
METTL3-Mediated LncRNA EN_42575 m6A Modification Alleviates CPB2 Toxin-Induced Damage in IPEC-J2 Cells
 ,
,
Abstract
:1. Introduction
2. Results
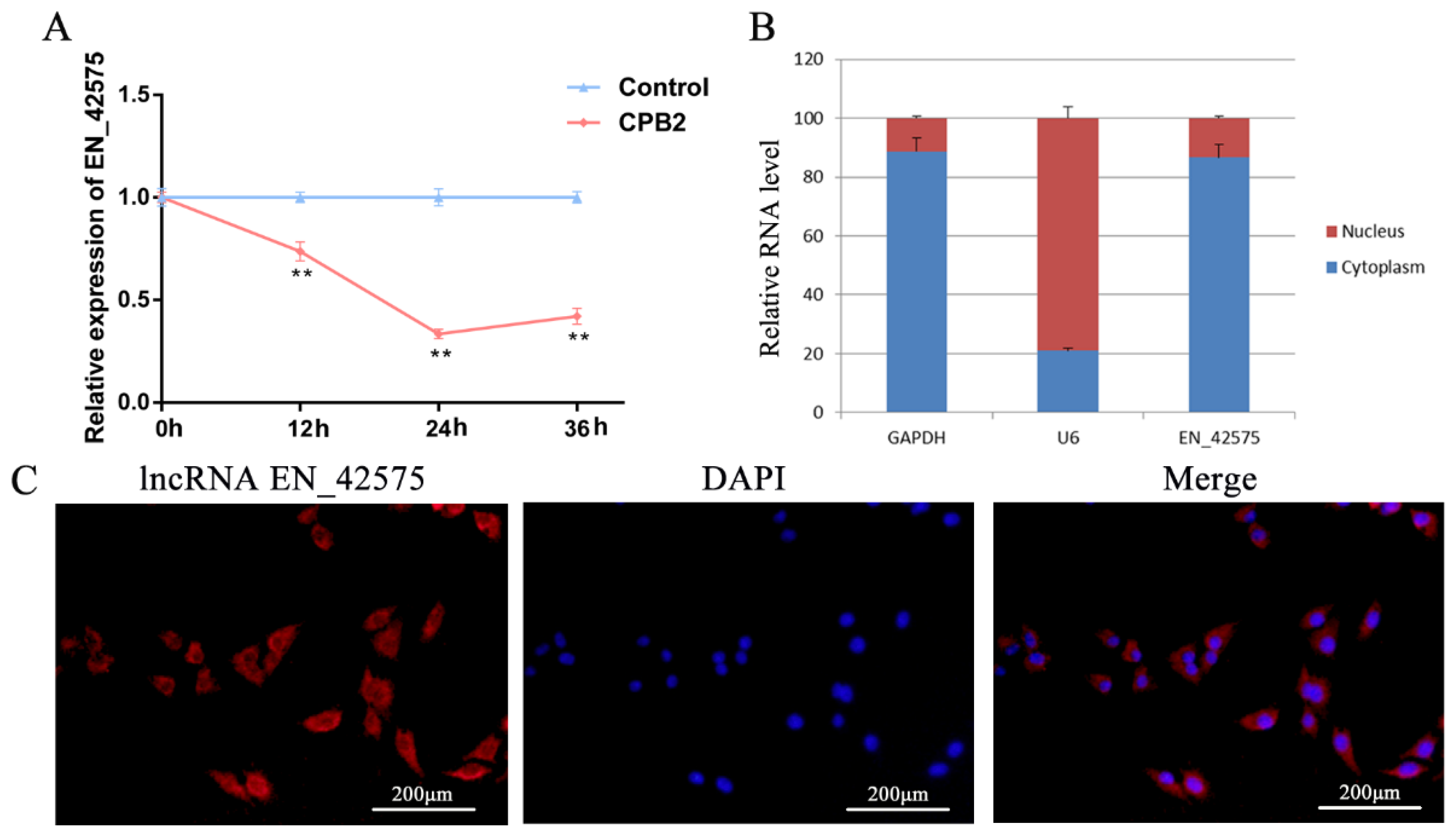
2.1. LncRNA EN_42575 Expression Pattern Analysis
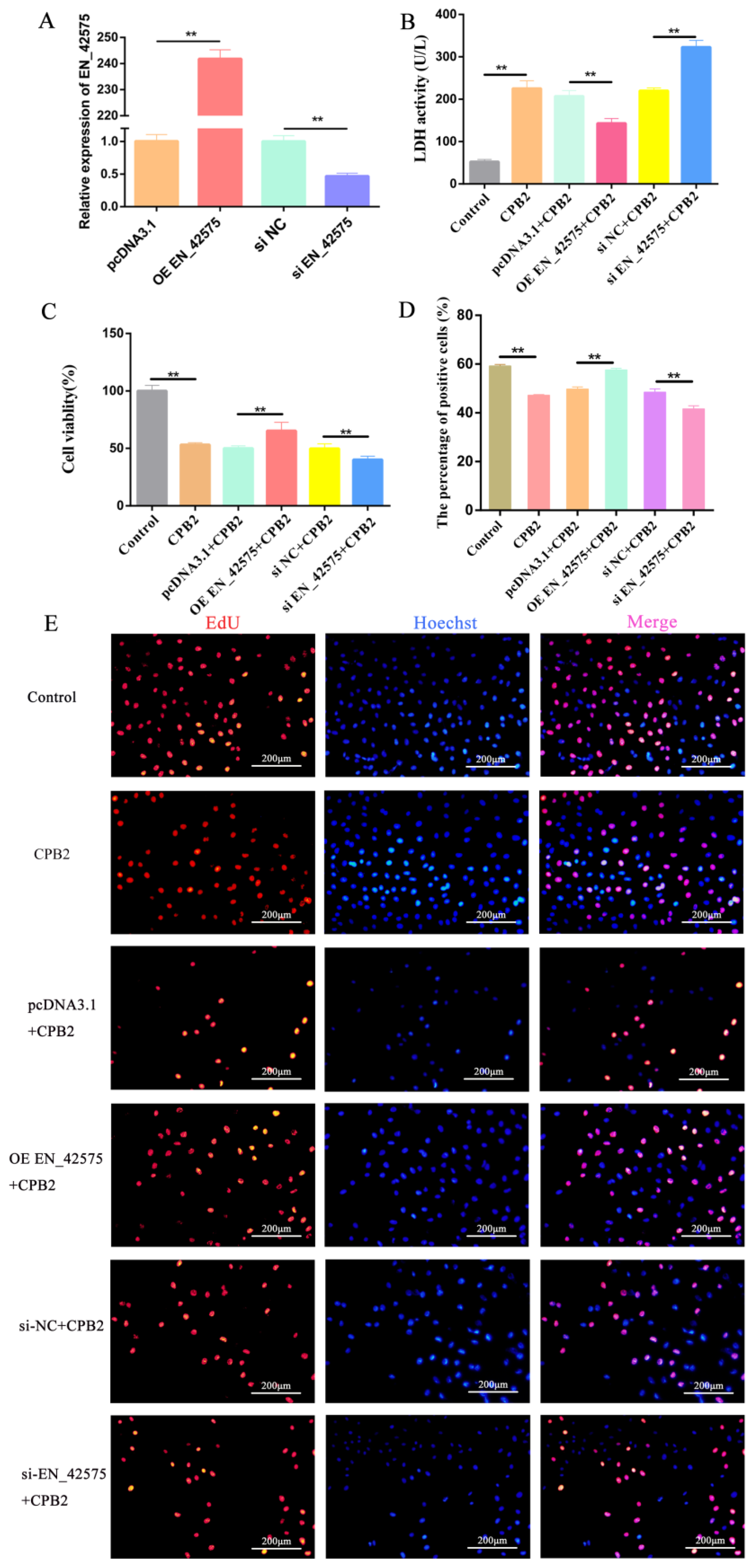
2.2. LncRNA EN_42575 Promotes CPB2-Induced Proliferation of IPEC-J2 Cells
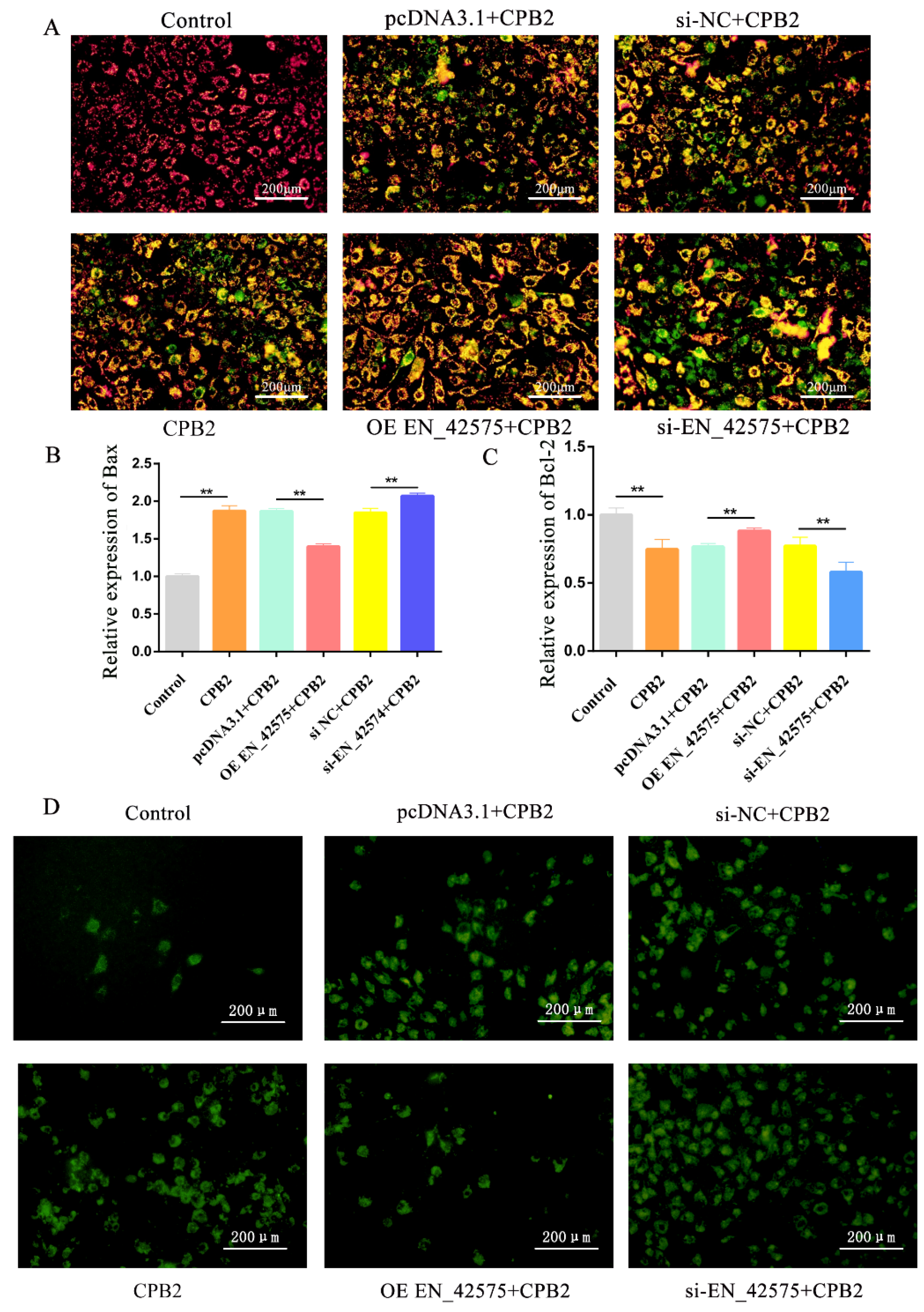
2.3. LncRNA EN_42575 Inhibits CPB2 Toxin-Induced Apoptosis and Oxidative Damage
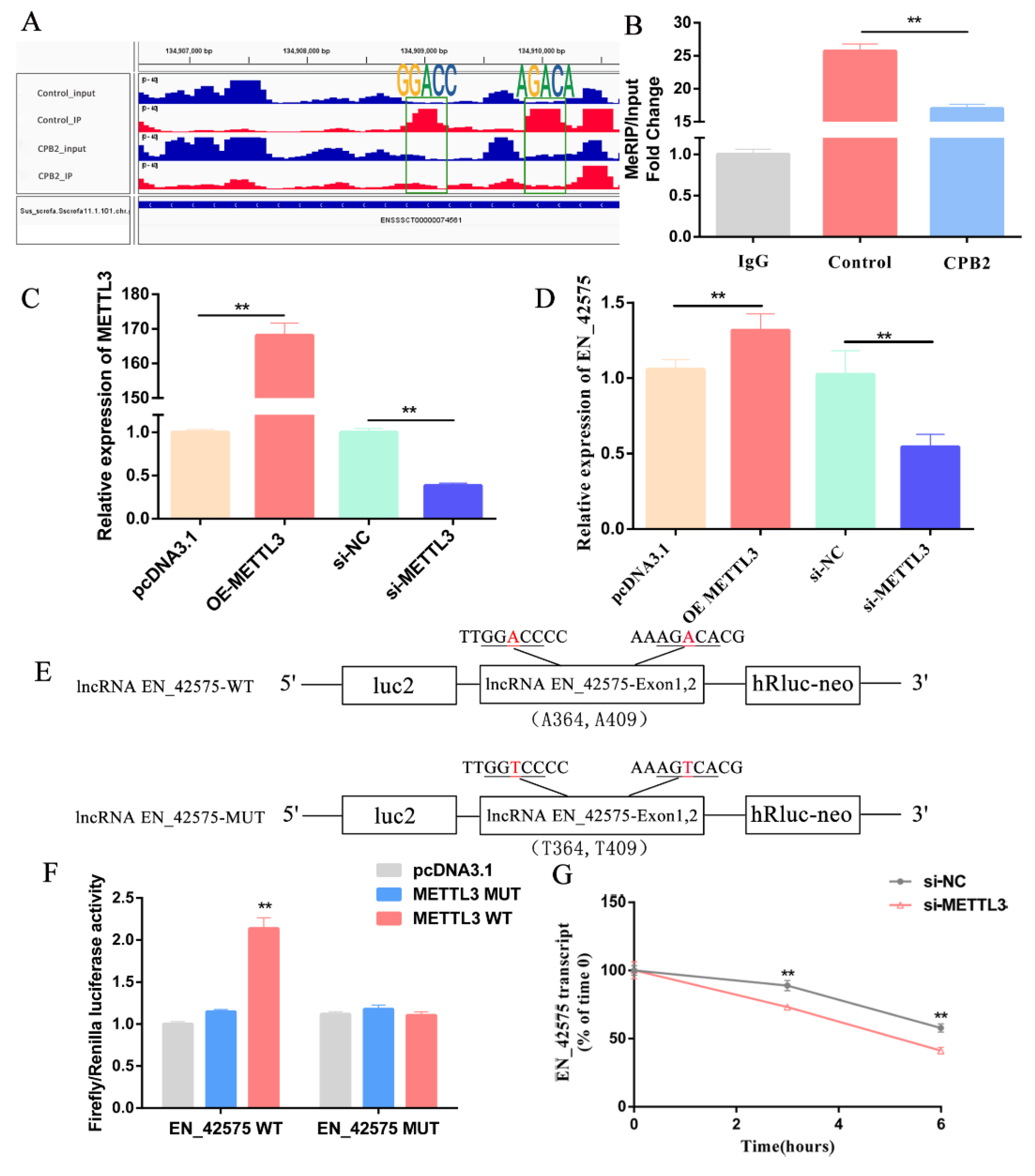
2.4. LncRNA EN_42575 Is a Target of METTL3
2.5. Functional Analysis of lncRNA EN_42575 Target Genes
3. Discussion
4. Materials and Methods
4.1. Cell Culture and CPB2 Toxin Treatment
4.2. Cell Transfection
4.3. Detection of Lactate Dehydrogenase Activity (LDH)
4.4. CCK-8 and Δψm Measurements
4.5. Reactive Oxygen Species (ROS) Determination
4.6. Cell Proliferation Assay
4.7. RT-qPCR
4.8. MeRIP-qPCR
4.9. Dual-Luciferase Reporter Assay
4.10. Single-RNA Fluorescence In Situ Hybridization (FISH)
4.11. RNA Stability
4.12. RNA Pull-Down and Mass Spectroscopy Analyses
4.13. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Zhu, S.; Wang, J.Z.; Chen, D.; He, Y.T.; Meng, N.; Chen, M.; Lu, R.X.; Chen, X.H.; Zhang, X.L.; Yan, G.A.-O. An oncopeptide regulates m(6)a recognition by the m(6)a reader igf2bp1 and tumorigenesis. Nat. Commun. 2020, 3, 1685. [Google Scholar] [CrossRef] [Green Version]
- Yang, G.; Sun, Z.; Zhang, N.A.-O. Reshaping the role of m6a modification in cancer transcriptome: A review. Cancer Cell Int. 2020, 29, 353. [Google Scholar] [CrossRef]
- Gu, J.; Zhan, Y.; Zhuo, L.; Zhang, Q.; Li, G.; Li, Q.; Qi, S.; Zhu, J.; Lv, Q.; Shen, Y.; et al. Biological functions of m(6)a methyltransferases. Cell Biosci. 2021, 11, 15. [Google Scholar] [CrossRef] [PubMed]
- Tong, J.; Flavell, R.A.; Li, H.B. Rna m(6)a modification and its function in diseases. Front. Med. 2018, 12, 481–489. [Google Scholar] [CrossRef] [Green Version]
- Elsabbagh, R.A.; Rady, M.; Watzl, C.; Abou-Aisha, K.; Gad, M.Z. Impact of n6-methyladenosine (m(6)a) modification on immunity. Cell Commun. Signal. 2022, 20, 140. [Google Scholar] [CrossRef]
- Liu, H.; Xu, Y.; Yao, B.; Sui, T.; Lai, L.; Li, Z. A novel n6-methyladenosine (m6a)-dependent fate decision for the lncrna thor. Cell Death Dis. 2020, 13, 613. [Google Scholar] [CrossRef]
- Pacholewska, A.A.-O.; Sung, M.A.-O. Lncrna expression predicts mrna abundance. Epigenomics 2019, 11, 1121–1128. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.; Zhou, Q.; Wang, C.Q.; Zhu, L.; Jin, H. Lncrnas regulate metabolism in cancer. Int. J. Biol. Sci. 2020, 16, 1194–1206. [Google Scholar] [CrossRef]
- Ding, X.; Jia, X.; Wang, C.; Xu, J.; Gao, S.-J.; Lu, C. Correction to: A dhx9-lncrna-mdm2 interaction regulates cell invasion and angiogenesis of cervical cancer. Cell Death Differ. 2019, 12, 26. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Cen, S. Roles of lncrnas in influenza virus infection. Emerg. Microbes Infect. 2020, 9, 1407–1414. [Google Scholar] [CrossRef] [PubMed]
- Jiang, B.; Liu, M.; Li, P.; Zhu, Y.; Liu, Y.; Zhu, K.; Zuo, Y.; Li, Y. Rna-seq reveals a novel porcine lncrna mphosph9-ot1 induces cxcl8/il-8 expression in etec infected ipec-j2 cells. Front. Cell. Infect. Microbiol. 2022, 25, 996841. [Google Scholar] [CrossRef] [PubMed]
- Lan, Y.; Liu, B.; Guo, H. The role of m 6 a modification in the regulation of tumor related lncrnas. Mol. Ther. Nucleic Acids 2021, 9, 768–779. [Google Scholar] [CrossRef]
- Yang, J.; Yang, Q.; Zhang, J.; Gao, X.; Luo, R.; Xie, K.; Wang, W.; Li, J.; Huang, X.; Yan, Z.; et al. N6-methyladenosine methylation analysis of long noncoding rnas and mrnas in ipec-j2 cells treated with clostridium perfringens beta2 toxin. Front. Immunol. 2021, 12, 769204. [Google Scholar] [CrossRef] [PubMed]
- Songer, J.G. Clostridial enteric diseases of domestic animals. Clin. Microbiol. Rev. 1996, 9, 216–234. [Google Scholar] [CrossRef]
- Straw, B.E.; Zimmerman, J.J.; D’Allaire, S. Diseases of swine. Can. Vet. J. 2000, 41, 145. [Google Scholar]
- Kreft, B.; Dalhoff, K.; Sack, K. Necrotizing enterocolitis: A historical and current review. Med. Klin. 2000, 95, 435–441. [Google Scholar] [CrossRef]
- Jäggi, M.; Wollschläger, N.; Abril, C.; Albini, S.; Brachelente, C.; Wyder, M.; Posthaus, H. Retrospective study on necrotizing enteritis in piglets in switzerland. Schweiz Arch Tierheilkd 2009, 151, 369–375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allaart, J.G.; van Asten, A.J.; Vernooij, J.C.; Gröne, A. Beta2 toxin is not involved in in vitro cell cytotoxicity caused by human and porcine cpb2-harbouring clostridium perfringens. Vet. Microbiol. 2014, 25, 132–138. [Google Scholar] [CrossRef]
- Athira, C.K.; Milton, A.A.P.; Reddy, A.; Mekhemadhom Rajendrakumar, A.; Abhishek; Verma, M.R.; Kumar, A.; Nagaleekar, V.K.; Agarwal, R.K. Diversity of toxin-genotypes among clostridium perfringens isolated from healthy and diarrheic neonatal cattle and buffalo calves. Anaerobe 2018, 49, 99–102. [Google Scholar] [CrossRef]
- Bacciarini, L.N.; Boerlin, P.; Straub, R.; Frey, J.; Gröne, A. Immunohistochemical localization of clostridium perfringens beta2-toxin in the gastrointestinal tract of horses. Vet. Pathol. 2003, 40, 376–381. [Google Scholar] [CrossRef] [Green Version]
- Diab, S.S.; Kinde, H.; Moore, J.; Shahriar, M.F.; Odani, J.; Anthenill, L.; Songer, G.; Uzal, F.A. Pathology of clostridium perfringens type c enterotoxemia in horses. Vet. Pathol. 2012, 49, 255–263. [Google Scholar] [CrossRef]
- Garcia, J.P.; Beingesser, J.; Fisher, D.J.; Sayeed, S.; McClane, B.A.; Posthaus, H.; Uzal, F.A. The effect of clostridium perfringens type c strain cn3685 and its isogenic beta toxin null mutant in goats. Vet. Microbiol. 2012, 15, 412–419. [Google Scholar] [CrossRef] [Green Version]
- Lebrun, M.; Filée, P.; Mousset, B.; Desmecht, D.; Galleni, M.; Mainil, J.G.; Linden, A. The expression of clostridium perfringens consensus beta2 toxin is associated with bovine enterotoxaemia syndrome. Vet. Microbiol. 2007, 25, 151–157. [Google Scholar] [CrossRef]
- Gao, X.; Yang, Q.; Huang, X.; Yan, Z.; Zhang, S.; Luo, R.; Wang, P.; Wang, W.; Xie, K.; Jiang, T.; et al. Effects of clostridium perfringens beta2 toxin on apoptosis, inflammation, and barrier function of intestinal porcine epithelial cells. Microb. Pathog. 2020, 147, 104379. [Google Scholar] [CrossRef]
- Luo, R.; Yang, Q.; Huang, X.; Yan, Z.; Gao, X.; Wang, W.; Xie, K.; Wang, P.; Gun, S. Clostridium perfringens beta2 toxin induced in vitro oxidative damage and its toxic assessment in porcine small intestinal epithelial cell lines. Gene 2020, 759, 144999. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Fan, H.; Jin, J.; Gao, S.; Huang, R.; Wu, S.; Bao, W.A.-O. Insight into mechanisms of pig lncrna fut3-as1 regulating E. coli f18-bacterial diarrhea. PLoS Pathog. 2022, 13, e1010584. [Google Scholar] [CrossRef]
- Chen, J.; Zhang, C.; Zhang, N.; Liu, G. Porcine endemic diarrhea virus infection regulates long noncoding rna expression. Virology 2019, 15, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Zhang, J.; Yang, Q.; Huang, X.; Yan, Z.; Wang, P.; Gao, X.; Li, J.; Li, N.; Gao, Y.; et al. Lncrna en-90756 promotes cpb2-induced proliferation and inhibits apoptosis in ipec-j2 cells by affecting the jak-stat signaling pathway activation. Front. Microbiol. 2023, 13, 1082025. [Google Scholar] [CrossRef]
- Dominissini, D.; Moshitch-Moshkovitz, S.; Schwartz, S.; Salmon-Divon, M.; Ungar, L.; Osenberg, S.; Cesarkas, K.; Jacob-Hirsch, J.; Amariglio, N.; Kupiec, M.; et al. Topology of the human and mouse m6A RNA methylomes revealed by m6A-seq. Nature 2012, 485, 201–206. [Google Scholar] [CrossRef] [PubMed]
- Nie, Y.; Tian, G.G.; Zhang, L.; Lee, T.; Zhang, Z.; Li, J.A.-O.; Sun, T. Identifying cortical specific long noncoding rnas modified by m(6)a rna methylation in mouse brains. Epigenetics 2021, 16, 1260–1276. [Google Scholar] [CrossRef]
- Song, Y.; Pan, Y.; Wu, M.; Sun, W.; Luo, L.; Zhao, Z.; Liu, J.A.-O. Mettl3-mediated lncrna m(6)a modification in the osteogenic differentiation of human adipose-derived stem cells induced by nel-like 1 protein. Stem Cell Rev. Rep. 2021, 17, 2276–2290. [Google Scholar] [CrossRef] [PubMed]
- Yoneda, R.; Ueda, N.; Uranishi, K.; Hirasaki, M.; Kurokawa, R. Long noncoding rna pncrna-d reduces cyclin d1 gene expression and arrests cell cycle through rna m(6)a modification. J. Biol. Chem. 2020, 24, 5626–5639. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Yan, B.; Wang, X.; Li, Q.; Kan, X.; Wang, J.; Sun, Y.A.-O.; Wang, P.; Tian, L.; Liu, M. Alkbh5-mediated m6a modification of lncrna kcnq1ot1 triggers the development of lscc via upregulation of hoxa9. J. Cell. Mol. Med. 2022, 26, 385–398. [Google Scholar] [CrossRef]
- Xue, L.; Li, J.; Lin, Y.; Liu, D.; Yang, Q.; Jian, J.; Peng, J.A.-O. M(6) a transferase mettl3-induced lncrna abhd11-as1 promotes the warburg effect of non-small-cell lung cancer. J. Cell Physiol. 2021, 236, 2649–2658. [Google Scholar] [CrossRef]
- Huang, Y.A.-O.; Qiao, Y.; Zhao, Y.; Li, Y.; Yuan, J.; Zhou, J.; Sun, H.A.-O.; Wang, H.A.-O. Large scale rna-binding proteins/lncrnas interaction analysis to uncover lncrna nuclear localization mechanisms. Brief. Bioinform. 2021, 5, bbab195. [Google Scholar] [CrossRef]
- Miao, H.; Wang, L.; Zhan, H.A.-O.; Dai, J.; Chang, Y.A.-O.; Wu, F.; Liu, T.; Liu, Z.; Gao, C.; Li, L.A.-O.; et al. A long noncoding rna distributed in both nucleus and cytoplasm operates in the pycard-regulated apoptosis by coordinating the epigenetic and translational regulation. PLoS Genet. 2019, 15, e1008144. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Feng, T.; Lian, Y.; Zhang, G.; Garen, A.; Song, X. Role of human noncoding rnas in the control of tumorigenesis. Proc. Natl. Acad. Sci. USA 2009, 106, 12956–12961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, G.; Cui, Y.; Zhang, G.; Garen, A.; Song, X. Regulation of proto-oncogene transcription, cell proliferation, and tumorigenesis in mice by psf protein and a vl30 noncoding rna. Proc. Natl. Acad. Sci. USA 2009, 29, 16794–16798. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.L. Linking long noncoding rna localization and function. Trends Biochem. Sci. 2016, 41, 761–772. [Google Scholar] [CrossRef] [PubMed]
- Hartford, C.C.R.; Lal, A. When long noncoding becomes protein coding. Mol. Cell. Biol. 2020, 27, e00528-19. [Google Scholar] [CrossRef] [Green Version]
- Lan, Y.; Xiao, X.; He, Z.; Luo, Y.; Wu, C.; Li, L.; Song, X. Long noncoding rna occ-1 suppresses cell growth through destabilizing hur protein in colorectal cancer. Nucleic Acids Res. 2018, 20, 5809–5821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative pcr and the 2(-delta delta c(t)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name/Id | Primer Sequence (5′-3′) | Product Length | Type | |
|---|---|---|---|---|
| ENSSSCG00000042575 | Forward | TGAATCAGCAGATACGGGCA | 82 | MeRIP-qPCR/RT-qPCR |
| Reverse | GAAACTTGTACGGGCATCCA | |||
| Bax | Forward | GCTGACGGCAACTTCAACTG | 202 | RT-qPCR |
| Reverse | GCGTCCCAAAGTAGGAGAGG | |||
| Bcl-2 | Forward | GGTGAACTGGGGGAGGATTG | 130 | RT-qPCR |
| Reverse | GTGCCGGTTCAGGTACTCAG | |||
| METTL3 | Forward | CCACTTCTGGTGGCCCTAAG | 104 | RT-qPCR |
| Reverse | CGCCAGATCAGAAAGGTGGT | |||
| GAPDH | Forward | AGTATGATTCCACCCACGGC | 139 | RT-qPCR |
| Reverse | TACGTAGCACCAGCATCACC | |||
| U6 | Forward | TTATGGGTCCTAGCCTGAC | 224 | RT-qPCR |
| Reverse | CACTATTGCGGGTCTGC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, J.; Yang, Q.; Huang, X.; Yan, Z.; Wang, P.; Gao, X.; Li, J.; Gun, S. METTL3-Mediated LncRNA EN_42575 m6A Modification Alleviates CPB2 Toxin-Induced Damage in IPEC-J2 Cells. Int. J. Mol. Sci. 2023, 24, 5725. https://doi.org/10.3390/ijms24065725
Yang J, Yang Q, Huang X, Yan Z, Wang P, Gao X, Li J, Gun S. METTL3-Mediated LncRNA EN_42575 m6A Modification Alleviates CPB2 Toxin-Induced Damage in IPEC-J2 Cells. International Journal of Molecular Sciences. 2023; 24(6):5725. https://doi.org/10.3390/ijms24065725
Chicago/Turabian StyleYang, Jiaojiao, Qiaoli Yang, Xiaoyu Huang, Zunqiang Yan, Pengfei Wang, Xiaoli Gao, Jie Li, and Shuangbao Gun. 2023. "METTL3-Mediated LncRNA EN_42575 m6A Modification Alleviates CPB2 Toxin-Induced Damage in IPEC-J2 Cells" International Journal of Molecular Sciences 24, no. 6: 5725. https://doi.org/10.3390/ijms24065725