
Alteration of Gut Microbes in Benign Prostatic Hyperplasia Model and Finasteride Treatment Model

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Validation of the Animal Model
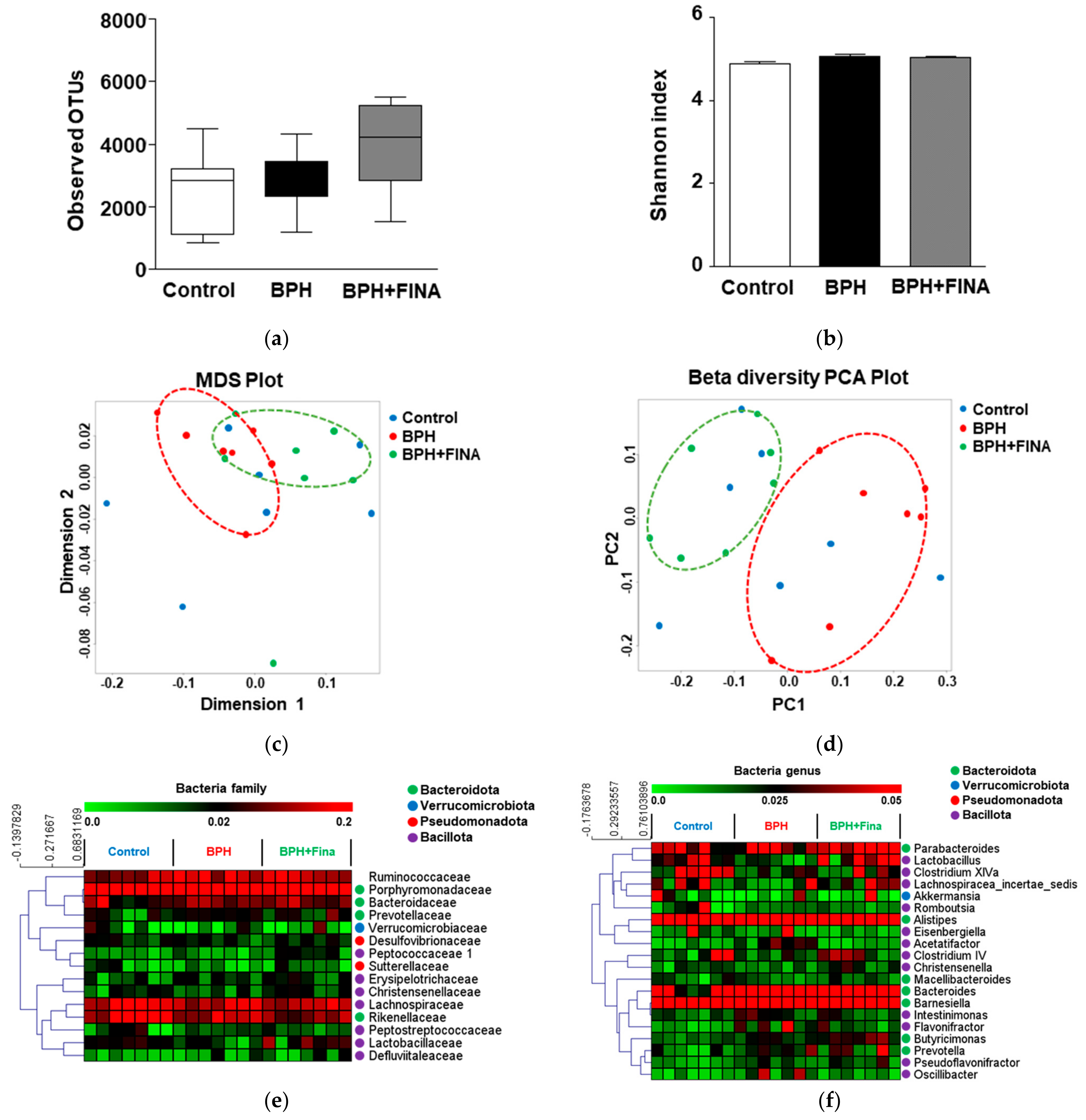
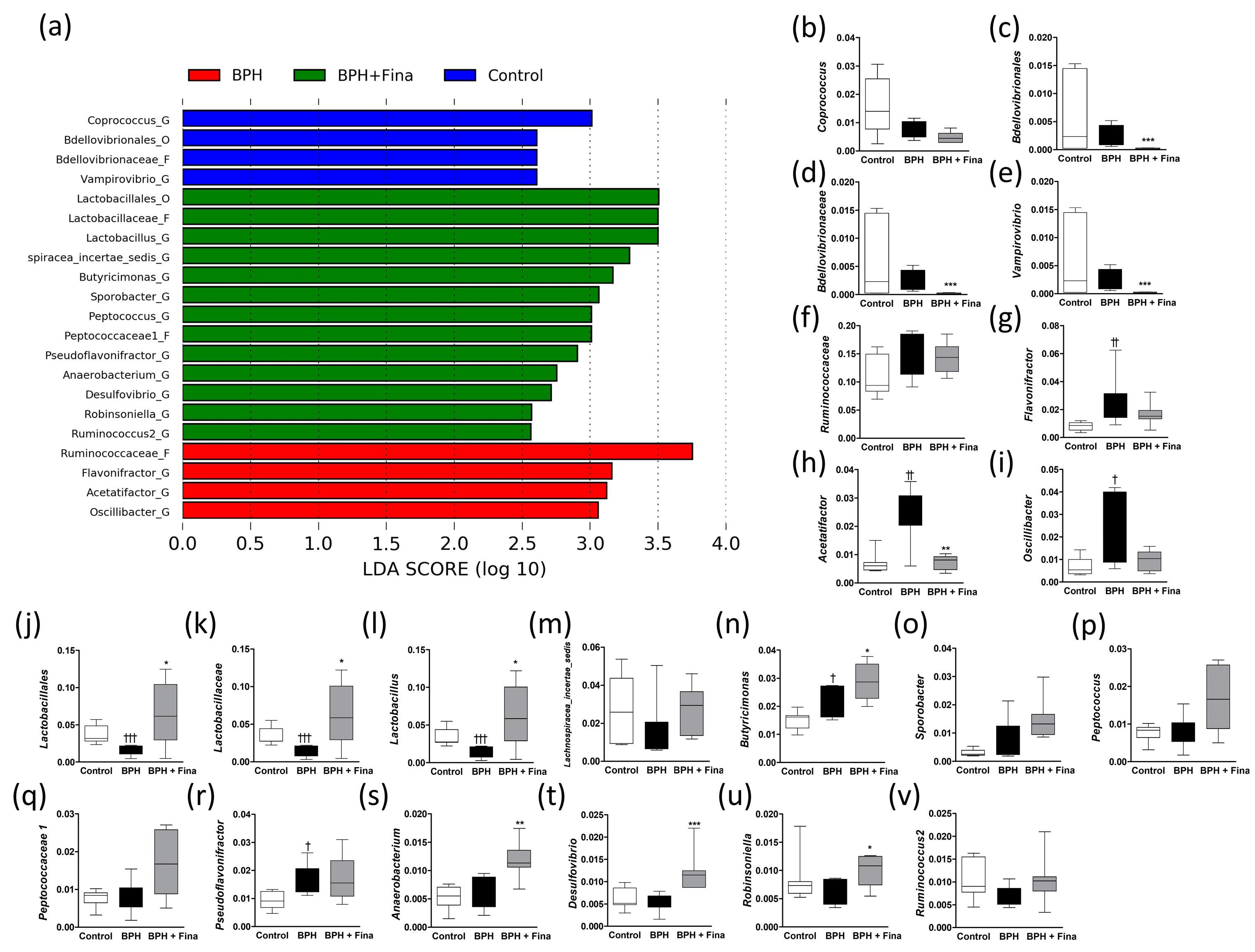
2.2. Effect of BPH Induction and Finasteride Treatment on Gut Microbiota Composition
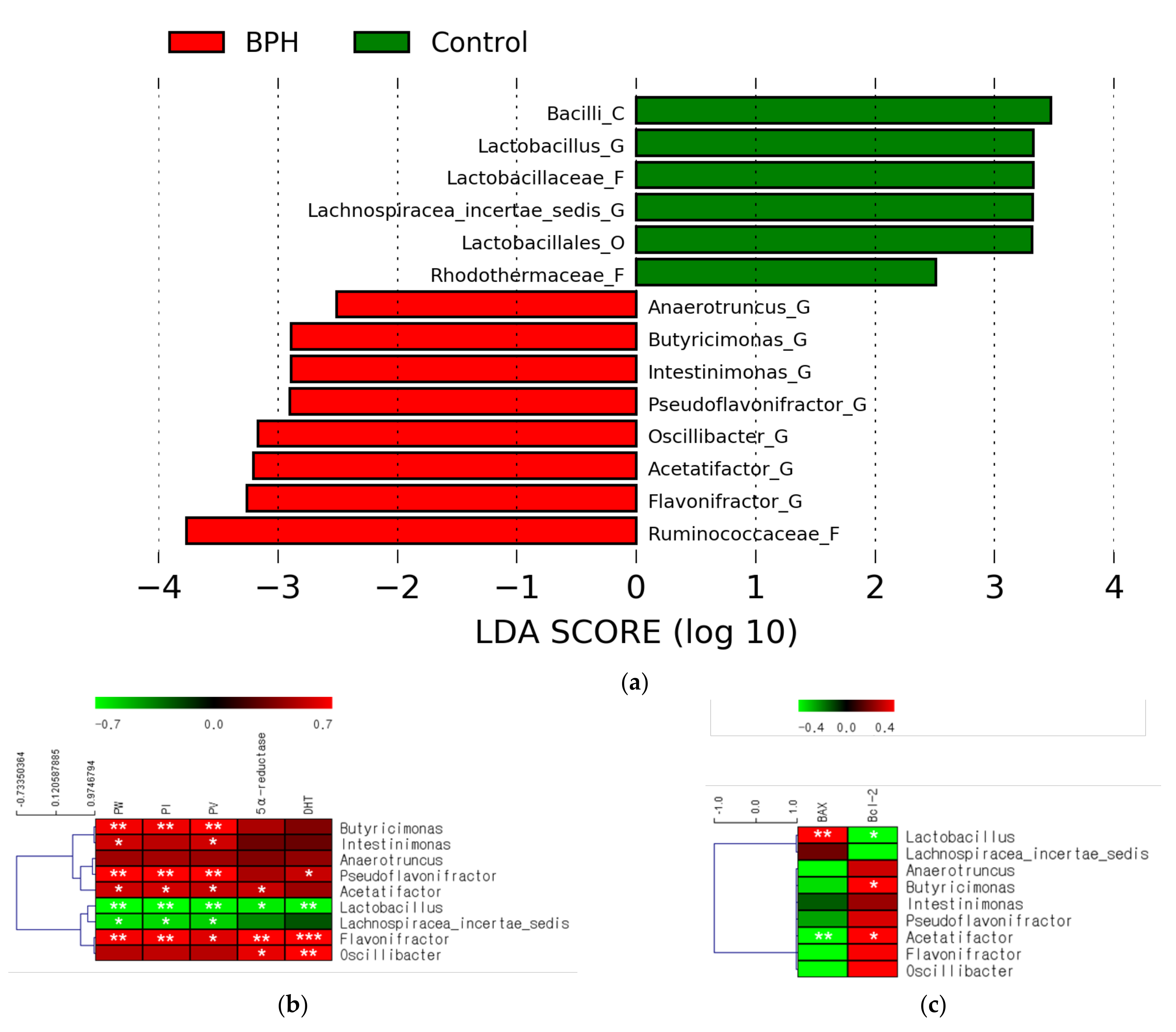
2.3. Effects of BPH Induction on Gut Microbiota
2.4. Effects of Finasteride Treatment on Gut Microbiota
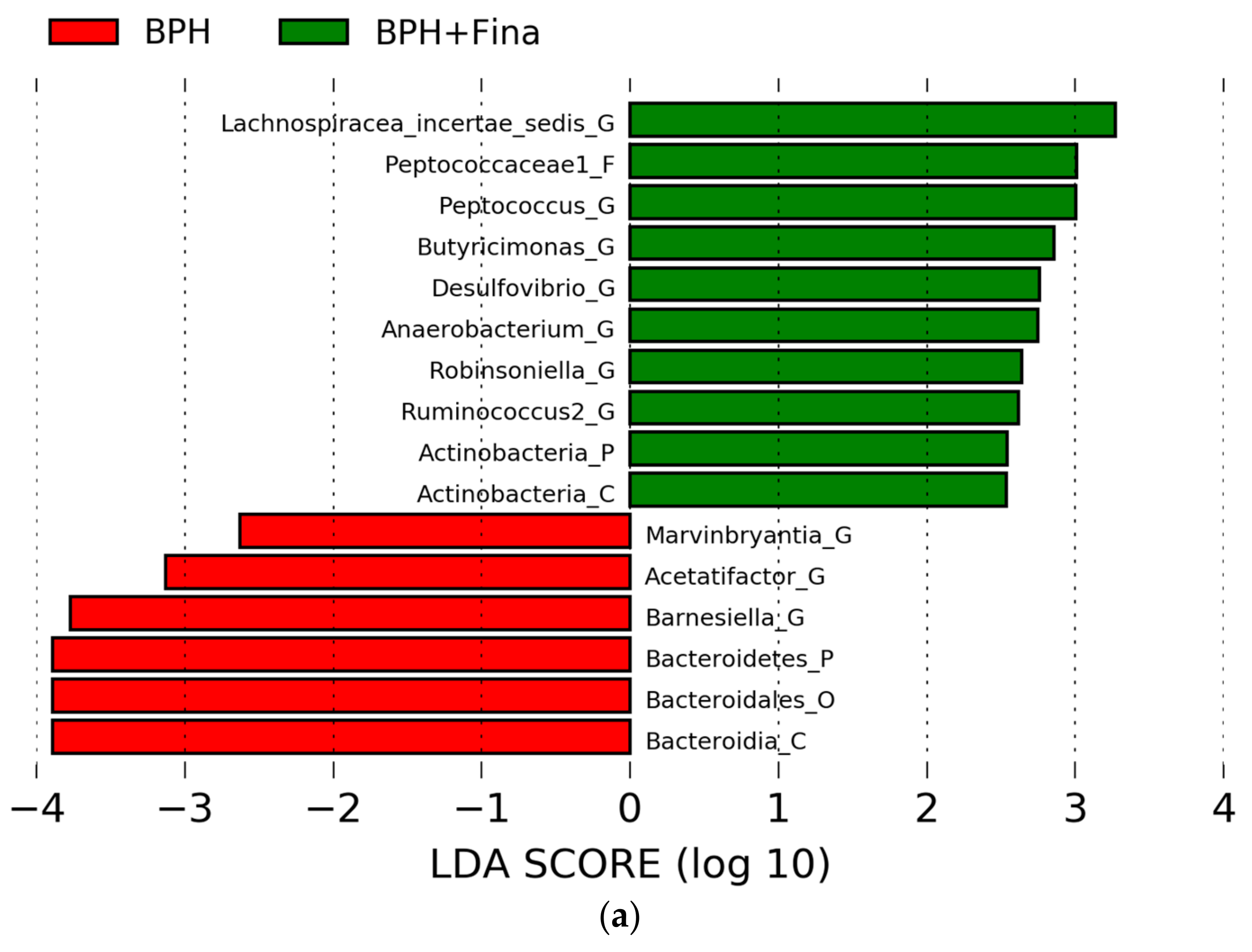
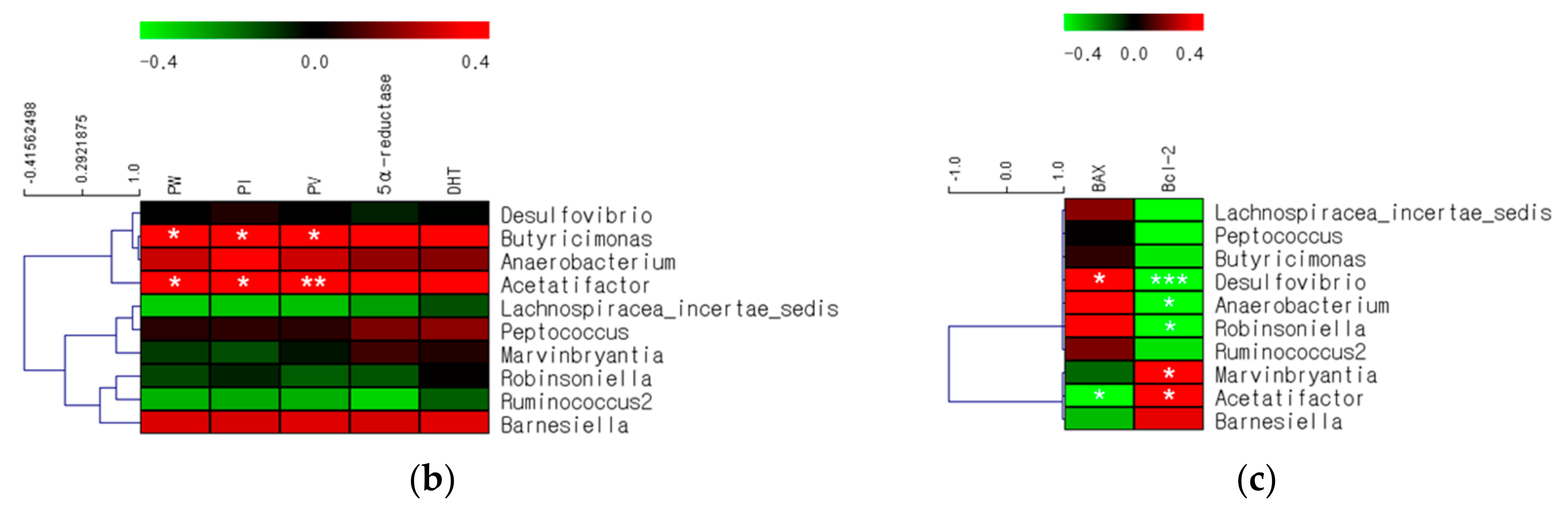
2.5. Comparison of Gut Microbes after BPH Induction and Finasteride Treatment
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. BPH Induction and Finasteride Treatment Model
4.3. Gut Microbiota Analysis
4.4. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Segal, J.P.; Mak, J.W.Y.; Mullish, B.H.; Alexander, J.L.; Ng, S.C.; Marchesi, J.R. The gut microbiome: An under-recognised contributor to the COVID-19 pandemic? Ther. Adv. Gastroenterol. 2020, 13. [Google Scholar] [CrossRef]
- Palmer, C.; Bik, E.M.; DiGiulio, D.B.; Relman, D.A.; Brown, P.O. Development of the human infant intestinal microbiota. PLoS Biol. 2007, 5, e177. [Google Scholar] [CrossRef] [Green Version]
- Shreiner, A.B.; Kao, J.Y.; Young, V.B. The gut microbiome in health and in disease. Curr. Opin. Gastroenterol. 2015, 31, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Ejtahed, H.-S.; Soroush, A.-R.; Angoorani, P.; Larijani, B.; Hasani-Ranjbar, S. Gut Microbiota as a Target in the Pathogenesis of Metabolic Disorders: A New Approach to Novel Therapeutic Agents. Horm. Metab. Res. 2016, 48, 349–358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kassabian, V.S. Sexual function in patients treated for benign prostatic hyperplasia. Lancet 2003, 361, 60–62. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.W.H.; Chan, E.M.C.; Lai, Y.K. The global burden of lower urinary tract symptoms suggestive of benign prostatic hyperplasia: A systematic review and meta-analysis. Sci. Rep. 2017, 7, 7984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Lourdes Arruzazabala, M.; Molina, V.; Más, R.; Carbajal, D.; Marrero, D.; González, V.; Rodríguez, E. Effects of coconut oil on testosterone-induced prostatic hyperplasia in Sprague-Dawley rats. J. Pharm. Pharmacol. 2007, 59, 995–999. [Google Scholar] [CrossRef] [PubMed]
- Tarter, T.; Miller, J. Combination therapy with dutasteride and tamsulosin for the treatment of symptomatic enlarged prostate. Clin. Interv. Aging 2009, 4, 251–258. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.-D.; Yang, Q.; Ceniccola, K.; Bianco, F.; Andrawis, R.; Jarrett, T.; Frazier, H.; Patierno, S.R.; Lee, N.H. Androgen Receptor-Target Genes in African American Prostate Cancer Disparities. Prostate Cancer 2013, 2013, 763569. [Google Scholar] [CrossRef] [Green Version]
- Cantlay, A.; Ni Raghallaigh, H. Benign prostatic hyperplasia. InnovAiT Educ. Inspir. Gen. Pr. 2015, 8, 238–245. [Google Scholar] [CrossRef] [Green Version]
- An, J.; Kong, H. Comparative application of testosterone undecanoate and/or testosterone propionate in induction of benign prostatic hyperplasia in Wistar rats. PLoS ONE 2022, 17, e0268695. [Google Scholar] [CrossRef]
- Lazier, C.B.; Thomas, L.N.; Douglas, R.C.; Vessey, J.P.; Rittmaster, R.S. Dutasteride, the dual 5alpha-reductase inhibitor, inhibits androgen action and promotes cell death in the LNCaP prostate cancer cell line. Prostate 2004, 58, 130–144. [Google Scholar] [CrossRef]
- Angwafor, F., 3rd; Anderson, M.L. An open label, dose response study to determine the effect of a dietary supplement on dihydrotestosterone, testosterone and estradiol levels in healthy males. J. Int. Soc. Sports Nutr. 2008, 5, 12. [Google Scholar] [CrossRef] [Green Version]
- Shariat, S.F.; Ashfaq, R.; Roehrborn, C.G.; Slawin, K.M.; Lotan, Y. Expressıon of survıvın and apoptotıc bıomarkers ın benıgn prostatıc hyperplasıa. J. Urol. 2005, 174, 2046–2050. [Google Scholar] [CrossRef] [PubMed]
- Kramer, G.; Mitteregger, D.; Marberger, M. Is benign prostatic hyperplasia (BPH) an immune inflammatory disease? Eur. Urol. 2007, 51, 1202–1216. [Google Scholar] [CrossRef]
- Abreu, D.; Campos, E.; Seija, V.; Arroyo, C.; Suarez, R.; Rotemberg, P.; Guillama, F.; Carvalhal, G.; Campolo, H.; Machado, M.; et al. Surgical site infection in surgery for benign prostatic hyperplasia: Comparison of two skin antiseptics and risk factors. Surg. Infect. 2014, 15, 763–767. [Google Scholar] [CrossRef]
- Porter, C.M.; Shrestha, E.; Peiffer, L.B.; Sfanos, K.S. The microbiome in prostate inflammation and prostate cancer. Prostate Cancer Prostatic Dis. 2018, 21, 345–354. [Google Scholar] [CrossRef]
- Traish, A.M. Post-finasteride syndrome: A surmountable challenge for clinicians. Fertil. Steril. 2020, 113, 21–50. [Google Scholar] [CrossRef]
- Martín, R.; Miquel, S.; Ulmer, J.; Kechaou, N.; Langella, P.; Bermúdez-Humarán, L.G. Role of commensal and probiotic bacteria in human health: A focus on inflammatory bowel disease. Microb. Cell Factories 2013, 12, 71. [Google Scholar] [CrossRef] [Green Version]
- Inglin, R.C. Combined Phenotypic-Genotypic Analyses of the Genus Lactobacillus and Selection of Cultures for Biopreservation of Fermented Food. Ph.D. Thesis, ETH Zurich, Zürich, Switzerland, 2017. [Google Scholar] [CrossRef]
- Yin, S.; Xu, D.; Zhang, M.; Zhang, P.; Guan, Y.; Kzhyshkowska, J.; Liang, C. Urine flora imbalance and new biomarkers in prostate cancer and benign prostatic hyperplasia. Arch. Med. Sci. 2021, 1–11. [Google Scholar] [CrossRef]
- Mändar, R.; Punab, M.; Korrovits, P.; Türk, S.; Ausmees, K.; Lapp, E.; Preem, J.K.; Oopkaup, K.; Salumets, A.; Truu, J. Seminal microbiome in men with and without prostatitis. Int. J. Urol. 2017, 24, 211–216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takeda, K.; Okumura, K. Effects of a Fermented Milk Drink Containing Lactobacillus casei Strain Shirota on the Human NK-Cell Activity. J. Nutr. 2007, 137, S791–S793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horinaka, M.; Yoshida, T.; Kishi, A.; Akatani, K.; Yasuda, T.; Kouhara, J.; Wakada, M.; Sakai, T. Lactobacillus strains induce TRAIL production and facilitate natural killer activity against cancer cells. FEBS Lett. 2009, 584, 577–582. [Google Scholar] [CrossRef] [Green Version]
- Tsai, K.-Y.; Wu, D.-C.; Wu, W.-J.; Wang, J.-W.; Juan, Y.-S.; Li, C.-C.; Liu, C.-J.; Lee, H.-Y. Exploring the Association between Gut and Urine Microbiota and Prostatic Disease including Benign Prostatic Hyperplasia and Prostate Cancer Using 16S rRNA Sequencing. Biomedicines 2022, 10, 2676. [Google Scholar] [CrossRef]
- Kläring, K.; Hanske, L.; Bui, N.; Charrier, C.; Blaut, M.; Haller, D.; Plugge, C.M.; Clavel, T. Intestinimonas butyriciproducens gen. nov., sp. nov., a butyrate-producing bacterium from the mouse intestine. Int. J. Syst. Evol. Microbiol. 2013, 63, 4606–4612. [Google Scholar] [CrossRef] [Green Version]
- Mikami, A.; Ogita, T.; Namai, F.; Shigemori, S.; Sato, T.; Shimosato, T. Oral Administration of Flavonifractor plautii, a Bacteria Increased With Green Tea Consumption, Promotes Recovery From Acute Colitis in Mice via Suppression of IL-17. Front. Nutr. 2021, 7, 610946. [Google Scholar] [CrossRef]
- Pathak, P.; Xie, C.; Nichols, R.G.; Ferrell, J.M.; Boehme, S.; Krausz, K.W.; Patterson, A.D.; Gonzalez, F.J.; Chiang, J.Y.L. Intestine farnesoid X receptor agonist and the gut microbiota activate G-protein bile acid receptor-1 signaling to improve metabolism. Hepatology 2018, 68, 1574–1588. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.; Hong, S.N.; Paik, N.Y.; Kim, T.J.; Kim, E.R.; Chang, D.K.; Kim, Y.H. CD1d Modulates Colonic Inflammation in NOD2-/- Mice by Altering the Intestinal Microbial Composition Comprising Acetatifactor muris. J. Crohns Colitis 2019, 13, 1081–1091. [Google Scholar] [CrossRef]
- Yuan, G.-H.; Zhang, Z.; Gao, X.-S.; Zhu, J.; Guo, W.-H.; Wang, L.; Ding, P.; Jiang, P.; Li, L. Gut microbiota-mediated tributyltin-induced metabolic disorder in rats. RSC Adv. 2020, 10, 43619–43628. [Google Scholar] [CrossRef]
- Kim, J.; Kim, H.; Park, J.; Cho, H.; Kwak, M.; Kim, B.; Yang, S.; Lee, J.; Kim, D.; Joo, K.; et al. SAT-184 The potential function of gut bacteria, Oscillibacter, on the uremia of chronic kidney disease patients. Kidney Int. Rep. 2020, 5, S78. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Ouyang, M.; Gao, X.; Wang, S.; Fu, C.; Zeng, J.; He, X. Phocea, Pseudoflavonifractor and Lactobacillus intestinalis: Three Potential Biomarkers of Gut Microbiota That Affect Progression and Complications of Obesity-Induced Type 2 Diabetes Mellitus. Diabetes Metab. Syndr. Obes. Targets Ther. 2020, 13, 835–850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.; An, J.; Kim, J.; Choi, D.; Song, Y.; Lee, C.-K.; Kong, H.; Kim, S.B.; Kim, K. A Novel Bacterium, Butyricimonas virosa, Preventing HFD-Induced Diabetes and Metabolic Disorders in Mice via GLP-1 Receptor. Front. Microbiol. 2022, 13, 858192. [Google Scholar] [CrossRef] [PubMed]
- An, J.; Lee, H.; Lee, S.; Song, Y.; Kim, J.; Park, I.H.; Kong, H.; Kim, K. Modulation of Pro-inflammatory and Anti-inflammatory Cytokines in the Fat by an Aloe Gel-based Formula, QDMC, Is Correlated with Altered Gut Microbiota. Immune Netw. 2021, 21, e15. [Google Scholar] [CrossRef]
- Martin-Nuñez, G.M.; Cornejo-Pareja, I.; Clemente-Postigo, M.; Tinahones, F.J. Gut Microbiota: The Missing Link Between Helicobacter pylori Infection and Metabolic Disorders? Front. Endocrinol. 2021, 12, 639856. [Google Scholar] [CrossRef]
- Conley, M.N.; Wong, C.P.; Duyck, K.M.; Hord, N.; Ho, E.; Sharpton, T.J. Aging and serum MCP-1 are associated with gut microbiome composition in a murine model. PeerJ 2016, 4, e1854. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Feng, D.; Law, H.K.-W.; Wu, Y.; Zhu, G.-H.; Huang, W.-Y.; Kang, Y. Integrative Analysis of Gut Microbiota and Fecal Metabolites in Rats after Prednisone Treatment. Microbiol. Spectr. 2021, 9, e0065021. [Google Scholar] [CrossRef]
- Banala, U.K.; Das, N.P.I.; Toleti, S.R. Microbial interactions with uranium: Towards an effective bioremediation approach. Environ. Technol. Innov. 2020, 21, 101254. [Google Scholar] [CrossRef]
- Coutinho, C.M.L.M.; Coutinho-Silva, R.; Zinkevich, V.; Pearce, C.B.; Ojcius, D.M.; Beech, I. Sulphate-reducing bacteria from ulcerative colitis patients induce apoptosis of gastrointestinal epithelial cells. Microb. Pathog. 2017, 112, 126–134. [Google Scholar] [CrossRef] [Green Version]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
An, J.; Song, Y.; Kim, S.; Kong, H.; Kim, K. Alteration of Gut Microbes in Benign Prostatic Hyperplasia Model and Finasteride Treatment Model. Int. J. Mol. Sci. 2023, 24, 5904. https://doi.org/10.3390/ijms24065904
An J, Song Y, Kim S, Kong H, Kim K. Alteration of Gut Microbes in Benign Prostatic Hyperplasia Model and Finasteride Treatment Model. International Journal of Molecular Sciences. 2023; 24(6):5904. https://doi.org/10.3390/ijms24065904
Chicago/Turabian StyleAn, Jinho, Youngcheon Song, Sangbum Kim, Hyunseok Kong, and Kyungjae Kim. 2023. "Alteration of Gut Microbes in Benign Prostatic Hyperplasia Model and Finasteride Treatment Model" International Journal of Molecular Sciences 24, no. 6: 5904. https://doi.org/10.3390/ijms24065904