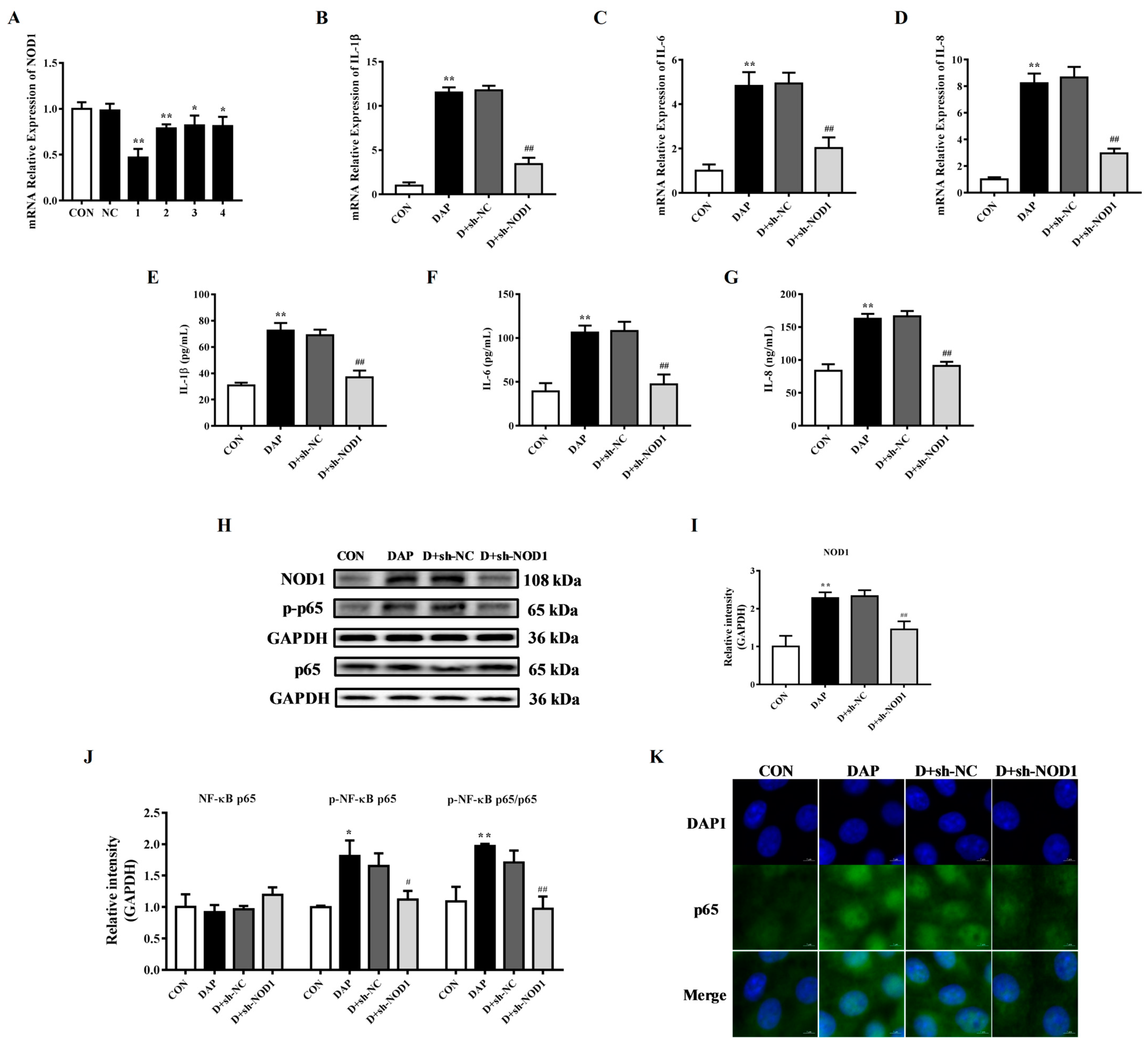
2.6. Effects of NOD1 Interference on the Inflammatory Response in BMECs Induced by iE-DAP
2.7. Effects of NOD1 Interference on the Tight Junction Disruption in BMECs Induced by iE-DAP
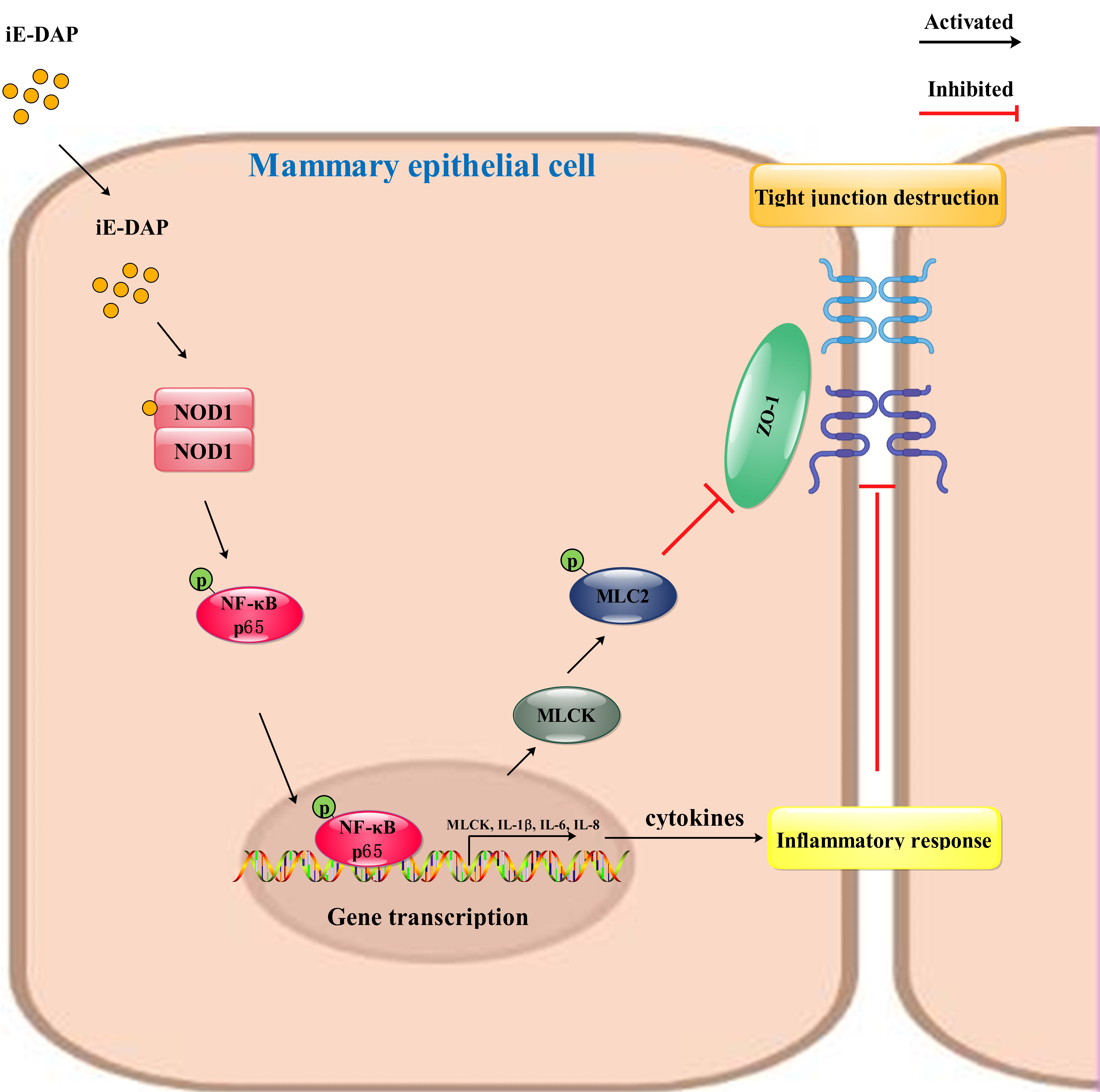
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Cell Culture
4.3. Experimental Design
4.4. Cell Viability Assay
4.5. Inflammatory Cytokine Concentration Assay
4.6. Real-Time Quantitative PCR Assay
4.7. Western Blotting Assay
4.8. Transepithelial Electrical Resistance and Paracellular Permeability Assay
4.9. Cellular Immunofluorescence Assay
4.10. Short Hairpin RNA Transfection Assay
4.11. Data Statistical Assay
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hertl, J.A.; Groehn, Y.T.; Leach, J.D.G.; Bar, D.; Bennett, G.J.; Gonzalez, R.N.; Rauch, B.J.; Welcome, F.L.; Tauer, L.W.; Schukken, Y.H. Effects of clinical mastitis caused by gram-positive and gram-negative bacteria and other organisms on the probability of conception in New York State Holstein dairy cows. J. Dairy Sci. 2010, 93, 1551–1560. [Google Scholar] [CrossRef] [PubMed]
- Hogeveen, H.; Steeneveld, W.; Wolf, C.A. Production diseases reduce the efficiency of dairy production: A review of the results, methods, and approaches regarding the economics of mastitis. Annu. Rev. Resour. Econ. 2019, 11, 289–312. [Google Scholar] [CrossRef] [Green Version]
- Fogsgaard, K.K.; Bennedsgaard, T.W.; Herskin, M.S. Behavioral changes in freestall-housed dairy cows with naturally occurring clinical mastitis. J. Dairy Sci. 2015, 98, 1730–1738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, X.; Li, S.; Fu, Y.; Zhang, N. Targeting gut microbiota as a possible therapy for mastitis. Eur. J. Clin. Microbiol. Infect. Dis. 2019, 38, 1409–1423. [Google Scholar] [CrossRef]
- Kai, Z.; Chang, G.; Xu, T.; Lei, X.; Guo, J.; Di, J.; Shen, X. Lipopolysaccharide derived from the digestive tract activates inflammatory gene expression and inhibits casein synthesis in the mammary glands of lactating dairy cows. Oncotarget 2016, 7, 9652–9665. [Google Scholar]
- Wang, Y.; Zhang, W.; Ma, N.; Wang, L.; Dai, H.; Bilal, M.S.; Roy, A.C.; Shen, X. Overfeeding with a high-concentrate diet activates the NOD1-NF-kappa B signalling pathway in the mammary gland of mid-lactating dairy cows. Microb. Pathog. 2019, 128, 390–395. [Google Scholar] [CrossRef]
- Chamaillard, M.; Hashimoto, M.; Horie, Y.; Masumoto, J.; Su, Q.; Saab, L.; Ogura, Y.; Kawasaki, A.; Fukase, K.; Kusumoto, S. An essential role for NOD1 in host recognition of bacterial peptidoglycan containing diaminopimelic acid. Nat. Immunol. 2003, 4, 702–707. [Google Scholar] [CrossRef]
- Girardin, S.E.; Travassos, L.H.; Hervé, M.; Blanot, D.; Boneca, I.G.; Philpott, D.J.; Sansonetti, P.J.; Menginlecreulx, D. Peptidoglycan molecular requirements allowing detection by Nod1 and Nod2. J. Biol. Chem. 2003, 278, 41702–41708. [Google Scholar] [CrossRef] [Green Version]
- Bi, D.; Gao, Y.; Chu, Q.; Cui, J.; Xu, T. NOD1 is the innate immune receptor for iE-DAP and can activate NF-kappa B pathway in teleost fish. Dev. Comp. Immunol. 2017, 76, 238–246. [Google Scholar] [CrossRef]
- Caruso, R.; Warner, N.; Inohara, N.; Nunez, G. NOD1 and NOD2: Signaling, host defense, and inflammatory disease. Immunity 2014, 41, 898–908. [Google Scholar] [CrossRef] [Green Version]
- Gong, Q.; Long, Z.; Zhong, F.L.; Teo DE, T.; Jin, Y.; Yin, Z.; Boo, Z.Z.; Zhang, Y.; Zhang, J.; Yang, R.; et al. Structural basis of RIP2 activation and signaling. Nat. Commun. 2018, 9, 4993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roy, A.C.; Chang, G.J.; Ma, N.N.; Wang, Y.; Roy, S.; Liu, J.; Aabdin, Z.U.; Shen, X.Z. Sodium butyrate suppresses NOD1-mediated inflammatory molecules expressed in bovine hepatocytes during iE-DAP and LPS treatment. J. Cell. Physiol. 2019, 234, 19602–19620. [Google Scholar] [CrossRef] [PubMed]
- Bonnefont, C.M.; Rainard, P.; Cunha, P.; Gilbert, F.B.; Toufeer, M.; Aurel, M.R.; Rupp, R.; Foucras, G. Genetic susceptibility to S. aureus mastitis in sheep: Differential expression of mammary epithelial cells in response to live bacteria or supernatant. Physiol. Genom. 2012, 44, 403–416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korhonen, H.; Marnila, P.; Gill, H.S. Milk immunoglobulins and complement factors. Br. J. Nutr. 2000, 84, S75–S80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilbert, F.B.; Cunha, P.; Jensen, K.; Glass, E.J.; Foucras, G.; Robert-Granie, C.; Rupp, R.; Rainard, P. Differential response of bovine mammary epithelial cells to Staphylococcus aureus or Escherichia coli agonists of the innate immune system. Vet. Res. 2013, 44, 40. [Google Scholar] [CrossRef] [Green Version]
- Bertero, A.; Augustyniak, J.; Buzanska, L.; Caloni, F. Species-specific models in toxicology: In vitro epithelial barriers. Environ. Toxicol. Pharmacol. 2019, 70, 103203. [Google Scholar] [CrossRef]
- Xu, T.; Dong, Z.; Wang, X.; Qi, S.; Li, X.; Cheng, R.; Liu, X.; Zhang, Y.; Gao, M.Q. IL-1 beta induces increased tight junction permeability in bovine mammary epithelial cells via the IL-1-ERK1/2-MLCK axis upon blood-milk barrier damage. J. Cell. Biochem. 2018, 119, 9028–9041. [Google Scholar] [CrossRef]
- Wall, S.K.; Wellnitz, O.; Hernández-Castellano, L.E.; Ahmadpour, A.; Bruckmaier, R.M. Supraphysiological oxytocin increases the transfer of immunoglobulins and other blood components to milk during lipopolysaccharide- and lipoteichoic acid–induced mastitis in dairy cows. J. Dairy Sci. 2016, 99, 9165–9173. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, K.; Oyama, S.; Numata, A.; Rahman, M.M.; Kumura, H. Lipopolysaccharide disrupts the milk-blood barrier by modulating claudins in mammary alveolar tight junctions. PLoS ONE 2013, 8, e62187. [Google Scholar] [CrossRef] [Green Version]
- Stelwagen, K.; Singh, K. The role of tight junctions in mammary gland function. J. Mammary Gland. Biol. Neoplasia 2014, 19, 131–138. [Google Scholar] [CrossRef]
- Park, H.-Y.; Kunitake, Y.; Hirasaki, N.; Tanaka, M.; Matsui, T. Theaflavins enhance intestinal barrier of Caco-2 Cell monolayers through the expression of AMP-activated protein kinase-mediated Occludin, Claudin-1, and ZO-1. Biosci. Biotechnol. Biochem. 2015, 79, 130–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walsh, S.V.; Hopkins, A.M.; Nusrat, A. Modulation of tight junction structure and function by cytokines. Adv. Drug Deliv. Rev. 2000, 41, 303–313. [Google Scholar] [CrossRef]
- Wang, J.J.; Wei, Z.K.; Zhang, X.; Wang, Y.N.; Fu, Y.H.; Yang, Z.T. Butyrate protects against disruption of the blood-milk barrier and moderates inflammatory responses in a model of mastitis induced by lipopolysaccharide. Br. J. Pharmacol. 2017, 174, 3811–3822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe, A.; Yagi, Y.; Shiono, H.; Yokomizo, Y.; Inumaru, S. Effects of intramammary infusions of interleukin-8 on milk protein composition and induction of acute-phase protein in cows during mammary involution. Can. J. Vet. Res. 2008, 72, 291–296. [Google Scholar]
- Al-Sadi, R.; Guo, S.; Ye, D.; Rawat, M.; Ma, T.Y. TNF-alpha modulation of intestinal tight junction permeability is mediated by NIK/IKK-alpha axis activation of the canonical NF-kappaB pathway. Am. J. Pathol. 2016, 186, 1151–1165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gadina, M.; Gazaniga, N.; Vian, L.; Furumoto, Y. Small molecules to the rescue: Inhibition of cytokine signaling in immune-mediated diseases. J. Autoimmun. 2017, 85, 20–31. [Google Scholar] [CrossRef] [PubMed]
- Aghamohammadi, M.; Haine, D.; Kelton, D.F.; Barkema, H.W.; Hogeveen, H.; Keefe, G.P.; Dufour, S. Herd-level mastitis-associated costs on Canadian dairy farms. Front. Vet. Sci. 2018, 5, 100. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.; Aabdin, Z.U.; Wang, Y.; Ma, N.; Dai, H.; Shi, X.; Shen, X. Glutamine pretreatment protects bovine mammary epithelial cells from inflammation and oxidative stress induced by γ-d-glutamyl-meso-diaminopimelic acid (iE-DAP). J. Dairy Sci. 2021, 104, 2123–2139. [Google Scholar] [CrossRef]
- Nguyen, D.A.; Neville, M.C. Tight junction regulation in the mammary gland. J. Mammary Gland. Biol. Neoplasia 1998, 3, 233–246. [Google Scholar] [CrossRef]
- Itoh, M.; Bissell, M.J. The organization of tight junctions in epithelia: Implications for mammary gland biology and breast tumorigenesis. J. Mammary Gland. Biol. Neoplasia 2003, 8, 449–462. [Google Scholar] [CrossRef] [Green Version]
- Duffy, H.S.; John, G.R.; Lee, S.C.; Brosnan, C.F.; Spray, D.C. Reciprocal regulation of the junctional proteins claudin-1 and connexin43 by interleukin-1beta in primary human fetal astrocytes. J. Neurosci. 2000, 20, Rc114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, R.; Han, X.; Uchiyama, T.; Watkins, S.K.; Yaguchi, A.; Delude, R.L.; Fink, M.P. IL-6 is essential for development of gut barrier dysfunction after hemorrhagic shock and resuscitation in mice. Am. J. Physiol. Gastroint. Liver Physiol. 2003, 285, G621–G629. [Google Scholar] [CrossRef] [PubMed]
- Phyn, C.V.C.; Stelwagen, K.; Davis, S.R.; McMahon, C.D.; Dobson, J.M.; Singh, K. Tight junction protein abundance and apoptosis during involution of rat mammary glands. J. Cell. Physiol. 2017, 232, 2075–2082. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Cai, J.; Zhao, F.; Liu, J. Low-quality rice straw forage increases the permeability of mammary epithelial tight junctions in lactating dairy cows. J. Sci. Food Agric. 2019, 99, 2037–2041. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Yu, C.; Chen, Y.; Liu, S.; Azevedo, P.; Gong, J.O.K.; Yang, C. Citral alleviates peptidoglycan-induced inflammation and disruption of barrier functions in porcine intestinal epithelial cells. J. Cell. Physiol. 2022, 237, 1768–1779. [Google Scholar] [CrossRef]
- Akbari, P.; Braber, S.; Varasteh, S.; Alizadeh, A.; Garssen, J.; Fink-Gremmels, J. The intestinal barrier as an emerging target in the toxicological assessment of mycotoxins. Arch. Toxicol. 2017, 91, 1007–1029. [Google Scholar] [CrossRef] [Green Version]
- Nighot, M.; Al-Sadi, R.; Guo, S.; Rawat, M.; Nighot, P.; Watterson, M.D.; Ma, T.Y. Lipopolysaccharide-induced increase in intestinal epithelial tight permeability is mediated by Toll-like receptor 4/myeloid differentiation primary response 88 (MyD88) activation of myosin light chain kinase expression. Am. J. Pathol. 2017, 187, 2698–2710. [Google Scholar] [CrossRef] [Green Version]
- Shen, L.; Black, E.D.; Witkowski, E.D.; Lencer, W.I.; Guerriero, V.; Schneeberger, E.E.; Turner, J.R. Myosin light chain phosphorylation regulates barrier function by remodeling tight junction structure. J. Cell Sci. 2006, 119, 2095–2106. [Google Scholar] [CrossRef] [Green Version]
- Kulms, D.; Schwarz, T. NF-κB and cytokines. Vitam. Horm. 2006, 74, 283–300. [Google Scholar]
- Senftleben, U.; Cao, Y.; Xiao, G.; Greten, F.R.; Krähn, G.; Bonizzi, G.; Chen, Y.; Hu, Y.; Fong, A.; Sun, S.C.; et al. Activation by IKKalpha of a second, evolutionary conserved, NF-kappa B signaling pathway. Science 2001, 293, 1495–1499. [Google Scholar] [CrossRef]
- Nighot, M.; Rawat, M.; Al-Sadi, R.; Castillo, E.F.; Nighot, P.; Ma, T.Y. Lipopolysaccharide-induced increase in intestinal permeability is mediated by TAK-1 activation of IKK and MLCK/MYLK gene. Am. J. Pathol. 2019, 189, 797–812. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.; Liu, B.; Hu, G.; Kan, X.; Li, Y.; Gong, Q.; Xu, D.; Ma, H.; Cao, Y.; Huang, B.; et al. Vanillin protects the blood-milk barrier and inhibits the inflammatory response in LPS-induced mastitis in mice. Toxicol. Appl. Pharm. 2019, 365, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Ying, C.; Hong, W.; Nianhui, Z.; Chunlei, W.; Kehe, H.; Cuiling, P. Nontoxic concentrations of OTA aggravate DON-induced intestinal barrier dysfunction in IPEC-J2 cells via activation of NF-kappaB signaling pathway. Toxicol. Lett. 2019, 311, 114–124. [Google Scholar] [CrossRef] [PubMed]
- Roy, A.C.; Chang, G.; Roy, S.; Ma, N.; Gao, Q.; Shen, X. gamma-d-Glutamyl-meso-diaminopimelic acid induces autophagy in bovine hepatocytes during nucleotide-binding oligomerization domain 1-mediated inflammation. J. Cell Physiol. 2021, 236, 5212–5234. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, J.; Huang, J.; Chang, G.; Roy, A.C.; Gao, Q.; Cheng, X.; Shen, X. Sodium butyrate attenuated iE-DAP induced inflammatory response in the mammary glands of dairy goats fed high-concentrate diet. J. Sci. Food Agric. 2020, 101, 1218–1227. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Li, X.; Han, Z.; Meng, M.; Shi, X.; Wang, L.; Chen, M.; Chang, G.; Shen, X. iE-DAP Induced Inflammatory Response and Tight Junction Disruption in Bovine Mammary Epithelial Cells via NOD1-Dependent NF-κB and MLCK Signaling Pathway. Int. J. Mol. Sci. 2023, 24, 6263. https://doi.org/10.3390/ijms24076263
Wang Y, Li X, Han Z, Meng M, Shi X, Wang L, Chen M, Chang G, Shen X. iE-DAP Induced Inflammatory Response and Tight Junction Disruption in Bovine Mammary Epithelial Cells via NOD1-Dependent NF-κB and MLCK Signaling Pathway. International Journal of Molecular Sciences. 2023; 24(7):6263. https://doi.org/10.3390/ijms24076263
Chicago/Turabian StyleWang, Yan, Xuerui Li, Zhengqiang Han, Meijuan Meng, Xiaoli Shi, Lairong Wang, Mengru Chen, Guangjun Chang, and Xiangzhen Shen. 2023. "iE-DAP Induced Inflammatory Response and Tight Junction Disruption in Bovine Mammary Epithelial Cells via NOD1-Dependent NF-κB and MLCK Signaling Pathway" International Journal of Molecular Sciences 24, no. 7: 6263. https://doi.org/10.3390/ijms24076263